(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
|
|
|
- Nadzieja Wróblewska
- 6 lat temu
- Przeglądów:
Transkrypt
 O udzieleniu patentu europejskiego ogłoszono: 22.06.11 Europejski Biuletyn Patentowy 11/2 EP 97444 B1 (13) (1) T3 Int.Cl. C07K 14/70 (06.01) C12N 1/8 (06.01) A01K 67/027 (06.")
1 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: (97) O udzieleniu patentu europejskiego ogłoszono: Europejski Biuletyn Patentowy 11/2 EP B1 (13) (1) T3 Int.Cl. C07K 14/70 (06.01) C12N 1/8 (06.01) A01K 67/027 (06.01) (4) Tytuł wynalazku: Sposób zastosowania transgenicznego zwierzęcia o podwyższonej odpowiedzi immunologicznej () Pierwszeństwo: HU HU (43) Zgłoszenie ogłoszono: w Europejskim Biuletynie Patentowym nr 09/37 (4) O złożeniu tłumaczenia patentu ogłoszono: Wiadomości Urzędu Patentowego 11/11 (73) Uprawniony z patentu: Agricultural Biotechnology Center, Gödöllö, HU Eötvös Lorand University, Budapest, HU (72) Twórca(y) wynalazku: PL/EP T3 ZSUZSANNA BÖSZE, Gödöllö, HU IMRE KACSKOVICS, Budakeszi, HU JUDIT CERVENAK, Budakeszi, HU LÁSZLÓ HIRIPI, Gödöllö, HU BALÁZS BENDER, Bátonyterenye, HU (74) Pełnomocnik: rzecz. pat. Urszula Bartnik POLSERVICE KANCELARIA RZECZNIKÓW PATENTOWYCH SP. Z O.O. ul. Bluszczańska Warszawa Uwaga: W ciągu dziewięciu miesięcy od publikacji informacji o udzieleniu patentu europejskiego, każda osoba może wnieść do Europejskiego Urzędu Patentowego sprzeciw dotyczący udzielonego patentu europejskiego. Sprzeciw wnosi się w formie uzasadnionego na piśmie oświadczenia. Uważa się go za wniesiony dopiero z chwilą wniesienia opłaty za sprzeciw (Art. 99 (1) Konwencji o udzielaniu patentów europejskich).
2 /2862 EP B1 Opis [0001] Niniejszy wynalazek dotyczy dziedziny immunologii. Bardziej szczegółowo, wynalazek dotyczy transgenicznego (Tg) zwierzęcia innego niż człowiek, zdolnego do rozwijania wzmożonej humoralnej odpowiedzi immunologicznej przeciwko antygenowi w porównaniu z nietransgenicznym zwierzęciem kontrolnym tego samego gatunku, przy czym to transgeniczne zwierzę inne niż człowiek posiada konstrukt genetyczny zapewniający wzmożoną aktywność noworodkowego receptora Fc związanego z klasą I MHC (FcRn). [0002] Niniejszy wynalazek zapewnia nieterapeutyczny sposób rozwijania wzmożonej humoralnej odpowiedzi immunologicznej, przy czym wspomniany sposób obejmuje wykorzystanie tego transgenicznego zwierzęcia innego niż człowiek i antygenu będącego przedmiotem zainteresowania. [0003] W odpowiedzi na antygen z limfocytów B rozwijają się komórki plazmatyczne B, w swoistej humoralnej reakcji immunologicznej, osiągającej szczyt około 1-2 tygodni po ekspozycji na antygen. Drugie zetknięcie z antygenem powoduje wydzielanie przeciwciał o zwiększonym powinowactwie do antygenu, zwiększony pik miana swoistych przeciwciała w surowicy i stały poziom przeciwciał w surowicy. Utrzymywanie się w surowicy poziomu swoistych przeciwciał wymaga ciągłego wydzielania Ig przez komórki plazmatyczne i ochrony ich przed szybką eliminacją. O ile IgM, IgA i IgE są usuwane z organizmu względnie szybko, czas półtrwania
3 4 1 2 IgG w surowicy jest wydłużony. W roku 198 Brambell opisał ulegający wysyceniu układ receptorowy, pośredniczący w transporcie matczynej IgG (Brambell, 198); następnie wysnuł on wniosek o istnieniu podobnego lub identycznego receptora, chroniącego IgG przed katabolizmem i sprawiającego, że IgG żyje najdłużej ze wszystkich białek osocza (Brambell i wsp., 1964) Istotnie, w późniejszym okresie wykazano, że receptor Brambella (FcRB) pośredniczy w przekazywaniu IgG w okresie przed urodzeniem i(lub) w okresie noworodkowym (w tej funkcji jest on określany terminem noworodkowy receptor Fc, w skrócie, od angielskiego neonatal Fc receptor - FcRn), i pośredniczy w ochronie IgG przed katabolizmem (Junghans, 1997). [0004] FcRn po raz pierwszy zidentyfikowano u gryzoni jako receptor przenoszący immunoglobuliny macierzyste od matki do noworodka poprzez jelito noworodka (Rodewald, 1976; Simister i Rees, 198). Od czasu pierwszego jego opisania w jelicie noworodka szczura przez Simistera i Mostova (Simister i Mostov, 1989), różne badania wykazały, że FcRn odgrywa centralną rolę w regulowaniu transportu IgG w obrębie komórki i pomiędzy komórkami o różnym pochodzeniu (Antohe i wsp., 01; Claypool i wsp., 04; Dickinson i wsp., 1999; Kobayashi i wsp., 02; McCarthy i wsp., 00; Ober i wsp., 04; Spiekermann i wsp., 02). Służy on również do ochrony IgG i albuminy, dwóch rozpuszczalnych białek najobficiej występujących w surowicy, przed rozpadem, przedłużając w ten sposób ich czas półtrwania (Ghetic i wsp., 1996; Israel i wsp., 1996; Junghans i Anderson, 1996). Początkowo sądzono, że w tym mechanizmie
4 1 2 pośredniczą głównie komórki śródbłonka, wyściełające naczynia krwionośne (Borvak i wsp., 1998), niedawno przeprowadzone badania sugerują jednak, że proces ten zachodzi także w innych komórkach (Akilesh i wsp., 07; Lu i wsp., 07). Wewnątrz tych komórek FcRn znajduje się głównie we wczesnych/ulegających recyklingowi endosomach, gdzie napotyka na IgG i albuminę internalizowane na drodze endocytozy płynnej fazy. Kwaśne środowisko endosomów ułatwia interakcję. Związane IgM i albumina ulegają recyklingowi (ponownie wchodzą w obieg), przemieszczając się na powrót ku powierzchni i ulegając usunięciu z komórki, natomiast ligandy niezwiązane są przenoszone w dół i ulegają rozpadowi w lizosomach (Anderson i wsp., 06; Roopenian i Akilesh, 07). Ostatnie dane potwierdzają nową koncepcję, a mianowicie założenie, że FcRn odgrywa główną rolę w fagocytozie zależnej od IgG (Vidarsson i wsp., 06). [000] Funkcjonalna cząsteczka FcRn jest heterodimerem złożonym z łańcucha alfa przypominającego klasę I MHC (lub łańcucha ciężkiego) i beta 2-mikroglobuliny ( 2m; nazwa alternatywna: łańcuch lekki) (Simister i Mostov, 1989), które wiążą IgG i albuminę w sposób zależny od ph (Chaudhury i wsp., 03; Simister i Mostov, 1989), aczkolwiek w różnych miejscach wiązania (Andersen i wsp., 06; Chaudhury i wsp., 06). [0006] FcRn klonuje się z szeregu gatunków ssaków, takich jak na przykład szczur (Simister i Mostov, 1989), mysz (Ahouse i wsp., 1993), człowiek (Story i wsp., 1994), bydło (Kacskovics i wsp., 00) opos (Adamski i wsp., 00), owca (Mayer i wsp., 02)
5 6 1 2 świnia (Schnulle i Hurley, 03; Zhao i wsp., 03), wielbłąd i pies (Kacskovics i wsp., 06b). Niedawno autorom niniejszego wynalazku udało się sklonować łańcuch alfa FcRn królika i określić jego właściwości. Jakkolwiek większość funkcji FcRn opisano w odniesieniu do myszy, badania u innych ssaków sugerują, że rola FcRn w katabolizmie IgG ma podobne, kluczowe znaczenie u wszystkich badanych ssaków, takich jak gryzonie, człowiek i naczelne (Ghetie i Ward, 02), świnia (Harmsen i wsp., 0) i bydło (Kacskovics i wsp., 06a). [0007] Niedawno wykazano, w dwóch różnych modelach mysich, że nadmierna ekspresja bydlęcych łańcuchów alfa FcRn (bfcrn) istotnie wydłuża czas półtrwania mysiej IgG u tych zwierząt (Bender i wsp., 07; Lu i wsp., 07), co wskazuje, że bfcrn tworzy funkcjonalny kompleks z mysim 2m (m 2m) i wiąże się z mysimi i ludzkimi IgG. Autorzy niniejszego wynalazku stwierdzili również, że nadmierna ekspresja bfcrn u myszy transgenicznych (Tg) (wykorzystywano takie same myszy, jakie wykorzystywali Bender i wsp., 07) umożliwiała tym zwierzętom wytwarzanie po immunizacji istotnie podwyższonego poziomu antygenowo swoistych IgG i IgM. [0008] W WO ujawniono wytwarzanie przeciwciał monoklonalnych, przy czym komórki wytwarzające przeciwciała transfekuje się kwasem nukleinowym kodującym gen FcRn. Zmodyfikowane komórki wykazują ekspresję FcRn a wytwarzanie przeciwciał jest podwyższone. Autorzy nie zapewniają ujawnienia ani uzyskiwania transgenicznych zwierząt, ani samych zwierząt, wspominają jednak o korzyściach z
6 7 1 2 wykorzystania zwierzęcia innego niż człowiek, z co najmniej jedną (dodatkową) kopią kwasu nukleinowego kodującą FcRn celem regulacji w górę poziomu przeciwciał we krwi. Opis jest jednak czysto teoretyczny; na podstawie tego opisu można sądzić, że autorzy skupili się na wytwarzaniu przeciwciał monoklonalnych in vivo, na przykład w płynie puchlinowym myszy. Autorzy nie wspominają również o korzyściach płynących z nadmiernej ekspresji FcRn w zakresie wzmagania odpowiedzi immunologicznej. [0009] Jasne jest, że w stanie techniki nadal istnieje zapotrzebowanie na poprawę w zakresie dostępnych immunoglobulin do zastosowania diagnostycznego, naukowego i terapeutycznego, w dużych ilościach i o wysokiej jakości. W celu spełnienia tego zapotrzebowania niniejszy wynalazek zapewnia transgeniczne zwierzęta, jakie reagują na antygenowa ekspozycję wybitnie podwyższonym poziomem swoistej humoralnej reakcji immunologicznej. [00] Nieoczekiwanie stwierdzono, że uzyskanie myszy BAC Tg, o zależnej od liczby kopii, nadmiernej ekspresji bydlęcego łańcucha alfa FcRn (kodowanego przez bydlęcy gen FCGRT) powodowało nie tylko to, że bydlęcy łańcuch alfa FcRn tworzył kompleks funkcjonalny z mysim 2m i istotnie zwiększał czas półtrwania podawanej egzogennie mysiej i ludzkiej IgG, lecz te transgeniczne zwierzęta wykazywały także wybitne nasilenie humoralnej odpowiedzi immunologicznej po immunizacji, w porównaniu z ich odpowiednikami - zwierzętami kontrolnymi typu dzikiego. Większość antygenowo swoistych przeciwciał stanowiła IgG, lecz
7 8 1 2 miano IgM było również podwyższone w czasie wtórnej odpowiedzi immunologicznej. Przy analizowaniu możliwych przyczyn autorzy niniejszego wynalazku wykryli istotne zwiększenie liczby antygenowo swoistych komórek B i komórek dendrytycznych oraz masywny napływ neutrofili do wtórnych narządów limfoidalnych po immunizacji u transgenicznych myszy bfcrn w porównaniu z ich odpowiednikami - zwierzętami kontrolnymi typu dzikiego. Należy zauważyć, że podobne, choć mniej zaznaczone zmiany obserwowano u zwierząt kontrolnych typu dzikiego. Wskutek tego można było wykazać, że podwyższenie poziomu antygenowo swoistych IgG i IgM u zwierząt Tg bfcrn było rezultatem nie tylko ulepszonej ochrony IgG, lecz także większej ekspansji antygenowo swoistych, klonalnych komórek B i w następstwie tego intensywniejszej syntezy immunoglobulin w porównaniu z ich odpowiednikami - zwierzętami kontrolnymi typu dzikiego. Te wyniki wskazują na nową rolę FcRn w odpowiedzi immunologicznej. W niniejszym wynalazku po raz pierwszy ujawnia się, że wskutek stanu istotnie zwiększonej, nadmiernej ekspresji FcRn, niewystępującej u zwierząt w stanie fizjologicznym, osiąga się duży wpływ na odpowiedź immunologiczną po immunizacji, przez co system jest szczególnie użyteczny do wytwarzania różnych typów przeciwciał przeciwko szeregowi antygenów. Krótki opis wynalazku [0011] Wynalazek w jego najszerszym sensie omówiono szczegółowo w zastrzeżeniu niezależnym. [0012] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym wzmożenie
8 9 1 2 humoralnej odpowiedzi immunologicznej obejmuje wytwarzanie podwyższonego poziomu immunoglobulin po immunizacji antygenem, przy czym FcRn wykazuje swoiste powinowactwo do immunoglobulin, wytwarzanych przez zwierzę po immunizacji. [0013] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym to wzmożenie humoralnej odpowiedzi immunologicznej obejmuje wzmożenie ekspansji antygenowo swoistych klonalnych komórek B. [0014] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym to wzmożenie humoralnej odpowiedzi immunologicznej obejmuje stymulację napływu neutrofili do wtórnych narządów limfoidalnych. [001] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym to wzmożenie humoralnej odpowiedzi immunologicznej obejmuje stymulację napływu komórek dendrytycznych i(lub) makrofagów do wtórnych narządów limfoidalnych. [0016] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym do tych wtórnych narządów limfoidalnych należy śledziona. [0017] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym antygenem jest słaby immunogen. [0018] W jednym z przykładów wykonania przedmiotem niniejszego wynalazku jest sposób wytwarzania immunoglobuliny, obejmujący co najmniej jeden etap, w którym zastosowany protokół dostosowuje się do faktu, że zwierzę Tg rozwija taki sam poziom humoralnej
9 1 2 odpowiedzi immunologicznej po mniejszej liczbie ekspozycji na antygen w porównaniu z nietransgenicznym zwierzęciem kontrolnym tego samego gatunku. [0019] W jeszcze innym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób wytwarzania immunoglobulin, obejmujący co najmniej jeden etap, w którym zastosowany protokół dostosowuje się do faktu, że zwierzę Tg rozwija taki sam poziom humoralnej odpowiedzi immunologicznej szybciej niż nietransgeniczne zwierzę kontrolne tego samego gatunku. [00] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym zwierzęciem jest ssak inny niż człowiek. [0021] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym ssakiem jest gryzoń. [0022] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym gryzoniem jest mysz. [0023] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym ssakiem jest królik. [0024] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym ssak jest wybrany z grupy złożonej z bydła, świni, wielbłąda, kozy, owcy, psa, osła i konia. [002] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym transgenicznego ssaka uzyskuje się ze szczepu użytecznego do wytwarzania przeciwciał monoklonalnych
10 lub zmodyfikowanego genetycznie tak, żeby stał się odpowiedni do wytwarzania przeciwciał monoklonalnych. [0026] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym transgeniczne zwierzę uzyskuje się ze szczepu użytecznego do wytwarzania przeciwciał poliklonalnych lub zmodyfikowanego genetycznie tak, żeby stał się odpowiedni do wytwarzania przeciwciał poliklonalnych. [0027] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym immunoglobuliną jest IgG. [0028] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym immunoglobuliną jest IgM. [0029] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym zwierzę jest transgeniczne pod względem wytwarzania immunoglobulin ludzkich lub humanizowanych. [00] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym sekwencja kwasu nukleinowego kodująca łańcuch alfa białka FcRn jest zmutowana [0031] i w którym mutacja powoduje, że miejsce wiązania z albuminami tego białka FcRn staje się niefunkcjonalne. [0032] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, zastosowanie lub zwierzę, w którym sekwencja kwasu nukleinowego koduje chimerowe białko FcRn. [0033] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym wzmożoną
11 aktywność FcRn zapewnia się przez zintegrowanie do genomu zwierzęcia więcej niż jednej funkcjonalnej kopii sekwencji kwasu nukleinowego kodującej FcRn. [0034] W korzystnym przykładzie wykonania przedmiotem niniejszego wynalazku jest sposób, w którym konstrukt genetyczny zawiera bydlęcy insert klonu BAC nr 128E04 lub króliczy insert klonu BAC nr 262E02. Szczegółowy opis wynalazku [003] W niniejszym zgłoszeniu opisano sposób wytwarzania zwierzęcia Tg innego niż człowiek, zdolnego do rozwijania wzmożonej humoralnej odpowiedzi immunologicznej przeciwko antygenowi w porównaniu z nietransgenicznym zwierzęciem kontrolnym tego samego gatunku, obejmujący wprowadzanie do organizmu tego zwierzęcia innego niż człowiek konstruktu genetycznego, zapewniającego wzmożoną aktywność związanego z klasą I MHC noworodkowego receptora Fc (FcRn). [0036] Bender i wsp. (Bender i wsp., 04) ujawnili zwierzę Tg, uzyskane przez wprowadzenie genu kodującego łańcuch alfa bydlęcego białka FcRn do organizmu myszy FVB/N z zastosowaniem bydlęcego klonu BAC nr 128E04, w celu zbadania regulacji transkrypcyjnej i czynności bydlęcego FcRn in vivo. [0037] W US06/03194 i Petkova i wsp. (Petkova i wsp., 06) ujawniono Tg myszy knock-out, zawierające homozygotyczne przerwanie FcRn i ludzki transgen FcRn (hfcrn). Ten dodatek istotnie wydłuża czas półtrwania podawanej egzogennie IgG ludzkiej, przy czym ekspresja hfcrn jest podobna do endogennego poziomu ekspresji. Autorzy wspominają również, bez podawania jakichkolwiek szczegółów, że ekspresja FcRn na poziomie istotnie
12 wyższym od endogennego mogłaby być korzystna. Spekuluje się, że poziom ekspresji można zwiększać od do 0 razy w stosunku do endogennego poziomu ekspresji poprzez zastosowanie silnych wektorów ekspresyjnych. W tych opisach ujawniono kilka odrębnych konstruktów, używanych do uzyskania zwierząt Tg. W pierwszym przypadku użyto 33-kb ludzkiego klonu kosmidu, obejmującego pełny gen hfcrn plus sekwencje flankujące -kb i -kb 3. Autorzy określili takie zwierzę Tg jako transgeniczną linię 32 genomowego hfcrn. W drugim przypadku, linia transgeniczna 276 cdna" obejmuje konstrukt genetyczny, zawierający łańcuch alfa hfcrn, klonowany do wektora E, zawierającego wzmacniacz CMV i kurzy promotor -aktyny. W trzecim przypadku, ich zwierzęta transgeniczne posiadają konstrukt genetyczny, obejmujący 34-kb fragment XhoI, w którego skład wchodzi pełny gen hfcrn z biblioteki ludzkiego pochodzenia bakteryjnych chromosomów sztucznych (Bacterial Artificial Chromosomes - BAC) firmy Genome Systems, Inc. Autorzy, jak widać, wytworzyli kilka linii myszy Tg przez bezpośrednią manipulację genetyczną (jak w przypadku myszy C7BL/6J i BXSB/MpJ) lub konwencjonalne krzyżowanie (na przykład ze szczepami MRL/MpJ i NZM24). [0038] Lu i wsp. ujawnili myszy Tg, posiadające w sutkach bfcrn, które uzyskali przez zastosowanie konstruktu ze swoistymi wobec sutków elementami regulatorowymi (Lu i wsp., 06). Uzyskano kilka linii myszy o różnej liczbie kopii bfcrn, w zakresie od 1 do 1, i z liczbą kopii bydlęcego 2m w zakresie od 1 do. Badano ekspresję i dokumentowano zmienność poziomu
13 IgG w surowicy. Ten system zapewnia jednak tylko ograniczoną ekspresję bfcrn po wystąpieniu laktacji, w komórkach nabłonkowych sutka w gronkach, i był wykorzystywany do badania transportu IgG w sutku w okresie laktacji. Zwierząt tych nie wykorzystywano do analizy odpowiedzi immunologicznej. Konstruktami stosowanymi do uzyskiwania tych myszy Tg były dwa wektory ekspresyjne, w których kozi promotor betakazeiny reguluje transkrypcję genów będących przedmiotem zainteresowania i genów wprowadzonych. Jeden wektor zawiera sekwencje kodujące łańcuch ciężki ( ) (pbc1-bfcrn), a drugi kodujące łańcuch lekki (pbc1-b 2m) bydlęcego FcRn. Konstruktami genetycznymi wprowadzanymi do organizmów zwierząt Tg są fragmenty uzyskiwane przez trawienie enzymami NotI i SalI, przez co powstaje łańcuch ciężki o długości 16,9 kilo par zasad (kb) i łańcuch lekki o długości 16,1 kb, które podawano w formie mikroiniekcji do zapłodnionych jaj myszy Kunming White w równym stężeniu. [0039] Yoshida i wsp. (Yoshida i wsp., 06) zbadali rolę mysiego FcRn (mfcrn) w nabłonku jelitowym w zakresie odporności przeciwko drobnoustrojom. Uzyskali oni linie FcRn myszy Tg, w których mfcrn i m 2m ulegały swoistej ekspresji przez komórki nabłonka jelita z wykorzystaniem swoistego tkankowo promotora genu wiążącego jelitowe kwasy tłuszczowe (IFABP) do uzyskania myszy IFABP-mFcRnTg/m 2mTg. Wywołano nadmierną ekspresję także 2m, żeby zapewnić, że nie zachodziło zjawisko ograniczenia substratu w odniesieniu do ekspresji transgenu FcRn. Również w tym przypadku zwierzęta Tg poddano dalszemu krzyżowaniu
14 1 1 2 wstecznemu z myszami FcRn-/- i następnie na BALB/c lub C7BL/6 (CD4.2+). [0040] W dokumencie WO01708 opisano klonowanie i określanie charakterystyki cdna FcRn przeżuwaczy oraz potencjalne ich zastosowanie do uzyskiwania zwierząt transgenicznych (ewentualnie w skojarzeniu z transgenem beta2-mikroglobuliny). [0041] W dokumencie EP opisano sposób wytwarzania przeciwciała, oparty na zwiększonym wytwarzaniu przeciwciał obserwowanym po transfekcji komórki wytwarzającej przeciwciała (na przykład hybrydoma) z zastosowaniem wektora ekspresyjnego FcRn. [0042] W dokumencie WO9919 opisano transgen kierujący ekspresją cząsteczki immunostymulującej (na przykład IL-6) swoiście w błonie śluzowej. [0043] W rozumieniu niniejszego opisu, termin zwierzę inne niż człowiek obejmuje zwierzęta z rzędu kręgowców, korzystnie ssaka. Zwierzęta inne niż człowiek, do których niniejszy wynalazek ma zastosowanie, można wybierać na podstawie kryteriów funkcjonalnych jako zwierzęta zdolne do produkcji immunoglobulin lub ich funkcjonalnych równoważników. W korzystnych przykładach wykonania do zwierząt według wynalazku należą między innymi myszy, szczury, inne gryzonie, króliki, świnie, wielbłądowate, owce, kozy, bydło, osły i konie. W jeszcze korzystniejszym przykładzie wykonania zwierzęciem jest mysz, królik, świnia, owca, koza i bydło. W zakresie niniejszego wynalazku pozostaje wykorzystanie dowolnego zwierzęcia innego niż człowiek, powszechnie stosowanego w stanie techniki do wytwarzania immunoglobulin: przeciwciał
15 monoklonalnych lub poliklonalnych surowic odpornościowych. [0044] W rozumieniu niniejszego opisu, wyrażenie zwierzę kontrolne tego samego gatunku lub nietransgeniczne zwierzę kontrolne tego samego gatunku stosuje się do określenia zwierzęcia, którego nie poddano procedurom związanym z uzyskiwaniem zwierzęcia Tg. Zwierzę kontrolne tego samego gatunku jest jednak leczone i traktowane w sposób podobny, jak zwierzę Tg w czasie procedur koniecznych do wytworzenia przeciwciał poliklonalnych lub monoklonalnych, takich jak na przykład pierwszorazowa immunizacja, immunizacja lub immunizacje przypominające, jeżeli są konieczne, i w trakcie całego procesu wytwarzania przeciwciał. [004] W rozumieniu niniejszego opisu termin immunoglobuliny odnosi się do glikoprotein z nadrodziny immunoglobulin, działających jak przeciwciała. Terminy przeciwciało i immunoglobulina są w niniejszym opisie stosowane wymiennie. Przeciwciała są to białka gospodarza, zawierające jeden z głównych efektorów adaptacyjnego układu immunologicznego. Ich funkcja jest wykorzystywana, ponieważ są one powszechnie stosowane jako odczynnik diagnostyczny i naukowe. Stają się one również ważnym narzędziem terapeutycznym w arsenale klinicysty, służąc do leczenia chorób. Przeciwciała wykorzystuje się do analizy, oczyszczania i wzbogacania, jak również do pośredniczenia w reakcjach fizjologicznych lub do ich modulowania. Zdolność przeciwciał do wiązania antygenu z dużym stopniem powinowactwa i swoistości doprowadziła do ich
16 powszechnego wykorzystania w rozmaitych dyscyplinach naukowych i medycznych. Interakcja antygen-przeciwciało ma kluczowe znaczenie dla naturalnej funkcji biologicznej przeciwciała, jak również dla jego zastosowania jako odczynnika naukowego lub terapeutycznego. [0046] Jeżeli chodzi o strukturę, przeciwciała są globulinami (w regionie elektroforezy białek). U ssaków istnieje pięć typów przeciwciał: IgA, IgD, IgE, IgG, i IgM (skrót Ig oznacza immunoglobulinę i jest także stosowany w niniejszym opisie do określenia przeciwciał). Klasyfikuje się je według różnic w zakresie domen stałych ich łańcuchów ciężkich. Poszczególne klasy immunoglobulin różnią się swymi właściwościami biologicznymi i ewoluowały w kierunku wywierania efektu na różne antygeny. IgA znajduje się w okolicach zawierających śluz (na przykład w jelicie, w drogach oddechowych lub w drogach moczowo-płciowych); zapobiega ona kolonizacji okolic pokrytych błoną śluzową przez patogeny. IgD działa głównie jako receptor antygenów na komórkach B. IgE wiąże się z alergenami i wyzwala uwalnianie histaminy z komórek tucznych (mechanizm leżący u podłoża alergii), jak również zapewnia ochronę przed robakami jelitowymi. IgM ulega ekspresji na powierzchni komórek B i jest także wydzielana w celu eliminowania patogenów we wczesnych stadiach odporności zależnej od komórek B. IgG odpowiada za większość opartej na przeciwciałach odporności przeciwko patogenom atakującym organizm. [0047] W korzystnym przykładzie wykonania niniejszego wynalazku wytwarzaną immunoglobuliną jest IgG,
17 jakkolwiek zakres wynalazku nie jest ograniczony do izotypu (klasy) IgG; bierze się pod uwagę również to, że IgG może zawierać modyfikacje, jak omówiono poniżej. [0048] IgG można klasyfikować na różne formy lub podklasy, które są efektem duplikacji genów u różnych ssaków. Istnieje wiele podklas IgG u gryzoni, ludzi, udomowionych przeżuwaczy, koni, świń, wielbłądowatych i świnek morskich, przy czym poszczególne podklasy wykazują odmienne właściwości biologiczne. Ludzkie IgG1 i IgG3 znajdują się na przykład w komórkach fagocytarnych ze względu na ich duże powinowactwo do Fc R1. IgG1 i IgG3 mogą być zatem szczególnie istotne: (a) w usuwaniu małych kompleksów IgG-Ag i (b) w dodatnim lub ujemnym stymulowaniu rozwoju komórek B i wytwarzania przeciwciał. IgG3 jest silnym aktywatorem dopełniacza, a IgG1 jest główną IgG surowicy, preferencyjnie transportowaną przez łożysko (Janeway Jr. i wsp., 01). Dobrze wiadomo, że zmienność podklas u ssaków bywa bardzo różna. Króliki posiadają tylko jeden gen IgM, natomiast myszy i ludzkie wykazują ekspresję czterech IgG, a konie - siedmiu. Bydło posiada trzy IgG, u świń zaś istnieje pięć domniemanych genów podklas IgG. Jest prawdopodobne, że analogiczne różnice funkcji biologicznych i ekspresji względnej będą obecne w zakresie podklas immunoglobulin u nie tak dobrze przebadanych ssaków (Butler, 06). [0049] Niniejszy wynalazek można wykorzystywać do zwiększania wytwarzania dowolnego, jednego lub więcej niż jednego, podtypu IgG bez swoistych ograniczeń. Możliwe jest jednak ściślejsze powiązanie izotypu IgG wytworzonego przez zwierzę-biorcę z białkiem FcRn,
18 19 1 obejmującym swoisty, egzogenny łańcuch alfa FcRn, nie zaś z białkiem FcRn obejmującym drugi egzogenny łańcuch alfa FcRn. Wykorzystanie transgenu swoistego łańcuch alfa FcRn do wzbogacenia ilości lub procentowej zawartości konkretnego podtypu IgG w surowicy odpornościowej, wytwarzanej przez selekcję transgenu FcRn o optymalnej zdolności wiązania i aktywności w odniesieniu do tego podtypu IgG, pozostaje w zakresie niniejszego wynalazku. [000] W innym korzystnym przykładzie wykonania niniejszego wynalazku wytwarzaną immunoglobuliną jest IgM. IgM stanowi około % całości immunoglobulin osocza i jest główną składową przeciwciał fazy wczesnej, wytwarzanych przeciwko antygenom błony komórkowej, wykazującym złożoną antygenowość, przeciwko drobnoustrojom zakaźnym i antygenom rozpuszczalnym. Jeżeli chodzi o strukturę, IgM in vivo ma strukturę pentameru. Pięć podjednostek, tworzących pentamerową strukturę IgM, ma strukturę czterołańcuchową, podobną do IgG. Jedną z głównych różnic w porównaniu ze strukturą IgG jest to, że IgM cechuje się zawartością łańcucha ciężkiego. Dodatkowo, immunoglobuliny te różnią się tym, że łańcuch zawiera o jedną domenę 2 więcej w swym regionie stałym, niż łańcuch, a IgM zawiera łańcuch polipeptydowy zwany łańcuchem J, który nie jest obecny w IgG i, jak się uważa, ułatwia polimeryzację łańcuchów przed wydzieleniem IgM z komórek produkujących przeciwciała. W stanie techniki znane są próby wytworzenia metodami rekombinacyjnymi IgM funkcjonalnie równoważnych produktowi naturalnemu (zob. na przykład US ). W przeciwieństwie do
19 1 2 metod, w których usiłowano zwiększyć wytwarzanie IgM przez wprowadzenie do organizmu zwierzęcia heterologicznych lub w inny sposób zmodyfikowanych genów immunoglobulin, przedmiotem niniejszego wynalazku jest rozwiązanie alternatywne, w którym zapewnia się zwierzę Tg, którego organizm zawiera konstrukt genetyczny zapewniający wzmożoną aktywność FcRn; takie zwierzę z kolei wytwarza w odpowiedzi na antygen podwyższony poziom endogennego IgM. [001] W rozumieniu niniejszego opisu termin humoralna odpowiedź immunologiczna odnosi się do procesu w organizmach żywych, w którym wytwarzane są przeciwciała w odpowiedzi na cząsteczki i organizmy, które na koniec neutralizują i(lub) eliminują. Swoistość odpowiedzi przeciwciał jest regulowana przez komórki T i(lub) B poprzez receptory związane z błoną, które wiążą antygen o pojedynczej swoistości. Po związaniu odpowiedniego antygenu i otrzymaniu rozmaitych innych sygnałów aktywujących, limfocyty B dzielą się, w wyniku czego powstają komórki B pamięci, jak również klony końcowo różnicujące się do wydzielających przeciwciał komórek plazmatycznych; każdy klon produkuje przeciwciała rozpoznające epitop antygenowy identyczny z epitopem rozpoznawanym przez jego receptor antygenowy. Limfocyty B pamięci pozostają w uśpieniu do momentu, gdy zostaną potem aktywowane przez swoisty dla nich antygen. Te limfocyty stanowią komórkową podstawę pamięci i późniejszego nasilenia odpowiedzi wytwarzania przeciwciał po ponownej ekspozycji na swoisty antygen. [002] W odniesieniu do humoralnej odpowiedzi immunologicznej termin wzmożony odnosi się do
20 odpowiedzi immunologicznej, w której poziom i poliklonalność przeciwciał powstających w odpowiedzi na dany antygen jest istotnie wyższy, rozwój odpowiedzi immunologicznej jest istotnie szybszy lub odpowiedź immunologiczna w odpowiedzi na dany antygen jest silniejsza w porównaniu z takimi samymi właściwościami przeciwciał powstających w odpowiedzi na dany antygen lub z odpowiedzią immunologiczną powstającą wtedy, gdy kontrolne zwierzę tego samego gatunku styka się z tym samym antygenem. [003] W rozumieniu niniejszego opisu termin immunizacja odnosi się do procesu, poprzez który osobnik jest eksponowany na czynnik, często skojarzony z adiuwantami, przy czym ten proces wzmacnia układ immunologiczny osobnika przeciwko temu czynnikowi. Czynnik określa się mianem immunogenu lub antygenu. Immunizacja oznacza to samo, co inokulacja i szczepienie, jako że w inokulacji i szczepieniu, podobnie jak w immunizacji, wykorzystuje się żywy czynnik zakaźny. Oprócz procesu immunizacji wstępnej, stwierdzono, że skuteczność immunizacji można ulepszyć przez okresowe powtarzanie wstrzyknięcia czynnika: proces ten określa się mianem immunizacji przypominającej. [004] W odniesieniu do immunizacji wyrażenie podwyższony poziom immunoglobuliny odnosi się do stężenia w surowicy konkretnej immunoglobuliny, które jest istotnie wyższe, niż poziom tej samej swoistej immunoglobuliny w surowicy kontrolnego zwierzęcia tego samego gatunku, co mierzy się poprzez siłę odpowiedzi immunologicznej, na przykład na podstawie szczytowego
21 uzyskiwanego w wyniku tego procesu miana przeciwciała. Ten podwyższony poziom jest zwykle o co najmniej 0% wyższy, korzystnie o 7% wyższy, korzystniej o 0% wyższy, jeszcze korzystniej nawet o % wyższy. [00] Wynik immunizacji w dużym stopniu zależy od natury antygenu. Powszechnie przyjmuje się, że każda substancja, która może wywoływać odpowiedź immunologiczną, jest immunogenna i nazywa się ją immunogenem. Istnieje wyraźne praktyczne rozróżnienie między immunogenem a antygenem. Antygen definiuje się jako dowolną substancję, która może wiązać się ze swoistym przeciwciałem. Wszystkie antygeny mogą zatem wywoływać produkcję swoistych przeciwciał, lecz niektóre z nich muszą w tym celu być połączone z immunogenem. Oznacza to, że chociaż wszystkie immunogeny są antygenami, nie wszystkie antygeny są immunogenne. Antygenami najczęściej stosowanymi w immunologii doświadczalnej są białka; przeciwciała przeciwko białkom są niezwykle użyteczne w biologii doświadczalnej i medycynie. Oczyszczone białka nie zawsze są jednak wysoce immunogenne; w celu uzyskania odpowiedzi immunologicznej muszą być podawane z adiuwantem. Węglowodany, kwasy nukleinowe i inne typy cząsteczek są potencjalnymi antygenami, lecz często odpowiedź immunologiczną będą wywoływać tylko wtedy, gdy są połączone z nośnikiem białkowym. Immunogenność antygenów białkowych determinuje zatem skuteczność absolutnie każdej odpowiedzi immunologicznej. Droga podania antygenu wpływa zarówno na wielkość, jak i typ uzyskiwanej odpowiedzi immunologicznej. Najczęstszymi drogami podawania antygenów do organizmu są
22 wstrzyknięcia do tkanki poprzez podskórne, śródskórne, domięśniowe, dożylne lub dootrzewnowe wstrzyknięcie lub przetoczenie. Podawanie doustne polega na dostarczeniu antygenu do przewodu pokarmowego, natomiast podawanie donosowe lub wziewne dostarcza antygen do dróg oddechowych. [006] Przeciwciała mogą rozpoznawać jako antygeny niemal każdego rodzaju cząsteczki biologiczne, w tym proste metabolity pośrednie, cukry, lipidy, autakoidy i hormony, jak również makrocząsteczki takie, jak złożone węglowodany, fosfolipidy, kwasy nukleinowe i białka. Tylko jednak makrocząsteczki są zdolne do stymulowania limfocytów B do zapoczątkowywania humoralnej odpowiedzi immunologicznej. Drobnocząsteczkowe substancje chemiczne, takie jak dinitrofenol, mogą wiązać się z przeciwciałami, lecz nie są w stanie same aktywować komórek B (to znaczy nie są immunogenne). W celu uzyskania przeciwciał swoistych wobec takich drobnocząsteczkowych substancji chemicznych immunolodzy zwykle przed immunizacją łączą je z makrocząsteczkami. W tych przypadkach drobnocząsteczkowa substancja chemiczna zwana jest haptenem, a makrocząsteczka zwana jest nośnikiem. Kompleks hapten-nośnik, w przeciwieństwie do wolnego haptenu, może działać jako immunogen (Abbas i Lichtman, 03). Podobne i inne protokoły immunizacji są powszechnie znane w stanie techniki; specjalista będzie w stanie łatwo wybrać odpowiednią strategię i wykorzystać wynalazek w praktyce. [007] Pierwotna odpowiedź immunologiczna zwykle powoduje tylko niewielkie, nieistotne podwyższenie
23 poziomu zarówno swoistych przeciwciał wytwarzanych przeciwko wprowadzonemu immunogenowi/antygenowi, jak i poziomu immunoglobulin całkowitych. Wielkość odpowiedzi immunologicznej zależy od podanej dawki immunogenu. Poniżej pewnej dawki progowej większość białek nie wywołuje żadnej odpowiedzi immunologicznej. Powyżej dawki progowej dochodzi do stopniowego zwiększania odpowiedzi w miarę zwiększania dawki antygenu, do osiągnięcia szerokiego poziomu plateau, po którym następuje zmniejszenie przy bardzo dużych dawkach antygenu. Ogólnie do wtórnej i późniejszej odpowiedzi immunologicznej dochodzi przy mniejszych dawkach antygenu; osiąga się wtedy wyższe wartości plateau, co jest oznaką pamięci immunologicznej. Dlatego właśnie zwykle do uzyskania istotnie podwyższonego miana swoistego przeciwciała niezbędna jest immunizacja przypominająca. Liczba immunizacji przypominających może być różna w zależności od kilku czynników, takich jak immunogenność antygenu, typ adiuwantu immunologicznego, droga i schemat immunizacji (Stills, 0). [008] Ponieważ większość antygenów ma charakter wysoce złożony, prezentują one liczne epitopy, które są rozpoznawane przez dużą liczbę limfocytów. Każdy limfocyt jest aktywowany do proliferacji i różnicowania do komórek plazmatycznych; uzyskaną odpowiedzią przeciwciał jest wytwarzanie przeciwciał poliklonalnych (PAb). Ważną cechą adaptacyjnej odpowiedzi immunologicznej jest pamięć immunologiczna. Pamięć immunologiczna powstaje wskutek immunizacji wstępnej,
24 2 1 2 czyli pierwotnej, która wywołuje pierwotną odpowiedź immunologiczną. [009] Większość białek jest słabo immunogenna lub nieimmunogenna, gdy są one podawane same. Silna adaptacyjna odpowiedź immunologiczna na antygeny białkowe zawsze wymaga wstrzyknięcia antygenu w mieszaninie zwanej adiuwantem. Adiuwant jest to dowolna substancja, która zwiększa immunogenność substancji z nią zmieszanych. Adiuwanty różnią się od nośników białkowych tym, że nie tworzą stabilnego sprzężenia z immunogenem. Po stymulacji adiuwantem, immunogeny także immunogeny dające słabą reakcję czy nawet niedające reakcji będą wyzwalać istotną odpowiedź immunologiczną po podaniu dawki przypominającej immunogenu wymagającego adiuwantu w nieobecności adiuwantu (Janeway Jr. i wsp., 01; Leenaars i Hendriksen, 0; Lipman i wsp., 0; McCullough i Summerfield, 0; Schunk i Macallum, 0; Stills, 0). [0060] Odpowiedź na każdą kolejną immunizację jest coraz silniejsza, tak więc drugo-, trzeciorzędowa i kolejnego rzędu odpowiedzi będą miały rosnące nasilenie. Wielokrotna ekspozycja na antygen celem uzyskania stanu wzmożonej odporności zwana jest hiperimmunizacją. Wiele przeciwciał stosowanych w technikach klinicznych i immunochemicznych są to przeciwciała poliklonalne, wytwarzane przez hiperimmunizację odpowiedniego zwierzęcia, takiego jak na przykład gryzonie, królik, koza, osioł lub owca, zawiesiną odpowiedniego antygenu. Na szczycie wytwarzania przeciwciał zbiera się surowicę; tą metodą
25 można uzyskać stężenie swoistej immunoglobuliny G (IgG) wynoszące od około 1 do mg/ml surowicy. Mimo że dowiedziono, że przeciwciała monoklonalne (mab) są źródłem o dobrze poznanych właściwościach, małej immunogenności i dużej skuteczności, nadal jednak jest miejsce na przeciwciała poliklonalne. Podobnie jak naturalny układ immunologiczny w odpowiedzi na patogeny wykorzystuje raczej przeciwciała poliklonalne niż mab, przeciwciała poliklonalne mogą być również w wielu przypadkach korzystne w immunoterapii biernej. Do korzyści przeciwciał poliklonalnych należą ich potencjalnie zwiększona siła działania w zakresie tworzenia kompleksów immunologicznych, użyteczność w zwalczaniu chorób zakaźnych, wywoływanych przez różne szczepy patogenów lub chorób wymagających do powodzenia leczenia neutralizacji wielu epitopów. Przeciwciała poliklonalne są również korzystne w technikach immunochemicznych, ponieważ są zazwyczaj względnie łatwe w wytwarzaniu i znacznie tańsze w produkcji. Przeciwciała poliklonalne można ponadto uzyskiwać u rozmaitych gatunków, takich jak królik, koza, owca, osioł, kura i inne, co zapewnia użytkownikom szeroki wybór przy opracowywaniu planów doświadczeń. Ilość przeciwciał swoistych w preparacie poliklonalnym stanowi jednak niekiedy zaledwie niewielką część całkowitej zawartości białek przeciwciał. Głównym celem wytwarzania przeciwciał w tych celach jest zatem uzyskanie wysokiego miana surowic odpornościowych o dużym powinowactwie. [0061] W protokołach hiperimmunizacji, powszechnie stosowanych w stanie techniki, do utrzymania wysokiego
26 poziomu IgG niezbędne są regularne immunizacje przypominające, ponieważ u zdrowych zwierząt katabolizm immunoglobulin jest zwiększony, jak wykazano już bardzo dawno w badaniach metabolizmu IgG (Andersen i Bjorneboe;, 1964). Przedmiotem niniejszego wynalazku jest natomiast skuteczniejszy protokół uzyskiwania stabilnie podwyższonego poziomu wytwarzania immunoglobulin, w którym wysoki poziom IgG utrzymuje się przez względnie długi czas bez kolejnych immunizacji lub przy znacznie rzadszych immunizacjach. [0062] W wytwarzaniu przeciwciał poliklonalnych można zidentyfikować pewną liczbę etapów o kluczowym znaczeniu, które mogą wpływać na wynik doświadczenia na zwierzętach na przykład na wyniki testów immunologicznych oraz na ból i cierpienie zwierząt. Gdy antygen, przeciwko któremu produkuje się przeciwciała, jest słabo immunogenny, do indukcji skutecznej odpowiedzi immunologicznej układ immunologiczny wymaga bodźca. W tym celu można stosować adiuwanty, i można ukierunkowywać odpowiedź immunologiczną bardziej w stronę odpowiedzi komórkowej lub bardziej w stronę odpowiedzi humoralnej. Mimo, że opisano ponad 0 adiuwantów, rutynowo do wytwarzania przeciwciał poliklonalnych wykorzystuje się zaledwie kilka adiuwantów (na przykład kompletny adiuwant Freunda (FCA), niekompletny adiuwant Freunda, sole glinu, Quil A, Iscoms, Montanide, TiterMax, RIBI itd.). Do wytwarzania przeciwciał poliklonalnych często stosuje się FCA, ponieważ pozwala on uzyskać wysokie miano przeciwciał przeciwko niemal wszystkim typom antygenów. Wielu badaczy opisywało jednak ciężkie objawy
27 niepożądane po wstrzyknięciu na przykład adiuwantów FCA, TiterMax i RIBI. Nasilenie zmian patologicznych zależy nie tylko od adiuwantu, lecz także od typu stosowanego antygenu. Ponadto alternatywne adiuwanty często nie indukują skutecznej odpowiedzi przeciwciał. Stwierdzono, że dodatkowy wpływ na zakres wywołanych zmian ma objętość wstrzyknięcia (Leenaars i Hendriksen, 0). Po uwzględnieniu tych czynników, przedmiotem niniejszego wynalazku jest skuteczniejszy protokół, umożliwiający rzadszą immunizację, co jest dużą korzyścią w odniesieniu do dobrostanu zwierzęcia, oprócz waloru ekonomicznego, ponieważ ten protokół zmniejsza ból i dyskomfort zwierząt, pozwalając jednocześnie uzyskać optymalną odpowiedź immunologiczną. [0063] Zgodnie z tym, przedmiotem niniejszego wynalazku jest opisany w zastrzeżeniach sposób wytwarzania immunoglobulin poliklonalnych, obejmujący wykorzystanie zwierzęcia Tg, którego organizm zawiera konstrukt genetyczny zapewniający wzmożoną aktywność FcRn, według dowolnego ustalonego protokołu immunizacji, umożliwiającego wytwarzanie immunoglobulin poliklonalnych. Zwierzę transgeniczne korzystnie uzyskuje się ze szczepu użytecznego do wytwarzania przeciwciał poliklonalnych lub zmodyfikowanego genetycznie tak, żeby stał się bardziej odpowiedni do wytwarzania przeciwciał poliklonalnych. [0064] W przeciwieństwie do PAb, przeciwciała monoklonalne (MAb) są to przeciwciała wytwarzane przez pojedynczy klon limfocytów B. W połowie lat 70 XX wieku Kohler i Milstein opracowali technikę wytwarzania
28 przeciwciał monoklonalnych o pożądanej swoistości, za co otrzymali nagrodę Nobla (Kohler i Milstein, 197). Po kilkudziesięciu latach zbierania doświadczeń i wykorzystywania technik z dziedziny MAb specjalista powinien być odpowiednio biegły w zakresie wszelkich aspektów wytwarzania przeciwciał monoklonalnych. W celu uzyskania przeciwciała monoklonalnego swoistego wobec zdefiniowanego antygenu, mysz, szczura lub królika immunizuje się tym antygenem. Można stosować komórki B, takie jak splenocyty, limfocyty węzłów chłonnych, inne limfocyty krwi obwodowej lub limfocyty pochodzące z innych tkanek zwierzęcia. Gospodarza-ssaka można poddawać dodatkowym immunizacjom w celu dalszego zwiększenia liczebności populacji komórek B o pożądanej swoistości antygenowej B, jak również w celu wzmożenia swoistości antygenowej. Te izolowane komórki B poddaje się następnie fuzji z odpowiednią, unieśmiertelnioną linią komórkową. Linie szpiczaka są najlepszymi partnerami fuzyjnymi komórek B, ponieważ komórki mają tendencję do ulegania fuzji i tworzenia stabilnych hybryd, skuteczniejszych niż komórki. Komórka nieśmiertelna jest to komórka limfoblastoidalna lub komórka szpiczaka plazmocytowego, na przykład komórka szpiczaka, która jest złośliwą komórką wytwarzającą przeciwciała. Przesiewa się nadsącze hybrydom, żeby wybrać optymalne hybrydomy o korzystnych właściwościach wiązania antygenu. Wybrane hybrydomy klonuje się i poddaje kriokonserwacji. [006] W tym kontekście w odniesieniu do przeciwciał monoklonalnych, określenie zwierzę odnosi się do każdego ssaka innego niż człowiek, w tym między innymi
29 1 2 do królików, myszy, szczurów, kóz, owiec, świnek morskich, świń i krów. [0066] Oprócz technik konwencjonalnych lub klasycznych, zastosowanie technologii rekombinacyjnych zapoczątkowało nową erę w wytwarzaniu przeciwciał poliklonalnych i monoklonalnych: obecnie możliwe jest poddawanie przeciwciał obróbce (na przykład zmniejszanie przeciwciała do funkcjonalnej wielkości lub wytwarzanie przeciwciał humanizowanych) (Lonberg, 0; Peterson, 0). [0067] Zgodnie z tym, przedmiotem niniejszego wynalazku jest określony w zastrzeżeniach sposób wytwarzania immunoglobulin monoklonalnych, obejmujący wykorzystanie zwierzęcia Tg, którego organizm zawiera konstrukt genetyczny zapewniający wzmożoną aktywność FcRn według dowolnego ustalonego protokołu immunizacji, umożliwiający wytwarzanie immunoglobulin monoklonalnych. Transgeniczny ssak inny niż człowiek korzystnie pochodzi ze szczepu nadającego się do wytwarzania przeciwciał monoklonalnych lub zmodyfikowanego genetycznie tak, żeby stał się bardziej odpowiedni do wytwarzania przeciwciał monoklonalnych. [0068] W rozumieniu niniejszego opisu, termin transgeniczny w kontekście wyrażenia zwierzę transgeniczne odnosi się do zwierzęcia innego niż człowiek, posiadającego gen lub inną sekwencję kwasu nukleinowego, której nie dałoby się uzyskać na drodze normalnej hodowli ani krzyżowania. Termin gen odnosi się do fragmentu kwasu nukleinowego, wykazującego ekspresję mrna, funkcjonalnego RNA lub swoistego białka, w tym do sekwencji regulatorowych. Termin gen
30 natywny odnosi się do genu takiego, jak występuje w naturze. Termin transgen odnosi się do genu, który wprowadzono do genomu przez transformację i który jest stabilnie utrzymywany. W tym kontekście, termin transformacja stosuje się w niniejszym opisie w szerokim znaczeniu do opisania wprowadzania obcego DNA do komórki. Termin ten ma także obejmować wszelkie inne, funkcjonalnie równoważne metody wprowadzania obcego DNA do komórki, takie jak na przykład mikroiniekcja, transfekcja, infekcja, transdukcja lub fuzja komórki donorowej i komórki akceptorowej. Transgeny mogą obejmować na przykład geny heterologiczne lub homologiczne do genów konkretnego zwierzęcia, które ma być transformowane. Dodatkowo transgeny mogą obejmować geny natywne, wprowadzane do organizmu nienatywnego, lub geny chimerowe. Termin gen endogenny odnosi się do genu natywnego w jego naturalnej lokalizacji w genomie organizmu. [0069] Wprowadzenia transgenu do zwierzęcia-biorcy dokonuje się zwykle przez zastosowanie konstruktu genetycznego, zawierającego transgen będący przedmiotem zainteresowania. W rozumieniu niniejszego opisu, termin konstrukt genetyczny ma obejmować sztucznie stworzony, rekombinowany DNA, zawierający sekwencje kwasu nukleinowego, który, po wprowadzeniu do komórki biorcy, zapewnia ekspresję tych wprowadzonych sekwencji kwasu nukleinowego. Konstrukt genetyczny może obejmować sekwencje kodujące i sekwencje regulatorowe. Termin sekwencja kodująca odnosi się do sekwencji DNA lub RNA, kodującej swoistą sekwencję aminokwasową, i nie obejmuje sekwencji niekodujących. Termin sekwencje
31 regulatorowe odnosi się do sekwencji nukleotydowych znajdujących się w górę od sekwencji kodującej (sekwencje niekodujące ), w obrębie sekwencji kodującej lub w dół sekwencji kodującej (sekwencje niekodujące 3 ), wpływających na transkrypcję, obróbkę lub stabilność RNA albo translację powiązanych sekwencji kodujących. Do sekwencji regulatorowych należą wzmacniacze, promotory, sekwencje liderowe translacji, introny i sekwencje sygnału poliadenylacji. Należą do nich sekwencje naturalne i syntetyczne, jak również sekwencje, które mogą być kombinacją sekwencji syntetycznych i naturalnych. Niektóre sekwencje regulatorowe użyteczne w niniejszym wynalazku będą obejmować między innymi promotory konstytutywne, promotory swoiste tkankowo, promotory swoiste dla stadium rozwojowego, promotory indukowalne i promotory wirusowe. [0070] Termin promotor odnosi się do sekwencji nukleotydowej, zwykle położonej w górę ( ) w stosunku do jej sekwencji kodującej, regulującej ekspresję sekwencji kodującej poprzez zapewnienie rozpoznawania polimerazy RNA i innych czynników niezbędnych do poprawnej transkrypcji. Termin promotor obejmuje promotor minimalny, którym jest krótka sekwencja DNA, złożona zwykle z TATA-box i inne sekwencje, służące do dokładnego określenia miejsca inicjacji transkrypcji, do których dodawane są elementy regulatorowe celem kontroli ekspresji. Pojęcie promotor odnosi się także do sekwencji nukleotydowej, obejmującej promotor minimalny plus elementy regulatorowe, zdolne do regulowania ekspresji sekwencji kodującej lub
32 funkcjonalnego RNA. Tego typu sekwencja promotorowa składa się z elementów proksymalnych i elementów położonych bardziej dystalnie w górę: te ostatnie elementy często są określane mianem wzmacniaczy. Zgodnie z tym, wzmacniacz jest to sekwencja DNA, która może stymulować aktywność promotora i może być elementem wrodzonym promotora lub elementem heterologicznym, wprowadzonym celem podwyższenia poziomu lub zwiększenia swoistości tkankowej promotora. Jest on zdolny do działania w obu orientacjach (normalnej lub odwróconej) oraz do działania nawet wtedy, gdy ulegnie przesunięciu w górę lub w dół od promotora. Zarówno wzmacniacze, jak i inne położone w górę elementy promotora wiążą się ze swoistymi dla sekwencji białkami wiążącymi DNA, które pośredniczą w ich działaniu. Promotory mogą pochodzić w całości z genu natywnego lub mogą być złożone z różnych elementów, pochodzących z różnych promotorów występujących w naturze, albo nawet mogą składać się z syntetycznych segmentów DNA. Promotor może również zawierać sekwencje DNA, biorące udział w wiązaniu czynników białkowych, regulujących skuteczność inicjacji transkrypcji w odpowiedzi na warunki fizjologiczne lub rozwojowe. Regulacja transkrypcji może być także zależna od obecności różnych środków chemicznych, hormonów, induktorów i tym podobnych. Specjalista będzie w stanie łatwo wybrać i zestawić sekwencje regulatorowe najlepiej nadające się do danego zastosowania. [0071] Technologia transgenów jest powszechnie wykorzystywana w badaniach naukowych; opracowano kilka
33 różnych protokołów. Mimo, że w stanie techniki znane są doniesienia o ograniczeniach konwencjonalnych technik transgenicznych, poziom ekspresji transgenu wykazuje różnice między liniami (Palmiter i wsp., 1984), a niekiedy nawet między zwierzętami z tej samej linii (Dobie i wsp., 1996; Sutherland i wsp., 00), zwłaszcza wtedy, gdy stosuje się cdna, nie zaś fragmenty genomu. Zróżnicowana ekspresja transgenu zależy od miejsc integracji transgenów do genomów gospodarz i może niekorzystnie wpływać na ekspresję transgenu (Opsahl i wsp., 03). Ograniczenia mikroiniekcji transgenu na bazie plazmidu można przezwyciężać przez wykorzystanie układów klonowania z DNA o wielkości mniejszej niż milion par zasad, takich jak YAC (sztuczny chromosom drożdży), BAC (sztuczny chromosom bakteryjny) lub PAC (sztuczny chromosom faga P1). Techniki te są dobrze sprawdzone przy wytwarzaniu myszy Tg (metody opisali Giraldo i Montoliu, 01). Specjalista będzie w stanie przeprowadzić etapy niezbędne do wytworzenia i określenia charakterystyki opisywanego tu zwierzęcia Tg na podstawie stanu techniki i niniejszego ujawnienia. [0072] W korzystnym przykładzie wykonania opisywany tu konstrukt genetyczny obejmuje wstawkę (insert) bydlęcego klonu BAC nr 128E04. Klon BAC nr 128E04 jest to klon z bydlęcej biblioteki BAC, na którą składa się DNA z grzebienia płciowego płodu męskiego byka rasy Holstein wysoko plasującego się w hierarchii, uzyskany z bydlęcej biblioteki BAC INRA, (Eggen i wsp., 01). Wstawkę klonu nr 128E04 definiuje się jako segment chromosomu 18 bydlęcego genomu między pozycjami
34 3 1 2 nukleotydowymi i 3624 tak, jak są one przedstawione w NCBI Map Viewer, Bos taurus (krowa) Build 3.1 (na podstawie Btau 3.1), stan aktualizacji z dnia 11 sierpnia 07. [0073] W korzystnym przykładzie wykonania opisywany tu konstrukt genetyczny obejmuje wstawkę króliczego klonu BAC nr 262E02. Klon BAC nr 262E02 izolowano z biblioteki króliczego BAC ((Rogel-Gaillard i wsp., 01). Bibliotekę BAC skonstruowano w wektorze pbelobac11, DNA o dużej masie cząsteczkowej wytworzono z krwinek białych królika nowozelandzkiego. Biblioteka króliczego BAC jest prowadzona przez centrum zasobów zwierząt domowych INRA i jest publicznie dostępna. [0074] Opisywana tu swoista modyfikacja, obecna u zwierzęcia Tg, zapewnia nadmierną ekspresję genu kodującego łańcuch alfa białka o aktywności FcRn. W rozumieniu niniejszego opisu, termin nadmierna ekspresja odnosi się do poziomu ekspresji, który jest wyższy niż poziom oczekiwany na podstawie dwóch kopii genomowych genu będącego przedmiotem zainteresowania u danego gatunku. Nadmierną ekspresję można oceniać na kilku poziomach procesów biochemicznych, na przykład na poziomie transkrypcji, translacji, modyfikacji posttranslacyjnych itd., pod warunkiem że podwyższony poziom ekspresji powoduje wykrywalne zmiany czynności FcRn w porównaniu z podstawowym poziomem ekspresji u zwierzęcia typu dzikiego. Białko kodowane przez gen będący przedmiotem zainteresowania może na przykład ulegać ekspresji w większej ilości, jak w kilku opisywanych tu przykładach, łańcuch alfa białka o
35 aktywności FcRn jest wytwarzany na podwyższonym poziomie. [007] Nadmierną ekspresję można osiągać kilkoma różnymi środkami, które są dobrze znane specjaliście z dziedziny biologii molekularnej. Przykładowymi sposobami uzyskania nadmiernej ekspresji genu będącego przedmiotem zainteresowania jest zwiększenie liczby kopii genu, zwiększenie siły wiązania regionu promotora, regulacja w górę elementów wzmacniających albo, przeciwnie, zahamowanie lub zablokowanie elementów represorowych, i tym podobne. [0076] U korzystnych opisywanych tu zwierząt transgenicznych nadmierna ekspresja nie zależy od miejsca integracji, zależy natomiast od liczby zintegrowanych kopii. Specjalista będzie jednak w stanie określić, kiedy korzystne lub konieczne będzie zastosowanie którychkolwiek z dostępnych elementów genetycznych w celu uzyskania ograniczonej ekspresji transgenu. W stanie techniki znane są liczne przykłady indukowalnych elementów regulatorowych, jak również elementów regulatorowych zależnych od tkanek, narządów, stadium rozwoju itd., umożliwiających precyzyjne regulowanie ekspresji genu będącego przedmiotem zainteresowania i są one do dyspozycji specjalisty. [0077] Jak tu opisano, nadmierną ekspresję można osiągnąć przez integrację więcej niż jednej funkcjonalnej kopii genu będącego przedmiotem zainteresowania do genomu zwierzęcia Tg według wynalazku. Korzystnie, od zwierząt izoluje się fragmenty DNAy zawierające gen łańcucha alfa FcRn. Takie duże fragmenty DNA można izolować przez
36 przesiewanie biblioteki kosmidów, YAC lub BAC, i tym podobnych, uzyskanych z genomowego DNA zwierzęcia innego niż człowiek. Klony YAC mogą zawierać fragmenty DNA o wielkości do 2 milionów par zasad; klony BAC mogą zawierać fragment DNA mniejszej wielkości (około - kb). Zwierzę stanowiące źródło DNA może należeć do dowolnego gatunku, na przykład do gatunku odgrywającego istotną rolę w wytwarzaniu przeciwciał poliklonalnych lub monoklonalnych na skalę handlową, takiego jak mysz, szczur, królik, owca, koza, bydło, świnia, osioł i koń. Jest oczywiste, że dobór źródła transgenu ma charakter nieograniczający: na podstawie niniejszego ujawnienia specjalista jest w stanie określić, czy dany gen nadaje się do wykorzystania w sposobach według niniejszego wynalazku. [0078] Procedury wprowadzania transgenu do organizmu zwierzęcia-biorcy i wyboru zwierząt Tg są dobrze znane specjaliście. Pokrótce, transgeniczne wektory, zawierające gen łańcucha alfa FcRn, wprowadza się do komórki lub komórek biorcy, po czym integruje do genomu komórki lub komórek biorcy przez integrację losową lub ukierunkowaną. W przypadku integracji losowej, wektor transgeniczny, zawierający locus FcRn, można wprowadzać do komórki zwierzęcia-biorcy standardowymi technikami transgenicznymi. Wektor transgeniczny można na przykład bezpośrednio wstrzykiwać do przedjądrza zapłodnionego oocytu. Wektor transgeniczny można również wprowadzać poprzez koinkubację spermy z wektorem transgenicznym przed zapłodnieniem oocytu. Zwierzęta transgeniczne mogą się rozwijać z zapłodnionych oocytów.
37 [0079] Innym sposobem wprowadzania wektora transgenicznego jest transgeneza lentiwirusowa. Ta niedawno opracowana metoda aczkolwiek ograniczona co do wielkości transgenu okazała się bardzo skuteczna w tworzeniu zwierząt Tg wielu gatunków, takich na przykład mysz, szczur, świnia, i jest narzędziem szczególnie obiecującym przy opracowywaniu strategii terapii genowej (Pfeifer, 06). Innym sposobem wprowadzania wektora transgenicznego jest transfekcja zarodkowych komórek macierzystych i następnie wstrzykiwanie genetycznie zmodyfikowanych zarodkowych komórek macierzystych do rozwijających się zarodków. Na koniec z zarodków uzyskuje się chimerowe zwierzęta Tg, u których transgen FcRn jest zintegrowany do genomu co najmniej niektórych komórek somatycznych. W przypadku integracji ukierunkowanej, do komórek odpowiedniego zwierzęcia-biorcy, takich jak zarodkowe komórki macierzyste lub już zróżnicowane komórki somatyczne, można wprowadzać wektor transgeniczny. [0080] W swoistych przykładach wykonania integracja transgenu może doprowadzić do utraty odpowiedniego locus endogennego łańcucha alfa FcRn poprzez wykorzystanie procedur rekombinacji homologicznej. Zamiast tego natywny locus FcRn można usunąć na drodze knock-out niezależnie od wprowadzenia transgenu. Pożądane zwierzę można uzyskać na przykład poprzez klasyczne sposoby hodowli i krzyżowania, wykorzystując rodziców z genotypami z knock-out. Zastąpienie endogennego locus FcRn nie jest jednak konieczne do spełnienia celu wynalazku. Specjalista będzie w stanie określić, czy obecność genu endogennego wywiera
38 negatywny wpływ na wytwarzanie immunoglobulin na podwyższonym poziomie. Kiedy wprowadza się wiele kopii genu kodującego łańcuch alfa białka o aktywności FcRn, żeby uzyskać nadmierną ekspresję genu, w normalnych warunkach nie ma jednak potrzeby stosowania takiej strategii zastępowania. [0081] Jeżeli istnieje potrzeba wzbogacenia swoistego izotypu IgG, który wiąże się ściślej z FcRn z egzogennego łańcucha alfa FcRn, nie zaś jego endogennego odpowiednika, wtedy korzystna jest delecja (knock-out) endogennego FcRn i(lub) zastąpienie go przez egzogenny łańcuch alfa FcRn. Ważnym przykładem jest nadprodukcja ludzkiego IgG u zwierząt Tg, posiadających ludzki chromosomalny DNA, kodujący łańcuch ciężki i lekki ludzkich genów immunoglobulin. Można przewidywać, że u zwierząt nadmierna ekspresja na przykład bydlęcego łańcuch alfa FcRn i delecja łańcucha alfa FcRn gospodarza jest korzystna w odniesieniu do wzbogacania w ludzką IgG kosztem IgG gospodarza, która słabiej wiąże się z egzogennym łańcuchem alfa FcRn i przez to jest szybciej usuwana z organizmu zwierzęcia. Podobnym przykładem jest sytuacja, gdy korzystne jest wzbogacenie w jeden z izotypów IgG gospodarza kosztem innych izotypów IgG. [0082] Wybrane komórki można następnie poddawać fuzji z pozbawionymi jądra komórkami jednostki transferu jądrowego, na przykład oocytami. Fuzję przeprowadza się konwencjonalnymi technikami, które są dobrze znane w stanie techniki (zob. na przykład (Cibelli i wsp., 1998). Pozbawienie oocytów jąder i transfer jądrowy można również przeprowadzać metodami
39 mikrochirurgicznymi, stosując pipety do wstrzyknięć (zob. na przykład Wakayama i wsp., 1998)). Uzyskane komórki jajowe hoduje się następnie w odpowiedniej pożywce i przenosi do zsynchronizowanych biorców celem uzyskania zwierząt Tg. Ewenrualnia wybrane, zmodyfikowane genetycznie zarodkowe komórki macierzyste można wstrzykiwać do rozwijających się zarodków, które następnie rozwijają się do zwierząt chimerowych. [0083] Opisuje się również transgeniczne zwierzę inne niż człowiek, takie, że zwierzę Tg posiada wielokrotne kopie transgenu. Teoretycznie nie przewiduje się ograniczenia liczby kopii transgenu wprowadzanych do organizmu zwierzęcia Tg. Specjalista będzie w stanie łatwo określić, czy nadmierna ekspresja genu kodującego łańcuch alfa białka o aktywności FcRn zapewnia względnie dużą ekspresję FcRn, zapewniającą korzystne efekty wynalazku bez jednoczesnej ryzykownie dużej ekspresji genu, która zakłócałaby homeostazę i funkcjonalność komórki. W przedstawionych tu przykładach nie obserwowano modyfikacji fenotypowych wśród różnych linii Tg zwierząt, posiadających do kopii pełnego genu bfcrn (Bender i wsp., 07). Dodatkowo Lu i wsp. (Lu i wsp., 07) przedstawili dane dotyczące linii Tg posiadających do 1 kopii cdna bfcrn pod kontrolą promotora swoistego dla sutka, bez istotnych modyfikacji fenotypowych. [0084] W rozumieniu niniejszego opisu, termin aktywność FcRn stosuje się do określenia serii zdarzeń zachodzących in vivo. Jak już omówiono przy opisie stanu techniki, FcRn po raz pierwszy zidentyfikowano u gryzoni jako receptor przekazujący
40 immunoglobuliny macierzyste od matki do noworodka poprzez jelito noworodkowe. Badania wykazały następnie, że FcRn odgrywa centralną rolę w regulowaniu transportu IgG w obrębie komórek różnego pochodzenia i pomiędzy takimi komórkami. W kontekście niniejszego opisu termin aktywność FcRn odnosi się do głównie do ochrony IgG przed rozpadem. Zgodnie z tym aktywność FcRn, w rozumieniu niniejszego opisu, definiuje się jako zdolność do wiązania IgG-Fc i ochrony IgG przed rozpadem. [008] Ponadto, co po raz pierwszy ujawnia się w niniejszym opisie, termin aktywność FcRn w rozumieniu niniejszego opisu odnosi się także do zdolności FcRn do nasilania humoralnej odpowiedzi immunologicznej, a bardziej szczegółowo, do nasilania swoistej antygenowo klonalnej ekspansji komórek B i w następstwie tego, syntezy IgM i IgG. [0086] Aktywność FcRn można na przykład określać na jednym lub więcej niż jednym etapie procesu ochrony IgG, której mechanizm, jak się uważa, jest regulowany przede wszystkim przez komórki śródbłonka, wyściełające naczynia krwionośne. Wewnątrz tych komórek FcRn znajduje się głównie we wczesnych/ulegających recyklingowi endosomach, gdzie napotyka IgG internalizowaną przez endocytozę fazy płynnej. Kwaśne środowisko endosomów ułatwia interakcję. Związana IgG i albumina ulegają recyklingowi z powrotem na powierzchnię i są uwalniane z komórki, natomiast niezwiązane ligandy są transportowane w dół i ulegają rozpadowi w lizosomach.
41 [0087] Niektóre z etapów można jednak modelować i definiować niezależnie od ogólnego znaczenia terminu. [0088] Zdolność wiązania IgG będącej przedmiotem zainteresowania lub cząsteczki zawierającej domenę stałą IgG lub jej fragment Fc z FcRn można określać w różnych testach in vitro. W WO 97/34631 ujawniono szczegółowo rozmaite metody. [0089] Jednym ze sposobów określania wiązania między IgG a FcRn jest test in vitro, w którym wykorzystuje się izolowany kompleks FcRn i 2m, zachowujący wykazywaną in vivo funkcję wiązania IgG-Fc przy ph około 6, z uwolnieniem związanego IgG-Fc zachodzącego po przesunięciu wartości ph do około 7,2. Termin około w rozumieniu niniejszego opisu w odniesieniu do ph oznacza oznaczoną wartość ph ±0,2. Kompleks FcRn/ 2m jest związany z podłożem stałym, takim jak studzienka do mikromiareczkowania, filtr, błona, kolumna lub kulki. Materiałami często wykorzystywanymi jako podłoże stałe są nylon, polistyren, polipropylen i agaroza. Zetknięcie składników cząsteczkowych osiąga się zwykle przez dodanie składników razem do roztworu wodnego, zwykle odpowiednio zbuforowanego, i pozostawienie składników do przereagowania ze sobą przez określony z góry czas. W zależności od planu badania między dodawaniem kolejnych składników konieczne mogą być etapy płukania. Te składniki służą do określenia, że doszło do wiązania między składnikami, i można je ustawiać według kilku dobrze znanych formatów testu, takich jak testy wiązania kompetycyjnego, testy bezpośredniego wiązania, testy kanapkowe i tym podobne. Gdy wszystkie konieczne etapy
42 wiązania się odbędą, przystępuje się do wykrywania z zastosowaniem sposobów zwykle wykorzystywanych w stanie techniki, na przykład przez wykrywanie sygnału, który może być sygnałem radioaktywnym, enzymatycznym, fluorescencyjnym lub innym dobrze znanym sygnałem. Zmierzony sygnał jest wprost proporcjonalny do zmierzonego sygnału (jak w testach wiązania bezpośredniego) lub jest kompetycyjny. [0090] Ogólny sposób ustalania wiązania między IgG a FcRn można dogodnie adaptować tak, żeby umożliwić identyfikację wiązania odpowiedniej pary łańcucha alfa FcRn i(lub) 2m, tworzącej idealny heterodimer FcRn, z IgG będącą przedmiotem zainteresowania. W tym przypadku w teście stosuje się łańcuch alfa FcRn pochodzący z różnych gatunków lub będący wynikiem mutacji oraz 2m wiążący się z największym powinowactwem z IgG będącą przedmiotem zainteresowania. Najpierw stosuje się IgG, o której wiadomo, że wiąże się z FcRn (na przykład ludzka IgG, która wiąże się z bfcrn), w warunkach odpowiednich do wiązania. Po tym wstępnym etapie prowadzi się doświadczenia, w których początkowo zastosowany łańcuch alfa FcRn będzie zastępowany łańcuchem alfa FcRn z różnych gatunków lub łańcuchem alfa FcRn uzyskiwanym na drodze mutagenezy in vitro, po czym oceniane będzie wiązanie z daną IgG. Najlepszy łańcuch alfa FcRn można zidentyfikować przez porównanie wiązania kompleksu powinowactwa z wiązaniem z początkowym kompleksem wiązania. Podobny sposób można zastosować do zidentyfikowania najlepszego z możliwych 2m do połączenia w parę z określonym z góry łańcuchem alfa FcRn. Zamiast lub oprócz tego można zastosować
43 44 metody o wysokiej przepustowości do jednoczesnego przebadania kilku par kombinacji alfa-fcrn i 2m. 1 2 Izolowany kompleks FcRn i 2m, zachowujący in vivo czynność wiązania IgG-Fc, korzystnie wytwarza się przez syntezę in vitro z poddanych obróbce kwasów nukleinowych, kodujących odpowiednie białka. Białka można wytwarzać oddzielnie, po czym łączyć razem celem uzyskania kompleksu. Segmenty DNA, kodujące takie białka, można włączać do wektora rekombinacyjnego w takiej pozycji, żeby wektor stał się zdolny do ekspresji białka. Techniki takiej manipulacji segmentów DNA, poprzez na przykład metody inżynierii genetycznej z wykorzystaniem endonukleaz restrykcyjnych, będą znane specjalistom po zapoznaniu się z niniejszym opisem i piśmiennictwem, takim jak na przykład publikacja Ausubel i wsp., Specjaliści zdają sobie sprawę z tego, że istnieją dodatkowe sposoby, które można wykorzystać do uzyskania kompleksu nadającego się do wykorzystania w opisanej tuż powyżej metodzie in vitro. Zamiast tego składniki endogennego kompleksu można izolować z odpowiedniego źródła komórkowego. [0091] Powinowactwo IgG będącej przedmiotem zainteresowania do FcRn można mierzyć na podstawie pomiarów powierzchniowego rezonansu plazmonowego (SPR), stosując na przykład BIAcore 00 (BIAcore Inc.), jak opisano uprzednio (Karlsson i wsp., 1991; Popov i wsp., 1996). W tym sposobie cząsteczki FcRn sprzęga się z chipem sensorowym BIAcore (na przykład z chipem CM firmy Pharmacia) i mierzy się wiązanie danej IgG z unieruchomionym FcRn przy pewnej prędkości przepływu, żeby uzyskać sensorogramy, stosując oprogramowanie do
44 4 1 2 oceny B1A 2.1., na podstawie czego można obliczyć prędkość on i off danej IgG, jej domen stałych lub ich fragmentów, wobec FcRn. [0092] Względne powinowactwo IgG będącej przedmiotem zainteresowania lub jej fragmentów do FcRn można również zmierzyć z użyciem na przykład testu wiązania kompetycyjnego. IgG będącą przedmiotem zainteresowania dodaje się w różnych ilościach do studzienek 96- studzienkowej płytki, w której unieruchamia się FcRn. Następnie do każdej studzienki dodaje się stałą ilość wyznakowanej radioaktywnie IgG będącej przedmiotem zainteresowania. Sporządza się wykres procentowej radioaktywności związanej frakcji w stosunku do ilości IgG typu dzikiego, i na podstawie nachylenia krzywej można obliczyć jej względne powinowactwo. Powinowactwo IgG będącej przedmiotem zainteresowania lub jej fragmentów do FcRn można również mierzyć z użyciem testu saturacji i analizy Scatcharda lub innymi sposobami, na przykład metodą obliczenia regresji nieliniowej (dopasowanie krzywej). [0093] Transfer danej IgG lub jej fragmentów między komórkami przez FcRn można mierzyć z użyciem testu transferu in vitro, stosując wyznakowaną radioaktywnie IgG lub jej fragmenty i komórki wykazujące ekspresję FcRn, i porównując radioaktywność jednej strony pojedynczej warstwy komórek z radioaktywnością drugiej strony. [0094] Innym testem, umożliwiającym zidentyfikowanie FcRn z ochrony IgG in vivo, jest test hodowli komórkowej. Wytwarza się, lub identyfikuje z uprzednio istniejących komórek ssaka, komórki ssaka, wykazujące
45 funkcjonalną ekspresję FcRn w hodowli. Komórkami nadającymi się do zastosowania w tym teście są komórki zdolne do katabolizmu IgG, przy czym ekspresja FcRn w tych komórkach powoduje zmniejszenie katabolizmu. Funkcjonalna ekspresja FcRn, w kontekście testu in vivo, wskazuje, że łańcuch alfa FcRn tworzy kompleks z 2m, wiąże się z IgG będącą przedmiotem zainteresowania i chroni związaną IgG przed rozpadem. W celu zidentyfikowania odpowiedniego łańcucha alfa FcRn, serię komórek wykazujących ekspresję różnych łańcuchów alfa FcRn i IgG będącą przedmiotem zainteresowania styka się z komórkami. Komórki inkubuje się w odpowiednich warunkach, umożliwiających normalną czynność komórek, po czym bada pod kątem katabolizmu IgG. Istotne zmniejszenie katabolizmu IgG w komórkach wykazujących ekspresję FcRn w porównaniu z katabolizmem IgG w komórkach kontrolnych, które nie wykazują ekspresji FcRn, wskazują, że dany łańcuch alfa FcRn chroni IgG przed rozpadem. Ten test wykrywa czynność FcRn poprzez wiązanie z IgG, internalizację i ochronę. Do testu hodowli komórkowej można zastosować linie komórkowe znane w stanie techniki, jak również komórki pierwotne gatunku wybranego do wytwarzania przeciwciał po immunizacji. Ten test ma kluczowe znaczenie w badaniu hybrydowych cząsteczek łańcucha alfa FcRn, złożonych z części pozakomórkowej o lepszym wiązaniu z IgG i segmentu przezbłonowego i cytoplazmatycznego, swoistego dla gatunku, który będzie zastosowany do immunizacji. [009] Jak wspomniano w odniesieniu do opisu stanu techniki, FcRn jest cząsteczką heterodimerową, złożoną
46 47 z łańcucha alfa podobnego do klasy I MHC i mikroglobuliny ( 2m). Łańcuch alfa FcRn (zatwierdzony symbol: FCGRT, zatwierdzona nazwa: receptor Fc, IgG, transporter łańcucha alfa, fragment Fc receptora immunoglobuliny G; noworodkowy receptor Fc, łańcuch alfa FcRn) nadaje cząsteczce jej swoistą charakterystykę, natomiast 2m jest powszechnie występującym składnikiem, obecnym w kilku różnych kompleksach białkowych. Dla specjalisty jest oczywiste, że w sposobach według niniejszego wynalazku muszą być obecne obie podjednostki heterodimeru, w odpowiednich ilościach. Cechą definiującą uzyskiwania podwyższonego poziomu immunoglobulin według wynalazku jest nadmierna ekspresja genu kodującego łańcuch alfa cząsteczki FcRn, jak opisano powyżej. Oczywiste jest jednak, że do uzyskania funkcjonalnego FcRn konieczne są ekwimolarne ilości drugiego łańcucha 2m. Przy wykorzystaniu wynalazku specjalista powinien zatem wziąć pod uwagę dostępność komórkową 2m. Korelacja między oczekiwanym poziomem heterodimeru FcRn na podstawie modyfikacji genetycznych dokonanych u zwierzęcia Tg a zmierzoną aktywnością FcRn powinna wskazywać na ewentualne problemy wynikające z niedostatecznej podaży 2m. Metody oceny obu czynników są dostępne dla specjalisty w stanie techniki i w niniejszym opisie. Podobnie, gdy jest to konieczne, specjalista może zwiększać poziom 2m na przykład przez zastosowanie metod transgenicznych znanych w stanie techniki. W swoistych przykładach wykonania niniejszy wynalazek można ulepszyć przez zapewnienie zwierząt Tg z nadmierną
47 48 ekspresją zarówno łańcucha alfa FcRn, jak i łańcucha mikroglobuliny z tych samych lub różnych konstruktów genetycznych i(lub) przez te same lub różne konstrukty genetyczne. [0096] Jak jasno wynika z powyższego rozdziału, w którym omówiono aktywność FcRn, skład heterodimeru może być istotnym czynnikiem przy ustalaniu jego aktywności. Zgodnie z tym przedmiotem niniejszego wynalazku jest również dowolna korzystna kombinacja łańcucha alfa i 2m, pozwalająca uzyskać podwyższony poziom immunoglobuliny w surowicy zwierzęcia Tg. Implikacje charakterystyki wiązania i swoistego powinowactwa FcRn omówiono szczegółowo powyżej; te właściwości heterodimeru FcRn są jednak również określone przez skład podjednostkowy. Jak omówiono w opisie stanu techniki, siła wiązania między bydlęcym FcRn a ludzką IgG jest większa, niż między ludzkim FcRn a ludzką IgG. Bardzo ważnym aspektem jest zatem zapewnienie cząsteczek FcRn o charakterystyce wiązania ulepszonej w stosunku do natywnej cząsteczki FcRn zwierzęcia Tg. Cel ten można w szczególności osiągnąć przez badanie i wybór optymalnej kombinacji łańcucha alfa i 2m FcRn do wytwarzania immunoglobuliny u zwierzęcia Tg. Można to osiągnąć przez wprowadzenie odpowiedniego łańcucha alfa lub 2m z różnych gatunków zwierząt lub z tego samego gatunku. Specjalista będzie w stanie wprowadzić konieczne modyfikacje u zwierzęcia Tg po określeniu charakterystyki wiązania, jak omówiono tu szczegółowo. Zgodnie z tym, gdy niniejsze zgłoszenie odnosi się do zwierzęcia Tg, ma to oznaczać zwierzę, w którego genom nie tylko włączono gen kodujący łańcuch alfa białka o
48 aktywności FcRn, lecz, jeżeli jest to konieczne, gen kodujący łańcuch alfa białka o aktywności FcRn i gen kodujący 2-mikroglobulinę. [0097] Opisano także osiąganie wzmożenia aktywności FcRn przez wprowadzenie do organizmu zwierzęcia innego niż człowiek konstruktu genetycznego kodującego 2m. Specjalista będzie w stanie określić warunki konieczne do osiągnięcia tego wzmożenia. Wszystkie rozważania, omówione w odniesieniu do łańcucha alfa FcRn, będą się odnosić, mutatis mutandi, do modyfikacji obejmujących 2-mikroglobulinę ( 2m). [0098] Opisano także zastosowanie zmutowanych wersji łańcucha alfa lub 2m, o ile modyfikacje powodują powstanie funkcjonalnej cząsteczki FcRn, spełniającej opisane powyżej wymagania co do aktywności. Techniki mutacji są dobrze znane specjaliście z dziedziny biologii molekularnej. Przy definiowaniu odpowiedniego białka FcRn, zamiast wymogów dotyczących aktywności, można określić homologię strukturalną między mutantem a białkiem natywnym. Należy tu podkreślić, że homologia strukturalna jest dostępną, lecz nieograniczającą cechą transgenu. Łańcuch alfa białka FcRn może wykazywać tożsamość sekwencji wynoszącą około 60% lub więcej z sekwencją bydlęcego białka FcRn, stosowanego według niniejszego wynalazku. Terminy tożsamość sekwencji, homologia i wariant stosuje się w niniejszym zgłoszeniu wymiennie. Gdy jeden aminokwas określa się jako homologiczny do drugiego, oznacza to, że jedna z omawianych sekwencji aminokwasowych wykazuje co najmniej 60%, korzystnie co najmniej 70%, korzystniej co najmniej 80%, jeszcze korzystniej co najmniej 90%, i
49 0 1 2 jeszcze korzystniej co najmniej 9% tożsamość sekwencji z drugą sekwencja. W celu wykazania, że sekwencje są, na przykład, w co najmniej 60% identyczne, można wykorzystywać program FastDB EMBL lub bazy danych SWISSPROT. Do określania tej homologii można także stosować inne algorytmy i ich skomputeryzowane przykłady wykonania, dobrze znane w stanie techniki. [0099] W swoistym przykładzie wykonania zmutowana wersja łańcucha alfa białka FcRn jest pozbawiona funkcjonalnego miejsca wiązania z albuminami. Jak omówiono powyżej, FcRn zawiera dwa niezależne miejsca wiązania immunoglobuliny i albuminy. Albumina jest to cząsteczka o masie 67 kd, stanowiąca dwie trzecie masy białek surowicy. Transportuje ona wiele różnych cząsteczek, takich jak kwasy tłuszczowe, kwasy żółciowe, eikozanoidy, witaminy, hormony, jony, toksyny i leki. Albumina odpowiada również w większości za koloidalne ciśnienie osmotyczne krwi i jest głównym białkiem surowicy buforującym ph. W normalnych warunkach całkowity poziom IgG mieści się w zakresie -1 mg/ml (Manz i wsp., 0), a poziom albuminy wynosi mg/ml (Beers i Berkow, 1999). Około 7% całkowitego koloidalnego ciśnienia osmotycznego osocza jest skutkiem zawartości w niej albuminy, a 2% - globulin (głównie IgG) (Guyton, 1991). Jako że masa cząsteczkowa IgG jest niemal trzykrotnie większa niż albuminy (-160 w porównaniu do 67 kda), proste obliczenie ich stosunku molekularnego (który liczy się przy koloidalnym ciśnieniu osmotycznym) wskazuje, że zwiększenie o trzy jednostki IgG jest identyczne ze zwiększeniem o jedną jednostkę albuminy, jeżeli chodzi
50 1 1 2 o wpływ na ciśnienie onkotyczne osocza (jest to ciśnienie, które można przypisać frakcji białkowej surowicy). Nadmierna ekspresja FcRn w celu uzyskania większej ilości antygenowo swoistej IgG może być korzystna, może jednak prowadzić do szkodliwych zaburzeń, zwłaszcza jeżeli współistnieje z wyższym od prawidłowego poziomem albuminy. Jako że FcRn wiąże IgG i albuminę w dwóch różnych miejscach (Andersen i wsp., 06; Chaudhury i wsp., 06), a (jak wykazało niedawne badanie) delecja wskutek mutacji miejsca wiązania z albuminami w łańcuchu alfa FcRn nie prowadzi do utraty wiązania IgG in vitro (Andersen i wsp., 06), można stwierdzić, że nie istnieje kompetycja między tymi dwoma ligandami. [00] Nawet w przypadku braku tego efektu kompetycji, w zależności od liczby kopii FcRn obecnych u zwierzęcia Tg, poziom albuminy w surowicy może zwiększać się do poziomu tak wysokiego, że może to powodować problemy u zwierzęcia. Zgodnie z tym, zwierzę posiadające zmutowany FcRn z niefunkcjonalnym miejscem wiązania z albuminami będzie w stanie funkcjonować lepiej według wynalazku w zakresie wytwarzania korzystnych przeciwciał. Specjalista będzie w stanie łatwo określić typ swoistej mutacji, powodującej niewłaściwe działanie miejsca wiązania z albuminami, jak również przeprowadzić konieczne procedury z zakresu biologii molekularnej i inne etapy niezbędne do uzyskania zwierzęcia Tg bez zbędnego obciążenia pracami doświadczalnymi. [01] Jak już wspomniano w tym opisie, ogólny stan zdrowia zwierzęcia ma duże znaczenie w wykorzystywaniu
51 2 1 2 wynalazku w wykonalny ekonomicznie sposób. Wykazano, że zwierzęta Tg według niniejszego wynalazku żyją ponad rok bez objawów patologicznych. [02] W innym przykładzie wykonania niniejszego wynalazku, w sposobie według wynalazku wykorzystuje się zwierzę Tg, posiadające białko o aktywności FcRn, w skład którego wchodzi chimerowy łańcuch alfa, zawierający endogenną wewnątrzkomórkową domenę białka FcRn zwierzęcia Tg w skojarzeniu z pozakomórkową domeną obcego pochodzenia. Składnik - domenę pozakomórkową dobiera się w sposób opisany powyżej. W tym przykładzie wykonania chimerowy łańcuch alfa mógłby być pomocny w przezwyciężaniu sytuacji, gdy białko kodowane przez transgen nie działa poprawnie dlatego, że jego właściwości przekazywania sygnałów wewnątrzkomórkowych nie wykazują zgodności z właściwościami zwierzęciabiorcy. Spowodowałoby to brak funkcjonalności heterodimeru FcRn; ulepszona zdolność wiązania immunoglobulin obcego receptora powoduje jednak, że warto stosować taką chimerę. Specjalista będzie w stanie łatwo zaadaptować metody inżynierii genetycznej znane ze stanu techniki do wytworzenia chimerowego receptora według tego przykładu wykonania. [03] W innym opisywanym tu aspekcie zwierzę Tg uzyskuje się ze zwierzęcia, które jest już transgeniczne pod względem wytwarzania immunoglobulin, które są ludzkie lub humanizowane. Ze stanu techniki wiadomo, jak ważne jest wytwarzanie ludzkich lub humanizowanych immunoglobulin u zwierząt Tg, i od niedawna uzyskuje się takie zwierzęta, różnych gatunków. Jedna z technik, które bada się celem
52 3 1 2 uzyskania mab o małej immunogenności do terapii in vivo u ludzi, obejmuje zastosowanie myszy Tg wykazujących ekspresję repertuarów ludzkich sekwencji genów przeciwciał. W przyszłości być może uda się rozszerzyć tę technikę i stosować ją nie tylko u gryzoni, lecz także u hodowlanych zwierząt Tg (takich jak na przykład bydło, króliki) do bezpośredniego tworzenia i produkcji surowic poliklonalnych o sekwencji ludzkiej (Lonberg, 0). W US opisano wytwarzanie bydła Tg, obejmujące genetyczną modyfikację, powodującą unieczynnienie i utratę ekspresji jego endogennych przeciwciał i ekspresję przeciwciał ksenogennych, korzystnie przeciwciał ludzkich. Uzyskuje się to przez unieczynnienie ekspresji łańcucha ciężkiego bydlęcej IgM i ewentualnie przez unieczynnienie ekspresji łańcucha lekkiego bydlęcej Ig, i następnie przez wprowadzenie sztucznego chromosomu, powodującego ekspresję przeciwciał innych niż bydlęce, korzystnie przeciwciał ludzkich. W US ujawniono inne podejście do zwiększania ekspresji immunoglobulin u zwierząt transgenicznych innych niż ludzie przez nadmierną ekspresję inhibitora apoptozy, którego ekspresja jest kierowana przez promotor swoisty dla komórek B swoiście w komórce B zwierzęcia, zwiększając przez to przeżycie komórek B. W WO ujawniono humanizowane przeciwciała wytwarzane przez zwierzę Tg inne niż człowiek, które poddano manipulacji genetycznej tak, żeby posiadało jedno lub więcej niż jedno humanizowane locus immunoglobulinowe, zdolne do ulegania rearanżacji genów i konwersji genów u zwierzęcia Tg innego niż człowiek, w celu uzyskania
53 4 1 2 rozmaitych humanizowanych immunoglobulin. Niedawno opisano króliki Tg, wykazujące ekspresję repertuaru humanizowanych przeciwciał (Thorey i wsp., 06). [04] W niniejszym zgłoszeniu opisano istotne korzyści u tych zwierząt Tg w zakresie ulepszonego wytwarzania immunoglobulin. Dobór i wprowadzenie transgenu FcRn według procedur opisanych w niniejszym opisie mogłyby na przykład zapewnić, że wytwarzanie immunoglobulin u tych zwierząt Tg jest optymalizowane przez wykorzystanie najskuteczniejszego partnera wiązania FcRn dla immunoglobulin wytwarzanych przez zwierzębiorcę, niezależnie od tego, czy są to immunoglobuliny endogenne, czy wytwarzane transgenicznie przeciwciała ludzkie lub humanizowane. Wynalazek obejmuje zatem zastosowanie zwierząt już transgenicznych, wytwarzających immunoglobuliny ludzkie lub humanizowane, jako punktu wyjścia do wprowadzenia jednej lub więcej niż jednej kopii łańcucha alfa FcRn i zapewnienia korzyści płynących z metod według niniejszego wynalazku. Ewentualnie, podwójnie transgeniczne zwierzę o korzystnym fenotypie można uzyskać przez skrzyżowanie zwierzęcia Tg, wytwarzającego humanizowane immunoglobuliny, ze zwierzęciem uzyskanym w sposób tu opisywany. [0] W niniejszym wynalazku zapewnia się określony w zastrzeżeniach sposób uzyskiwania podwyższonego poziomu immunoglobulin w surowicy zwierzęcia, obejmujący uzyskanie wyżej opisanego zwierzęcia Tg i zastosowanie tego zwierzęcia w dowolnym ustalonym protokole immunizacji do wytwarzania immunoglobulin.
54 1 2 [06] W kolejnym aspekcie niniejszego wynalazku, wzmożenie odpowiedzi immunologicznej obejmuje nasilenie ekspansji antygenowo swoistych, klonalnych komórek B. Po raz pierwszy opisuje się tu rolę FcRn w modulowaniu odpowiedzi immunologicznej poprzez klonalną ekspansję komórek B i syntezę immunoglobulin. Jest to najbardziej uderzające i nieoczekiwane odkrycie. Większość badań, w których opisywano status odporności humoralnej u zwierząt pozbawionych funkcjonalnego FcRn, bądź we wcześniejszych badaniach z myszami z knock-out 2m, bądź ostatnio u zwierząt z knock-out w łańcuchu alfa FcRn, nie wykazywała zakłóceń syntezy IgG; niski poziom IgG w surowicy tych zwierząt definiowano na podstawie zaburzonej ochrony IgG (Junghans i Anderson, 1996; Roopenian i wsp., 03). W opisach innych badań donoszono jednak, że u zwierząt pozbawionych 2m synteza IgG jest zmniejszona, autorzy nie doszli jednak do definitywnego wyjaśnienia tego faktu (Ghetie i wsp., 1996; Israel i wsp., 199). [07] W niniejszym wynalazku wykazano, że 2 dni po immunizacji OVA występowało dwukrotnie więcej komórek wydzielających IgM swoiste wobec OVA i trzykrotnie więcej komórek wydzielających IgG swoiste wobec OVA u myszy Tg w porównaniu ze zwierzętami typu dzikiego (ang. wild type wt). Wzmożoną ekspansję klonalną antygenowo swoistych komórek B można częściowo wyjaśnić tym, że śledziony myszy Tg były istotnie większe w porównaniu z kontrolnymi zwierzętami wt, co także obserwowano. Za tezą o bardziej nasilonej reakcji immunologicznej przemawiają także profile odporności komórkowej, w których stwierdzano podobne zmiany w
55 6 1 2 proporcjach składników komórkowych po immunizacji u myszy Tg i wt, lecz myszy Tg wykazywały zmiany znacznie bardziej radykalne. W obu grupach stwierdzano proporcjonalne zmniejszenie liczby komórek wykazujących ekspresję B2, CD3, równolegle z istotnym, choć umiarkowanym wzrostem CD11b+ / CD11 c+/mhc11, i istotnie masywnym napływem komórek posiadających CD11b high / Gr-1 high, co sugeruje, że dominującą populacją komórkową, która napłynęła do śledziony w czasie wtórnej odpowiedzi immunologicznej były neutrofile i, w mniejszym stopniu, makrofagi i(lub) komórki dendrytyczne. Zgodnie z tym, w innym aspekcie sposobów według niniejszego wynalazku wzmożenie odpowiedzi immunologicznej obejmuje pobudzenie napływu neutrofili, makrofagów i(lub) komórek dendrytycznych do wtórnych narządów limfoidalnych. [08] Obwodowe, czyli wtórne narządy limfoidalne są to miejsca, gdzie zapoczątkowywana jest adaptacyjna odpowiedź immunologiczna i gdzie utrzymywane są limfocyty. Do tych narządów należą węzły chłonne, śledziona i narządy limfoidalne błony śluzowej. Zgodnie z tym, w korzystnych przykładach wykonania, wtórnym narządem limfoidalnym jest śledziona i węzły chłonne. [09] Chociaż w monocytach, makrofagach i komórkach dendrytycznych (Sachs i wsp., 06; Stirling i wsp., 0; Zhu i wsp., 01) obserwowano ekspresję FcRn, jego funkcja w tych komórkach jest nadal niejasna i przez to niejasna jest rola FcRn w prezentacji antygenu, co mogłoby mieć bezpośrednie konsekwencje w odniesieniu do syntezy immunoglobulin. Z drugiej strony, późniejsze badanie wykazało, że FcRn odgrywa
56 7 1 2 główną rolę w zależnej od IgG fagocytozie w leukocytach o wielokształtnym jądrze i w monocytach (Vidarsson i wsp., 06). Jako że ekspresję bfcrn wykrywano w neutrofilach i makrofagach pochodzących z otrzewnej myszy Tg, można postawić hipotezę, że nadmierna ekspresja FcRn w tych komórkach odpowiada za wzmożoną fagocytozę antygenów (w postaci kompleksów immunologicznych), a nawet za prezentację antygenów, co następnie powoduje powstanie we wtórnych narządach limfoidalnych większej liczby antygenowo swoistych komórek plazmatycznych, wytwarzających IgM i IgG. Jeżeli ta teoria jest poprawna, to wzmożenie byłoby oczywiste po powstaniu IgG, i nie wcześniej. Istotnie, podwyższenie miana IgM stwierdzano dopiero po pojawieniu się IgG swoistych wobec OVA, głównie we wtórnej odpowiedzi immunologicznej. Analiza profilu odporności komórkowej śledziony wykazała jednak, że dominującym typem komórek, które napływają do śledziony po immunizacji, są neutrofile. Z innego niedawno opublikowanego doniesienia (Maletto i wsp., 06) wynika, że neutrofile są główną populacją komórek zawierających antygen w narządach limfoidalnych, gdy tylko dochodzi do swoistej odpowiedzi immunologicznej i tworzy się kompleks immunologiczny IgG-antygen. Wykazano również, że neutrofile we wtórnych narządach limfoidalnych wykazują ekspresję głównie TNF-alfa i przyczyniają się do jakości ustalonej, wtórnej odpowiedzi immunologicznej. To odkrycie zapewnia wiarygodne wyjaśnienie tego, dlaczego u badanych przez autorów myszy wt i Tg stwierdza się istotny napływ komórek z CD11b high i Gr-1 high. Jeżeli weźmie się pod
57 8 1 2 uwagę, że myszy Tg wytwarzają znacznie więcej IgG swoistej wobec OVA i wykazują nadmierną ekspresję FcRn w tych komórkach, można również wyjaśnić, dlaczego myszy Tg wykazują większy napływ tych komórek do śledziony, znaczniejszą ekspansję klonalną antygenowo swoistych komórek B i w następstwie tego silniejszą odpowiedź przeciwciał w porównaniu z ich odpowiednikami - zwierzętami kontrolnymi typu dzikiego. [01] Ważnym aspektem niniejszego wynalazku jest to, że po mniejszej liczbie ekspozycji na antygen rozwija się taki sam poziom humoralnej odpowiedzi immunologicznej, jak u nietransgenicznego zwierzęcia kontrolnego tego samego gatunku. Ta cecha niniejszego wynalazku jest bardzo korzystna, gdy w tym samym czasie potrzebne są duże ilości surowic odpornościowych i gdy dużą liczbę zwierząt immunizuje się równolegle, niekiedy drogimi antygenami. Oszczędności mogą być znaczne, nawet jeżeli możliwe jest wyeliminowanie tylko jednego wstrzyknięcia przypominającego. [0111] Innym ważnym aspektem niniejszego wynalazku jest to, że taki sam poziom humoralnej odpowiedzi immunologicznej rozwija się szybciej w porównaniu z nietransgenicznym zwierzęciem kontrolnym tego samego gatunku. Specjalista będzie w stanie łatwo zaadaptować jakikolwiek odpowiedni protokół immunizacji, przy stosowaniu zwierzęcia w sposobie według wynalazku, i określić korzystny moment do zebrania wytworzonych przeciwciał. Gdy zwierzę wykorzystywane do wytwarzania przeciwciał wykorzystuje się wielokrotnie (jak w przypadku kozy, owcy, innych dużych zwierząt), ta cecha wynalazku będzie prowadzić do ulepszenia czasu cyklu
58 9 1 2 zbierania surowic odpornościowych, czego implikacje handlowe są ewidentne. Gdy zwierzę na koniec protokołu uśmierca się (jak w przypadku królików), korzyść handlowa jest nawet większa: producent przeciwciał, specjalizujący się w zapewnianiu określonych surowic odpornościowych wobec antygenów dostarczonych przez klienta, będzie w stanie szybciej dostarczyć zamówioną surowicę odpornościową, zapewniając klientowi korzyść pożądaną w dzisiejszym świecie, gdzie rozwój nauki ulega przyśpieszeniu i istnieje znaczna konkurencja. Krótki opis rysunków [0112] Fig. 1. Struktura i określanie charakterystyki bydlęcego transgenu BAC 128E04 A: Schematyczne przedstawienie fragmentu genomu BAC 128E04 z względnym położeniem bfcgrt i pięcioma domniemanymi genami kodującymi białka: FLT3LG, LOC39196 i LOC273, LOC11234, LOC1123. B: Brak naruszenia zintegrowanego transgenu wykrywano metodą PCR. I: DNA genomowe z transgenicznej linii nr 9; II: DNA genomowe z transgenicznej linii nr 14; III: DNA genomowe z transgenicznej linii nr 19 i IV: kontrolne matryce genomu DNA BAC 128E04. Przedziały: MM: drabinka o wielkości 1kb, 1: BAC 128E04, koniec, 2: FLT3LG, 3: LOC39196, 4: FCGRT, : LOC273, 6: LOC1 1234, 7: LOC1123, 8: PCR swoistego dla końca 3 BAC 128E04. Należy zauważyć, że LOC1 1234, LOC1123 i fragmenty swoiste dla końca 3 nie zostały powielone z transgenicznej linii nr 9 DNA genomowego. Fig. 2. Metafaza szerzy się z fibroblastów pochodzących z transgenicznych linii nr 14 i nr 19. Analiza FISH
59 wykazała, że fluorescencyjnie wyznakowany klon BAC 128E04m zawierający bfcgrt hybrydyzuje do całkowicie odmiennych segmentów chromosomów, w fibroblastach myszy, odpowiednio, nr 14 i nr 19, co wyklucza możliwość, że fenotyp transgenicznych szczepów myszy nr 14 i nr 19 jest zależny od miejsca integracji. Fig. 3. Standardowe krzywe zależności Ct od log liczby kopii (Q) w przypadku, odpowiednio, mysiej -aktyny i łańcuch alfa bfcrn (bfcgrt), służące do określenia bezwzględnej liczby kopii bfcrn u myszy Tg (linie nr 14 i nr 19) z zastosowaniem ilościowej PCR w czasie rzeczywistym. Ct jest to cykl, w którym wartość fluorescencji przecina wartość progową przy zastosowaniu PCR w czasie rzeczywistym. Wykrywanie przeprowadzono z zastosowaniem technologii fluorogennej nukleazy k (TaqMan, (Lee i wsp., 1993) na systemie ABI Prism7000 Sequence Detection System (Applied Biosystems Foster City CA, USA). Fig. 4. Ekspresja mrna swoistego wobec bfcrn zależna od liczby kopii transgenu. A: RT-PCR, odpowiednio, z próbek tkanki hemizygotycznych transgenicznych linii nr 9 (1, 4, 7, ), nr 14 (2,, 8, 11), nr 19 (3, 6, 9, 12). Przedziały: MW: drabinka o wielkości 1kb, 1-3 płuco; 4-6 wątroba; 7-9 jelito noworodka; -12 sutek. B: Analiza Northern służąca wykrywaniu ekspresji mrna łańcucha alfa bfcrn w próbkach RNA całkowitego z wątroby. Przedział 1: mysz typu dzikiego; 2: próbka bydlęca, 3-4: hemi- i homozygotyczne myszy linii nr 14 odpowiadają dwóm i czterem kopiom transgenu bfcrn; -6: hemi- i homozygotyczne myszy linii nr 19 odpowiadają pięciu i dziesięciu kopiom transgenu bfcrn. C: Ocena
60 ilościowa zależnej od liczby kopii transgenu ekspresji bfcrn, wykrywanej przez analizę Northern. Kolumny odpowiadają gęstości optycznej (średniej), natomiast słupki błędu odpowiadają standardowemu błędowi średniej. Istotność statystyczną oznacza się następująco: *, p<0,0; **, p<0,01. Fig.. Zależny od liczby kopii bfcrn poziom białka w płucu myszy wt i Tg. Western blot całkowitego białka komórkowego ( g/ścieżkę) poprzez zastosowanie oczyszczonej metodą powinowactwa króliczej surowicy odpornościowej (B4 - stabilnie transfekowany łańcuchem alfa bfcrn ekstrakt z linii komórek MAC-T (Kacskovics i wsp., 06a); WT - mysz typu dzikiego; TG2 - mysz hemizygotyczna, linia nr 14; TG - mysz hemizygotyczna, linia nr 19). Po lewej ukazano markery masy cząsteczkowej w kilodaltonach. W surowicy oczyszczonej metodą swoistego powinowactwa wobec łańcucha alfa bfcrn (Mayer i wsp., 02) wykryto około 40-kDa prążek w próbkach tkanki transgenicznego płuca, podobny do prążka obecnego w transfekowanym bfcrn (B4) wyciągu z komórek stanowiących kontrolę dodatnią. Drugi prążek (strzałka linią przerywaną) odpowiada prawdopodobnie mniej obficie występującej, nieglikozylowanej formie receptora. Porównanie ilości rekombinowanego łańcucha alfa FcRn w TG2 i TG, odpowiadających, odpowiednio, dwóm i pięciu kopiom transgenu, wykazuje, że ekspresja zależna od liczby kopii jest oczywista nie tylko na poziomie mrna, lecz również na poziomie białka. Fig. 6. Farmakokinetyka IgG mysiej (A) i ludzkiej (B) u myszy Tg bfcrn (linia nr 14, homozygota, zawiera 4 kopie bfcrn) i u myszy wt. Sporządzono wykres
61 modelowanych danych (symulacja na podstawie średniej geometrycznej pierwszorzędowych parametrów farmakokinetycznych), jak również obserwowanego średniego stężenia przeciwciała w surowicy ( g/ml) u trojga z pięciorga zwierząt (młode, dorosłe zwierzęta z tego samego miotu) w funkcji czasu (godzin po wstrzyknięciu) IgG mysiej ( mg/masy ciała) i ludzkiej ( i mg/masy ciała). Wstawki przedstawiają wartości czasu półtrwania wstrzykniętych IgG, obliczone z użyciem profesjonalnego oprogramowania WinNonLin w modelu dwukompartmentowym. Czas półtrwania IgG u zwierząt Tg był istotnie dłuższy w porównaniu z myszami wt. Próbki badano w dwóch powtórzeniach; słupki błędu odpowiadają standardowemu błędowi średniej (SEM). Statystyczną istotność oznaczono w następujący sposób: * oznacza p<0,0; ** oznacza p<0,01. Fig. 7. Nadmierna ekspresja bfcrn powoduje znaczne nasilenie odpowiedzi immunologicznej u myszy Tg, bez zakłócania jej natury. Myszy Tg i wt immunizowano i.p. OVA w CFA i 14 dni później eksponowano na OVA w 1FA. Seryjne próbki surowicy analizowano pod kątem IgM i IgG swoistych wobec OVA. W przypadku myszy Tg miana IgM były wyższe we wtórnej odpowiedzi immunologicznej w porównaniu z poziomem IgM w pierwotnej odpowiedzi immunologicznej, i istotnie wyższe w porównaniu z poziomem u myszy wt. Poziom istotności wskazuje na różnicę między myszami Tg a myszami wt (A). Miano IgG swoistej wobec OVA było niemal trzykrotnie wyższe u myszy Tg w porównaniu z mianem u zwierząt wt w czasie wtórnej odpowiedzi immunologicznej. Istotność poziomu wskazuje na różnicę między myszami Tg a myszami wt (B).
62 Po 32 dniach od immunizacji analizowano miana izotypów IgG swoistej wobec OVA. Myszy Tg wytwarzały istotnie wyższe miana izotypów IgG (z wyjątkiem IgG3), niemniej ich proporcje były takie same, jak u myszy wt, co wskazuje, że nadmierna ekspresja bfcrn zwiększa wytwarzanie IgG swoistych wobec OVA bez zakłócania go (C). Produkcja IgG całkowitej odzwierciedla miano IgG swoistej wobec OVA: myszy Tg wytwarzały istotnie większą ilość IgG niż ich rodzeństwo wt. Należy zauważyć, że wykazywały one wyższy poziom IgG nawet przed immunizacją (D). Ukazane wartości stanowią średnią SEM. * oznacza p<0,0; ** oznacza p<0,01; *** oznacza p<0,001; brak znamienności - p>0,0. Analizowano skuteczność wychwytu kompleksu immunologicznego FITC-OVA metodą FACS (E). Linia zaznaczona czarnymi kropkami wskazuje autofluorescencję komórek P388, linia szara wskazuje wychwyt OVA-FITC niepołączonego w kompleks immunologiczny, linia czarna wskazuje wychwyt kompleksu immunologicznego OVA-FITC wytworzonego z surowic wt, natomiast pole wypełnione na szaro oznacza wychwyt kompleksu immunologicznego OVA- FITC wytworzonego z surowic Tg. Autorzy nie stwierdzili wzmożonej fagocytozy przy stosowaniu 2, l surowicy w porównaniu z wychwytem OVA-FITC (2, l). Przy zastosowaniu l surowicy od myszy Tg obserwowano jednak istotne wzmocnienie, natomiast przy zastosowaniu takiej samej ilości ( l) od myszy wt nie stwierdzano nasilenia fagocytozy. I wreszcie, 40 l surowicy od myszy Tg i wt powodowało takie samo zwiększenie fagocytozy, co niepołączona w kompleks immunologiczny postać OVA-FITC (40 l).
63 Fig. 8. Myszy Tg i wt podobnie odpowiedziały na immunizację FITC-dekstranem. Myszy Tg (linia nr 14) i wt immunizowano dootrzewnowo FITC-dekstranem i poddawano je podobnej ekspozycji dwa tygodnie później. Autorzy obserwowali przede wszystkim IgM swoiste wobec FITC, nie stwierdzili natomiast różnic między zwierzętami Tg i wt. Analizując odpowiedź immunologiczną w czasie przebiegu immunizacji autorzy zauważyli zwiększenie wytwarzania IgM po drugiej immunizacji zarówno u myszy Tg, jak i wt. Fig. 9. Komórki śledziony myszy (po trzy samce w każdej grupie) immunizowanych OVA+CFA, eksponowanych w dniu 14 na OVA+IFA, zbierane w dniu 2. W śledzionie myszy Tg liczba komórek wytwarzających IgM swoiste wobec OVA uległa podwojeniu, a komórek wytwarzających IgG swoiste wobec OVA zwiększeniu ponadtrzykrotnemu w porównaniu z liczbą u zwierząt wt (A). 2 dni po immunizacji OVA doszło do zwiększenia masy śledziony (B) i liczby komórek (C) u myszy zarówno wt, jak i Tg, jednak wielkość śledziony i liczba komórek były istotnie większe u myszy Tg niż u myszy wt. Przedstawione wartości są to średnie SD (* oznacza p<0,0; *** oznacza p<0,01; p<0,001). Fig.. Analiza dystrybucji komórek śledziony u myszy zdrowych i u myszy immunizowanych OVA (2 dni po immunizacji), wt i Tg. Autorzy stwierdzili, że po immunizacji odsetek komórek z antygenami CD4R/B2 (limfocyty B) i I-A/I-E (klasa MHC II) uległ istotnemu zmniejszeniu (p<0,0). Nie stwierdzano różnic ani mniej radykalnego zmniejszenia w odniesieniu do komórek z markerem CD3 (limfocyt T), odpowiednio, u myszy wt i
64 6 1 2 Tg. Histogramy ukazują odsetek komórek (średnia SD) z antygenami CD4RB2 (limfocyty B), CD3 (limfocyty T) i I-A/I-E (klasa MHC II) (pole zakreskowane) lub swoiste izotypowo kontrole (linia czarna). Ukazano jedno typowe doświadczenie z dwóch przeprowadzonych (n = 3 myszy na grupę). Fig. 11. Immunizacja spowodowała masywny napływ neutrofili do śledziony. W celu dokładniejszego poznania charakterystyki komórek migrujących do śledziony 2 dni po immunizacji autorzy analizowali je metodą cytometrii przepływowej, stosując technikę podwójnego znakowania. Stwierdzono, że liczba komórek z antygenami CD11 b high i Gr-1 high uległa po immunizacji zwiększeniu około pięciokrotnemu u myszy wt i dziewięciokrotnemu u myszy Tg (A). Stwierdzono także, że odsetek komórek z antygenami CD11 b i antygenami klasy MHC II (makrofagi, komórki dendrytyczne) (B) i antygenami CD11b i CD11c (komórki dendrytyczne) (komórki C-bramkowane) był istotnie podwyższony u myszy Tg w porównaniu z ich kontrolami wt. Wykresy gęstości przedstawiają jedno typowe doświadczenie z dwóch przeprowadzonych (n = 3 myszy na grupę). Wartości przedstawione w wykresie słupkowym jest to średnia SD; * oznacza p<0,0; ** oznacza p<0,01. Fig. 12. Swoista wobec bfcrn ekspresja RT-PCR w neutrofilach i makrofagach pochodzących z komórek otrzewnej. Swoiste wobec bfcrn amplikony z powodzeniem powielono z próbek myszy Tg. Znaczniki: M1 drabinka DNA BenchTop (0 par zasad) - (Promega); I nieoczyszczone komórki otrzewnej myszy wt; 2 - nieoczyszczone komórki otrzewnej myszy Tg; 3 -
65 66 1 oczyszczone neutrofile (CD11b high, Gr-1 high ), makrofagi i komórki dendrytyczne (CD11b high, Gr-1 low ) myszy Tg, również analizowane metodą cytometrii przepływowej, 4 - powielanie z plazmidu zawierającego cdna łańcucha alfa bfcrn. Grot strzałki wskazuje na prążek DNA o długości 48 par zasad, swoisty wobec bfcrn. Wykres gęstości przedstawia jedno typowe doświadczenie z dwóch przeprowadzonych. Fig. 13. Obecność komórek zawierających OVA-FITC w śledzionie. Immunizowanym OVA (6 dni po immunizacji) i nieimmunizowanym myszom transgenicznym (homozygota nr 14) podano dootrzewnowo UVA-FITC. Pięć godzin po leczeniu analizowano komórki śledziony metodą cytometrii przepływowej i stwierdzono, że w śledzionie myszy immunizowanych OVA znajdowało się 1,1 1,4% komórek OVA-FITC-dodatnich, natomiast u zwierząt nieimmunizowanych nie wykryto istotnych ilości komórek OVA-FITC-dodatnich (2,1 0,3%). Obrazy konfokalne przedstawiają komórki śledziony myszy immunizowanych OVA i myszy nieimmunizowanych. Jądra komórek zabarwiono przenikalną dla komórek sondą DNA fluoryzującą w czerwieni - DRAQ, natomiast FITC-OVA uwidaczniają się jako jasne plamki (A). U zwierząt immunizowanych OVA wśród komórek FITC-dodatnich, 61,2,4% stanowiły 2 komórki B2-dodatnie (limfocyty B), 18, 0,6% - komórki CD11b high i Gr-1 high (neutrofile), a 13, 2,1% - komórki CD11b- i CD11c-dodatnie (dendrytyczne) (B). OVA-FITC ulega internalizacji do typowego neutrofila z jądrem wielopłatowym, natomiast OVA-FITC jest wykrywany na powierzchni komórki, która posiada duże jądro o okrągłym kształcie i cienki rąbek cytoplazmy; jest to
66 przypuszczalnie limfocyt B (C). Wykresy gęstości i histogramy przedstawiają jedno typowe doświadczenie (n = 3 myszy na grupę); dane wskazują odsetek komórek (średnia SD). Paski skali oznaczają m. Fig. 14. Analiza RT-PCR transgenicznych myszyzałożycieli HIV-P2-FcRn. RT-PCR z primerami planowanymi dla promotora P2, oczekiwana wielkość fragmentu: 79 par zasad, B RT-PCR z primerem planowanym dla bydlęcego egzonu 4 FcRn, oczekiwana wielkość fragmentu: 161 par zasad. Próbki: 1. płuco myszy HIV-P2-FcRn; 2 wątroba myszy HIV-P2-FcRn; 3 jelito myszy HIV-P2-FcRn; 4. śledziona myszy vhiv-p2-fcrn;. DNA genomowe myszy HIV-P2-FcRn; 6. bydlęce DNA genomowe; 7. DNA genomowe myszy kontrolnych; 8. cdna wątroby bydlęcej; 9. cdna myszy kontrolnych,. Kontrola ujemna. Fig. 1. Nadmierna ekspresja bfcrn poważnie wpływa na metabolizm albuminy. Poziom albuminy w surowicy myszy pozbawionych FcRn (FcRn KO), wt, homozygotycznych nr 14 i nr 19 (wykazujących ekspresję, odpowiednio, 4 i kopii bfcrn). Przy porównywaniu poziomu albuminy u myszy KO i WT myszy (p<0,01), myszy WT i TG4 lub TG myszy (p<0,001) obserwowano istotne różnice. Fig. 16. Eliminacja IgG króliczej u myszy FcRn -/- (KO) i u myszy FcRn -/- transgenicznych pod względem bydlęcego FcRn (KO_bFcRn). A. Multipleksowa PCR celem zidentyfikowania myszy F2 z genotypem bfcrn /mfcrn -/- (oznaczony kwadratem). Myszy F2 uzyskano poprzez skrzyżowanie myszy F1, urodzonych po skrzyżowaniu szczepów rodzicielskich bfcrn/mfcrn X mfcrn -/- neo+. Primery opisano w przykładzie 12. Oczekiwana wielkość fragmentów: bfcrn: 6 bp; neo: 34 par zasad; mfcrn
67 par zasad. Przedziały: 1. bfcrn+, neo+, mfcrn+; 2. bfcrn+, neo+, mfcrn - ; 3. bfcrn+, neo+, mfcrn+; 4. bfcrn -, neo+, mfcrn - ;. bfcrn -, neo+, mfcrn - ; 6. bfcrn+, neo+, mfcrn+; 7. bfcrn+, neo -, mfcrn+; 8. bfcrn+ neo+, mfcrn+; 9. bfcrn -, neo+, mfcrn+;. bfcrn+, neo+, mfcrn+; 11. bfcrn 4, neo+, mfcrn+; 12. bfcrn+, neo -, mfcrn+; 13. Bydlęce DNA genomowe; 14. bfcrn +, neo -, mfcrn + ; 1. Kontrola ujemna B. Farmakokinetyka króliczej IgG u myszy FcRn -/- (KO) i u myszy FcRn -/- transgenicznych pod względem bydlęcego FcRn (KO_bFcRn). Królicza IgG była szybko usuwana u myszy FcRn -/- (czas półtrwania: 1 godzin), natomiast u myszy bfcrn/fcrn -/- była chroniona (czas półtrwania: 67 godzin). Zwierzętom wstrzykiwano i.v. g króliczej IgG. Leczeniu poddawano trzy myszy/grupę; dane odpowiadają dwóm niezależnym doświadczeniom. Dane przedstawiają wartości średnie; słupki błędu odpowiadają standardowemu błędowi średniej (SEM). Fig. 17. Analiza PCR królika transgenicznego bfcrn. PCR, z primerami opracowanymi do wykrywania bydlęcego 4. egzonu FCGRT. Oczekiwana wielkość wynosi 160 par zasad. Próbki: 1. DNA genomowe królika 38/JT; 2. Bydlęce DNA genomowe; 3. DNA genomowe myszy FVB/n; 4. Próba ślepa, kontrolna; MM drabinka o wielkości 1kb. Fig. 18. Określanie charakterystyki BAC królika 262E02. Primery i warunki PCR, stosowane do oceny obecności genów królika, opisano w tabeli 7 w przykładzie 1. PRZYKŁADY Przykład 1 - Izolowanie i określanie charakterystyki klonów BAC, zawierających gen łańcucha alfa hfcra (bfcgrt) (przykład przygotowawczy)
68 69 Izolowanie klonów BAC, zawierających hfcgrt 1 2 [0113] BAC 90 wyizolowano z bydlęcej biblioteki BAC, uzyskanej z DNA z limfocytów dorosłego samca (2 lata) Bos taurus Jersey (uzyskanych z bazy danych Resource Center/Primary Database, German Human Genome Project (RZDP), Instytut Maxa Plancka Genetyki Molekularnej, Berlin, Niemcy: Zidentyfikowano bfcgrt-dodatni klon BAC poprzez skrining PCR z użyciem primerów swoistych dla mrna łańcucha alfa bfcrn [6 42 hp. GenBank AF1396) powielono PCR BFclS: - CAGTACCACTTCACCGCCGTGT-3 (SEQ ID nr 1); BFclas: - CTT-GGAGCGCTTCGAGGAAGAG-3 (SEQ ID nr 2)]. W kolejnym etapie DNA bfcgrt sekwencjonowano z użyciem primerów, zapewniających annealing z egzonami genu (Kacskovics i wsp. 00). W celu analizy bfcgrt w górę, region flankujący DNA BAC strawiono BamHI, i strawiony DNA rozdzielono na żelu agarozowym. Analiza Southern blot przy zastosowaniu jako sondy fragmentu DNA z domeny ai pozwoliła wykryć prążek dodatni o długości 9 kb. Fragment BamHI o długości 9 kb subklonowano do wektora pgem-11zf(+). W wyniku dodatkowego procesu subklonowania uzyskano 2 kb lub segmenty promotora z egzonami od 1 do 3 w tym samym wektorze. Następnie przeprowadzono pełne sekwencjonowanie wstawki za pomocą zestawu Ready Reaction Kit (AB1. 373A-Stretch, Perkin Elmer) z Cybergene Company (Huddinge, Szwecja) w cyklu AB1 Prism BigDye Terminator Cycle. [0114] BAC 189HO2 i 128EO4 izolowano z użyciem swoistych wobec bfcgrt primerów: FcRnF: - CGGCCACCTCTAT-CACATTT-3 (SEQ ID nr 3) i FcRnR: - TGCATTGACCACACTTGGTT-3 (SEQ ID nr 4) (GenBank NW_
69 ) z bydlęcej biblioteki BAC (Eggen i wsp. 01). Wielkość wstawki izolowanych klonów BAC analizowano poprzez trawienie klonów endonukleazą restrykcyjną Notl. Do określenia wielkości regionów granicznych i 3 hfcgrt we wstawkach zastosowano system Expand Long Template PCR System (Roche). Opracowano dwa zestawy primerów: primery pbac-górny ( - ACCTCTTTCTCCGCACCCGACATAG, SEQ ID nr. U ) i bfcrn-antysensowny (GTTCAAGTCCAAAGGCAGGCTATCT, SEQ ID nr 6) powielały region overhang. Z drugiej strony jako primery antysensowne stosowano bfcrnsensowny (CCTTTACCCACACCCACTCCCCACA, SEQ ID nr 7) i pbac-niższy (AGAAGTTCGT-GCCGCCGCCGTAGTA, SEQ ID nr 8, U ) do powielenia regionu overhang 3 bfcgrt. Określanie charakterystyki bfcgrt [011] Około 1800 par zasad regionu w górę od początku transkrypcji, jak również cały gen bfcgrt, sekwencjonowano i porównano ze złożonymi w depozycie NCBI bydlęcymi sekwencjami genomowymi poprzez zastosowanie programu BLAST. Sekwencjonowany fragment poddano skriningowi pod kątem rozproszonych powtórzeń i sekwencji DNA o małej złożoności, stosując program RepeatMasker (Smit i wsp ) (http: repeatmasker.org). Baza danych rozproszonych powtórzeń, sprawdzana przez program RepeatMasker, opiera się na bazie danych powtórzeń (Repbase Uptake; (Jurka i wsp. 0). Dane wykazywały dużą zgodność ze złożoną w depozycie NCBI krowią sekwencją; pierwsze 3477 par zasad (do niesekwencjonowanego regionu 2, kb) wykazuje 99% tożsamość z klonem NW_92938S (18 contigów
70 genomowych chromosomu Bos taurus), jakkolwiek wykazuje przerwę o długości 28 par zasad między 02 a 2287 parą zasad, w pierwszym intronie. Druga część analizowanej sekwencji zawierała 1 78<t par zasad (po niesekwencjonowanym segmencie o długości 2, par zasad), wykazujących 99% tożsamość z tym samym contigiem genomowym. Ten fragment zawiera także przerwę o długości 167 par zasad (między 13 a 1370 parą zasad), należącą do intronu 6. Dwa niezidentyfikowane segmenty zawierają powtarzające się sekwencje (powtórzenia proste), co wyjaśnia brak homologii. Na podstawie tego dopasowania długość brakującego odcinka obliczono na 26 par zasad. Stwierdzono zatem, że sekwencja wykazuje dużą homologię z contigiem genomu bydła NW_92938, między a parą zasad. [0116] Analizowano także miejsce inicjacji transkrypcji poprzez dopasowanie dostępnych sekwencji cdna łańcucha alfa bfcrn (sekwencje referencyjne, jak również sekwencje z bazy danych EST; NCBI) z sekwencją genomową bfcgrt. Porównywano organizację genomową bydlęcych, ludzkich, szczurzych i mysich genów FCGRT za pomocą NCBI Map Viewer (dane nieukazane). Określanie charakterystyki bfccrt-dodatnich klonów BAC [0117] Izolowano trzy różne klony BAC - 90, 189H02 i 128E04 zawierające gen łańcucha alfa bydlęcego noworodkowego receptora Fc (bfcgrt) i jego własne środowisko genomowe. Po trawieniu NotI, wstawki genomowe z wektorów BAC analizowano metodą elektroforezy żelowej w polu pulsacyjnym, która ujawniła, że klon 189H02, klon 128E04 i klon 90
71 zawierają wstawki genomu bydlęcego o wielkości, odpowiednio, około 1 kb, 0 kb i 90 kb. [0118] Następnie metodą PCR dużego zasięgu określono wielkość regionów granicznych genu bfcgrt. Dane wykazują, że BAC 90c i 189HO2 zawierają regiony flankujące i 3 o wielkości, odpowiednio, 8, kb i 14 kb, które, jak można sądzić na podstawie ich wielkości, mogą nie zawierać wszystkich elementów regulatorowych, zapewniających niezależną od miejsca integracji, swoistą tkankowo ekspresję genu bfcgrt. Powielanie metodą PCR klonu 128HO2 dwoma zestawami primerów nie ujawniło żadnych produktów. Jako że system Expand Long Template PCR System jest sprawną metodą powielania, generującą do 2 kb amplikonów z faga DNA, autorzy wywnioskowali, że regiony graniczne i 3 bfcgrt w tym klonie mają długość powyżej 2 kb, do mikroiniekcji wybrali zatem BAC 128E04. [0119] Dane niedawno opublikowane na stronie internetowej Bovine Genome Resource ( guide/cow/) i sekwencje końca i 3 BAC 128E04 umożliwiły autorom określenie kontekstu genomowego i dokładnej wielkości regionów granicznych bfcgrt: region regulatorowy rozciąga się na długość do 44 kb, natomiast region 3 - do 0 kb. Dane ujawniły, że BAC 128E04 zawierał pięć domniemanych genów kodujących białka (FLT3LG. LOC LOC273, LOCS11234, LOC1123) i bfcgrt (Fig. 1A). Przykład 2 Wytwarzanie i genotypowanie transgenicznych myszy, posiadających bydlęcy BAC 128E04 (przykład porównawczy)
72 Wytwarzanie wstawki genomowej o wielkości 2 kb z klonu BAC 128EO4 do mikroiniekcji i uzyskiwanie transgenicznych myszy, posiadających bydlęcy BAC 128E04 [01] Wytwarzanie DNA BAC (klon 128E04) do mikroiniekcji przeprowadzono z zastosowaniem niewielkich modyfikacji lub zgodnie z protokołem opublikowanym przez Schedl i wsp. (Schedl i wsp., 1996). Oczyszczony BAC (oczyszczanie plazmidów Qiagen dla plazmidów o bardzo małej liczbie kopii) poddano trawieniu Not 1 (enzymy) celem uwolnienia wstawki, którą izolowano w preparatywnym 1% żelu agarozowym w polu pulsacyjnym. Warstwę żelu, zawierającą wstawkę, umieszczono na żelu LMP (ang. low melting point o niskiej temperaturze topnienia), który trawiono z użyciem Gelase (Epicentre). Do oczyszczenia wstawki z agarozy zastosowano kolumnę Microcon YM0 (Millipore). Wstawkę eluowano w buforze odpowiednim do mikroiniekcji ( mm Tris-HCl, ph 7,, 0,1 mm EDTA, ph 8,0, 0 mm NaCl z dodatkiem 0,03 mm spwmine/0,07mm spermidyny - (SIGMA) albo bez tego dodatku. Stężenie DNA doprowadzono do wartości 0,4 ng/ l, stosując bufor do mikroiniekcji (mm Tris-HCl, ph 7,, 0,1 mm EDTA, 0 mm NaCl) i wstrzyknięto do zapłodnionych oocytów myszy FVB/N. Biorcami były samice CD1 w wieku tygodni. Zwierzęta doświadczalne nabyto w firmie Charles Rivers Laboratories Hungary Ltd. (Budapeszt). Określanie charakterystyki transgenicznych myszy [0121] W celu wykrywania obecności bfcgrt u myszy, izolowano DNA genomowe z bioptatów ogona miotów urodzonych po transferze zarodków oraz potomstwa G1 i G2 założycieli i przeprowadzano ich badania przesiewowe
73 74 1 pod postacią dwóch amplifikacji PCR. Dwie pary primerów opracowano na podstawie sekwencji bfcgrt klonu BAC 90a. Pierwsza para primerów była złożona z bfcsuf: - CTCCTTTGTCTTGGGCACTT-3 (SEQ ID nr 9) jako primeru sensownego i BFcL -GCCGCGGATC-CCTTCCCTCTG -3 (SEQ ID nr ) primeru antysensownego, w wyniku czego uzyskano produkt o wielkości 600 par zasad (pary zasad ), natomiast drugą parą primerów była para Fcrnfpr/in -AAAGTTTCTCGAGAGAGGCAGAGAC-3 (SEQ ID nr 11) jako primer sensowny i Fcmrpr/in - TAGTTACAGAGCCTGGATAGGCTGA-3 (SEQ ID nr 12) jako primer antysensowny, w wyniku czego uzyskano produkt o wielkości 4 par zasad (pary zasad ). Wyniki doświadczeń z transgenezą przedstawiono w tabeli 1. Tabela 1. Uzyskiwanie transgenicznych myszy, posiadających bydlęcy BAC 128E04 Wstrzyknięty konstrukt 128E04 Liczba zarodków Podano w mikroiniekcji 0 Podano na drodze transferu 360 Liczba zwierząt Urodzonych 41 Transgenicznych 6 2 Skuteczność Liczba zwierząt 14,6 mikroiniekcji urodzonych/liczba zwierząt Tg [0122] Jak ukazano w tabeli 1, urodziło się w sumie 41 młodych; genotypowano je poprzez analizę DNA z ogona na obecność bfcgrt. Od sześciu zwierząt-założycieli uzyskano trzy niezależne linie Tg. Dwie z tych trzech linii - nr 14, nr 19 wykazywały mendlowski wzór dziedziczenia transgenu w pierwszym pokoleniu (17 i 12 z całkowitej liczby i 34 miotów wykazywały obecność transgenu), natomiast trzecia linia, nr 9, wykazywała pewien stopień mozaikowatości u zwierzęcia-założyciela.
74 7 1 Myszy Tg były nie do odróżnienia od ich rodzeństwa z tego samego miotu: nie różniły się masą ciała ani ogólnym stanem zdrowia. Analiza integralności transgenu o dużym zasięgu [0123] Oceniano integralność transgenu w trzech liniach Tg poprzez swoiste pary primerów opracowanych dla końców i 3 BAC 128E04 i dla pięciu domniemanych genów kodujących białka, zlokalizowanych na wstrzykiwanym 13AC, na podstawie mapy genomu bydlęcego (GenBank MapViewer Build 3.1 (na podstawie Btau 3.1) - bydlęcy chromosom 18, region między parami zasad ). Sekwencje primerów i warunki opisano w tabeli 2. Tabela 2. Primery i warunki PCR do oceny braku odchyleń w strukturze zintegrowanego BAC 128E04 Domniemany gen BAC128EOH koniec FLT3LG LOC39196 Primer -TTT AGC TGC ATC GGG ATC TT-3 (SEQ ID nr 13) -GGA GTG ATG GCA TTT GGT TT-3 (SEQ ID nr 14) -TCG GAG ATG GAG AAA CTG CT-3 (SEQ ID nr 1) -CTG GAC GAA GCG AAG ACA G-3 (SEQ ID nr 16) -AGA ACG TGC GTA CCA AAA GC-3 (SEQ ID nr 17) -AGC GGT TGT ACT TTC GGA TG-3 (SEQ ID nr 18) Temp. topnienia ( C) Produkt (par zasad)
75 76 bfcgrt -CCA AGT TTG CCC TGA ACG-3 (SEQ ID nr 19) -GTG TGG GCA GGA GTA GAG GA-3 (SEQ ID nr ) LOC273 -AGT GGT CCT GGG ATT GAC AG-3 (SEQ ID nr 21) -TCA CTG AGT CCC GTA TGT GC-3 (SEQ ID nr 22) LOC CTA CGT GTG CGC CGT 61 2 GAC-3 (SEQ ID nr 23) -AAT CAG CTT CTC CAC GCA CT-3 (SEQ ID nr 24) LOC1123 -GTT GTT CAC ACC AGG GAA CC-3 (SEQ ID nr 2) -CCT TTG CCA TTG TAG ATG TAG C-3 (SEQ ID nr 26) BAC128EOH -AGT CGT GTC CGA CTC koniec 3 TTT GC-3 (SEQ ID nr 27) -CAG CCT GTC TGG TGT TCT GA-3 (SEQ ID nr 28) [0124] Sekwencje BAC128EOH i 3 BAC128EOH uzyskano od A. Eggen (Inra, Jouy-en-Josas, Francja). Wszystkie pary primerów dały takie same produkty PCR, jak BAC 128E04 i bydlęce DNA genomowe, co wskazuje na integrację nienaruszonego BAC, z wyjątkiem DNA linii nr 9, w której PCR LOC 11234, LOC 2223 i koniec 3 BAC128E04 nie dał w wyniku produktów PCR (Fig. 1B). Można zatem wyciągnąć wniosek, że w tej linii Tg brak fragmentu o szacunkowej długości kb od końca 3 zintegrowanego transgenu BAC. Utrata fragmentów genomu z obu końców ( i 3 ) dużych transgenów jest częstym zjawiskiem (Raguz i wsp., 1998). Niemniej, żeby uniknąć
76 możliwości zaburzenia ekspresji bfcgrt z powodu nieobecności elementów regulatorowych o nieznanych właściwościach, które mogłyby znajdować się w brakującej części DNA BAC, linii nr 9 nie włączono do dalszych badań. Lokalizacja chromosomalna transgenu [012] W celu wykluczenia możliwości, że bydlęcy klon BAC 128E04 uległ przypadkowo integracji w identycznym segmencie mysiego chromosomu w obu liniach transgenicznych i przez to fenotyp linii transgenicznych myszy nr 14 i nr 19 był skutkiem insercyjnej mutagenezy niezidentyfikowanego genu lub genów w miejscach integracji transgenu, przeprowadzono fluorescencyjną hybrydyzację in situ (FISH) w celu wizualizacji integracji genomowej transgenu 128E04. DNA BAC 128E04 wyznakowano przez nick-translację biotyno- 14-dATP (zestaw do znakowania BioNick, Invitrogen, USA). Chromosomy mitotyczne uzyskano z fibroblastów traktowanych winblastyną, izolowanych z zarodków homozygot, odpowiednio, nr 14 i nr 19, w wieku 13, dnia, zgodnie ze standardowymi protokołami, obejmującymi obróbkę hipotoniczną i utrwalenie w metanolu:kwasie octowym (3:1). [0126] FISH przeprowadzono w zasadzie tak, jak opisywano uprzednio (Hayes i wsp., 1992). Biotynylowaną sondę poddano denaturacji i pozostawiono do hybrydyzacji do zdenaturowanych, rozchodzących się chromosomów przez noc w temperaturze 37 C. Miejsca hybrydyzacji na chromosomach powielono z użyciem przeciwciała przeciwko biotynie, uzyskanego u kozy (Vector Laboratories Inc, Burlingame, CA, USA), i
77 wizualizowano przez dalszą inkubację ze sprzężoną z fluoresceiną króliczą antykozią IgG (Nordic Immunologic Laboratories, Tilburg, Holandia). Preparaty chromosomów poddano przeciwbarwieniu diamidyno-2-fenylindolem (DAPI; Vector Laboratories, Burlingame, CA, USA) i obserwowano za pomocą mikroskopu epifluorescencyjnego Nikon Eclipse E600 (Nikon Instruments; Kawasaki, Japonia). Obrazy fluorescencyjne rejestrowano za pomocą aparatu Cohu 49 CCD (Cohu, Inc.; San Diego, CA, USA) i przetwarzano na obraz cyfrowy z użyciem oprogramowania MacProbe 4.3 FISH (Applied Imaging; Newcastle upon Tyne, Wielka Brytania) na komputerze Apple Macintosh G4. [0127] Wynik analizy FISH ujawnił, że fluorescencyjnie wyznakowany BAC 128E04 hybrydyzował u szczepów myszy nr 14 i nr 19 do zupełnie różnych segmentów chromosomów. Wyklucza to możliwość, żeby ich fenotypy były zależne od miejsca integracji. Pojedyncze plamki w chromosomach wskazują, że wielokrotne kopie (2 w linii nr 14 i w linii nr 19) integracji transgenu najprawdopodobniej występowały w postaci powtórzeń tandemowych (Fig. 2). Przykład 3 - Efekt liczby kopii bydlęcego genu FCGRT (bfcgrt) na ekspresję FcRn (przykład stanu wyjściowego) Określanie liczby kopii transgenu z użyciem ilościowej polimerazowej reakcji łańcuchowej w czasie rzeczywistym [0128] PCR w czasie rzeczywistym jest ilościową, dokładną metodą określania liczby kopii i zygotyczności transgenów u zwierząt Tg (Tesson i wsp., 02). Liczbę kopii transgenu BAC 128E04 ustalono przez bezwzględne określenie liczby bfcgrt i genów -aktyny myszy (wzorzec wewnętrzny) w następujący sposób.
78 79 1 [0129] Liczbę kopii transgenu BAC 128E04 określono metodą TaqMan, stosując system ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster City, CA, USA). Sekwencje oligonukleotydowe stanowiące primer i sondę opracowano z użyciem programu Primer Express v2.0 (Applied Biosystems), stosując parametry domyślne (primery i sondy przedstawiono w tabeli 3). Do ekstrakcji DNA z próbek z ogona zwierząt hemizygotycznych zastosowano konwencjonalną metodę fenolowo-chloroformową z dodatkowym etapem ekstrakcji chloroformem. Tabela 3. Primery i sondy do określania liczby kopii transgenu bfcgrt Mysia - Sonda VIC-TGGCTTTCTGAACTTGACAACATTAT-TAMRA (SEQ aktyna ID nr 29) Forward TTCACCTGCCCTGAGTGTTTC (SEQ ID nr ) Reverse TGAAGGTCTCAAACATGATCTGTAGA (SEQ ID nr 31) Bydlęce Sonda FAM-CACAGTCAAGAGTGGCGACGAGCAC-TAMRA (SEQ FCGRT ID nr 32) Forward GCACCACGCAGCGGTAGT (SEQ ID nr 33) Reverse CCTTCTACGCCTGGTCATCAC (SEQ ID nr 34) [01] Geny mysiej -aktyny i geny bfcgrt oznaczano ilościowo w poszczególnych próbkach poprzez bezwzględne oznaczenie ilościowe z użyciem krzywych kalibracji. W wyniku zastosowania krzywych standardowych, z wykorzystaniem pięciu punktów, z rozcieńczeniem w zakresie do 32 razy, uzyskano dużą liniowość przy użyciu zestawów primerów. Przed oznaczeniem ilościowym walidowano liniowość i skuteczność ilościowego oznaczenia metodą PCR. Próbki oceniano w dwóch powtórzeniach (Fig. 3).
79 80 [0131] Do określania stężenia DNA jako wzorzec 1 wewnętrzny stosowano endogenny gen -aktyny, obecny w dwóch kopiach w każdej komórce. Do wykreślenia krzywej kalibracji dla genu -aktyny stosowano mysie DNA genomowe. Bezwzględne oznaczenie ilościowe genu bfcgrt przeprowadzano na podstawie standardowej krzywej, uzyskanej z seryjnych rozcieńczeń BAC 128E04 z dodatkiem mysiego DNA genomowego. Krzywe standardowe umożliwiły autorom określanie liczby kopii genu bfcgrt na podstawie następujących obliczeń: dokładna ilość DNA określała liczbę diploidalnych genomów w próbkach, natomiast krzywa kalibracji genu bfcgrt określała liczbę jego kopii w próbkach DNA hemizygotycznych zwierząt z linii nr 14 i nr 19. Po przeprowadzeniu tych obliczeń ustalono, że liczba kopii genu bfcgrt wynosiła 2 i u hemizygotycznych zwierząt, odpowiednio, z linii nr 14 i nr 19 myszy Tg (Tabela 4). Tabela 4. Wartości bezwzględnej liczby komórek i bezwzględnej liczby kopii FcRn w próbkach uzyskano, odpowiednio, z krzywych kalibracji mysiej -aktyny i bfcrn (Fig. 3). Szacunkowa liczba kopii transgenu bfcrn u zwierząt hemizygotycznych wynosiła 2 w linii nr 14 i w linii nr 19. Linia nr Gen Ct Bezwzględna liczba genów w próbce (AGN) Bezwzględna liczba komórek w próbce (ACN) Liczba kopii bfcrn (ACNb- FcRn/ACN) Szacunkowa liczba kopii transgenu bfcrn u diploidalnego zwierzęcia hemizygotycznego 14 Mysia 31, , ,248 - aktyna bfcrn 28, , Mysia, ,722
80 aktyna bfcrn 27,0 468,18,13 Oznaczenie liczby kopii na poziomie transkrypcji przez odwrotną transkryptazę-pcr i analizę Northern [0132] Ekstrahowano RNA całkowite przez zastosowanie RNAzol B (TEL-TEST INC) z wątroby, płuca i sutka samic w wieku 6 tygodni oraz z jelita noworodków. Dwa mikrogramy RNA poddano odwrotnej transkrypcji, stosując enzym odwrotnej transkryptazy wirusa mysiej białaczki Moloneya (M-MLV) i primer adapterowy (dt)17, zgodnie z zaleceniem producenta (Acces RT-PCR System; Promega). Przeprowadzono PCR do uzyskania swoistego wobec bfcgrt amplikonu o długości 367 par zasad (pary zasad , Kacskovics i wsp., 00) z użyciem par primerów: B7 -. GGCGACGAGCACCACTAC-3 (SEQ ID nr 3) i B8 - GATTCCCGGAGGTCWCACA-3 (SEQ ID nr 36) (gdzie W może stanowić A lub T). Powielony segment rozdzielano przez elektroforezę na 1% żelu agarozowym i barwiono bromkiem etydowym. [0133] Ponieważ w wielu komórkach nabłonkowych wykryto transkrypty FcRn przeżuwaczy (Kacskovics i wsp., 06b; Mayer i wsp., 04; Mayer i wsp., 02), podobnie jak w komórkach śródbłonka naczyniowego (Kacskovics i wsp., 06a), ekspresję łańcucha alfa bfcrn analizowano w płucu, wątrobie i sutku dorosłych samic Tg w fazie laktacji oraz w jelitach noworodków, stosując RT-PCR. mrna bydlęcego łańcucha alfa FcRn ulegało ekspresji we wszystkich wybranych tkankach hemizygotycznych zwierząt z linii nr 9, nr 14 i nr 19 (Fig. 4A). [0134] W celu oceny zależności ekspresji transgenu od liczby kopii i porównania liczby kopii z ilością mrna endogennego bydlęcego łańcucha alfa FcRn, próbki RNA
81 wątroby od bydła i od osobników z linii nr 14 i nr 19 analizowano metodą Northern blot. RNA całkowite izolowano z wątroby młodych, dorosłych samic myszy i g RNA całkowitego poddano frakcjonowaniu ze względu na wielkość na 1% żelu agarozowym/2,2 M formaldehydzie, przeniesiono na błonę Hybond N+ (Amersham) i hybrydyzowano z wyznakowaną 32 P sondą cdna, syntetyzowaną metodą PCR z użyciem opisanych powyżej primerów B7-B8. Jako wzorzec wewnętrzny zastosowano sygnał RNA 18S do oszacowania ładunku RNA na żelach. Uzyskane sygnały oceniano za pomocą PhosphorImager i oznaczano ilościowo za pomocą STORM (Molecular Dynamics). Porównania między gęstością sygnału swoistego mrna bfcrn między myszami Tg z dwiema, czterema, pięcioma i dziesięcioma kopiami dokonano, stosując test t-studenta. [013] Wyniki wykazują wyższy poziom ekspresji mrna transgenu w linii nr 19. Poziom ekspresji mrna w wątrobie hemizygotycznych myszy Tg z linii nr 14, posiadających dwie kopie transgenu BAC, osiągał 90% poziomu obserwowanego w wątrobie bydlęcej (Fig. 4B). [0136] Analiza ilościowa i ocena statystyczna intensywności sygnału od dwojga lub trojga zwierząt hemi- i homozygotycznych z obu linii Tg wykazała, że ilość mrna bydlęcego łańcucha alfa FcRn w wątrobie myszy Tg ściśle koreluje z ich liczbą kopii transgenu i różnice były istotne na poziomie prawdopodobieństwa p<0,0 (Fig. 4C). Ten wynik, wraz z tym, że poziom ekspresji mrna łańcucha alfa FcRn w wątrobie myszy Tg z dwiema kopiami transgenu był podobny do poziomu mrna w wątrobie bydlęcej, wskazuje, że BAC 128E04 zawiera
82 wszystkie konieczne elementy regulatorowe, zapewniające zależną od liczby kopii, niezależną od pozycji ekspresję bfcgrt. Wykrywanie białka łańcuch alfa bfvrn w płucu myszy transgenicznych [0137] Badano ekspresję bydlęcego łańcucha alfa FcRn na poziomie białka poprzez analizę Western. Ekstrakty białka rozwinięto na poliakrylamidowym, denaturującym żelu Tris-glicyna: plamki sondowano z użyciem oczyszczonej metodą powinowactwa króliczej surowicy odpornościowej (uzyskanej przeciwko peptydowi CLEWKEPPSMRLKAR, odpowiadającemu wysoce zakonserwowanym resztom aminokwasowym łańcucha alfa bfcrn plus N-końcowa Cys do sprzęgania z KLH (Mayer i wsp., 02)). Związane przeciwciało przeciwko łańcuchowi alfa bfcrn wykrywano z użyciem sprzężonego z peroksydazą chrzanową koziego przeciwciała antykróliczego i na podstawie wzmożonej chemiluminescencji, stosując jako substrat roztwór na bazie luminolu. Jako kontroli dodatniej użyto bydlęcej linii komórek nabłonkowych (B4) stabilnie transfekowanej łańcuchem alfa bfcrn (Kacskovics i wsp., 06a). [0138] W próbkach hemizygotycznego płuca z obu linii Tg o masie odpowiadającej znanej masie cząsteczkowej łańcucha alfa bfcrn (Kacskovics i wsp., 00) - wykrywano białko o długości 40 kd, które nie występowało u myszy typu dzikiego, użytych jako kontrola ujemna (Fig. ). Masa cząsteczkowa transgenicznego łańcucha alfa FcRn odpowiadała rekombinowanemu białku, wytwarzanemu przez bydlęcą linię komórek nabłonkowych B4, które stabilnie
83 transfekowano bfcrn (Kacskovics i wsp., 06a). Dane te ponadto potwierdziły wyniki analizy Northern blot, wskazując, że próbka od myszy z linii nr 19 z ekspresją kopii transgenu bfcrn zawiera znacznie więcej białka bfcrn, niż wykrywano u myszy z linii nr 14 z ekspresją 2 kopii transgenu (Fig. ). [0139] Dane te dowodzą, że klon BAC o długości 2 kb zawiera wszystkie konieczne genetyczne elementy regulatorowe, niezbędne do powtarzalnej, tkankowo swoistej ekspresji na fizjologicznym poziomie łańcucha alfa bfcrn (Fig. 4A), co podkreśla tezę, że zachowanie bydlęcych transgenów u myszy powinno się w miarę możliwości porównywać z wzorcami ekspresji tego genu w tkankach bydlęcych. Przykład 4 Badania in vivo z analizą czasu półtrwania mysiej i ludzkiej IgG u myszy transgenicznych pod względem bydlęcego BAC (przykład stanu wyjściowego) [0140] W celu analizy ekspresji łańcucha alfa bfcrn u myszy Tg, jak również zbadania, czy bydlęcy łańcuch alfa FcRn i mysi 2m są w stanie tworzyć funkcjonalny receptor, analizowano eliminację mysiej i ludzkiej IgG. Tę ostatnią poddano badaniom, ponieważ niedawno wykazano, że bydlęcy FcRn wiąże ludzką IgG znacznie lepiej, niż IgG bydlęcą (Kacskovics i wsp., 06a). [0141] Po wstępnym pobraniu krwi, dobranym pod względem wieku, masy ciała i płci (samce) homozygotom nr 14 i myszom kontrolnym (3- na grupę) podano w formie mikroiniekcji dożylnie mg/kg masy ciała mysiej IgG1 przeciwko OVA (mab Sigma), albo mg/masy ciała lub mg/masy ciała ludzkiej IgG (Gammonativ przeznaczony do stosowania dożylnego autorzy otrzymali w darze od firmy
84 8 1 2 Octapharma, Sztokholm, Szwecja) w 0mg/ml roztworze soli fizjologicznej; w czasie następnych 216 godzin okresowo pobierano próbki krwi (0 l/pobranie) ze splotu zaoczodołowego. Do oceny stężenia w osoczu mysiej IgG1 anty-ova w trakcie doświadczenia zastosowano ilościową ELISA, wykorzystując OVA (Sigma) jako odczynnik wychwytujący i sprzężoną z HRP, oczyszczoną metodą powinowactwa poliklonalną, kozią antymysią IgG (swoistą wobec łańcucha ) (Southern Biotech Associates Inc., Birmingham, AL, USA) jako odczynnik wykrywający. Stężenie ludzkiej IgG w surowicy określano w ilościowym teście ELISA, jak opisano uprzednio (Kacskovics i wsp., 06a). Do oceny stężenia w osoczu mysiej IgG1 skierowanej przeciwko owalbuminie w trakcie doświadczenia zastosowano ilościową ELISA, wykorzystując owalbuminę (Sigma) jako odczynnik wychwytujący i sprzężoną z HRP, oczyszczoną metodą powinowactwa poliklonalną, kozią antymysią IgG (swoistą wobec łańcucha ) (Southern Biotech Associates Inc., Birmingham, AL, USA) jako odczynnik wykrywający. Przeciwciało sprzężone z peroksydazą wykrywano, stosując jako substrat TMB (3,3,, - tetrametylobenzydynę; Sigma). Próbki badano w dwóch powtórzeniach. Stężenie Ig podawano w odniesieniu do wzorca referencyjnego. [0142] Analizę średniego stężenia IgG myszy w pierwszych dniach przeprowadzono przez dopasowanie danych do modelu dwukompartmentowego, stosując profesjonalne oprogramowanie WinNonLin, wersja 4.1 (Pharsight, Mountain View, CA, USA).
85 [0143] Krzywe usuwania mysiej IgG były dwufazowe, przy czym faza 1 (faza alfa) odpowiadała zrównoważeniu między kompartmentem śródnaczyniowym i pozanaczyniowym, a faza 2 (faza beta) odpowiadała powolnej eliminacji. Modelowanie matematyczne faz 1 i 2 do godziny 216 wykazało dobrą korelację z ogólnym schematem zależnej od FcRn farmakokinetyki IgG (Lobo i wsp., 04), autorzy obliczyli zatem fazę alfa i beta czasu półtrwania migg w tym przedziale czasowym. Szacowana faza alfa czasu półtrwania była podobna u myszy wt i Tg i wynosiła około godzin. Stwierdzano natomiast istotną różnicę (p<0,0) w fazie beta czasu półtrwania, ponieważ trwała ona 12,4 3,2 godziny (średnia SEM) u zwierząt wt i 16,1 7,8 godziny u zwierząt Tg, na podstawie analiz modelowania dwukompartmentowego (tabela ; Fig. 6A). [0144] W przypadku higg, podobnie do danych dotyczących eliminacji migg, modelowanie matematyczne faz 1 i 2 do godziny 216 danych pochodzących z tego badania wykazało dobrą korelację z ogólnym schematem zależnej od FcRn farmakokinetyki IgG; autorzy obliczyli zatem fazę alfa i beta higg w tym przedziale czasowym. Na podstawie analiz modelowania dwukompartmentowego faza alfa trwała około godzin u myszy zarówno typu dzikiego, jak i transgenicznych, bez różnic między doświadczeniami z dawką i mikrogramów na gram. W przypadku faz beta autorzy obserwowali istotne różnice (p<0,0) między myszami wt i Tg. Obliczone fazy beta trwały, odpowiednio, 6 3,6 i 171, 16,2 godziny po wstrzyknięciu dawki mikrogramów na gram, w porównaniu z 8,2 3,7 godziny i 181 7,7 godziny po
86 87 1 wstrzyknięciu dawki mikrogram na gram, u myszy wt i Tg (tabela ; Fig. 6B). Zgodnie z oczekiwaniem, wartości pola pod krzywą (AUC) były istotnie różne, jako że w doświadczeniu, w którym autorzy wstrzykiwali mikrogramów na gram, były one mniej więcej podwojone w porównaniu z doświadczeniem z dawką mikrogramów na gram; z drugiej strony ani faza alfa, ani faza beta czasu półtrwania nie wykazywały istotnych różnic po wstrzyknięciu lub g higg na gram. [014] Gdy autorzy porównywali dane dotyczące eliminacji migg z danymi dotyczącymi eliminacji higg w tych doświadczeniach (do porównania wzięto doświadczenie z dawką mikrogramów IgG na gram), obserwowali, że higg ulegało eliminacji znamiennie szybciej (p<0,0) u myszy wt (6 w porównaniu do 12, godzin, odpowiednio, w przypadku IgG ludzkiej i mysiej), nie stwierdzano jednak różnic w danych dotyczących eliminacji mysiej i ludzkiej IgG u zwierząt Tg (tabela ). Tabela. Parametry farmakokinetyczne IgG mysiej i ludzkiej u transgenicznych myszy typu dzikiego i bfcrn. Wartości odpowiadają średniej SEM; MRT średni czas przebywania leku w organizmie; AUC pole pod krzywą WTmIgG TGmIgG Dawka g/g m. ciała Faza alfa (h) czasu półtrwania Faza beta (h) czasu półtrwania MRT (h),2 0,6 12,4 3,2 167,7,6 4,6 0,2 16,1 7,8 227,2 11,9 AUC (h* g/ l) 12444, ,3 78,8 n WThIgG 8,2 1,8 6 3,7 139,8 4,8 731,2 376, 4
87 88 TGhIgG 12,7 1,8 171, 16,2 227,9 21, ,9 WThIgG,8,7 8,2 3,7 11,8,8 1774, TGhIgG 13,2 7, 181 7,7 26,1,9 788, 18,4 4 1 [0146] We wniosku można stwierdzić, że te badania eliminacji IgG u myszy transgenicznych pod względem BAC potwierdziły, że nie ma istotnego niedoboru 2m, jak również to, że bydlęcy łańcuch alfa FcRn tworzy funkcjonalny kompleks z mysim 2m. Eliminacja migg1 I była znamiennie zmniejszona u myszy Tg, co wskazuje, że ilość mysiego i hybrydowego FcRn, dostępnego do ochrony podawanych przeciwciał, przyczynia się do stopnia ochrony przed usuwaniem in vivo. Dodatkowo nie stwierdzano różnic między danymi dotyczącymi eliminacji mysiej i ludzkiej IgG u zwierząt Tg, co wskazuje, że hybrydowy receptor m 2m z łańcuchem alfa bfcrn może także chronić higg1. Przykład - Immunizacja owalbuminą [0147] Homozygoty nr 14, hemizygoty nr 19 (kodującym, odpowiednio, 4 i kopie bfcgrt) oraz dobrane pod względem wieku i płci (samce) myszy wt (po pięć na grupę) immunizowano dootrzewnowo g owalbuminy (OVA, Sigma-Aldrich, Budapeszt, Węgry) w kompletnym adiuwancie Freunda (CFA) i eksponowano je 14 dni później na g OVA z niekompletnym adiuwantem Freunda (IFA, Sigma-Aldrich, Budapeszt, Węgry). [0148] Na wszystkie procedury doświadczalne wyraziła zgodę Komisja Opieki nad Zwierzętami i Etyki
88 Wykorzystania Zwierząt Ośrodka Rolnictwa i Biotechnologii w Gödöllő, Węgry. Miana antygenowo swoistych IgM i IgG [0149] W celu wykrycia konsekwencji immunologicznych nadmiernej ekspresji bfcrn, oprócz ochrony IgG, myszy wt i Tg (nr 14 i nr 19) immunizowano, eksponowano 2 tygodnie później OVA i oznaczano miana IgM i IgG przeciwko OVA w ich surowicy. Próbki krwi (0 l/pobranie) ze splotu zaoczodołowego pobierano przez 6 dni. Surowice testowano pod kątem IgM i IgG swoistych wobec OVA i FITC. Do oceny stężenia w surowicy mysich IgM i IgG skierowanych przeciwko OVA i przeciwko FITC w trakcie doświadczenia zastosowano ilościowy test ELISA z wykorzystaniem OVA i FITCalbuminy (Sigma-Aldrich, Budapeszt, Węgry) jako odczynników wychwytujących i sprzężonych z HRP, oczyszczonych metodą powinowactwa poliklonalnych, kozich, antymysich IgM i kozich antymysich IgG (swoistych wobec łańcuchów i ) (Southern Biotech Associates Inc., Birmingham, AL, USA) jako odczynników wykrywających. Przeciwciało sprzężone z peroksydazą wykrywano, stosując jako substrat TMB (Sigma-Aldrich, Budapeszt, Węgry). Zastosowano seryjne rozcieńczenia poszczególnych badanych próbek surowicy i odnotowano stężenie Ig na podstawie wartości absorbancji przy 40 nm, interpolowanych z liniowej części krzywek zależności odpowiedzi od dawki. Próbki badano w dwóch powtórzeniach. Do oceny istotności statystycznej średnich wartości z grup terapeutycznych zastosowano dwustronny test t-studenta. Wartości uznawano za znamiennie różne, gdy p<0,0.
89 [0] Wyniki wykazały brak różnic między zwierzętami wt i Tg w trakcie pierwotnej odpowiedzi immunologicznej, lecz po immunizacji przypominającej miana swoistych wobec OVA IgM i IgG były wyraźnie różne. W trakcie wtórnej odpowiedzi przeciwciał, miana IgM u zwierząt wt wykazywały typową krzywą i nieco mniejszy pik IgM w porównaniu z pikiem w pierwotnej odpowiedzi immunologicznej. Z drugiej strony, w przypadku myszy Tg miana IgM były wyższe we wtórnej odpowiedzi immunologicznej w porównaniu z pierwotną odpowiedzią immunologiczną, i istotnie wyższe, niż u myszy wt (Fig. 7A). Jeżeli chodzi o miana IgG, miana IgG swoistej wobec OVA były niemal trzykrotnie wyższe u myszy Tg w porównaniu ze zwierzętami wt w czasie wtórnej odpowiedzi immunologicznej (Fig. 7B). Profil podklasy IgG [011] Następnie oznaczono profil podklasy IgG swoistych wobec OVA immunoglobulin surowicy. W 32 dni po immunizacji surowice myszy wt i Tg (nr 14) poddano badaniom pod kątem swoistych wobec OVA izotypów IgG. Do oceny stężenia w osoczu izotypów IgG przeciwko OVA zastosowano ilościowy test ELISA, z wykorzystaniem OVA jako odczynników wychwytujących i sprzężonych z HRP, oczyszczonych metodą powinowactwa poliklonalnych kozich, antymysich IgG1, IgG2a, IgG2b i IgG3 (Southern Biotech Associates Inc., Birmingham, AL, USA) jako odczynników wykrywających. Przeciwciało sprzężone z peroksydazą wykrywano, stosując jako substrat TMB. Stosowano seryjne rozcieńczenia każdej z badanych próbek surowicy i odnotowywano stężenie Ig na podstawie wartości absorbancji przy długości fali 40 nm,
90 interpolowanej z liniowej części krzywej zależności odpowiedzi od dawki. Próbki badano w dwóch powtórzeniach. [012] Stwierdzono, że zwierzęta immunizowane OVA-CFA i następnie OVA-IFA wykazywały dominację podklasy IgG1 przeciwciał przeciwko OVA. Dane wykazywały, że myszy Tg generowały istotnie wyższe miana swoistych wobec OVA IgG1, IgG2a i IgG2b. Jakkolwiek myszy Tg generowały nieco wyższy poziom IgG3 w porównaniu z myszami wt, różnica nie była znamienna ze względu na duże odchylenie standardowe wyników. Przedstawiane tu dane wykazały także, że izotypy IgG swoiste wobec OVA miały podobne proporcje, jak u myszy wt (Fig. 7C). To wskazuje, że poza różnicami ilościowymi, ekspresja bfcrn u myszy Tg nie zmienia swoistej wobec OVA odpowiedzi immunologicznej. Poziom IgG całkowitej [013] Do oceny stężenia w osoczu mysiej IgG w trakcie doświadczenia zastosowano ilościową ELISA, z wykorzystaniem niewyznakowanego, oczyszczonego metodą powinowactwa, koziego, antymysiego przeciwciała poliklonalnego (kozia antymysia IgG (H+L); Southern Biotechnology Associates, Inc., Birmingham, AL, USA) jako odczynnika wychwytującego i sprzężonej z peroksydazą chrzanową, oczyszczonej metodą powinowactwa, poliklonalnej, koziej, antymysiej IgG (swoistej wobec łańcucha ; Southern Biotechnology Associates, Inc., Birmingham, AL, USA) jako odczynnika wykrywającego. Przeciwciało sprzężone z peroksydazą wykrywano, stosując jako substrat TMB (Sigma). Próbki badano w dwóch powtórzeniach.
91 [014] Analiza poziomu IgG całkowitej wykazała, że myszy Tg (nr 14) wytwarzały znamiennie większe ilości IgG w porównaniu z myszami wt, nawet przed immunizacją (2,4 ± 0,4 i 4,8 ± 0, mg/ml, średnia ± SEM, odpowiednio, u myszy wt i Tg; p<0,01). Po immunizacji poziom IgG całkowitej wykazywał ciągły wzrost i osiągał swój szczyt w dniach 28 i 36, odpowiednio, u zwierząt wt i Tg. Godne uwagi jest to, że autorzy stwierdzili zaznaczoną i znamienną różnicę najwyższego poziomu IgG, który wynosił 14,8 ± 2,6 mg/ml (średnia ± SEM) i 39,9 ± 2,7 mg/ml, odpowiednio, u myszy wt i Tg (p<0,001) (Fig. 7D). Test fagocytozy [01] W celu oceny funkcjonalnej integralności IgG swoistej wobec OVA, wytworzonej u myszy Tg, wytworzono kompleksy OVA-przeciwciało anty-ova poprzez inkubację OVA-FITC z coraz większą ilością surowicy myszy Tg i wt, produkt zebrano po 3 dniach od immunizacji OVA, i przeprowadzono test fagocytozy na makrofagach linii mysiej P388D1. Znakowanie OVA znacznikiem FITC przeprowadzono według procedury opisanej w instrukcji producenta (Molecular Probes, Eugene, OR, USA). Makrofagi mysiej linii komórkowej P388D1 hodowano w temperaturze 37 C w pożywce RPMI 1640 (Sigma-Aldrich, Budapeszt, Węgry) z dodatkiem % płodowej surowicy cielęcej (Sigma-Aldrich, Budapeszt, Węgry) i M 2- merkaptoetanolu (Sigma-Aldrich, Budapeszt, Węgry). 2x komórek/studzienkę traktowano w temperaturze 37 C przez 60 minut 1 ng octanu mirystynianu forbolu (PMA, ng/ml, Sigma-Aldrich, Budapeszt, Węgry) w 24- studzienkowej płytce do hodowli komórkowej. W celu
92 93 uzyskania kompleksu OVA-przeciwciało anty-ova, 0 g OVA-FITC (1 g/ l) poddano wstępnej inkubacji w 1 2 temperaturze 37 C przez 60 minut z 2,, i 40 l surowicy od immunizowanych OVA myszy Tg i wt (3 dni po immunizacji). Fagocytozę prowadzono w temperaturze 37 C przez 90 minut poprzez inkubację potraktowanych PMA komórek P388D1 z kompleksami OVA-przeciwciało anty-ova. Po intensywnym płukaniu celem usunięcia niezwiązanych białek, komórki analizowano metodą cytometrii przepływowej (FACS). Do przerwania reakcji FITC pozakomórkowe-wyznakowana OVA zastosowano błękit trypanu. [016] Analizowano skuteczność wychwytu kompleksów immunologicznych metodą FACS. Przy zastosowaniu 2, l surowicy, bądź myszy Tg, bądź wt, nie stwierdzono wzmożenia fagocytozy w porównaniu z wychwytem nieopsonizowanego OVA-FITC. Istotne wzmożenie obserwowano jednak po zastosowaniu l surowicy myszy Tg, ale nie myszy wt. I wreszcie, 40 l surowicy od myszy Tg i wt wywołało takie samo wzmożenie fagocytozy w porównaniu z wychwytem nieopsonizowanego antygenu (Fig. 7E). Dane te wskazują, że surowica myszy Tg zawiera znamiennie więcej swoistego wobec OVA przeciwciała w dniu 3 po immunizacji, i że przeciwciało to było funkcjonalnie nienaruszone. Przykład 6 Myszy Tg i wt podobnie reagują na immunizację FITC-dekstranem [017] W celu analizy odpowiedzi immunologicznej, generującej głównie IgM, myszy Tg i wt immunizowano dootrzewnowo FITC-dekstranem i eksponowano na jego działanie 2 tygodnie. Jako że FITC-dekstran jest
93 haptenem (FITC) sprzężonym z typowym antygenem typu II niezależnym od komórek T (dekstran) (Mond i wsp., 199), w surowicy obserwowano głównie swoistą wobec FITC IgM, przy bardzo małej ilości IgG swoistej wobec FITC. Nie było różnic między zwierzętami Tg i wt pod względem swoistej wobec FITC odpowiedzi immunologicznej. Analiza odpowiedzi immunologicznej w trakcie immunizacji wykazała zwiększenie wytwarzania IgM po drugiej immunizacji, zarówno u myszy Tg, jak i wt (Fig. 8). Przykład 7 - Efekt nadmiernej ekspresji bfcrn na komórki wytwarzające IgM i IgG swoiste wobec OVA Analiza komórek B swoistych wobec OVA testem ELISPOT [018] Podwyższenie miana swoistych wobec OVA IgM u myszy Tg sugerowało, że nadmierna ekspresja bfcrn modyfikuje odpowiedź immunologiczną w odniesieniu do klonalnej ekspansji komórek B, niezależnie od tego, że przyczynia się do zwiększenia ochrony IgG. W celu sprawdzenia tej obserwacji, analizowano najpierw liczbę komórek plazmatycznych wydzielających IgM anty-ova i IgG anty-ova w śledzionie w dniu 2 po immunizacji poprzez analizę komórek B swoistych wobec OVA testem ELISPOT. Homozygoty nr 14 i myszy wt (po trzy na grupę) immunizowano dootrzewnowo g OVA w CFA i eksponowano 14 dni później na g OVA z IFA. 2 dni po pierwszej immunizacji zwierzęta uśmiercano i analizowano komórki ich śledziony pod kątem komórek wytwarzających IgM i IgG swoiste wobec OVA. Do badania ELISPOT, płytki MultiScreen-HTS (Millipore, Bedford, MA, USA) powlekano g/ml OVA w PBS w temperaturze pokojowej przez 3 godziny. Płytki płukano następnie
94 9 sześciokrotnie PBS i blokowano pożywką RPMI, 1 2 zawierającą % FCS i merkaptoetanol (0 M) przez minut w temperaturze pokojowej. Do studzienek dodawano seryjne rozcieńczenia (od x komórek/studzienkę) komórek śledziony. Płytki inkubowano w temperaturze 37 C z % CO 2 przez noc i płukano sześciokrotnie PBS- Tween, po czym do każdej studzienki dodano sprzężoną z peroksydazą chrzanową kozią, antymysią IgG (swoistą dla łańcucha ciężkiego ; rozcieńczenie 1:4000; Southern Biotechnology). Po jednogodzinnej inkubacji w temperaturze pokojowej płytki przepłukano sześciokrotnie PBS-Tween. Płytki inkubowano następnie w obecności substratu chromogennego 3-amino-9- etylokarbazolu (AEC; Sigma) i H 2 O 2 w temperaturze pokojowej i reakcję zakończono przez płukanie wodą. Zliczano jednostki tworzące plamki (ang. spot-forming units - SFU) na studzienkę w czytniku ImmunoScan ELIspot (Cellular Technology Ltd., USA) i oceniano z użyciem oprogramowania ImmunoSpot, wersja 3.2 (Cellular Technology Ltd., USA). [019] Wynik wykazał, że w śledzionie liczba komórek wytwarzających IgM swoistą wobec OVA uległa podwojeniu (p<0,0), a liczba komórek wytwarzających IgG swoistą wobec OVA uległa zwiększeniu ponad trzykrotnemu (p<0,001) (Fig. 9A). Immunizacja powoduje masywny napływ neutrofili do śledziony [0160] Immunizacja OVA powoduje zwiększenie masy śledziony (Fig. 9B) i liczby jej komórek (Fig. 9C): obserwowano także powiększenie obwodowych węzłów chłonnych. Zjawisko to było obecne u myszy wt, jak
95 również u myszy Tg, lecz wielkość śledziony u zwierząt Tg była dwukrotnie większa, niż u kontrolnych zwierząt wy (p<0,001). [0161] Autorzy określili zatem charakterystykę dystrybucji typów komórek śledziony metodą cytometrii przepływowej. Do FACS izolowano splenocyty i najpierw inkubowano je z anty-cd32/cd16 (klon 2.4G2) przez minut. Następnie komórki inkubowano ze sprzężonymi z fluorochromem swoistymi Ab w temperaturze 4 C przez 0 minut w buforze barwiącym (PBS z 0,1% BSA i 0,1% azydkiem sodowym), przepłukano dwukrotnie, po czym analizowano za pomocą FACSCalibur zaopatrzonego w oprogramowanie CellQuest (BD Biosciences, San Jose, CA, USA). Sprzężone mab, CD4R/B2-PECy. 1-A/1-E-PE, GR- 1(Ly-6G)-PE i CD11b-A647 uzyskano z firmy BD Pharmingen (San Diego, CA, USA). CD3-A647, CD86-PE, CD11c-A647 i CD11b-PE kupiono w firmach, odpowiednio, Caltag (Burlingame, CA, USA), ebioscience (San Diego, CA, USA), Serotec (Dusseldorf, Niemcy) i ImmunoTools GmbH (Friesoythe, Niemcy). Izotypy kontrolne uzyskano z firm BD Pharmingen lub Serotec. [0162] Wyniki wykazały, że po immunizacji odsetek komórek z antygenami CD4R/B2 (limfocyty B) i 1-A/I-E (klasa MHC II) ulegał istotnemu zmniejszeniu (p<0,0). Nie było różnic, lub zmniejszenie było znacznie mniej zaznaczone, w odniesieniu do komórek z markerem CD3 (limfocyt T) u myszy, odpowiednio, wt i Tg. Te zjawiska występowały u myszy wt i Tg, lecz były wyraźniejsze u zwierząt Tg (Fig. ). [0163] Odsetek komórek z markerami CD11b i Gr-1 był po immunizacji wybitnie zwiększony, zarówno u myszy wt,
96 jak i Tg (p<0,001). Jeżeli chodzi o rozmiary tego wzrostu, autorzy stwierdzili około -krotne zwiększenie u myszy wt i 9-krotne zwiększenie u myszy Tg. Autorzy stwierdzili również, że większość komórek, które wykazywały ekspresję Gr-1, stanowiły Gr-1 high (Fig. 11A) z typowym położeniem granulocyta według parametrów rozproszenia światła kąt przedni/kąt boczny (FCS/SSC), co sugeruje, że były to neutrofile (dane nieukazane). To zwiększenie również było bardziej zaznaczone u myszy Tg (p<0,01) (Fig. 11A). Autorzy stwierdzili również, że odsetek komórek z antygenami CD11b i klasy II MHC (makrofagi, komórki dendrytyczne) (Fig. 11B) i z antygenami CD11b i CD11c (komórki dendrytyczne) (Fig. 11C komórki bramkowane) był znamiennie podwyższony (odpowiednio, p<0,0 i p<0,01) u myszy Tg w porównaniu z ich kontrolami wt. Te wyniki wyjaśniają proporcjonalne zmniejszenie liczby komórek z B2 i MHC-II, nie implikuje to jednak zmniejszenia całkowitej liczby komórek, ponieważ śledziona i liczba komórek śledziony były po immunizacji większe (Fig. 9C). Na podstawie tych analiz autorzy wydedukowali, że większość komórek, które napływają do śledziony w 2 dni po immunizacji, stanowią neutrofile, jakkolwiek stwierdzono także zwiększenie liczby komórek dendrytycznych. Przykład 8 - bfcrn ulega silnej ekspresji w neutrofilach, makrofagach i komórkach dendrytycznych w wysięku z jamy otrzewnej [0164] Ekspresję łańcucha alfa bfcrn analizowano najpierw w komórkach z wysięku z jamy otrzewnej, pochodzących od myszy dwa dni po dootrzewnowym podaniu
97 konkanawaliny A (ConA, 0 g/ mysz w PBS; Sigma- Aldrich, Budapeszt, Węgry). W celu uzyskania preparatu komórkowego wzbogaconego w neutrofile, myszom wstrzykiwano i.p. 2 ml % (masa/obj.) kazeiny (Sigma- Aldrich, Budapeszt, Węgry) w jałowej soli fizjologicznej i 6 godzin po wstrzyknięciu izolowano komórki otrzewnej. Następnie neutrofile oczyszczano przez odwirowanie Ficoll-Paque Plus (GE Healthcare, Uppsala, Szwecja) (400 x g przez minut w temperaturze pokojowej). Czystość neutrofili wynosiła ~80%, co stwierdzono metodą cytometrii przepływowej, stosując jako odczynniki anty-cd11b i anty-gr-1 (Fig. 12). [016] RNA całkowite ekstrahowano z tych komórek, stosując odczynnik Trizol (Invitrogen, Carlsbad, CA, USA) i 2 mikrogramy RNA poddawano odwrotnej transkrypcji, stosując enzym odwrotnej transkryptazy mysiego wirusa białaczki Moloneya (M-MLV) (Promega) i primer (dt)17-adapter z wykorzystaniem standardowych protokołów. Wykonano PCR, uzyskując swoisty dla łańcucha alfa bfcrn amplikon o długości 422 par zasad ( par zasad AF1396) z parami primerów: B3 - CGCAGCARTAYCTGASCTACAA-3 (SEQ ID nr 37, gdzie R może stanowić G lub A, Y może stanowić T lub C, a S może stanowić G lub C), i B4 -GGCTCCTTCCACTC-CAGGTT-3 (SEQ ID nr 38). Powielony segment oddzielano elektroforetycznie na 1% żelu agarozowym i barwiono bromkiem etydowym. [0166] Ekspresję BFcRn wykrywano przez powielanie PCR najpierw w komórkach z wysięku z jamy otrzewnej od myszy Tg i wt po potraktowaniu ich ConA. Ekspresję
98 bfcrn wykazano również w populacji komórkowej wzbogaconej w neutrofile i makrofagi, ponieważ oczyszczone komórki stanowiły ~78% CD11b high / Gr-1 hight i ~% CD 11b high / Gr-1 - (Fig. 12). [0167] Później opublikowane badanie wykazało, że FcRn odgrywa główną rolę w zależnej od IgG fagocytozie w leukocytach z jądrem wielokształtnym i monocytach (Vidarsson i wsp., 06), co sugeruje, że FcRn uczestniczy w prezentacji antygenu. Na podstawie danych autorów niniejszego zgłoszenia można postawić hipotezę, że nadmierna ekspresja bfcrn w tych komórkach odpowiada za wzmożoną fagocytozę antygenu i nawet prezentację antygenu, co z kolei powoduje zwiększenie liczby komórek plazmatycznych, produkujących antygenowo swoiste IgM i IgG, we wtórnych narządach limfoidalnych. Jeżeli ta teoria jest prawdziwa, to zwiększenie jest oczywiste wtedy, gdy dojdzie do wytwarzania IgG, i nie wcześniej. Istotnie, podwyższenie miana IgM stwierdzano tylko po pojawieniu się IgG swoistej wobec OVA, głównie we wtórnej odpowiedzi immunologicznej. Przykład 9 - Obecność limfocytów B, neutrofili i komórek dendrytycznych z OVA w śledzionie immunizowanych myszy [0168] Przeprowadzona przez autorów analiza komórkowa wykazała, że dominującym typem komórek, napływających do śledziony po immunizacji, były neutrofile. W innym niedawno opublikowanym doniesieniu napisano, że neutrofile stanowią główną populację komórek z antygenem we wtórnych narządach limfoidalnych, gdy tylko dochodzi do swoistej odpowiedzi immunologicznej i powstania kompleksu immunologicznego IgG-antygen,
99 0 1 2 przyczynia się on do jakości uzyskanej wtórnej odpowiedzi immunologicznej (Maletto i wsp., 06). W celu zbadania, czy napływ neutrofili w opisywanych tu doświadczeniach w istocie zależy od antygenowo swoistej odpowiedzi immunologicznej i posiadanych antygenów, transgenicznym myszom 6 dni po immunizacji OVA (gdy poziom IgG swoistej wobec OVA był nadal wysoki - Fig. 7B) i bez immunizacji podawano dootrzewnowo FITC-OVA. [0169] W tym doświadczeniu myszom (homozygoty nr 14), podawano dootrzewnowo OVA-FITC (7,4 mg/ml, 0 l/mysz). godzin po wstrzyknięciu OVA-FITC myszom wycinano śledzionę i analizowano komórki śledziony metodami cytometrii przepływowej i mikroskopii konfokalnej. Do cytometrii przepływowej, komórki poddawano opisanej powyżej obróbce, z użyciem następujących odczynników: CD4R/B2-PECy, GR-1 (Ly- 6G)-PE, CD11c-A647, CD11b-A647 i CD11b-PE. Do mikroskopii konfokalnej komórki barwiono przenikalną dla komórek sondą DNA fluoryzującą w czerwieni - DRAQ (Biostatus Ltd., Wielka Brytania) przez min w temperaturze pokojowej. Po etapie płukania rejestrowano obrazy fluorescencji i DIC (12x12 pikseli) za pomocą konfokalnego laserowego mikroskopu skaningowego Olympus FLUOView00 (Hamburg, Niemcy), przy dużym powiększeniu (obiektyw 63x). Do wzbudzenia barwników FITC i DRAQ zastosowano laser argonowy o długości fali 488 nm i linie lasera 632 He-Ne. [0170] Uzyskane przez autorów dane z cytometrii przepływowej wykazują, że u zwierząt immunizowanych OVA wśród komórek OVA-FITC-dodatnich (1,1 1,4%), 61,2,4% było B2-dodatnich (limfocyty B), 18, 0,6%
100 1 wykrywano jako CD11b high i Gr-1 high (neutrofile), a 13, 1 2 2,1% było CD11b i CD11c-dodatnich (komórki dendrytyczne). Autorzy nie wykryli istotniejszej liczby komórek OVA-FITC-dodatnich u zwierząt nieimmunizowanych (2,1 0,3%) (Fig. 13A i 13B). Uzyskane przez autorów dane z cytometrii przepływowej, dotyczące OVA-FITCdodatnich neutrofili i limfocytów B, zostały potwierdzone metodą mikroskopii konfokalnej. Autorzy wykryli typowe neutrofile o wielopłatowych jądrach, internalizujące OVA-FITC, natomiast inne komórki, przypuszczalnie limfocyty B (z dużym, okrągłym jądrem i cienkim rąbkiem cytoplazmy) były powleczone OVA-FITC (Fig. 13C). Autorzy nie stwierdzili istotnych różnic w zakresie wielkości śledziony i liczby komórek śledziony pomiędzy zwierzętami immunizowanymi a nieimmunizowanymi (dane nieukazane). To odkrycie pasuje bardzo dobrze do innych wyników autorów i zapewnia wyjaśnienie zaznaczonego napływu neutrofili CD11b- i Gr-1-dodatnich u wykorzystanych w doświadczeniach myszy wt i Tg, 2 dni po immunizacji. Po uwzględnieniu tego, że myszy Tg wytwarzały znacznie więcej IgG swoistej wobec OVA (Fig. 7B) i wykazywały nadmierną ekspresję FcRn w tych komórkach (Fig. 12), autorzy są również w stanie wyjaśnić, czemu myszy Tg wykazują większy napływ tych komórek do śledziony, większą ekspansję klonalną swoistych antygenowo komórek B i w następstwie tego silną odpowiedź przeciwciał. Przykład Wytwarzanie myszy transgenicznych pod względem bfcrn poprzez transgenezę lentiwirusową (przykład przygotowawczy)
101 2 1 2 [0171] Niedawno zastosowano pochodzące z HIV i EIAV wektory lentiwirusowe do uzyskania powtarzalnego, wysokiego odsetka transgenezy u myszy (Pfeifer i wsp., 02), szczurów (Lois i wsp., 02), kur (McGrew i wsp., 04), świń (Hofmann i wsp., 03; Whitelaw i wsp., 04) i bydła (Hofmann i wsp., 04). Wektor transferowy WPRE-P2_bFcRn wytworzono przez zastąpienie pierwotnego segmentu EF1a-EGFP wektora transferowego pwpts-egfp (Bovia i wsp., 03) sztucznie skonstruowanym segmentem promotora o długości 290 par zasad, genu łańcucha alfa bydlęcego FcRn (bfcrn) z sekwencją kodującą tego samego genu. Segment promotora bfcrn powielono metodą PCR, stosując polimerazę Deep Vent (New England Biolab, Beverly, MA, USA) z klonu BAC nr 128E04, zawierającego gen łańcucha alfa bfcrn, i zastosowano do uzyskania myszy transgenicznych pod względem bfcrn (zob. przykład 2). Primer forward zawiera miejsce XbaI (kursywą) (lenti-bore: -GGG TCT AGA ACA CCA AGG GCG GCA TCA-3, SEQ ID nr 39); primer reverse zawiera EcoRI (lenti-bore18: -GGG GAA TTC CGG CTC CCG TGA CTG GAG AC-3, SEQ ID nr 40). Wytworzony metodą PCR amplikon był to fragment sekwencji regulatorowej bfcgrt o długości 290 par zasad (GenBank: nukleotyd od 764 do 762 pary zasad klonu NW /Bt18 WGA2132 3/ contig genomowy chromosomu 18 Bos taurus, montaż referencyjny (na podstawie Btau_3.1). Segment kodujący łańcucha ciężkiego bfcrn powielono metodą PCR z użyciem polimerazy Deep Vent z klonu zawierającego cdna bfcrn (GenBank AF1396); uprzednio stosowano go w laboratorium autorów do wytwarzania stabilnie
102 3 1 2 transfekowanych komórek (Kacskovics i wsp., 06a). Primer forward zawiera miejsce EcoRI (BORE: -GGG GAA TTCTGG GGC CGC AGA GGG AAG G-3, SEQ ID nr 41); primer reverse zawiera miejsce MluI (lenti-bore19: -GGG ACG CGTGAG GCA GAT CAC AGG AGG AGA AAT-3, SEQ ID nr 42). W wyniku PCR uzyskano amplikon o długości 128 par zasad cdna łańcucha alfa bfcrn (GenBank: nukleotydy sekwencji referencyjnej NM_17667 bfcrn). Po oczyszczeniu oba amplikony (promotor i sekwencję kodującą bfcrn) strawiono EcoRI (Promega) i poddano ligacji ligazą T4 (Promega), uzyskując konstrukt P2- bfcrn, kodujący segment promotora łańcucha ciężkiego bfcrn o długości 290 par zasad i segment łańcucha ciężkiego bfcrn o długości 128 par zasad (SEQ ID nr 43). Promotor bfcrn-fragment cdna wprowadzono następnie do plazmidu wektora transferowego pwpts. Wektory powielono przez przemijającą transfekcję do komórek 293T, jak opisano uprzednio (Bovia i wsp., 03). Wektory użyte do stworzenia lentiwirusa uzyskano z Tronolab, Kantonalny Uniwersytet Medyczny, Genewa, Szwajcaria. Użyto następujących wektorów: pmd.g (konstrukt kopertowy, 6kbp); pcmv R8.91 (konstrukt pakujący, 12kbp); pwpts (konstrukt transferowy, 12,7kbp, w którym między miejsca ClaI a SalI wprowadzono promotor FcRn (P2)-sekwencję cdna FcRn. Miano lentiwirusa oszacowano przez transdukcję komórek Jurkat seryjnymi rozcieńczeniami nadsączy równolegle sporządzonego roztworu macierzystego wirusa. Ten wirus zawierał wektor transferowy pwpre-egfp. Komórki GFP+ zliczano w aktywowanym fluorescencją sorterze komórek (FACS). Miano mieściło się w zakresie 7-8 jednostek
103 4 1 transdukcji na mililitr. Myszy transgeniczne uzyskano przez wstrzyknięcie HIV-P2-FcRn lentiwirusa do przestrzeni wokół pęcherzyka źółtkowego zygot w stadium jednej komórki. Mikroiniekcje prowadzono na podłożach genetycznych FVB/N i Balb/c. Po wstrzyknięciu zygoty przenoszono do samic-biorców (tabela 6). Transgenicznych założycieli zidentyfikowano metodą PCR z użyciem następujących primerów: bfcsuf CTCCTTTGTCTTGGGCACTT 3 (SEQ ID nr 9) i bfcl GCCGCGGATCCCTTCCCTCTG 3 (SEQ ID nr ). Tabela 6. Dane na temat zarodków mysich po wstrzyknięciu lentiwirusa FVB/N Balb/c Zarodki po wstrzyknięciu i transferze Liczba samic-biorców 2 3 Udane transfery 2 1 Liczba noworodków 17 lub 18 4 Liczba żywych noworodków 13 4 PCR+ (transgeniczne) 1 [0172] W celu wykrycia ekspresji transgenu wykonano RT- PCR na próbkach RNA izolowanego z wątroby, śledziony, płuca i jelita noworodków myszy HIV-P2-FcRn, jak przedstawiono na Fig. 14. Włączono primery planowane dla promotora P2 (FcRnprf: CGGCCACCTCTATCACATTT, SEQ ID nr 3; FcRnprr: TGCATTGACCACACTTGGTT, SEQ ID nr 4; oczekiwana wielkość fragmentu: 79 par zasad), żeby wykluczyć możliwość zanieczyszczenia DNA genomowego w próbkach RNA, i wykrywano mrna swoiste wobec transgenu przy użyciu primerów planowanych dla egzonu 4 bydlęcego FcRn (bfcrnex4f: CCAAGTTT-GCCCTGAACG, SEQ ID nr 19; bfcrnex4r: GAGGCAGATCACAGGAGGAG, SEQ ID nr 44,
104 1 2 oczekiwana wielkość fragmentu: 161 par zasad). Wyniki wyraźnie wskazują na ekspresję mrna bfcrn. [0173] Transgenicznych założycieli hodowano indywidualnie przez krzyżowanie z myszami typu dzikiego, żeby uzyskać linie transgeniczne. Przykład 11 - Poziom albuminy również ulega podwyższeniu w surowicy myszy Tg (przykład stanu wyjściowego) [0174] Jako że myszy transgeniczne wykazują nadmierną ekspresję bfcrn, badano dwie kwestie: (1) czy bfcrn wchodzi w interakcje z mysią albuminą i (2) czy istnieje wpływ na metabolizm albuminy. Mierzono zatem poziom albuminy u myszy niewykazujących ekspresji FcRn (FcRn KO zakupione w firmie Jackson Laboratory, USA), zwierząt wt, homozygot nr 14 i nr 19 (ekspresja, odpowiednio, 4 i kopii bfcrn) przez zastosowanie swoistego dla albuminy testu ELISA kanapkowego. Na podstawie tego badania stwierdzono, że bfcrn wiąże się z albuminą i poważnie wpływa na jej poziom w surowicy (Fig. 1). Oznaczono także zawartość albuminy w mleku transgenicznych myszy i stwierdzono, że zawiera ono istotnie wyższe stężenie w porównaniu z kontrolami typu dzikiego (dane nieukazane). Te dane sugerują więc, że oprócz korzystnego efektu nadmiernej ekspresji FcRn na humoralną odpowiedź immunologiczną, podwyższone jest także stężenie albuminy, co mogłoby być szkodliwe, ponieważ podwyższa ponad normę ciśnienie osmotyczne we krwi, jakkolwiek nawet przy liczbie kopii wynoszącej nie zauważono żadnych szkodliwych efektów, a zwierzęta Tg żyły długo i w dobrym stanie zdrowia do 14 miesięcy.
105 6 1 2 Przykład 12 Badanie in vivo z analizą czasu półtrwania króliczej IgG u myszy FcRn KO - myszy transgenicznych pod względem bydlęcego BAC (przykład stanu wyjściowego) [017] W celu dokonania analizy, czy bfcrn wiąże się z króliczą IgG i chroni ją przed szybką eliminacją, autorzy analizowali czas półtrwania króliczej IgG u myszy FcRn KO - myszy transgenicznych pod względem bydlęcego BAC. Uzyskano transgeniczne pod względem bydlęcego FcRn, pozbawione mysiego łańcucha alfa FcRn myszy (bfcrn/mfcrn -/- ) przez skrzyżowanie homozygotycznych myszy bfcrn (nr 14) z 4 kopiami transgenu BAC 128E04 (Bender i wsp., 07) ze szczepem myszy homozygotycznym pod względem allelu knockout mfcrn. Ten szczep myszy nabyto w firmie Jackson Laboratories (USA) pod nazwą: B6,129X1-Fcgnt cm1der/der. Podwójnie transgeniczne myszy o pożądanym genotypie (bfcrn/mfcrn -/- ) wybrano na podstawie multipleksowej PCR z użyciem następujących primerów: NEO-F: GGA ATT CCC AGT GAA GGG C (SEQ ID nr 4); NEO-R: CGA GCC TGA GAT TGT CAA GTG TAT T (SEQ ID nr 46); FcRn wt-f: GGG ATG CCA CTG CCC TG (SEQ ID nr 47); bfcsuf: CTC CTT TGT CTT GGG CAC TT (SEQ ID nr 9); bfcl: GCC GCG GAT CCC TTC CCT CTG (SEQ ID nr ). Zidentyfikowano odpowiednie myszy F2, jak ukazano na Fig. 16A. [0176] Czas półtrwania króliczej IgG analizowano w zasadzie tak, jak opisano w przykładzie 4. Pokrótce, po wstępnym pobraniu krwi, dopasowanym pod względem wieku, masy ciała i płci (samce) myszom mfcrn -/- i bfcrn+ / +mfcrn -/- (po trzy na grupę) podawano przez mikroiniekcję dożylnie g króliczej IgG (Sigma) w
106 mg/ml roztworze soli fizjologicznej i w ciągu kolejnych 216 godzin, okresowo pobierano próbki krwi (0 l/pobranie) ze splotu zaoczodołowego. Do oceny stężenia w osoczu króliczej IgG w trakcie doświadczenia stosowano ilościowy test ELISA z wykorzystaniem koziej, antykróliczej IgG (swoistej wobec H+L) (Caltag) jako odczynnika wychwytującego i sprzężonej z HRP koziej, antykróliczej IgG (swoistej wobec H+L) (Vector Laboratories) jako odczynnika wykrywającego. Przeciwciało sprzężone z peroksydazą wykrywano, stosując jako substrat TMB (3,3,, -tetrametylobenzydyna: Sigma). Próbki badano w dwóch powtórzeniach. Stężenie króliczej IgG podawano w odniesieniu do wzorca referencyjnego. Królicza IgG ulegała szybko eliminacji u myszy FcRn -/- (czas półtrwania: 1 godzin), natomiast u myszy bfcrn/fcrn -/- była chroniona (czas półtrwania: 67 godzin) (Fig. 16B). Szybka eliminacja IgG u zwierząt FcRn -/- jest udokumentowana (Roopenian i wsp., 03), podobnie jak u zwierząt pozbawionych funkcjonalnego FcRn z powodu braku beta-2-mikroglobuliny (Ghetie i wsp., 1996; Israel i wsp., 1996: Junghans i Anderson, 1996). W doświadczeniu autorów obserwowano znamienne zmniejszenie eliminacji króliczej IgG (p<0,0001) u zwierząt bfcrn/mfcrn -/- z powodu aktywności bfcrn. Niemniej czas półtrwania króliczej IgG u tych zwierząt jest nadal znacznie krótszy niż czas półtrwania króliczej IgG u królika (czas półtrwania: godzin (Andersen i Bjorneboe, 1964: Dima i wsp., 1983; Sabiston i Rose, 1976) lub u myszy (czas półtrwania: 6 godzin) (Dima i wsp., 1983), co sugeruje, że interakcja króliczej IgG i kompleksu bfcrn/mysia beta-
107 mikroglobulina jest względnie słaba. W istocie, uprzednio wykrywano różnice w interakcjach IgG-FcRn pomiędzy poszczególnymi gatunkami (Ober i wsp., 01), co podkreśla konieczność doświadczalnej selekcji FcRn odpowiedniego do wydłużania czasu półtrwania IgG i wskutek tego jego poziomu po immunizacji. Przykład 13 - Uzyskiwanie przez krzyżowanie podwójnie transgenicznych królików, wytwarzających humanizowane immunoglobuliny i wykazujących nadmierną ekspresję bydlęcego FcRn (przykład przygotowawczy) [0177] Podwójnie transgeniczne linie królików, wytwarzających IgG ludzką i bydlęcy FcRn, dobrze nadają się do ulepszania poziomu immunoglobulin w surowicy. Już wykazano efekt nadmiernej ekspresji bydlęcego FcRn na czas półtrwania ludzkiej IgG, te podwójnie transgeniczne linie królików są zatem bardzo użyteczne w wytwarzaniu poliklonalnych surowic odpornościowych lub jako punkt wyjściowy do uzyskiwania króliczych przeciwciał monoklonalnych. [0178] Podwójnie Tg królika można uzyskać przez standardowe krzyżowanie. Mimo, że w poprzednim doświadczeniu wykazano, że bfcrn nie jest optymalny w odniesieniu do wiązania się z króliczą IgG, uzyskiwanie królików transgenicznych pod względem bfcrn byłoby nadal pożądanym etapem pośrednim w uzyskiwaniu podwójnie transgenicznych królików, wytwarzających zarówno ludzką IgG, jak i bydlęcy FcRn. W późniejszym etapie można skrzyżować dwie Tg linie królików, z których każda posiada jeden transgen, w celu uzyskania ostatecznego zwierzęcia.
108 9 1 2 [0179] Bydlęcy klon BAC 128E04 wprowadzono na drodze mikroiniekcji do przedjądrza zygot królików, uzyskując transgeniczne zwierzę założycielskie. Obecność bydlęcego genu FcRn wykrywano metodą PCR, z primerami zaplanowanymi tak, żeby uzyskać powielenie fragmentu o długości 160 par zasad z egzonu 4 bydlęcego FCGRT: bfcrnex4f: -CCAAGTTTGCCCTGAACG - 3 (SEQ ID nr 19) i bfcrnex4r: - GTGTGGGCAGGAGTAGAGGA - 3 (SEQ ID nr ). Fig. 17 ukazuje obecność fragmentu zgodnie z oczekiwaniami. [0180] Mioty F1 królika-założyciela będą nadawały się do krzyżowania z genetycznie zmodyfikowanymi królikami, wytwarzającymi humanizowane immunoglobuliny (Thorey i wsp., 06): w wyniku tego uzyskuje się podwójnie transgeniczne zwierzęta, wykazujące zwiększony poziom wytwarzania humanizowanych przeciwciał poliklonalnych. Przykład 14 - Uzyskiwanie podwójnie transgenicznych królików, wytwarzających humanizowane immunoglobuliny i wykazujących nadmierną ekspresję bydlęcego FcRn (przykład przygotowawczy) [0181] Zamiast wykorzystania sposobu z przykładu 13, linie podwójnie transgenicznych królików można tworzyć poprzez wykorzystanie transgenicznych królików, produkujących humanizowane immunoglobuliny (Thorey i wsp., 06) jako zwierząt-biorców w doświadczeniu z transgenezą podobną do opisanej w przykładzie 3. Stosując bydlęcy klon BAC 128E04, uzyskuje się linie transgenicznych królików poprzez mikroiniekcję do przedjądrza. Transgeniczne króliki identyfikuje się metodą Southern blot i(lub) PCR, stosując primery i sondy swoiste dla bydlęcego klonu BAC, i ocenia się
109 1 1 2 czas półtrwania wstrzykniętej ludzkiej IgG. Znacząco dłuższy czas półtrwania w surowicy królików wskazuje, że wprowadzony bydlęcy FcRn chroni ludzką immunoglobulinę, wytworzoną przez transgenicznego królika, przed rozpadem. Przykład 1 - Izolowanie i określanie charakterystyki klonu BAC, zawierającego króliczy gen łańcucha alfa FcRn (przykład stanu wyjściowego) [0182] Ponieważ w wielu z dawniejszych badań nad aktywnością FcRn jako model wykorzystywano królika, przewidywano, że króliczy FcRn ulega ekspresji w komórkach śródbłonka (oraz w innych tkankach) i przyczynia się do homeostazy IgG. Dodanie dodatkowych kopii króliczego genu FcRn powoduje wydłużenie czasu półtrwania IgG. Przyjmując takie założenia, dokonano klonowania i sekwencjonowania króliczego FcRn (SEQ ID nr 48). [0183] Równolegle izolowano króliczy klon BAC 262E02 z króliczej biblioteki BAC (Rogel-Gaillard i wsp., 01). Bibliotekę BAC skonstruowano w wektorze pbelobac11; DNA o dużej masie cząsteczkowej uzyskano z krwinek białych królika nowozelandzkiego. Królicza biblioteka BAC jest prowadzona przez ośrodek zasobów zwierząt domowych INRA i jest publicznie dostępna. Program Primer 3 zaplanował primery swoiste dla króliczego genu FCGRT: OCU_FCGRT_F: GGGACTCCCTCCTTCTTTGT (SEQ ID nr 49) i OCU_FCGRT_R: AGCACTTCGAGAGCTTCCAG (SEQ ID nr 0) ( na ulegającym ekspresji u królika znaczniku sekwencji (EST) EB37777 zidentyfikowano przez dopasowanie go z ludzkim genem
110 111 1 FCGRT (GenBank NM_0047), z użyciem programu Iccare ( Sekwencja EB EST jest identyczna, jak odpowiadająca jej część króliczego cdna FcRn (SEQ ID nr ). Króliczy klon BAC 262E02 analizowano na obecność potencjalnych genów na podstawie ortologicznego chromosomu Bos Taurus. Zaplanowano primery dla EST Oryctolagus cuniculus lub zastosowano primery swoiste dla genów bydlęcych w sąsiedztwie (w odległości 0 kb) genu FCGRT w kierunkach i 3. Na klonie BAC 262E02 zidentyfikowano następujące geny królicze: RPL13A, RPS11, FCGRT, RCN3, PRRG2 (zob. Fig. 18). Tabela 7. Primery i warunki PCR do oceny obecności ortologicznych genów bydlęcych na króliczym klonie 262E02 Domniemany gen Primer PRRG PRRG2 L: - GTTGTTCACACCAGGGAACC-3 (SEQ ID nr 2) PRRG2_R: - CCTTTGCCATTGTAGATGTAGC- 3 (SEQ ID nr 26) RCN3 RCN3_L: - GACGCCGAGACCTACAAGAA-3 (SEQ ID nr 1) RCN3_R: - CATGTGCGGGAACTCCTC-3 (SEQ ID nr 2) FCGRT FCGRT_L: - CTGAACGGTGAGGACTTCAT-3 (SEQ ID nr 3) FCGRT_R: - GCTCCTTCCACTCCAGGTT-3 (SEQ ID nr 4) Temp. topnienia ( C) Produkt (par zasad)
111 112 1 RPS11 RPS11_L: - AGATCGGCGACATCGTCA-3 (SEQ ID nr ) RPS11_R: TCTGGAACTGCTTCTTGGTG.3 (SEQ ID nr 6) RPL13A RPL13A_L: CATGAGGTGGGCTGGAAGTA-3 (SEQ ID nr 7) RPL13A- R: - TCCGGTAGTGGATCTTAGCC-3 (SEQ ID nr 8) [0184] Wykryto również obecność genu FLT3LG, na podstawie analizy slot-blot, gdy bydlęcy produkt PCR FLT3LG o długości 47 par zasad, wytworzony przez primery: FLT3LG_L: -TCGGAGATGGAGAAACTGCT-3 (SEQ ID nr 1) i FLT3LG_R: -CTGGACGAAGCGAAGACAG-3 (SEQ ID nr 16) poddano hybrydyzacji do króliczego BAC 262E02, uzyskując silnie dodatni sygnał w warunkach bardzo rygorystycznych. [018] Struktura króliczego klonu BAC 262E02 jest, jak się wydaje, bardzo podobna do struktury bydlęcego klonu BAC 128E04, można zatem oczekiwać, że zapewni równie dobre wyniki w doświadczeniach z transgenezą. Przykład 16 - Uzyskiwanie transgenicznych królików, wykazujących nadmierną ekspresję króliczego FcRn, i badania in vivo analizy czas półtrwania króliczej IgG (przykład przygotowawczy) [0186] Do zygoty króliczej wprowadzono poprzez mikroiniekcję do przedjądrza konstrukt genetyczny, zawierający króliczy klon BAC 262E02 o właściwościach podanych w przykładzie poprzednim lub konstrukt zawierający królicze cdna FcRn (SEQ ID nr 48) pod kontrolą dobrze znanego promotora bfcrn, uzupełniony w dół króliczego cdna FcRn heterologicznym intronem i
112 113 1 dostępnym w handlu regionem SV40 polia. Zamiast tego transgeniczne króliki można uzyskać przez wprowadzenie konstruktu cdna P2-króliczy FcRn do lentiwirusowego wektora transferowego, jak opisano w przykładzie, i wstrzyknięcie do przestrzeni wokół pęcherzyka żółtkowego zarodków króliczych. Transgeniczne króliki identyfikuje się metodą Southern blot i(lub) PCR, stosując swoiste primery/sondy. Czas półtrwania IgG u transgenicznych królików, wykazujących nadmierną ekspresję króliczego FcRn, będzie określony standardowymi metodami, na przykład metodami omówionymi powyżej. [0187] Tak uzyskane transgeniczne króliki, wykazujące nadmierną ekspresję króliczego łańcucha alfa FcRn, można korzystnie wykorzystywać do zwiększonego wytwarzania poliklonalnych surowic odpornościowych. Piśmiennictwo [0188] Abbas A. K. i Lichtman A. H. (03) Cellular and molecular immunology. WB Saunders Co, Philadelphia. 2 Adamski F. M., King A. T. i Demmer J. (00) Expression of the Fc receptor in the mammary gland during lactation in the marsupial Trichosurus vulpecula (brushtail possum). Mol Immunol 37, Ahouse J. J., Hagerman C. L., Mittal P., Gilbert D. J., Copeland N. G., Jerkins N. A. i Simister N. E. (1993)
113 114 Mouse MHC class I-like Fc receptor encoded outside the MHC. J Immunol 11, Akilesh S., Christianson G. J., Roopenian D. C. i Shaw A. S. (07) Neonatal FcR Expression in Bone Marrow- Derived Cells Functions to Protect Serum IgG from Catabolism. J Immunol 179, (Andersen J. T., Dee Qian J. i Sandlie I. (06) The conserved histidine 166 residue of the human neonatal Fc receptor heavy chain is critical for the phdependent binding to albumin. Eur J Immunol 36, Andersen S. B. i Bjorneboe M. (1964) Gamma Globulin Turnover in Rabbits before and During Hyperimmunization. J Exp Med 119, Anderson C. L., Chaudhury C., Kim J., Bronson C. L., Wani M. A. i Mohanty S. (06) Perspective - FcRn transports albumin: relevance to immunology and medicine. Trends Immunol 27, Antohe F., Radulescu L., Gafencu A., Ghetie V. i Simionescu M. (01) Expression of functionally active FcRn and the differentiated bidirectional transport of IgG in human placental endothelial cells. Hum Immunol 62, Ausubel F. M., Brent R., Kingston R. E., Moore D. D., Seidman J. G., Smith J. A. i Struhl K. (1998) Current protocols in Molecular Biology. John Wiley & Sons., Hoboken, NJ, USA.
114 11 Beers M. H. i Berkow R. (1999) The Merck Manual of Diagnosis and Therapy. Merck Research Laboratories, New York, N.J. Bender B., Bodrogi L., Mayer B., Schneider Z., Zhao Y., Hammarstrom L., Eggen A., Kacskovics I. i Bosze Z. (07) Position independent and copy-number-related expression of the bovine neonatal Fc receptor alphachain in transgenic mice carrying a 2 kb BAC genomic fragment. Transgenic Res 16, Bender B., Bodrogi L., Yaofeng Z., Kacskovics I., Hammarstrom L. i Bosze Z. (04) Generation of bovine FcRn alpha chain BAC transgenic mice. Tissue Antigens 64, Borvak J., Richardson J., Medesan C., Antohe F., Radu C., Simionescu M., Ghetie V. i Ward E. S. (1998) Functional expression of the MHC class 1-related receptor, FcRn, in endothelial cells of mice. Int Immunol, Bovia F., Salmon P., Matthes T., Kvell K., Nguyen T. H., Werner-Favre C., Barnet M., Nagy M., Leuba F., Arrighi J. F., Piguet V., Trono D. i Zubler R. H. (03) Efficient transduction of primary human B lymphocytes and nondividing myeloma B cells with HIV-1- derived lentiviral vectors. Blood 1, Brambell F. W. R. (198) The passive immunity of the young mammal. Biological Reviews 33,
115 116 Brambell F. W. R., Hemmings W. A. i Morris 1. G. (1964) A theoretical model of gammaglobulin catabolism. Nature 3, Butler J. E. (06) Why I agreed to do this. Dev Comp Immunol,1-17. Chaudhury C., Brooks C. L., Carter D. C., Robinson J. M. i Anderson C. L. (06) Albumin binding to FcRn: distinct from the FcRn-IgG interaction. Biochemistry 4, Chaudhury C., Mehnaz S., Robinson J. M., Hayton W. L., Pearl D. K., Roopenian D. C. i Anderson C. L. (03) The Major Histocompatibility Complex-related Fc Receptor for IgG (FcRn) Binds Albumin and Prolongs Its Lifespan. The Journal of Experimental Medicine 197, Cibelli J. B., Stice S. L., Golueke P. J., Kane J. J., Jerry J., Blackwell C., Ponce de Leon F. A. i Robl J. M. (1998) Transgenic bovine chimeric offspring produced from somatic cell-derived stem-like cells. Nat Biotechnol 16, Claypool S. M., Dickinson B. L., Wagner J. S., Johansen F. E., Venu N., Borawski J. A., Lencer W. I. i Blumberg R. S. (04) Bidirectional transepithelial IgG transport by a strongly polarized basolateral membrane Fcgamma-receptor. Mol Biol Cell 1, Cowan P. J., Tsang D., Pedic C. M., Abbott L. R., Shinkel T. A., d'apice A. J. i Pearse M. J. (1998) The
116 117 human ICAM-2 promoter is endothelial cell-specific in vitro and in vivo and contains critical Spl and GATA binding sites. J Biol Chem 273, Dickinson B. L., Badizadegan K., Wu Z., Ahouse J. C., Zhu X., Simister N. E., Blumberg R. S. i Lencer W. I. (1999) Bidirectional FcRn-dependent IgG transport in a polarized human intestinal epithelial cell line. J Clin Invest 4, Dima S., Medesan C., Mota G., Moraru I., Sjoquist J. i Ghetie V. (1983) Effect of protein A and its fragment B on the catabolic and Fc receptor sites of IgG. Eur J Immunol 13, Dobie K. W., Lee M., Fantes J. A., Graham E., Clark A. J., Springbett A., Lathe R. i McClenaghan M. (1996) Variegated transgene expression in mouse mammary gland is determined by the transgene integration locus. Proc Natl Acad Sci U S A 93, Eggen A., Gautier M., Billaut A., Petit E., Hayes H., Laurent P., Urban C., Plister-Genskow M., Eilertsen K. i Bishop M. D. (01) Construction and characterization of a bovine BAC library with four genome-equivalent coverage. Genet Sel Evol 33, Emanuel S. L. i Pestka S. (1993) Amplification of specific gene products from human serum. Genet Anal Tech Appl, Ghetie V., Hubbard J. G., Kim J. K., Tsen M. F., Lee Y. i Ward E. S. (1996) Abnormally short.<erl1m half-lives
117 118 of IgG in beta 2-microglobulin-deficient mice. Eur J Immunol 26, Ghetie V. i Ward E. S. (02) Transcytosis and catabolism of antibody. Immunol Res 2, Giraldo P. i Montoliu L. (01) Size matters: use of YACs, BACs and PACs in transgenic animals. Transgenic Res, Guyton A. C. (1991) Textbook of Medical Physiology. W.B. Saunders Company. 1 Hao Y. H., Yong H. Y., Murphy C. N., Wax D., Samuel M., Rieke A., Lai L., Liu Z., Durtschi D. C., Welbern V. R., Price E. M., McAllister R. M., Turk J. R., Laughlin M. H., Prather R. S. i Rucker E. B. (06) Production of endothelial nitric oxide synthase (enos) overexpressing piglets. Transgenic Res. Harmsen M. M., Van Solt C. B., Fijten H. P. i Van Setten M. C. (0) Prolonged in vivo residence times of Ilama single-domain antibody fragments in pigs by binding to porcine immunoglobulins. Vaccine 23, Hayes H., Petit E., Lemieux N. i Dutrillaux B. (1992) Chromosomal localization of the ovine beta-casein gene by non-isotopic in situ hybridization and R-banding. Cytogenet Cell Genet 61, Hofmann A., Kessler B., Ewerling S., Weppert M., Vogg B., Ludwig H., Stojkovic M., Boelhauve M., Brem G.,
118 119 Wolf E. i Pfeifer A. (03) Efficient transgenesis in farm animals by lentiviral vectors. EMBO Rep 4, Hofmann A., Zakhartchenko V., Weppert M., Sebald H., Wenigerkind H., Brem G., Wolf E. i Pfeifer A. (04) Generation of transgenic cattle by lentiviral gene transfer into oocytes. Biol Reprod 71, Israel E. J., Patel V. K., Taylor S. F., Marshak- Rothstein A. i Simister N. E. (199) Requirement for a beta 2-microglobulin-associated Fc receptor for acquisition of maternal IgG by fetal and neonatal mice. J Immunol 14, Israel E. J., Wilsker D. F., Hayes K. C., Schoenfeld D. i Simister N. E. (1996) Increased clearance of IgG in mice that lack beta 2-microglobulin: possible protective role of FcRn. Immunology 89, Janeway Jr. C. A., Travers P., Walport M. i Shlomchik M. J. (01) Immunobiology: the immune system in health and disease. Garland Publishing, New York, NY. Junghans R. P. (1997) Finally! The Brambell receptor (FcRB). Mediator of transmission of immunity and protection from catabolism for IgG. Immunol Res 16, Junghans R. P. i Anderson C. L. (1996) The protection receptor for IgG catabolism is the beta2-microglobulincontaining neonatal intestinal transport receptor. Proc Natl Acad Sci U S A 93, 12-6.
119 1 Jurka J., Kapitonov V. V., Pavlicek A., Klonowski P., Kohany O. i Walichiewicz J. (0) Repbase Update, a database of eukaryotic repetitive elements. Cytogenet Genome Res 1, Kacskovics I., Kis Z., Mayer B., West A. P., Jr., Tiangco N. E., Tilahun M., Cervenak L., Bjorkman P. J., Goldsby R. A., Szenci O. i Hammarstrom L. (06a) FcRn mediates elongated serum half-life of human IgG in cattle. Int Immunol 18, Kacskovics I., Mayer B., Kis Z., Frenyo L. V., Zhao Y., Muyldermans S. i Hammarstrom L. (06b) Cloning and characterization of the dromedary (Camelus dromedarius) neonatal Fc receptor (drfcrn). Dev Comp Immunol, Kacskovics I., Wu Z., Simister N. E., Frenyo L. V. i Hammarstrom L. (00) Cloning and characterization of the bovine MHC class I-like Fc preceptor. J Immunol 164, Karlsson R., Michaelsson A. i Mattsson L. (1991) Kinetic analysis of monoclonal antibody-antigen interactions with a new biosensor based analytical system. J Immunol Methods 14, Kobayashi N., Suzuki Y., Tsuge T., Okumura K., Ra C. i Tomino Y. (02) FcRn-mediated transcytosis of immunoglobulin G in human renal proximal tubular epithelial cells. Am J Physiol Renal Physiol 282, F38-6.
120 121 Kohler G. i Milstein C. (197) Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 26, Leenaars M. i Hendriksen C. F. (0) Critical steps in the production of polyclonal and monoclonal antibodidies:: evaluation and recommendations. Ilar J 46, Lin Z. i Floros J. (00) Protocol for genomic DNA preparation from fresh or frozen serum for PCR amplification. Biotechniques 29, 460-2,464,466. Lipman N. S., Jackson L. R., Trudel L. J. i Weis-Garcia F. (0) Monoclonal versus polyclonal antibodies: distinguishing characteristics, applications, and information resources. Ilar J 46, Lobo E. D., Hansen R. J. i Balthasar J. P. (04) Antibody pharmacokinetics and pharmacodynamics. J Pharm Sci 93, Lois C., Hong E. J., Pease S., Brown E. J. i Baltimore D. (02) Germline transmission and tissue-specific expression of transgenes delivered by lentiviral vectors. Science 29, Lonberg N. (0) Human antibodies from transgenic animals. Nat Biotechnol 23, Lu W., Zhao Z., Zhao Y., Hammarstrom L. i Li N. (06) Over-expression of the bovine neonatal Fc receptor gene in the mammary gland of transgenic mice. 8th World
121 122 Congress on Genetics Applied to Livestock Production, Belo Horizonte, MG, Brazylia. Lu W., Zhao Z., Zhao Y., Yu S., Zhao Y., Fan B., Kacskovics I., Hammarstrom L. i Li N. (07) Overexpression of the bovine FcRn in the mammary gland results in increased IgG levels in both milk and serum of transgenic mice. Immunology 122, Maletto B. A., Ropolo A. S., Alignani D.O., Liscovsky M. V., Ranocchia R. P., Moron V. G. i Pistoresi- Palencia M. C. (06) Presence of neutrophil-bearing antigen in lymphoid organs of immune mice. Blood 8, Manz R. A., Hauser A. E., Hiepe F. i Radbruch A. (0) Maintenance of serum antibody levels. Annu Rev Immunol 23, Mayer B., Kis Z., Kajan G., Frenyo L. V., Hammarstrom L. i Kacskovics I. (04) The neonatal Fc receptor (FcRn) is expressed in the bovine lung. Vet Immunol Immunopathol 98, Mayer B., Zolnai A., Frenyo L. V., Jancsik V., Szentirmay Z., Hammarstrom L. i Kacskovics I. (02) Redistribution of the sheep neonatal Fc receptor in the mammary gland around the time of parturition in ewes and its localization in the small intestine of neonatal lambs. Immunology 7, McCarthy K. M., Yoong Y. i Simister N. E. (00) Bidirectional transcytosis of IgG by the rat neonatal
122 123 Fc receptor expressed in a rat kidney cell line: a system to study protein transport across epithelia. J Cell Sci 113, McCullough K. C. i Summerfield A. (0) Basic concepts of immune response and defense development. Ilar J 46, McGrew M. J., Sherman A., Ellard F. M., Lillico S. G., Gilhooley H. J., Kingsman A. J., Mitrophanous K. A. i Sang H. (04) Efficient production of germline transgenic chickens using lentiviral vectors. EMBO Rep, Mond J. J., Lees A. i Snapper C. M. (199) T cellindependent antigens type 2. Annu Rev Immunol 13, Ober R. J., Martinez C., Lai X., Zhou J. i Ward E. S. (04) Exocytosis of IgG as mediated by the receptor, FcRn: an analysis at the single-molecule level. Proc Natl Acad Sci U S A 1, Ober R. J., Radu C. G., Ghetie V. i Ward E. S. (01) Differences in promiscuity for antibody-fcrn interactions across species: implications for therapeutic, antibodies. Int Immunol 13, Opsahl M. L., Springbett A., Lathe R., Colman A., McClenaghan M. i Whitelaw C. B. (03) Mono-allelic expression of variegating transgene locus in the mouse. Transgenic Res 12,
123 124 Palmiter R. D., Wilkie T. M., Chen H. Y. i Brinster R. L. (1984) Transmission distortion and mosaicism in an unusual transgenic mouse pedigree. Cell 36, Peterson N. C. (0) Advances in monoclonal antibody technology: genetic engineering of mice, cells, and immunoglobulins. Ilar J 46, Petkova S. B., Akilesh S., Sproule T. J., Christianson G. J., Al Khabbaz H., Brown A. C., Presta L. G., Meng Y. G. i Roopenian D. C. (06) Enhanced half-life of genetically engineered human IgG1 antibodies in a humanized FcRn mouse model: potential application in humorally mediated autoimmune disease. Int Immunol 18, Pfeifer A. (06) Lentiviral transgenesis- a versatile tool for basic research and gene therapy. Curr Gene Ther 6, Pfeifer A., Ikawa M., Dayn Y. i Verma 1. M. (02) Transgenesis by lentiviral vectors: lack of gene silencing in mammalian embryonic stem cells and preimplantation embryos. Proc Natl Acad Sci U S A 99, Popov S., Hubbard J. G., Kim J., Ober B., Ghetie V. i Ward E. S. (1996) The stoichiometry and affinity of the interaction of murine Fc fragments with the MHC class I-related receptor, FcRn. Mol Immunol 33, 21-. Raguz S., Hobbs C., Yague E., Ioannou P. A., Walsh F. S. i Antoniou M. (1998) Muscle-specific locus control
124 12 region activity associated with the human desmin gene. Dev Biol 1, Rodewald R. (1976) ph-dependent binding of immunoglobulins to intestinal cells of the neonatal rat. J Cell Biol 71, Rogel-Gaillard C., Piumi F., Billault A., Bourgeaux N., Save J. C., Urien C., Salmon J. i Chardon P. (01) Construction of a rabbit bacterial artificial chromosome (BAC) library: application to the mapping of the major histocompatibility complex to position 12q.1.1. Mamm Genome 12, 23-. Roopenian D. C. i Akilesh S. (07) FcRn: the neonatal Fc receptor comes of age. Nat Rev Immunol 7, Roopenian D. C., Christianson G. J., Sproule T. J., Brown A. C., Akilesh S., Jung N., Petkova S., Avanessian L., Choi E. Y., Shaffer D. J., Eden P. A. i Anderson C. L. (03) The MHC class I-like IgG receptor controls perinatal IgG transport, IgG homeostasis, and fate of IgG-Fc-coupled drugs. J Immunol 170, Sabiston B. H. i Rose J. E. (1976) Effect of cold exposure on the metabolism of immunoglobulins in rabbits. J Immunol 116, Sachs U. J., Socher l., Braeunlich C. G., Kroll H., Bein G. i Santoso S. (06) A variable number of tandem repeats polymorphism influences the transcriptional activity of the neonatal Fc receptor alpha-chain promoter. Immunology 119,83-9.
125 126 Sandford A. J. i Pare P. D. (1997) Direct PCR of small genomic DNA fragments from serum. Biotechniques 23, Schedl A., Grimes B. i Montoliu L. (1996) YAC transfer by microinjection. Methods Mol Biol 4, Schnulle P. M. i Hurley W. L. (03) Sequence and expression of the FcRn in the porcine mammary gland. Vet Immunol Immunopathol 91, Schunk M. K. i Macallum G. E. (0) Applications and optimization of immunization procedures. Ilar J 46, Simister N. E. i Mostov K. E. (1989) An Fc receptor structurally related to MHC class I antigens. Nature 337, Simister N. E. i Rees A. R. (198) Isolation and characterization of an Fc receptor from neonatal rat small intestine. Eur J Immunol 1, Smit A., Hubley R. i Green P. ( ) RepeatMasker Open-3.0. Spiekermann G. M., Finn P. W., Ward E. S., Dumont J., Dickinson B. L., Blumberg R. S. i Lencer W. I. (02) Receptor-mediated immunoglobulin G transport across mucosal barriers in adult life: functional expression of FcRn in the mammalian lung. J Exp Med 196, Stills H. F., Jr. (0) Adjuvants and antibody production: dispelling the myths associated with
126 127 Freund's complete and other adjuvants. Ilar J 46, Stirling C. M., Charleston B., Takamatsu H., Claypool S., Lencer W., Blumberg R. S. i Wileman T. E. (0) Characterization of the porcine neonatal Fc receptorpotential use for trans-epithelial protein delivery. Immunology 114, Story C. M., Mikulska J. E. i Simister N. E. (1994) A major histocompatibility complex class I-like Fc receptor cloned from human placenta: possible role in transfer of immunoglobulin G from mother to fetus. J Exp Med 180, Sutherland H. G., Kearns M., Morgan H. D., Headley A. P., Morris C., Martin D.I. i Whitelaw E. (00) Reactivation of heritably silenced gene expression in mice. Mamm Genome 11, Tesson L., Heslan J. M., Menoret S. i Anegon I. (02) Rapid and accurate determination of zygosity in transgenic animals by real-time quantitative PCR. Transgenic Res 11, Thorey I., Offner S., Ros F., Schueler N., Siewe B., Niersbach H., Buelow R., van Schooten W. i Platzer J. (06) Expression of a humanized antibody repertoire in transgenic rabbits. In International Congress of Immunogenomics and Immunomics, Budapest, Węgry. Vidarsson G., Stemerding A. M., Stapleton N. M., Spliethoff S. E., Janssen H., Rebers F. E., de Haas M.
127 128 i van de Winkel J. G. (06) FcRn: an IgG receptor on phagocytes with a novel role in phagocytosis. Blood 8, Wakayama T., Perry A. C., Zuccotti M., Johnson K. R. i Yanagimachi R. (1998) Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 394, Whitelaw C. B., Radcliffe P. A., Ritchie W. A., Carlisle A., Ellard F. M., Pena R. N., Rowe J., Clark A. J., King T. J. i Mitrophanous K. A. (04) Efficient generation of transgenic pigs using equine infectious anaemia virus (EIAV) derived vector. FEBS Lett 71, Yoshida M., Kobayashi K., Kuo T. T., Bry L., Glickman J. N., Claypool S. M., Kaser A., Nagaishi T., Higgins D. E., Mizoguchi E., Wakatsuki Y., Roopenian D. C., Mizoguchi A., Lencer W. I. i Blumberg R. S. (06) Neonatal Fc preceptor for IgG regulates mucosal immune responses to luminal bacteria. J Clin Invest 116, Zhao Y., Kacskovics I., Zhao Z. i Hammarstrom L. (03) Presence of the di-leucine motif in the cytoplasmic tail of the pig FcRn alpha chain. Vet Immunol Immunopathol 96, Zhu X., Meng G., Dickinson B. L., Li X., Mizoguchi E., Miao L., Wang Y., Robert C., Wu B., Smith P. D., Lencer W.I. i Blumberg R. S. (01) MHC class I-related neonatal Fc receptor for IgG is functionally expressed
128 129 in monocytes, intestinal macrophages, and dendritic cells. J Immunol 166, Wykaz sekwencji [0189] <1> AGRICULTURAL BIOTECHNOLOGY CENTER EOTVOS LORAND UNIVERSITY <1> TRANSGENICZNE ZWIERZĘ O ZWIĘKSZONEJ ODPOWIEDZI IMMUNOLOGICZNEJ I SPOSÓB JEGO UZYSKIWANIA <1> 3608/LT <160> 8 <170> PatentIn wersja 3.3 <2> 1 <211> 22 <212> DNA <213> Sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec mrna łańcucha alfa bfcrn <400> 1 cagtaccact tcaccgccgt gt <2> 2 <211> 22 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec mrna łańcucha alfa bfcrn <400> 2 cttggagcgc ttcgaggaag ag 22 <2> 3 <211> <212> DNA <213> sztuczna
129 1 <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn <400> 3 cggccacctc tatcacattt <2> 4 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn 1 2 <400> 4 tgcattgacc acacttggtt <2> <211> 2 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec pbac, U <400> acctctttct ccgcacccga catag <2> 6 <211> 2 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn <400> 6 gttcaagtcc aaaggcaggc tatct 2 <2> 7 <211> 2 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn
130 131 <400> 7 cctttaccca cacccactcc ccaca 2 1 <2> 8 <211> 2 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec pbac, U80929, <400> 8 agaagttcgt gccgccgccg tagta 2 <2> 9 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn <400> 9 ctcctttgtc ttgggcactt <2> <211> 21 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn <400> gccgcggatc ccttccctct g 21 <2> 11 <211> 2 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn <400> 11
131 132 aaagtttctc gagagaggca gagac <2> 12 <211> 2 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn <400> 12 tagttacaga gcctggatag gctga 2 <2> 13 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec BAC 128EOH, koniec <400> 13 tttagctgca tcgggatctt <2> 14 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec BAC 128EOH, koniec <400> 14 ggagtgatgg catttggttt <2> 1 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec FLT3LG <400> 1 tcggagatgg agaaactgct
132 133 <2> 16 <211> 19 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec FLT3LG 1 2 <400> 16 ctggacgaag cgaagacag 19 <2> 17 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec LOC39196 <400> 17 agaacgtgcg taccaaaagc <2> 18 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec LOC39196 <400> 18 agcggttgta ctttcggatg <2> 19 <211> 18 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcgrt <400> 19 ccaagtttgc cctgaacg 18 <2> <211>
133 134 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcgrt <400> gtgtgggcag gagtagagga <2> 21 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec LOC273 <400> 21 agtggtcctg ggattgacag <2> 22 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec LOC273 <400> 22 tcactgagtc ccgtatgtgc <2> 23 <211> 18 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec LOC <400> 23 ctacgtgtgc gccgtgac 18 <2> 24 <211> <212> DNA <213> sztuczna
134 13 <2> <223> Syntetyczny primer PCR swoisty wobec LOC11234 <400> 24 aatcagcttc tccacgcact <2> 2 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec LOC <400> 2 gttgttcaca ccagggaacc <2> 26 <211> 22 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec LOC1123 <400> 26 cctttgccat tgtagatgta gc 22 <2> 27 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec BAC 128EOH, koniec 3 <400> 27 agtcgtgtcc gactctttgc <2> 28 <211> <212> DNA <213> sztuczna <2>
135 136 <223> Syntetyczny primer PCR swoisty wobec BAC 128EOH, koniec <400> 28 cagcctgtct ggtgttctga <2> 29 <211> 26 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec mysiej betaaktyny <400> 29 tggctttctg aacttgacaa cattat 26 <2> <211> 21 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec mysiej betaaktyny <400> ttcacctgcc ctgagtgttt c 21 <2> 31 <211> 26 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec mysiej betaaktyny <400> 31 tgaaggtctc aaacatgatc tgtaga 26 <2> 32 <211> 2 <212> DNA <213> sztuczna <2>
136 137 <223> Syntetyczny primer PCR swoisty wobec bfcgrt 1 <400> 32 cacagtcaag agtggcgacg agcac 2 <2> 33 <211> 18 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcgrt <400> 33 gcaccacgca gcggtagt 18 <2> 34 <211> 21 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcgrt <400> 34 ccttctacgc ctggtcatca c 21 <2> 3 <211> 18 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcgrt <400> 3 ggcgacgagc accactac 18 <2> 36 <211> 19 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcgrt <400> 36
137 138 gattcccgga ggtcwcaca 19 <2> 37 <211> 22 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec łańcucha alfa bfcrn mrna <400> 37 cgcagcarta yctgasctac aa <2> 38 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec łańcucha alfa bfcrn mrna <400> 38 ggctccttcc actccaggtt <2> 39 <211> 27 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec promotora bfcrn <400> 39 gggtctagaa caccaagggc ggcatca 27 <2> 40 <211> 29 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec promotora bfcrn
138 139 <400> 40 ggggaattcc ggctcccgtg actggagac 29 <2> 41 <211> 28 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn <400> 41 ggggaattct ggggccgcag agggaagg <2> 42 <211> 33 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec bfcrn <400> 42 gggacgcgtg aggcagatca caggaggaga aat 33 <2> 43 <211> 423 <212> DNA <213> sztuczna <2> <223> Konstruct P2-bFcRn kodujący segment o długości 290 par zasad promotora łańcucha ciężkiego bfcrn i segment o długości 128 par zasad cdna łańcucha ciężkiego bfcrn <400> <2> 44 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec egzonu 4 bfcrn
139 140 <400> 44 gaggcagatc acaggaggag <2> 4 <211> 19 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec NEO <400> 4 ggaattccca gtgaagggc <2> 46 <211> 2 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec NEO <400> 46 cgagcctgag attgtcaagt gtatt 2 <2> 47 <211> 17 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec FcRn <400> 47 gggatgccac tgccctg 17 <2> 48 <211> 164 <212> DNA <213> Oryctolagus cuniculus <400> 48 4 <2> 49 <211> <212> DNA <213> sztuczna
140 141 <2> <223> Syntetyczny primer PCR swoisty wobec rfcrn 1 2 <400> 49 gggactccct ccttctttgt <2> 0 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec rfcrn <400> 0 agcacttcga gagcttccag <2> 1 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec RCN3 <400> 1 gacgccgaga cctacaagaa <2> 2 <211> 18 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec RCN3 <400> 2 catgtgcggg aactcctc 18 <2> 3 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec FCGRT
141 142 <400> 3 ctgaacggtg aggacttcat 1 2 <2> 4 <211> 19 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec FCGRT <400> 4 gctccttcca ctccaggtt 19 <2> <211> 18 <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec RPS11 <400> agatcggcga catcgtca 18 <2> 6 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec RPS <400> 6 tctggaactg cttcttggtg <2> 7 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec RPL13A <400> 7 catgaggtgg gctggaagta
142 143 <2> 8 <211> <212> DNA <213> sztuczna <2> <223> Syntetyczny primer PCR swoisty wobec RPL13A <400> 8 tccggtagtg gatcttagcc Zastrzeżenia patentowe 1. Nieterapeutyczny sposób rozwijania wzmożonej humoralnej odpowiedzi immunologicznej, obejmujący etap zetknięcia transgenicznego (Tg) zwierzęcia innego niż człowiek z antygenem będącym przedmiotem zainteresowania, przy czym to Tg zwierzę inne niż człowiek posiada konstrukt genetyczny, przy czym ten konstrukt genetyczny zapewnia ekspresję sekwencji kwasu nukleinowego, kodującej łańcuch białka FcRn, i przy czym ta sekwencja kwasu nukleinowego, kodująca łańcuch białka FcRn ulega nadmiernej ekspresji, przez co konstrukt genetyczny zapewnia zwiększoną ochronę IgG przed rozpadem i wzmożoną humoralną odpowiedź immunologiczną przeciwko antygenowi w porównaniu z nietransgenicznym zwierzęciem kontrolnym tego samego gatunku. 2. Sposób według zastrz. 1, w którym ten konstrukt genetyczny zapewnia ekspresję sekwencji kwasu
143 144 nukleinowego, kodującej bydlęcy łańcuch białka FcRn lub kodujący łańcuch wykazujący około 60% lub większą tożsamość sekwencji z bydlęcym białkiem FcRn. Agricultural Biotechnology Center Zastępca:
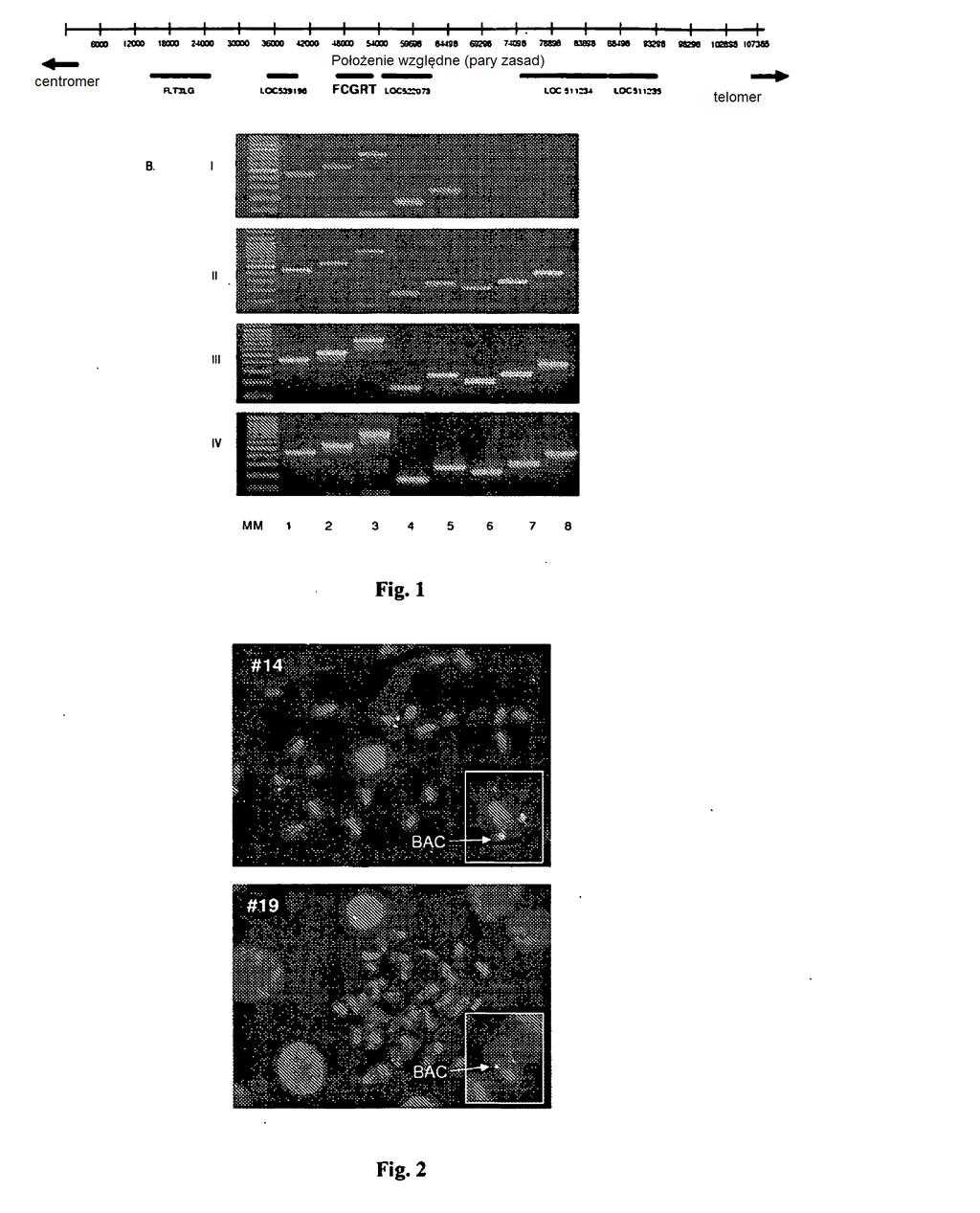
144 14
145 146
146 147
147 148
148 149
149
150 11
151 12
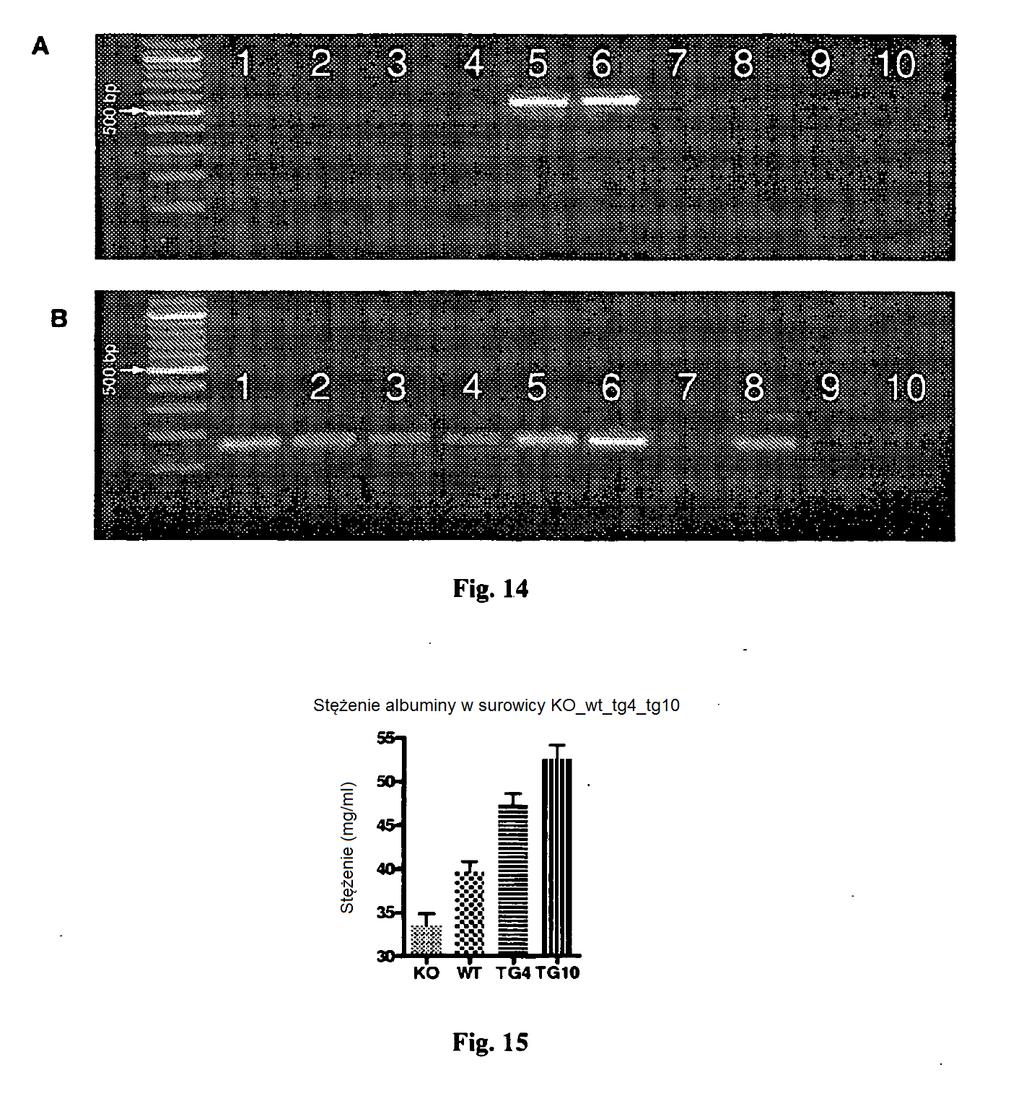
152 13
153 14
PODSTAWY IMMUNOLOGII Komórki i cząsteczki biorące udział w odporności nabytej (cz. III): Aktywacja i funkcje efektorowe limfocytów B
: Aktywacja i funkcje efektorowe limfocytów B") PODSTAWY IMMUNOLOGII Komórki i cząsteczki biorące udział w odporności nabytej (cz. III): Aktywacja i funkcje efektorowe limfocytów B Nadzieja Drela ndrela@biol.uw.edu.pl Konspekt wykładu Rozpoznanie antygenu
PODSTAWY IMMUNOLOGII Komórki i cząsteczki biorące udział w odporności nabytej (cz. III): Aktywacja i funkcje efektorowe limfocytów B Nadzieja Drela ndrela@biol.uw.edu.pl Konspekt wykładu Rozpoznanie antygenu
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE. Ewa Waszkowska ekspert UPRP
 WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
PODSTAWY IMMUNOLOGII Komórki i cząsteczki biorące udział w odporności nabytej (cz.i): wprowadzenie (komórki, receptory, rozwój odporności nabytej)
: wprowadzenie (komórki, receptory, rozwój odporności nabytej)") PODSTAWY IMMUNOLOGII Komórki i cząsteczki biorące udział w odporności nabytej (cz.i): wprowadzenie (komórki, receptory, rozwój odporności nabytej) Nadzieja Drela ndrela@biol.uw.edu.pl Konspekt do wykładu
PODSTAWY IMMUNOLOGII Komórki i cząsteczki biorące udział w odporności nabytej (cz.i): wprowadzenie (komórki, receptory, rozwój odporności nabytej) Nadzieja Drela ndrela@biol.uw.edu.pl Konspekt do wykładu
Nowoczesne systemy ekspresji genów
 Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg
 STRESZCZENIE Przewlekła białaczka limfocytowa (PBL) jest najczęstszą białaczką ludzi starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg kliniczny, zróżnicowane rokowanie. Etiologia
STRESZCZENIE Przewlekła białaczka limfocytowa (PBL) jest najczęstszą białaczką ludzi starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg kliniczny, zróżnicowane rokowanie. Etiologia
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1968711 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 05.01.2007 07712641.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1968711 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 05.01.2007 07712641.5
Dr. habil. Anna Salek International Bio-Consulting 1 Germany
 1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach
 FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach FOCUS Plus to dodatek dostępny dla standardowych pasz tuczowych BioMaru, dostosowany specjalnie do potrzeb ryb narażonych na trudne
FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach FOCUS Plus to dodatek dostępny dla standardowych pasz tuczowych BioMaru, dostosowany specjalnie do potrzeb ryb narażonych na trudne
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
oporność odporność oporność odporność odporność oporność
 oporność odporność odporność nieswoista bierna - niskie ph na powierzchni skóry (mydła!) - enzymy - lizozym, pepsyna, kwas solny żołądka, peptydy o działaniu antybakteryjnym - laktoferyna- przeciwciała
oporność odporność odporność nieswoista bierna - niskie ph na powierzchni skóry (mydła!) - enzymy - lizozym, pepsyna, kwas solny żołądka, peptydy o działaniu antybakteryjnym - laktoferyna- przeciwciała
Białka układu immunologicznego. Układ immunologiczny
 Białka układu immunologicznego Układ immunologiczny 1 Białka nadrodziny immunoglobulin Białka MHC 2 Białka MHC typu I Łańcuch ciężki (alfa) 45 kda Łańcuch lekki (beta 2 ) 12 kda Występują na powierzchni
Białka układu immunologicznego Układ immunologiczny 1 Białka nadrodziny immunoglobulin Białka MHC 2 Białka MHC typu I Łańcuch ciężki (alfa) 45 kda Łańcuch lekki (beta 2 ) 12 kda Występują na powierzchni
Lp. tydzień wykłady seminaria ćwiczenia
 Lp. tydzień wykłady seminaria ćwiczenia 21.02. Wprowadzeniedozag adnieńzwiązanychzi mmunologią, krótka historiaimmunologii, rozwójukładuimmun ologicznego. 19.02. 20.02. Wprowadzenie do zagadnień z immunologii.
Lp. tydzień wykłady seminaria ćwiczenia 21.02. Wprowadzeniedozag adnieńzwiązanychzi mmunologią, krótka historiaimmunologii, rozwójukładuimmun ologicznego. 19.02. 20.02. Wprowadzenie do zagadnień z immunologii.
Klonowanie molekularne Kurs doskonalący. Zakład Geriatrii i Gerontologii CMKP
 Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Odporność nabyta: Nadzieja Drela Wydział Biologii UW, Zakład Immunologii
 Odporność nabyta: Komórki odporności nabytej: fenotyp, funkcje, powstawanie, krążenie w organizmie Cechy odporności nabytej Rozpoznawanie patogenów przez komórki odporności nabytej: receptory dla antygenu
Odporność nabyta: Komórki odporności nabytej: fenotyp, funkcje, powstawanie, krążenie w organizmie Cechy odporności nabytej Rozpoznawanie patogenów przez komórki odporności nabytej: receptory dla antygenu
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Plan. Sztuczne systemy immunologiczne. Podstawowy słownik. Odporność swoista. Architektura systemu naturalnego. Naturalny system immunologiczny
 Sztuczne systemy immunologiczne Plan Naturalny system immunologiczny Systemy oparte na selekcji klonalnej Systemy oparte na modelu sieci idiotypowej 2 Podstawowy słownik Naturalny system immunologiczny
Sztuczne systemy immunologiczne Plan Naturalny system immunologiczny Systemy oparte na selekcji klonalnej Systemy oparte na modelu sieci idiotypowej 2 Podstawowy słownik Naturalny system immunologiczny
Inżynieria genetyczna- 6 ECTS. Inżynieria genetyczna. Podstawowe pojęcia Część II Klonowanie ekspresyjne Od genu do białka
 Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Tolerancja immunologiczna
 Tolerancja immunologiczna autotolerancja, tolerancja na alloantygeny i alergeny dr Katarzyna Bocian Zakład Immunologii kbocian@biol.uw.edu.pl Funkcje układu odpornościowego obrona bakterie alergie wirusy
Tolerancja immunologiczna autotolerancja, tolerancja na alloantygeny i alergeny dr Katarzyna Bocian Zakład Immunologii kbocian@biol.uw.edu.pl Funkcje układu odpornościowego obrona bakterie alergie wirusy
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1747298 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.7 (51) Int. Cl. C22C14/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1747298 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.7 (51) Int. Cl. C22C14/00 (2006.01)
Leczenie biologiczne co to znaczy?
 Leczenie biologiczne co to znaczy? lek med. Anna Bochenek Centrum Badawcze Współczesnej Terapii C B W T 26 Październik 2006 W oparciu o materiały źródłowe edukacyjnego Grantu, prezentowanego na DDW 2006
Leczenie biologiczne co to znaczy? lek med. Anna Bochenek Centrum Badawcze Współczesnej Terapii C B W T 26 Październik 2006 W oparciu o materiały źródłowe edukacyjnego Grantu, prezentowanego na DDW 2006
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii
 Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
LEKI CHEMICZNE A LEKI BIOLOGICZNE
 LEKI CHEMICZNE A LEKI BIOLOGICZNE PRODUKT LECZNICZY - DEFINICJA Art. 2 pkt.32 Ustawy - Prawo farmaceutyczne Substancja lub mieszanina substancji, przedstawiana jako posiadająca właściwości: zapobiegania
LEKI CHEMICZNE A LEKI BIOLOGICZNE PRODUKT LECZNICZY - DEFINICJA Art. 2 pkt.32 Ustawy - Prawo farmaceutyczne Substancja lub mieszanina substancji, przedstawiana jako posiadająca właściwości: zapobiegania
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2152872. (96) Data i numer zgłoszenia patentu europejskiego: 21.05.2008 08748815.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2152872. (96) Data i numer zgłoszenia patentu europejskiego: 21.05.2008 08748815.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 212872 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.0.08 0874881.1 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 212872 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.0.08 0874881.1 (97)
PL 204536 B1. Szczepanik Marian,Kraków,PL Selmaj Krzysztof,Łódź,PL 29.12.2003 BUP 26/03 29.01.2010 WUP 01/10
 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 204536 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 354698 (22) Data zgłoszenia: 24.06.2002 (51) Int.Cl. A61K 38/38 (2006.01)
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 204536 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 354698 (22) Data zgłoszenia: 24.06.2002 (51) Int.Cl. A61K 38/38 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886669 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 02.08.2007 07113670.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886669 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 02.08.2007 07113670.9
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii
 Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
Czy żywność GMO jest bezpieczna?
 Instytut Żywności i Żywienia dr n. med. Lucjan Szponar Czy żywność GMO jest bezpieczna? Warszawa, 21 marca 2005 r. Od ponad połowy ubiegłego wieku, jedną z rozpoznanych tajemnic życia biologicznego wszystkich
Instytut Żywności i Żywienia dr n. med. Lucjan Szponar Czy żywność GMO jest bezpieczna? Warszawa, 21 marca 2005 r. Od ponad połowy ubiegłego wieku, jedną z rozpoznanych tajemnic życia biologicznego wszystkich
Agenda: Wynalazek jako własnośd intelektualna. Co może byd wynalazkiem. Wynalazek biotechnologiczny. Ochrona patentowa
 dr Bartosz Walter Agenda: Wynalazek jako własnośd intelektualna Co może byd wynalazkiem Wynalazek biotechnologiczny Ochrona patentowa Procedura zgłoszenia patentowego Gdzie, kiedy i jak warto patentowad
dr Bartosz Walter Agenda: Wynalazek jako własnośd intelektualna Co może byd wynalazkiem Wynalazek biotechnologiczny Ochrona patentowa Procedura zgłoszenia patentowego Gdzie, kiedy i jak warto patentowad
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ 1. Gen to odcinek DNA odpowiedzialny
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ 1. Gen to odcinek DNA odpowiedzialny
TATA box. Enhancery. CGCG ekson intron ekson intron ekson CZĘŚĆ KODUJĄCA GENU TERMINATOR. Elementy regulatorowe
 Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych
 Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Część praktyczna: Metody pozyskiwania komórek do badań laboratoryjnych cz. I
 Ćwiczenie 1 Część teoretyczna: Budowa i funkcje układu odpornościowego 1. Układ odpornościowy - główne funkcje, typy odpowiedzi immunologicznej, etapy odpowiedzi odpornościowej. 2. Komórki układu immunologicznego.
Ćwiczenie 1 Część teoretyczna: Budowa i funkcje układu odpornościowego 1. Układ odpornościowy - główne funkcje, typy odpowiedzi immunologicznej, etapy odpowiedzi odpornościowej. 2. Komórki układu immunologicznego.
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
Dobierając optymalny program szczepień, jesteśmy w stanie zapobiec chorobom, które mogą być zagrożeniem dla zdrowia Państwa pupila.
 SZCZEPIENIA KOTÓW Działamy według zasady: Lepiej zapobiegać niż leczyć Wychodząc naprzeciw Państwa oczekiwaniom oraz dbając o dobro Waszych pupili opisaliśmy program profilaktyczny chorób zakaźnych psów,
SZCZEPIENIA KOTÓW Działamy według zasady: Lepiej zapobiegać niż leczyć Wychodząc naprzeciw Państwa oczekiwaniom oraz dbając o dobro Waszych pupili opisaliśmy program profilaktyczny chorób zakaźnych psów,
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445326 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.10.2011 11186353.6
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445326 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.10.2011 11186353.6
Geny, a funkcjonowanie organizmu
 Geny, a funkcjonowanie organizmu Wprowadzenie do genów letalnych Geny kodują Białka Kwasy rybonukleinowe 1 Geny Występują zwykle w 2 kopiach Kopia pochodząca od matki Kopia pochodząca od ojca Ekspresji
Geny, a funkcjonowanie organizmu Wprowadzenie do genów letalnych Geny kodują Białka Kwasy rybonukleinowe 1 Geny Występują zwykle w 2 kopiach Kopia pochodząca od matki Kopia pochodząca od ojca Ekspresji
TRANSKRYPCJA - I etap ekspresji genów
 Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
Wyklady IIIL 2016/ :00-16:30 środa Wprowadzenie do immunologii Prof. dr hab. med. ML Kowalski
 III rok Wydział Lekarski Immunologia ogólna z podstawami immunologii klinicznej i alergologii rok akademicki 2016/17 PROGRAM WYKŁADÓW Nr data godzina dzień tygodnia Wyklady IIIL 2016/2017 tytuł Wykladowca
III rok Wydział Lekarski Immunologia ogólna z podstawami immunologii klinicznej i alergologii rok akademicki 2016/17 PROGRAM WYKŁADÓW Nr data godzina dzień tygodnia Wyklady IIIL 2016/2017 tytuł Wykladowca
października 2013: Elementarz biologii molekularnej. Wykład nr 2 BIOINFORMATYKA rok II
 10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
Probiotyki, prebiotyki i synbiotyki w żywieniu zwierząt
 .pl Probiotyki, prebiotyki i synbiotyki w żywieniu zwierząt Autor: dr inż. Barbara Król Data: 2 stycznia 2016 W ostatnich latach obserwuje się wzmożone zainteresowanie probiotykami i prebiotykami zarówno
.pl Probiotyki, prebiotyki i synbiotyki w żywieniu zwierząt Autor: dr inż. Barbara Król Data: 2 stycznia 2016 W ostatnich latach obserwuje się wzmożone zainteresowanie probiotykami i prebiotykami zarówno
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2179743 (96) Data i numer zgłoszenia patentu europejskiego: 13.07.2009 09460028.5 (13) (51) T3 Int.Cl. A61K 38/18 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2179743 (96) Data i numer zgłoszenia patentu europejskiego: 13.07.2009 09460028.5 (13) (51) T3 Int.Cl. A61K 38/18 (2006.01)
Rozkład materiału z biologii dla klasy III AD. 7 godz / tyg rok szkolny 2016/17
 Rozkład materiału z biologii dla klasy III AD zakres rozszerzony LO 7 godz / tyg rok szkolny 2016/17 Biologia na czasie 2 zakres rozszerzony nr dopuszczenia 564/2/2012 Biologia na czasie 3 zakres rozszerzony
Rozkład materiału z biologii dla klasy III AD zakres rozszerzony LO 7 godz / tyg rok szkolny 2016/17 Biologia na czasie 2 zakres rozszerzony nr dopuszczenia 564/2/2012 Biologia na czasie 3 zakres rozszerzony
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1799953 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.08.2005 05770398.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1799953 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.08.2005 05770398.5
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1732433 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.01.2005 05702820.1
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1732433 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.01.2005 05702820.1
Zawartość. Wstęp 1. Historia wirusologii. 2. Klasyfikacja wirusów
 Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 223771 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.08 0886773.1 (13) (1) T3 Int.Cl. A47L 1/42 (06.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 223771 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.08 0886773.1 (13) (1) T3 Int.Cl. A47L 1/42 (06.01) Urząd
Nazwa programu: LECZENIE PIERWOTNYCH NIEDOBORÓW ODPORNOŚCI U DZIECI
 Załącznik nr 12 do zarządzenia Nr 59/2011/DGL Prezesa NFZ z dnia 10 października 2011 roku Nazwa programu: LECZENIE PIERWOTNYCH NIEDOBORÓW ODPORNOŚCI U DZIECI ICD 10 D80 w tym D80.0, D80.1, D80.3, D80.4,
Załącznik nr 12 do zarządzenia Nr 59/2011/DGL Prezesa NFZ z dnia 10 października 2011 roku Nazwa programu: LECZENIE PIERWOTNYCH NIEDOBORÓW ODPORNOŚCI U DZIECI ICD 10 D80 w tym D80.0, D80.1, D80.3, D80.4,
Analizy DNA in silico - czyli czego można szukać i co można znaleźć w sekwencjach nukleotydowych???
 Analizy DNA in silico - czyli czego można szukać i co można znaleźć w sekwencjach nukleotydowych??? Alfabet kwasów nukleinowych jest stosunkowo ubogi!!! Dla sekwencji DNA (RNA) stosuje się zasadniczo*
Analizy DNA in silico - czyli czego można szukać i co można znaleźć w sekwencjach nukleotydowych??? Alfabet kwasów nukleinowych jest stosunkowo ubogi!!! Dla sekwencji DNA (RNA) stosuje się zasadniczo*
STRESZCZENIE PRACY DOKTORSKIEJ
 mgr Bartłomiej Rospond POSZUKIWANIE NEUROBIOLOGICZNEGO MECHANIZMU UZALEŻNIENIA OD POKARMU - WPŁYW CUKRÓW I TŁUSZCZÓW NA EKSPRESJĘ RECEPTORÓW DOPAMINOWYCH D 2 W GRZBIETOWYM PRĄŻKOWIU U SZCZURÓW STRESZCZENIE
mgr Bartłomiej Rospond POSZUKIWANIE NEUROBIOLOGICZNEGO MECHANIZMU UZALEŻNIENIA OD POKARMU - WPŁYW CUKRÓW I TŁUSZCZÓW NA EKSPRESJĘ RECEPTORÓW DOPAMINOWYCH D 2 W GRZBIETOWYM PRĄŻKOWIU U SZCZURÓW STRESZCZENIE
Promotor: prof. dr hab. Katarzyna Bogunia-Kubik Promotor pomocniczy: dr inż. Agnieszka Chrobak
 INSTYTUT IMMUNOLOGII I TERAPII DOŚWIADCZALNEJ IM. LUDWIKA HIRSZFELDA WE WROCŁAWIU POLSKA AKADEMIA NAUK mgr Milena Iwaszko Rola polimorfizmu receptorów z rodziny CD94/NKG2 oraz cząsteczki HLA-E w patogenezie
INSTYTUT IMMUNOLOGII I TERAPII DOŚWIADCZALNEJ IM. LUDWIKA HIRSZFELDA WE WROCŁAWIU POLSKA AKADEMIA NAUK mgr Milena Iwaszko Rola polimorfizmu receptorów z rodziny CD94/NKG2 oraz cząsteczki HLA-E w patogenezie
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2127498 (96) Data i numer zgłoszenia patentu europejskiego: 14.02.2008 08716843.1 (13) (51) T3 Int.Cl. H05B 41/288 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2127498 (96) Data i numer zgłoszenia patentu europejskiego: 14.02.2008 08716843.1 (13) (51) T3 Int.Cl. H05B 41/288 (2006.01)
ROZPORZĄDZENIE KOMISJI (UE) / z dnia r.
 / z dnia r.") KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2259949 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2009 09727379.1 (13) (51) T3 Int.Cl. B60L 11/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2259949 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2009 09727379.1 (13) (51) T3 Int.Cl. B60L 11/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1787644 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 07.11.2006 06123574.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1787644 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 07.11.2006 06123574.3
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 213136 (96) Data i numer zgłoszenia patentu europejskiego: 14.03.2008 08723469.6 (13) (1) T3 Int.Cl. F24D 19/ (2006.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 213136 (96) Data i numer zgłoszenia patentu europejskiego: 14.03.2008 08723469.6 (13) (1) T3 Int.Cl. F24D 19/ (2006.01) Urząd
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2321656 (96) Data i numer zgłoszenia patentu europejskiego:.08.09 09807498.2 (13) (51) T3 Int.Cl. G01R /18 (06.01) G01R 19/
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2321656 (96) Data i numer zgłoszenia patentu europejskiego:.08.09 09807498.2 (13) (51) T3 Int.Cl. G01R /18 (06.01) G01R 19/
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 221611 (96) Data i numer zgłoszenia patentu europejskiego: 19.01. 000481.1 (13) (1) T3 Int.Cl. B28C /42 (06.01) B60P 3/16
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 221611 (96) Data i numer zgłoszenia patentu europejskiego: 19.01. 000481.1 (13) (1) T3 Int.Cl. B28C /42 (06.01) B60P 3/16
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2003466 (96) Data i numer zgłoszenia patentu europejskiego: 12.06.2008 08460024.6 (13) (51) T3 Int.Cl. G01S 5/02 (2010.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2003466 (96) Data i numer zgłoszenia patentu europejskiego: 12.06.2008 08460024.6 (13) (51) T3 Int.Cl. G01S 5/02 (2010.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 161679 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 24.06.0 064.7 (1) Int. Cl. B60R21/01 (06.01) (97) O udzieleniu
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 161679 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 24.06.0 064.7 (1) Int. Cl. B60R21/01 (06.01) (97) O udzieleniu
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1505553. (96) Data i numer zgłoszenia patentu europejskiego: 05.08.2004 04018511.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1505553. (96) Data i numer zgłoszenia patentu europejskiego: 05.08.2004 04018511.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 3 (96) Data i numer zgłoszenia patentu europejskiego: 0.08.04 0401811.8 (13) (1) T3 Int.Cl. G08C 17/00 (06.01) Urząd Patentowy
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 3 (96) Data i numer zgłoszenia patentu europejskiego: 0.08.04 0401811.8 (13) (1) T3 Int.Cl. G08C 17/00 (06.01) Urząd Patentowy
Wykład 14 Biosynteza białek
 BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609. (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609. (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609 (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.2 (13) (51) T3 Int.Cl. A61K 38/17 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609 (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.2 (13) (51) T3 Int.Cl. A61K 38/17 (2006.01)
ZAKŁAD DIAGNOSTYKI LABORATORYJNEJ I IMMUNOLOGII KLINICZNEJ WIEKU ROZOJOWEGO AM W WARSZAWIE.
 ZAKŁAD DIAGNOSTYKI LABORATORYJNEJ I IMMUNOLOGII KLINICZNEJ WIEKU ROZOJOWEGO AKADEMII MEDYCZNEJ W WARSZAWIE. ZAKŁAD DIAGNOSTYKI LABORATOTORYJNEJ WYDZIAŁU NAUKI O ZDROWIU AKADEMII MEDYCZNEJ W WARSZAWIE Przykładowe
ZAKŁAD DIAGNOSTYKI LABORATORYJNEJ I IMMUNOLOGII KLINICZNEJ WIEKU ROZOJOWEGO AKADEMII MEDYCZNEJ W WARSZAWIE. ZAKŁAD DIAGNOSTYKI LABORATOTORYJNEJ WYDZIAŁU NAUKI O ZDROWIU AKADEMII MEDYCZNEJ W WARSZAWIE Przykładowe
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1854925 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2005 05826699.0 (13) (51) T3 Int.Cl. E03D 1/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1854925 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2005 05826699.0 (13) (51) T3 Int.Cl. E03D 1/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1477128 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 14.05.2004 04076445.8 (51) Int. Cl. A61D1/02 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1477128 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 14.05.2004 04076445.8 (51) Int. Cl. A61D1/02 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2074843. (96) Data i numer zgłoszenia patentu europejskiego: 27.09.2007 07818485.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2074843. (96) Data i numer zgłoszenia patentu europejskiego: 27.09.2007 07818485.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 74843 (96) Data i numer zgłoszenia patentu europejskiego: 27.09.07 0781848.0 (13) (1) T3 Int.Cl. H04W 4/12 (09.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 74843 (96) Data i numer zgłoszenia patentu europejskiego: 27.09.07 0781848.0 (13) (1) T3 Int.Cl. H04W 4/12 (09.01) Urząd
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2190940 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.09.2008 08802024.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2190940 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.09.2008 08802024.3
CHOROBY AUTOIMMUNIZACYJNE
 CHOROBY AUTOIMMUNIZACYJNE Autoimmunizacja Odpowiedź immunologiczna skierowana przeciwko własnym antygenom Choroba autoimmunizacyjna Zaburzenie funkcji fizjologicznych organizmu jako konsekwencja autoimmunizacji
CHOROBY AUTOIMMUNIZACYJNE Autoimmunizacja Odpowiedź immunologiczna skierowana przeciwko własnym antygenom Choroba autoimmunizacyjna Zaburzenie funkcji fizjologicznych organizmu jako konsekwencja autoimmunizacji
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1711158 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 16.11.2004 04806793.8
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1711158 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 16.11.2004 04806793.8
Model Marczuka przebiegu infekcji.
 Model Marczuka przebiegu infekcji. Karolina Szymaniuk 27 maja 2013 Karolina Szymaniuk () Model Marczuka przebiegu infekcji. 27 maja 2013 1 / 17 Substrat Związek chemiczny, który ulega przemianie w wyniku
Model Marczuka przebiegu infekcji. Karolina Szymaniuk 27 maja 2013 Karolina Szymaniuk () Model Marczuka przebiegu infekcji. 27 maja 2013 1 / 17 Substrat Związek chemiczny, który ulega przemianie w wyniku
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 21737 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2010 10790844.4 (13) (1) T3 Int.Cl. A47L 1/42 (2006.01) A47L
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 21737 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2010 10790844.4 (13) (1) T3 Int.Cl. A47L 1/42 (2006.01) A47L
Pytania Egzamin magisterski
 Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
mikrosatelitarne, minisatelitarne i polimorfizm liczby kopii
 Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2047071 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.07.2007 07786251.4
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2047071 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.07.2007 07786251.4
Ocena ekspresji genu ABCG2 i białka oporności raka piersi (BCRP) jako potencjalnych czynników prognostycznych w raku jelita grubego
 jako potencjalnych czynników prognostycznych w raku jelita grubego") Aleksandra Sałagacka Ocena ekspresji genu ABCG2 i białka oporności raka piersi (BCRP) jako potencjalnych czynników prognostycznych w raku jelita grubego Pracownia Biologii Molekularnej i Farmakogenomiki
Aleksandra Sałagacka Ocena ekspresji genu ABCG2 i białka oporności raka piersi (BCRP) jako potencjalnych czynników prognostycznych w raku jelita grubego Pracownia Biologii Molekularnej i Farmakogenomiki
SPOSÓB PRZEDSTAWIANIA DOKUMENTACJI DOŁĄCZANEJ DO WNIOSKU O DOPUSZCZENIE DO OBROTU PRODUKTU LECZNICZEGO WETERYNARYJNEGO IMMUNOLOGICZNEGO
 Załącznik nr 2 SPOSÓB PRZEDSTAWIANIA DOKUMENTACJI DOŁĄCZANEJ DO WNIOSKU O DOPUSZCZENIE DO OBROTU PRODUKTU LECZNICZEGO WETERYNARYJNEGO IMMUNOLOGICZNEGO CZĘŚĆ I STRESZCZENIE DOKUMENTACJI I A Wniosek o dopuszczenie
Załącznik nr 2 SPOSÓB PRZEDSTAWIANIA DOKUMENTACJI DOŁĄCZANEJ DO WNIOSKU O DOPUSZCZENIE DO OBROTU PRODUKTU LECZNICZEGO WETERYNARYJNEGO IMMUNOLOGICZNEGO CZĘŚĆ I STRESZCZENIE DOKUMENTACJI I A Wniosek o dopuszczenie
Patentowanie wynalazków z dziedziny biotechnologii
 chroń swoje pomysły 3 Kongres Świata Przemysłu Farmaceutycznego Poznań, 16-17 czerwca 2011 Patentowanie wynalazków z dziedziny biotechnologii dr Izabela Milczarek Czym jest biotechnologia? Biotechnologia,
chroń swoje pomysły 3 Kongres Świata Przemysłu Farmaceutycznego Poznań, 16-17 czerwca 2011 Patentowanie wynalazków z dziedziny biotechnologii dr Izabela Milczarek Czym jest biotechnologia? Biotechnologia,
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680075 (13) T3 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2004
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680075 (13) T3 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2004
Immunologia komórkowa
 Immunologia komórkowa ocena immunofenotypu komórek Mariusz Kaczmarek Immunofenotyp Definicja I Charakterystyczny zbiór antygenów stanowiących elementy różnych struktur komórki, związany z jej różnicowaniem,
Immunologia komórkowa ocena immunofenotypu komórek Mariusz Kaczmarek Immunofenotyp Definicja I Charakterystyczny zbiór antygenów stanowiących elementy różnych struktur komórki, związany z jej różnicowaniem,
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1890471 (96) Data i numer zgłoszenia patentu europejskiego: 19.10.2006 06791271.7 (13) (51) T3 Int.Cl. H04M 3/42 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1890471 (96) Data i numer zgłoszenia patentu europejskiego: 19.10.2006 06791271.7 (13) (51) T3 Int.Cl. H04M 3/42 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1674111 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 08.07.2005 05760156.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1674111 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 08.07.2005 05760156.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 71811 (96) Data i numer zgłoszenia patentu europejskiego: 29.09.06 06791167.7 (13) (1) T3 Int.Cl. H04Q 11/00 (06.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 71811 (96) Data i numer zgłoszenia patentu europejskiego: 29.09.06 06791167.7 (13) (1) T3 Int.Cl. H04Q 11/00 (06.01) Urząd
Rozmnażanie i wzrost komórek sąściśle kontrolowane. Genetyczne podłoże nowotworzenia
 Rozmnażanie i wzrost komórek sąściśle kontrolowane Genetyczne podłoże nowotworzenia Rozmnażanie i wzrost komórek sąściśle kontrolowane Rozmnażanie i wzrost komórek sąściśle kontrolowane Połączenia komórek
Rozmnażanie i wzrost komórek sąściśle kontrolowane Genetyczne podłoże nowotworzenia Rozmnażanie i wzrost komórek sąściśle kontrolowane Rozmnażanie i wzrost komórek sąściśle kontrolowane Połączenia komórek
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1658064 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.6 (51) Int. Cl. A61K31/37 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1658064 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.6 (51) Int. Cl. A61K31/37 (2006.01)
KLONOWANIE DNA REKOMBINACJA DNA WEKTORY
 KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
Warszawa, dnia 3 sierpnia 2016 r. Poz. 1173
 Warszawa, dnia 3 sierpnia 2016 r. Poz. 1173 ROZPORZĄDZENIE MINISTRA ŚRODOWISKA 1) z dnia 18 lipca 2016 r. w sprawie określenia wzorów wniosków oraz zgłoszeń związanych z zamkniętym użyciem mikroorganizmów
Warszawa, dnia 3 sierpnia 2016 r. Poz. 1173 ROZPORZĄDZENIE MINISTRA ŚRODOWISKA 1) z dnia 18 lipca 2016 r. w sprawie określenia wzorów wniosków oraz zgłoszeń związanych z zamkniętym użyciem mikroorganizmów
WYKŁAD: Klasyczny przepływ informacji ( Dogmat) Klasyczny przepływ informacji. Ekspresja genów realizacja informacji zawartej w genach
 Klasyczny przepływ informacji. Ekspresja genów realizacja informacji zawartej w genach") WYKŁAD: Ekspresja genów realizacja informacji zawartej w genach Prof. hab. n. med. Małgorzata Milkiewicz Zakład Biologii Medycznej Klasyczny przepływ informacji ( Dogmat) Białka Retrowirusy Białka Klasyczny
WYKŁAD: Ekspresja genów realizacja informacji zawartej w genach Prof. hab. n. med. Małgorzata Milkiewicz Zakład Biologii Medycznej Klasyczny przepływ informacji ( Dogmat) Białka Retrowirusy Białka Klasyczny
Substancje o Znaczeniu Biologicznym
 Substancje o Znaczeniu Biologicznym Tłuszcze Jadalne są to tłuszcze, które może spożywać człowiek. Stanowią ważny, wysokoenergetyczny składnik diety. Z chemicznego punktu widzenia głównym składnikiem tłuszczów
Substancje o Znaczeniu Biologicznym Tłuszcze Jadalne są to tłuszcze, które może spożywać człowiek. Stanowią ważny, wysokoenergetyczny składnik diety. Z chemicznego punktu widzenia głównym składnikiem tłuszczów
Prezentuje: Magdalena Jasińska
 Prezentuje: Magdalena Jasińska W którym momencie w rozwoju embrionalnym myszy rozpoczyna się endogenna transkrypcja? Hipoteza I: Endogenna transkrypcja rozpoczyna się w embrionach będących w stadium 2-komórkowym
Prezentuje: Magdalena Jasińska W którym momencie w rozwoju embrionalnym myszy rozpoczyna się endogenna transkrypcja? Hipoteza I: Endogenna transkrypcja rozpoczyna się w embrionach będących w stadium 2-komórkowym
Cytokiny jako nośniki informacji
 Wykład 2 15.10.2014 Cytokiny jako nośniki informacji Termin cytokiny (z greckiego: cyto = komórka i kinos = ruch) określa dużą grupę związków o różnym pochodzeniu i budowie, będących peptydami, białkami
Wykład 2 15.10.2014 Cytokiny jako nośniki informacji Termin cytokiny (z greckiego: cyto = komórka i kinos = ruch) określa dużą grupę związków o różnym pochodzeniu i budowie, będących peptydami, białkami
Immulina wzmacnia odporność
 Immulina wzmacnia odporność Narodowe Centrum Badania Preparatów Naturalnych Immulina została opracowana przez zespół naukowców z Narodowego Centrum Badania Preparatów Naturalnych Uniwersytetu Missisipi
Immulina wzmacnia odporność Narodowe Centrum Badania Preparatów Naturalnych Immulina została opracowana przez zespół naukowców z Narodowego Centrum Badania Preparatów Naturalnych Uniwersytetu Missisipi
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 5 Droga od genu do
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 5 Droga od genu do
Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny
 Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny Zadanie 1 1 pkt. za prawidłowe podanie typów dla obydwu zwierząt oznaczonych literami A oraz B. A. ramienionogi, B. mięczaki A.
Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny Zadanie 1 1 pkt. za prawidłowe podanie typów dla obydwu zwierząt oznaczonych literami A oraz B. A. ramienionogi, B. mięczaki A.
PODSTAWY IMMUNOHISTOCHEMII. Determinanty antygenowe (epitopy) Surowice. Antygeny. Otrzymywanie przeciwciał poliklonalnych. poliwalentne monowalentne
 Surowice. Antygeny. Otrzymywanie przeciwciał poliklonalnych. poliwalentne monowalentne") PODSTAWY IMMUNOHISTOCHEMII Antygeny substancje obce dla organizmu, najczęściej o strukturze wielkocząsteczkowej zdolne do wywołania odpowiedzi immunologicznej (tu: produkcji przeciwciał) - IMMUNOGENNOŚĆ
PODSTAWY IMMUNOHISTOCHEMII Antygeny substancje obce dla organizmu, najczęściej o strukturze wielkocząsteczkowej zdolne do wywołania odpowiedzi immunologicznej (tu: produkcji przeciwciał) - IMMUNOGENNOŚĆ
Ocena jakościowa reakcji antygen - przeciwciało. Mariusz Kaczmarek
 Ocena jakościowa reakcji antygen - przeciwciało Mariusz Kaczmarek Antygeny Immunogenność - zdolność do wzbudzenia przeciwko sobie odpowiedzi odpornościowej swoistej; Antygenowość - zdolność do reagowania
Ocena jakościowa reakcji antygen - przeciwciało Mariusz Kaczmarek Antygeny Immunogenność - zdolność do wzbudzenia przeciwko sobie odpowiedzi odpornościowej swoistej; Antygenowość - zdolność do reagowania
Nazwa programu: LECZENIE PIERWOTNYCH NIEDOBORÓW ODPORNOŚCI U DZIECI
 Załącznik nr 11 do Zarządzenia Nr 41/2009 Prezesa NFZ z dnia 15 września 2009 roku Nazwa programu: LECZENIE PIERWOTNYCH NIEDOBORÓW ODPORNOŚCI U DZIECI ICD 10 D80 w tym D80.0, D80.1, D80.3, D80.4, D80.5,
Załącznik nr 11 do Zarządzenia Nr 41/2009 Prezesa NFZ z dnia 15 września 2009 roku Nazwa programu: LECZENIE PIERWOTNYCH NIEDOBORÓW ODPORNOŚCI U DZIECI ICD 10 D80 w tym D80.0, D80.1, D80.3, D80.4, D80.5,
Rośliny modyfikowane genetycznie (GMO)
") Rośliny modyfikowane genetycznie (GMO) Organizmy modyfikowane genetycznie Organizm zmodyfikowany genetycznie (międzynarodowy skrót: GMO Genetically Modified Organizm) to organizm o zmienionych cechach,
Rośliny modyfikowane genetycznie (GMO) Organizmy modyfikowane genetycznie Organizm zmodyfikowany genetycznie (międzynarodowy skrót: GMO Genetically Modified Organizm) to organizm o zmienionych cechach,
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1802536 (96) Data i numer zgłoszenia patentu europejskiego: 20.09.2004 04774954.4 (13) T3 (51) Int. Cl. B65D77/20 B65D85/72
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1802536 (96) Data i numer zgłoszenia patentu europejskiego: 20.09.2004 04774954.4 (13) T3 (51) Int. Cl. B65D77/20 B65D85/72
Czy immunoterapia nowotworów ma racjonalne podłoże? Maciej Siedlar
 Czy immunoterapia nowotworów ma racjonalne podłoże? Maciej Siedlar Zakład Immunologii Klinicznej Katedra Immunologii Klinicznej i Transplantologii Uniwersytet Jagielloński Collegium Medicum, oraz Uniwersytecki
Czy immunoterapia nowotworów ma racjonalne podłoże? Maciej Siedlar Zakład Immunologii Klinicznej Katedra Immunologii Klinicznej i Transplantologii Uniwersytet Jagielloński Collegium Medicum, oraz Uniwersytecki