(21) Numer zgłoszenia: (86) Data i numer zgłoszenia międzynarodowego: , PCT/US94/12401
|
|
|
- Julian Pawlak
- 6 lat temu
- Przeglądów:
Transkrypt
1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) (13) B1 (21) Numer zgłoszenia: Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: (86) Data i numer zgłoszenia międzynarodowego: , PCT/US94/12401 (51 ) IntCl7 C12N 15/86 C12N 15/12 C12N 15/11 C07K 14/82 A61K 48/00 A61P 35/00 (87) Data i numer publikacji zgłoszenia międzynarodowego: , W095/12660, PCT Gazette nr 20/95 (54) Zrekombinowane adenowirusy, kompozycje zawierające zrekombinowane adenowirusy, komórka gospodarza zainfekowana zrekombinowanym adenowirusem oraz sposób otrzymywania biologicznie aktywnego białka p53 typu dzikiego (30) Pierwszeństwo: , US,08/145,826 (73) Uprawniony z patentu: Board of Regents, The Uniwersity of Texas System, Austin, US (43) Zgłoszenie ogłoszono: BUP 18/96 (72) Twórcy wynalazku: Wei-Wei Zhang, Sugar Land, US Jack A. Roth, Houston, US (45) O udzieleniu patentu ogłoszono: WUP 11/03 (74) Pełnomocnik: Łazewska Sławomira, Sławomira Łazewska i Syn PL B1 (57) 1. Zrekombinowany adenowirus wywołujący ekspresję zdolnego do ekspresji białka p53 w ludzkich komórkach nowotworowych, znamienny tym, że zawiera wektor adenowirusowy, do którego wbudowano fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem eukariotycznym. 7. Kompozycja farmaceutyczna w postaci akceptowalnej farmakologicznie zawiesiny, znamienna tym, że zawiera zrekombinowany adenowirus, w którym do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem eukariotycznym. 9. Sposób przywracania funkcji białka p53 typu dzikiego w komórce, w której brak jest białka p53 typu dzikiego, znamienny tym, że komórkę doprowadza się do kontaktu ze zrekombinowanym adenowirusem, w którym do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53, połączony funkcjonalnie z promotorem eukariotycznym po czym prowadzi się w tej komórce ekspresję białka p53 typu dzikiego. Fig. 1

2 Zrekombinowane adenowirusy, kompozycje zawierające zrekombinowane adenowirusy, komórka gospodarza zainfekowana zrekombinowanym adenowirusem oraz sposób otrzymywania biologicznie aktywnego białka p53 typu dzikiego Zastrzeżenia patentowe 1. Zrekombinowany adenowirus wywołujący ekspresję zdolnego do ekspresji białka p53 w ludzkich komórkach nowotworowych, znamienny tym, że zawiera wektor adenowirusowy, do którego wbudowano fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem eukariotycznym. 2. Zrekombinowany adenowirus według zastrz. 1, znamienny tym, że wektor zawiera fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem IE wirusa cytomegalii. 3. Zrekombinowany adenowirus według zastrz. 2, znamienny tym, że wektor zawiera fragment DNA kodujący białko p53, promotor IE wirusa cytomegalii oraz wczesny sygnał poliadenylacji SV Zrekombinowany adenowirus według zastrz. 1, znamienny tym, że z wektora usunięto fragment DNA odpowiedzialny za replikację adenowirusa i w jego miejsce wbudowano fragment DNA kodujący białko p Zrekombinowany adenowirus według zastrz. 1, znamienny tym, że regiony E1 A i E1B wektora adenowirusowego usunięto a na ich miejsce wprowadzono fragment DNA kodujący białko p Zrekombinowany adenowirus według zastrz. 1, znamienny tym, że ma strukturę genomu jak na fig Kompozycja farmaceutyczna w postaci akceptowalnej farmakologicznie zawiesiny, znamienna tym, że zawiera zrekombinowany adenowirus, w którym do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem eukariotycznym. 8. Zrekombinowana komórka gospodarza w której zachodzi ekspresja E1, korzystnie komórka 293, znamienna tym, że jest zainfekowana zrekombinowanym adenowirusem, w którym do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem eukariotycznym. 9. Sposób przywracania funkcji białka p53 typu dzikiego w komórce, w której brak jest białka p53 typu dzikiego, znamienny tym, że komórkę doprowadza się do kontaktu ze zrekombinowanym adenowirusem, w którym do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53, połączony funkcjonalnie z promotorem eukariotycznym po czym prowadzi się w tej komórce ekspresję białka p53 typu dzikiego. 10. Sposób według zastrz. 9, znamienny tym, że komórka jest komórką rakową z mutacją w genie p Farmakologicznie akceptowalna kompozycja do przywracania in vivo oraz in vitro, funkcji białka p53 typu dzikiego w komórce, w której brak białka p53 typu dzikiego, zwłaszcza w ludzkiej komórce raka płuc lub ludzkiej komórce raka piersi, znamienna tym, że zawiera zrekombinowany adenowirus, w którym do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem eukariotycznym. * * * Wynalazek dotyczy dziedziny technologii rekombinacji. W pewnych swych aspektach dotyczy uproszczonych i efektywnych sposobów wytwarzania zrekombinowanych adenowi-

3 rusów. W innych aspektach przedstawione zostały nowe kompozycje i sposoby w których wykorzystywane są konstrukty adenowirusowe zawierające fragment DNA kodujący p53, włącznie ze sposobem otrzymywania biologicznie aktywnego białka p53 typu dzikiego. Znane obecnie metody leczenia raka, włącznie z radioterapią, leczeniem operacyjnym i chemioterapią, mają jak wiadomo ograniczoną efektywność. Rak płuc powoduje co roku śmierć osób w Stanach Zjednoczonych. Według ostatnich danych po wzięciu poprawki na wiek, śmiertelność z powodu raka płuc przewyższyła śmiertelność z powodu raka piersi u kobiet. Pomimo wprowadzenia programów ograniczających palenie, poziom śmiertelności na raka płuc pozostanie wysoki jeszcze w XXI wieku. Racjonalne opracowywanie nowych terapii raka płuc zależeć będzie od zrozumienia biologii raka płuc na poziomie komórkowym. Ustalono już, że wiele odmian raka spowodowanych jest nieprawidłowościami genetycznymi wynikającymi z nadekspresji jednego lub więcej genów, bądź z ekspresji abnormalngo lub zmutowanego genu lub genów. Na przykład ekspresja onkogenów w wielu przypadkach może spowodować rozwinięcie się raka. Onkogeny są to genetycznie zmienione geny, których zmutowany produkt ekspresji w jakiś sposób zakłóca normalne funkcjonowanie lub kontrolę komórkową. (Spandidos i inni, 1989). W przypadku większości onkogenów przebadanych do tej pory, wykryto możliwość ich aktywacji w wyniku mutacji, zazwyczaj mutacji punktowej w obszarze kodującym normalnego genu komórkowego, t.j. proto-onkogenu, co powoduje zastąpienie reszt aminokwasowych w produkcie białkowym otrzymanym w wyniku ekspresji tego genu. Ten zmieniony produkt ekspresji wykazuje anormalne funkcje biologiczne, które biorą udział w procesie neoplastyczym (Travalli i inni, 1990). Mutacje mogą powstać w różny sposób, jak na przykład mutacja w wyniku działania środków chemicznych lub naświetlania jonizującego. Zidentyfikowano już i opisano w różnym stopniu wiele onkogenów i rodzin onkogenów włącznie z ras, myc, neu, raf, erb, src, fm s, jun i abl (Travali i inni., 1990; Bishop 1987). Uważa się, że w czasie normalnego rozwoju komórek, proto-onkogeny powodujące rozwój komórek rakowych są równoważone przez powstrzymujące taki wzrost, geny hamujące rozwój guza. Do zachwiania tej równowagi mogą przyczynić się różne czynniki, co prowadzi do stanu neoplastycznego. Jednym z tych czynników mogą być mutacje w genach powstrzymujących rozwój guza (Weiberg, 1991). Ważnym genem powstrzymującym rozwój guza jest gen kodujący białko komórkowe, p53, fosfoproteinę komórkową, o masie 53 kd, kontrolujące proliferację komórek. Mutacje w genie p53 oraz utrata allelu w chromosomie 17p, gdzie gen ten jest zlokalizowany, są jednymi z najczęstszych zmian wykrywanych w złośliwych zmianach nowotworowych u ludzi. Białko p53 jest w dużym stopniu zachowywane ewolucyjnie i znajduje się w większości zwykłych tkanek. Wykazano, że białko p53 typu dzikiego bierze udział w cyklu komórkowym (Mercer, 1992), regulacji transkrypcji (Fields i inni, 1990 i Mietz i inni 1992), replikacji DNA (Wilcock i Lane 1991 oraz Bargonetti i inni 1991) oraz wywołaniu apoptozy (Yonish- Rouach i inni 1991 oraz Shaw i inni 1992) Znane są różne zmutowane allele p53, w których pojedyncze podstawienia zasad powodują syntezę protein o znacznie różniących się właściwościach regulacji rozwoju i w ostateczności prowadzić może do powstania tkanek złośliwych (Hollstein i inni 1991). Stwierdzono, iż gen p53 jest najczęściej nurtowanym genem w typowych przypadkach raka u ludzi (Hollstein i inni 1991; Zakut-Houri i inni 1985). Udokumentowana jest także nadekspresja p53 w guzach piersi (Casey i inni 1991). Jednym z najtrudniejszych zadań terapii genowej w przypadku raka jest zastosowanie supresorów genów powstrzymujących rozwój guza, jak na przykład p53. Znany jest fakt iż transfekcja p53 typu dzikiego do pewnych odmian komórek raka płuc i piersi może przywrócić kontrolę hamowania rozwoju komórek w liniach komórek (Casey i inni 1991; Takahasi i inni 1992). Pomimo, że transfekcja DNA nie jest skutecznym sposobem wprowadzania DNA do komórek pacjentów, to wyniki te służą wykazaniu, iż wprowadzenie genu p53 typu dzikiego do komórek rakowych o zmutowanym genie p53, może być efektywnym sposobem leczenia, o ile opracowane zostaną ulepszone metody wprowadzania genu p53.

4 Systemy wprowadzania genów możliwe do zastosowania w terapii genowej w celu hamowania rozwoju są opracowywane. Największe zainteresowanie wzbudzają sposoby przenoszenia genu oparte na wirusach z powodu wydajności wirusów w zarażaniu żywych komórek - proces w którym przenoszony jest materiał wirusowy. Dokonał się na tym polu pewien postęp, na przykład w generowaniu wektorów retrowirusowych skonstruowanych tak, aby dostarczały różne geny. Jakkolwiek główne problemy związane są z użyciem wektorów retrowirusowych do terapii genowej, jako że ich wydajność zależy od dostępności receptorów retrowirusowych w komórkach docelowych, są one także trudne do zatężania i oczyszczania oraz wydajnie włączają się one tylko do komórek replikujących się. Ostatnio zaproponowano użycie do niektórych sposobów przenoszenia systemów wektorów adenowirusowych, jednak znane sposoby przygotowania zrekombinowanych adenowirusów mają kilka wad. Sposoby te opierają się na transfekcji wektorami ekspresyjnymi i plazmidami adenowirusowymi komórek docelowych metodą fosforanu wapnia, a następnie przeprowadzaniu analizy łysinkowej, które komórki uległy transfekcji. Te sposoby i próby transfekcji są mało wydajne, i zwykle dają niski poziom namnażania wirusa. Pozostaje więc wyraźna potrzeba opracowania nowych metod wprowadzania do komórek genów, takich jak p53, powstrzymujących rozwój guza, w celu przywrócenia tej funkcji. Korzystne byłoby opracowanie sposobów wytwarzania zrekombinowanych adenowirusów, które nie wymagają transfekcji za pośrednictwem fosforanu wapnia oraz analizy łysinkowej. Adenowirusy ludzkie są to wirusy nowotworowe o podwójnej nici DNA o wielkości genomu około 36 kb (Tooza 1981). Jako modelowy system ekspresji genu u eukariontów, adenowirusy były szeroko badane i dokładnie opisane, co czyni je atrakcyjnymi układami do zastosowania jako systemy przenoszenia genów. Ta grupa wirusów jest łatwa do hodowli i modyfikacji, istnieje także szeroka gama komórek in vivo i in vitro, w których można je umieścić. W komórkach zainfekowanych litycznie adenowirusy mają zdolność blokowania syntezy białek komórkowych, wywoływania syntezy dużych ilości białek wirusowych oraz produkowania kopii wirusa. Genom adenowirusa składa się z kilku regionów. Region El genomu adenowirusa obejmuje E1 A oraz E1B, które kodują białka odpowiedzialne za nadzorowanie transkrypcji genomu wirusowego, jak również kilka genów komórkowych. Ekspresja E2, obejmująca E2A i E2B pozwala na syntezę białek o replikacyjnych funkcjach wirusowych, np. białka wiążącego DNA, polimerazy DNA i końcowego białka, które rozpoczyna replikację. Produkty genu E3 przeciwdziałają cytolizie cytotoksycznych komórek T oraz martwicy tkanek nowotworowych i wydają się ważne w rozprzestrzenianiu wirusa. Funkcje związane z białkami E4 to replikacja DNA, późna ekspresja genu oraz zamykanie komórki. Późnymi produktami genowymi jest większość białek kapsydu wirusa, które ulegają ekspresji jedynie po zakończeniu większości procesów, związanych z pojedynczym pierwszorzędowym transkryptem późnego promotora. Główny późny promotor (MLP) wykazuje wysoką wydajność podczas późnej fazy infekowania (Stratford-Perricaudet i Perricaudet 1991). Jako, że jedynie mała część genomu wirusowego wydaje się potrzebna in cis (Tooza 1981), otrzymane z adenowirusów wektory przedstawiają doskonały materiał do substytucji dużych fragmentów DNA, jeżeli używane są w połączeniu z liniami komórek takimi jak komórki 293. Linia ludzkich komórek embrionalnych nerki transformowana Ad5 (Graham i inni 1977) została opracowana w celu uzyskania koniecznych białek wirusowych in trans. Wynalazcy uznali więc, że cechy adeowirusów czynią je dobrymi kandydatami do stosowania in vivo w rakowych komórkach docelowych (Gninhaus & Horwitz 1992) Zazwyczaj systemy transferu genów przez adenowirusy oparte są na rekombinowanych adenowirusach, otrzymanych w wyniku inżynierii genetycznej, które czyni się niezdolnymi do replikacji, a pozostawia ich zdolność do infekcji. Polega to na delecji fragmentu ich genomu, jak na przykład El. W wyniku wprowadzenia wirusa względnie duże obce białka mogą ulegać ekspresji jeśli z genomu adenowirusowego usunie się dwa dodatkowe fragmenty. Na przykład, adenowirusy mające delecje w obszarach El i E3 są zdolne do przenoszenia do 10 Kb obcego DNA i mogą rosnąć do dużych rozmiarów w komórkach 293 (Stratford-Perricaudet

5 i Perricaudet 1991). Zaskakuje fakt, że po infekcjach andowirusowych może być zauważalna, ciągła ekspresja transgeniczna. Transfer genów za pomocą adenowirusów był ostatnio rozważany jako układ przenoszący geny do komórek eukariontów i do całych zwierząt. Na przykład przy leczeniu myszy z rzadką odmianą recesywnych zmian genetycznych, deskambrylazy ornitynowej (OTC), wykryto, że konstrukcje adenowirusowe mogą być użyte do dostarczenia normalnego enzymu OTC. Niestety, ekspresję normalnych poziomów enzymu OTC osiągnięto jedynie w 4 przypadkach na 17 (Stratford-Perricaudet i inni 1991). Defekt u większości myszy został więc poprawiony jedynie częściowo i nie doprowadził do zmian fizjologicznych ani fenotypowych. Ten rodzaj wyników nie zachęca więc zbytnio do stosowania wektorów adenowirusowych w terapii raka. Próby użycia adenowirusów do przenoszenia genu regulującego zwłóknienie torbielowe przewodnictwa transmembranowego (CFTR) do nabłonka płucnego szczurów również zakończyły się jedynie częściowym sukcesem, pomimo że nie było możliwe oszacowanie aktywności biologicznej przenoszonego genu w nabłonku zwierząt (Rosenfeld i inni 1992). W badaniach tych uzyskano transfer genu i ekspresję białka CFTR w komórkach powietrznych płuc, lecz nie powodowały efektu fizjologicznego. W artykule Rosenfeld i inni, Science 1991, przedstawili ekspresję w komórkach płuc białka α-l-antitrypsynowego, lecz znowu nie stwierdzili efektu fizjologicznego. Oszacowali jednak, że poziom ekspresji zaobserwowany przez nich był jedynie o 2% niższy od poziomu wymaganego do ochrony płuc u ludzi, tj. dużo niższy niż wymagany dla efektu fizjologicznego. Gen ludzkiej α,-antytrypsyny wprowadzono do wątroby zwykłych szczurów poprzez bezpośredni zastrzyk, tam podlegał on ekspresji i wydzielał wprowadzone proteiny ludzkie do plazmy tych szczurów (Jaffe i inni 1992). Jednak poziomy uzyskane były niestety niewystarczające aby przedstawiać wartość terapeutyczną. Wyniki tego rodzaju nie wskazują, że adenowirus zdolny jest kierować ekspresją białka w zrekombinowanych komórkach, w ilości wystarczającej aby uzyskać znaczący fizjologicznie efekt i dlatego wyniki te nie sugerują użyteczności systemu adenowirusowego w terapii raka. Co więcej przed tym wynalazkiem uważano, że p53 nie mógł być włączony do komórki takiej jak używane do przygotowania adenowirusów, jako że byłoby to toksyczne. Ponieważ E1B w adenowirusie wiąże się z p53 uważano to za kolejny powód, dla którego nie można łączyć adenowirusów i technologii p53. Wynalazek dotyczy rozwiązania tych i innych problemów poprzez dostarczenie wydajnych sposobów wytwarzania zrekombinowanych adenowirusów, takich jak konstrukt adenowirusowy zawierający fragment DNA kodujący p53, oraz wydajnych metod przywracania funkcji p53 typu dzikiego komórkom o uszkodzonym p53. Opisano według wynalazku zrekombinowane wektory i wiriony, a także sposoby użycia tych układów do ekspresji p53 w komórkach o funkcjach p53 odbiegających od normy, takich jak komórki rakowe. Opisany także zostanie uproszczony protokół reprodukcji zrekombinowanych adenowirusów wykorzystujący transfekcję DNA przy użyciu liopozomów, a następnie obserwację efektu cytopatycznego (CPE) oraz korzystnie analizę przy pomocy reakcji łańcuchowej polimerazy (PCR). Przedmiotem wynalazku są zrekombinowane adenowirusy wywołujące ekspresję zdolnego do ekspresji białka p53 w ludzkich komórkach nowotworowych, przy czym istotę wynalazku stanowi fakt, że omawiane adenowirusy zawierają wektory adenowirusowe, do których wbudowano fragmenty DNA kodujące białko p53, połączone funkcjonalnie z promotorami eukariotycznymi. W korzystnej odmianie wynalazku wektor zawiera fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem IE wirusa cytomegalii. W jeszcze korzystniejszej odmianie wynalazku wektor zawiera fragment DNA kodujący białko p53, promotor IE wirusa cytomegalii oraz wczesny sygnał poliadenylacji SV40. W innym korzystnym wariancie wynalazku z wektora usunięty jest fragment DNA odpowiedzialny za replikację adenowirusa i w jego miejsce wbudowany jest fragment DNA kodujący białko p53. W jeszcze innym korzystnym wariancie z wektora adenowirusowego usunięte są regiony El A i E1B, a na ich miejsce wprowadzony jest fragment DNA kodujący białko p53.

6 W kolejnym korzystnym wariancie wynalazku zrekombinowany adenowirus ma strukturę genomu jak na fig. 1. Przedmiotem wynalazku jest również kompozycja farmaceutyczna w postaci farmakologicznie akceptowalnej zawiesiny przy czy istotę wynalazku stanowi fakt, że zawiesina zawiera zrekombinowane adenowirusy, w których do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem eukariotycznym. Przedmiotem wynalazku jest również zrekombinowana komórka gospodarza, w której zachodzi ekspresja E1, korzystnie komórka 293, przy czym istotę wynalazku stanowi fakt, że omawiana komórka jest zainfekowana zrekombinowanym adenowirusem, w którym do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem eukariotycznym. Przedmiotem wynalazku jest również sposób przywracania funkcji białka p53 typu dzikiego w komórce, w której brak jest białka p53 typu dzikiego, przy czym istotę wynalazku stanowi fakt, że komórkę doprowadza się do kontaktu ze zrekombinowanym adenowirusem, w którym do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53, połączony funkcjonalnie z promotorem eukariotycznym, przy czym prowadzi się w tej komórce ekspresję białka p53 typu dzikiego. Korzystnie według wynalazku komórka jest komórką rakową z mutacją w genie p53. Przedmiotem wynalazku jest ponadto farmakologicznie akceptowalna kompozycja do przywracania in vivo oraz in vitro, funkcji białka p53 typu dzikiego w komórce, w której brak białka p53 typu dzikiego, zwłaszcza w ludzkiej komórce raka płuc lub ludzkiej komórce raka piersi, przy czym istotę wynalazku stanowi fakt, że omawiana kompozycja zawiera zrekombinowany adenowirus, w którym do wektora adenowirusowego wbudowano fragment DNA kodujący białko p53 połączony funkcjonalnie z promotorem eukariotycznym. W przypadku zrekombinowanego adenowirusa stanowiącego przedmiot wynalazku fragment DNA kodujący p53 w wektorze funkcjonalnie połączony z promotorem eukariotycznym może zawierać sekwencję genomową, przy czym dla ułatwienia uznaje się, że częściej używaną sekwencją będzie sekwencja cdna kodująca białko p53, jako że jest ona dostępniejsza i łatwiejsza w użyciu. Sekwencja DNA kodująca białko p53 w wektorze zwykle zawiera obszar niezbędny do ekspresji genu oraz sygnał poliadenylacji, taki jak wczesny sygnał poliadenylacji SV40 lub sygnał poliadenylacji genu protoaminy. W korzystnej odmianie wynalazku, uważa się za pożądane zlokalizowanie fragmentu DNA kodującego białko p53 połączonego funkcjonalnie z silnym, konstytutywnym promotorem takim jak region CMV, wirusowy LTR, RSV lub SV40 lub połączonego z genami, których ekspresja w komórkach ssaków jest wysoka, np. promotor obszar wydłużania czynnika-1 lub promotor aktynowy. Obecnie najkorzystniejszym promotorem jest promotor IE wirusa cytomegalii (CMV). Fragmenty DNA lub cdna kodujące białko p53 mogą być wprowadzone do zrekombinowanych adenowirusów według wynalazku poprzez wprowadzenie lub dodanie fragmentu DNA (cdna) kodującego białko p53 do genomu wirusowego o brakującym E1B. Jakkolwiek korzystnymi adenowirusami będą wirusy o uszkodzonej replikacji, w których wirusowy gen niezbędny do replikacji i/lub włączenia do genomu, został usunięty z wektorowego konstruktu adenowirusowego, co pozwoliło na wprowadzenie na jego miejsce obszaru p53. Każdy gen poza E1B, zarówno konieczny (np. E1A, E2 i E4), jak i nie wpływający (np. E3) na replikację może być usunięty i zastąpiony fragmentem DNA kodującym p53. Szczególnie korzystne są te wektory, w których regiony El A i E1B wektora adenowirusowego zostały usunięte i wprowadzono w ich miejsce fragment DNA kodujący białko p53, jak przedstawiono na schemacie genomu na fig. 1. Wynalazek nie ogranicza się do pozbawionego E1 wirusa ani do komórek, w których zachodzi ekspresja El. W wynalazku zastosowane mogą być również komplementarne kombinacje wirusów i komórek docelowych o ile wektor p53 nie zawiera E1B. Użyć można wirusów bez funkcjonalnego E2 komórek, w których zachodzi ekspresja E2, jak również wirusów bez E4 i komórek w których zachodzi ekspresja E4 i tym podobne. Jeśli zostanie usunięty lub

7 zastąpiony gen, który nie jest niezbędny do replikacji jak na przykład gen E3, defekt ten nie będzie musiał być równoważony przez komórkę gospodarza. W zastosowaniach według wynalazku, poza wymaganiem aby wektory adenowirusowe nie zawierały E1B, ich budowa nie jest uznawana za istotną. Adenowirusy mogą należeć do każdego z 42 różnych znanych serotypów lub podgrup A-F. W sposobie według wynalazku, adenowirusem stosowanym w celu otrzymania wektora adenowirusowego o brakach replikacyjnych jest typ 5 adenowirusów z podgrupy C. Wybrano ten adenowirus dlatego, że typ 5 jest adenowirusem ludzkim, na temat którego posiadamy dużą wiedzę biochemiczną i genetyczną oraz który był w przeszłości najczęściej używany w konstruktach wektorowych. Wynalazek odnosi się do nowych fragmentów DNA kodujących p53 lub wektorów ekspresyjnych i zrekombinowanych komórek je zawierających. Wszystkie one zawierają wektor adenowirusowy p53 otrzymany zgodnie ze sposobem według wynalazku. Fragmenty DNA według wynalazku zwykle zawierają w kierunku transkrypcji 5'-3', promotor wirusa cytomegalii IE, strukturalny gen ludzkiego p53 typu dzikiego oraz wczesny sygnał poliadenylacji SV40. W przypadku kompozycji farmaceutycznej stanowiącej przedmiot wynalazku zrekombinowane adenowirusy zawieszone są w akceptowalnych farmakologicznie roztworach lub buforach. Korzystne farmakologicznie akceptowalne roztwory zawierają obojętne roztwory soli fizjologicznej zbuforowane fosforanem, mleczanem, Tris-em i podobnymi buforami. Oczywiście pożądane będzie oczyszczenie adenowirusa w takim stopniu aby był on pozbawiony niepotrzebnych składników, takich jak uszkodzone interferujące cząstki adenowirusa lub endotoksyny i inne czynniki gorączkotwórcze, tak aby wirus ten nie powodował negatywnych reakcji w zwierzęciu lub osobie, która przyjmuje konstrukt wektorowy. Korzystnie oczyszczanie odbywa się przy użyciu objętościowego gradientu stężeniowego np. wirowanie w gradiencie chlorku cezu. W przypadku komórki gospodarza stanowiącej przedmiot wynalazku, komórka ta jest komórką eukariotyczną, zwłaszcza komórką ssaka, np. komórką 293, albo może być komórką o defekcie w genie p53, zainfekowaną adenowirusem według wynalazku. Ogólnie wynalazek dotyczy konstruktów wektorowych do wprowadzania genów p53 typu dzikiego, do komórek docelowych, takich jak komórki posiadające zmutowane lub anormalne geny p53, włącznie z komórkami złośliwymi. Te aspekty zawierają otrzymywanie układu ekspresji lub tanskrypcji genu, w którym gen p53 znajduje się pod kontrolą promotora, a jednostka jest włączona do wektora adenowirusowego w zrekombinowanym adenowirusie. Wynalazek jako całość jest niespodziewany i korzystny z wielu powodów. Po pierwsze dotąd przypuszczano, że wirus p53 nie może być wytwarzany w komórkach takich jak używane do przygotowania adenowirusów, jako że byłoby to toksyczne. Po drugie region E1B adenowirusa wiąże się z p53, wpływając w ten sposób na jego funkcje. Po trzecie stwierdzono, że otrzymany adenowirus jest nieoczekiwanie wydajny w powstrzymywaniu powstawania różnych komórek rakowych. Wreszcie po czwarte rakotwórczość komórek rakowych raka płuc zahamowana została przez leczenie Ad5CMV-p53, lecz nie wirusem kontrolnym - co dowodzi, że nowy sposób otrzymywania i dostarczania białka p53 ma zaskakującą skuteczność terapeutyczną. Adenowirusowe konstrukty wektorowe według wynalazku można zastosować do przeniesienia genów powstrzymujących rozwój guza takich jak p53, onkogenów antysensowych i innych pochodnych genów używanych do leczenia raka. Zrekomibnowany adenowirus, który po transfekcji komórek gospodarza, wywołuje ekspresję p53, może być stosowany do zapobiegania powstawaniu złośliwych lub przerzutowych nowotworów. Również może on być użyty do powstrzymywania niekontrolowanego wzrostu komórek o zmutowanym genie p53. Korzystnie adenowirus wywołujący ekspresję p53 hamuje rakotwórczość oraz rozwój komórek H358, lecz może być zastosowana każda inna komórka, która jest wskaźnikiem funkcji p53. Ponadto wektor adenowirusowy, zawierający fragment DNA kodujący p53, po zaszczepieniu w oskrzelach, przeciwdziała rozwojowi ortotopowego nowotworu płuc. Wirus Ad5CMV-p53 dał zachęcające rezultaty w testach na myszach. Po zainfekowaniu wirus powstrzymywał rakotwórczość komórek H358, które zwykle produkują znaczną masę nowo-

8 tworu. Ponadto po zaszczepieniu w oskrzelach, wirus zapobiegał także rozwojowi ortotopowego nowotworu płuc, powstałego w wyniku wprowadzenia do oskrzeli komórek H226r, co potwierdziło skuteczne oddziaływanie Ad5CVM-p53 na komórki raka płuc in vitro. Rakotwórczość komórek raka płuc została zahamowana w wyniku leczenia przy pomocy Ad5CMV-p53, lecz nie wirusa kontrolnego Ad5/RSV/GL2, co wskazuje na skuteczność leczniczą białka p53. Dla osób zaznajomionych ze stanem techniki zrozumiałym będzie, że inne metody wprowadzania wirusów zawarte są w metodzie według wynalazku. Innym aspektem wynalazku jest sposób otrzymywania biologicznie aktywnego białka p53 typu dzikiego, w komórkach i/lub organizmach eukariotycznych, znamienny tym, że komórkę styka się ze zrekombinowanym adenowirusem zawierającym fragment DNA kodujący p53, połączony funkcjonalnie z promotorem eukariotycznym, przy czym zrekombinowany adenowirus wprowadza się w ilości wystarczającej do ekspresji w komórce p53 typu dzikiego. Wektory adenowirusowe według wynalazku znajdą również zastosowanie poza aspektami mającymi bezpośredni związek z terapią genową. Możliwe zastosowania to na przykład analiza in vitro oraz badania mutacji w różnych genach p53 oraz otrzymywanie białek w wyniku rekombinacji, stosowanych na przykład do otrzymywania przeciwciał itp. W zastosowaniach innych niż związane z leczeniem ludzi, włącznie ze wszystkimi związanymi z dalszym definiowaniem aktywności molekularnej p53, inne pochodne wirusy mogą nawet być użyte do dostarczenia p53 do komórki. Stosowne mogą być wirusy należące do rodziny opryszczki np. wirus opryszczki zwykłej (HSV), wirus Epstein-Barr (EBV), wirus cytomegalii (CMV) oraz wirus choroby Aujeszkyego (PRV). Pomimo, że aspekty wynalazku przedstawione są przy pomocy konstruktu p53 stosowanego do przywracania normalnych funkcji komórek w leczeniu raka, to wynalazek może znaleźć zastosowanie w każdej sytuacji, w której pożądany jest wysoki poziom ekspresji białek powstrzymujących rozwój nowotworu w komórce docelowej lub w komórce gospodarza w wyniku użycia zrekombinowanych adenowirusów. Na przykład w zakresie sposobów leczenia raka, konkretnym przykładem, oprócz zastąpienia p53, który jest podkreślony przez wynalazcę, jest wprowadzenie genu nabłoniaka nerwowego z rozetkami prawdziwymi (rb), antysensowego onkogenu (c-myc lub c-ras) oraz innych pokrewnych genów w leczeniu raka. Podkreślić należy, że ponieważ zastosowany wektor adenowirusowy ma uszkodzoną replikacę, nie będzie on zdolny do replikacji w komórkach takich jak komórki rakowe, które zostały zainfekowane. Tak więc, jeśli w niektórych przypadkach istnieje potrzeba stałego leczenia, tak jak na początku terapii, może zaistnieć potrzeba ponownego wprowadzenia wirusa po upływie określonego okresu, na przykład 6 miesięcy lub rok. Rysunki są częścią opisu i są włączone dla lepszego pokazania różnych aspektów wynalazku. Wynalazek może być bardziej zrozumiały przy odwołaniu się do jednego lub więcej z tych rysunków w powiązaniu ze szczegółowym opisem poszczególnych aspektów przedstawionych dalej. Figura 1. Schemat otrzymywania zrekombinowanego adenowirusa p53. Kaseta ekspresyjna p53 wprowadzona została do pxcjl1 w miejsca Xba I i Cla I. Komórki 293 kotransfekowano wektorem ekspresyjnym p53 (pec53) i produktem rekombinacji plazmidu pjm17. Komórki po transfekcji były hodowane na pożywce, aż do wystąpienia efektu cytopatycznego. Identyfikacja nowo otrzymanych zrekombinowanych adenowirusów (Ad5CMV-p53) poprzez analizę DNA metodą PCR, używając matryc DNA przygotowanych z supernatantu komórek CPE wystawionych na działanie proteinazy K i fenolu. Figura 2A przedstawia mapę do analizy strukturalnej DNA Ad5CMV-p53. Mapa genomowego DNA Ad5CMV-p53, z lokalizacją kasety ekspresyjnej p53, starterów PCR i miejsc restrykcyjnych. Wielkość genomu wynosi około 34,4 kb, który podzielono na 10 jednostek mapowych (1 m.u. = 0,35 kb). Kaseta ekspresyjna p53 zastąpiła region El (1,3-9,2 m.u.) genomu Ad5. Starter 1znajduje się w pierwszej wstawce w dół od głównego promotora genu IE ludzkiego CMV. Starter 2 znajduje się w obszarze wczesnego sygnału poliadenylacji. Oba startery o wielkości bp od wstawki cdna p53 na obu końcach definiują produkt PCR o wielkości 1,40 kb. Startery 3 i 4 znajdują się odpowiednio w 1,40 m.u. i 13,4
 i 0,86 kb (Ad5).")
9 m.u. genomu Ad5, co definiuje produkt PCR o wielkości 0,86 kb, specyficzny dla genomu wirusowego. Figura 2B przedstawia obraz analizy produktów PCR w żelu agarozowym. W każdej reakcji użyto dwóch par starterów odpowiadających fragmentom DNA 1,4 kb (p53) i 0,86 kb (Ad5). Matryce DNA użyte w każdej z reakcji stanowi plazmid pec53 (ścieżka 1), DNA Ad5/RSV/GL2 (ścieżka 2), bez DNA (ścieżka 3) oraz DNA Ad4CMV-p53 (ścieżka 4). Ścieżka M zawiera standardy mas cząsteczkowych. Figura 2C przedstawia mapowanie restrykcyjne DNA Ad5CMV-p53. Próbki DNA Ad5CMV-p53 oczyszczone w gradiencie chlorku cezu trawiono odpowiednio: bez enzymu (U), z Hind III(H), Bam III (B), Eco RI (E) i Cla I (C) oraz analizowano w 1% żelu agarozowym. Ścieżka M zawiera standardy masy cząsteczkowej. Figura 3 A, 3B, 3C i 3D, przedstawiają obserwację efektu cytopatycznego w komórkach 293, spowodowanym infekcją zrekombinowanym adenowirusem. Figury 3A, 3B, 3C i 3D przedstawiają serię zdjęć komórek 293 z mikroskopu z kontrastem fazowym (x400), stanowiących kolejne stadiami. Figura 3A, obraz przed transfekcją; fig 3B, negatywna kontrola 12 dni po transfekcji; fig. 3C powstanie CPE 12 dnia po transfekcji; fig. 3D zakończenie CPE 14 dnia po transfekcji. Figura 4A, 4B, 4C i 4D przedstawiają obraz imunohistologiczny komórek zainfekowanych produktami rekombinacji adenowirusów, stanowiących serię obrazów immunohistologicznych komórek H358 w różnych stadiach. Infekcyjność Ad5CMV-p53 w komórkach H358. Komórki H358 zostały zainfekowane Ad5CMV-p53 lub Ad5/RSV/GL2 przy 50 PFU/komórkę przez 24 godziny. Jako próbną infekcję użyto samej pożywki. Zainfekowane komórki znakowano przeciwciałami. Figura 4A jest infekcją próbną sondowaną przeciwciałem anty-p53. Figura 4B przedstawia komórki zainfekowane wirusem kontrolnym Ad4/RSV/GL2 sondowane przeciwciałem anty-p53. Figura 4C to komórki zainfekowane Ad5CMV-p53 sondowane niespokrewnionym przeciwciałem (MOPC-21). Figura 4D to komórki zainfekowane Ad5CMV-p53 sondowane przeciwciałem anty-p53. Jako przeciwciało anty-p53 użyto Pab 1801, a do oznaczania użyto metody awidyno-biotynowej. Figura 5A przedstawia obraz żelu SDS-PAGE wybarwionego błękitem Coomassie umożliwiającym porównanie odpowiadających poziomów ekspresji egzogennych p53 w komórkach H358. Próbki komórek H358 zainfekowane Ad5CMV-p53 lub Ad4/RSV/GL2 przy 30 PFU/komórkę przygotowano 24 i 72 godziny po zainfekowaniu. Żel otrzymany metodą SDS-PAGE, wybarwiony błękitem Coomassie przedstawia względne ilości białek w próbkach. Ścieżki 1 i 4 zawierają próbki komórek zainfekowanych Ad4/RSV/GL2. Ścieżki 2 i 3 zawierają próbki komórek zainfekowanych dwoma pojedynczymi Ad5CMV-p53 w 24 po infekcji, ścieżki 5 i 6 zawierają próbki komórek zainfekowanych Ad5CMV-p53 zebrane 72 godz. po zainfekowaniu. Ścieżka 7 zawiera próbki zainfekowane H godz. po infekcji. Ścieżka M zawiera standardy masy cząsteczkowej w kda (GIBCO-BRL). Figura 5B przedstawia analizę Western blot żelu identycznego z otrzymanym w analizie SDS-PAGE na fig. 5A. Względne poziomy ekspresji p53 zanalizowano metodą Western blot przy użyciu anty-p53. Pierwszorzędowymi przeciwciałami były monoklonowe przeciwciała przeciw białku p53 (Pab 1801, Oncogene Science Inc.) i β-aktynie (Amersham Inc.). Sprzężone z HRP drugorzędowe przeciwciało i ECL pochodziły z Amershem Inc., komórek H358 zainfekowanych wirusem z analizy Western Blotting. Western blot na fig. 5B został wykonany na równoważnej aparaturze i w podobnej skali jak żel przedstawiony na fig. 5 A. Figura 6 przedstawia zależność od czasu ekspresji p53 określoną metodą Western Blotting. Płytki titracyjne z komórkami H358 zainfekowano Ad5CMV-p53 przy 10 PFU/komórkę. Lizaty komórkowe przygotowano w oznaczonych momentach po zainfekowaniu. Western blot znakowano równocześnie przeciwciałami anty-p53 i antyaktynie. Ścieżki oznaczone C reprezentują kontrolę negatywną. Histogram przedstawia względne ilości p53 oznaczone densytometrycznie. Figura7A przedstawia krzywą wzrostu zainfekowanych wirusem ludzkich komórek raka płuc z linii komórek H358. Komórki wysiano w ilości 105 na szalkę (60 mm), 6 szalek na każdą linię komórkową. Po 24 godzinach, komórki były infekowane Ad5CMV-p53 lub Ad5/RSV/GL2 przy 10 m. o. i. (z ang. multiplicity of infection, czyli wielokrotność infekcji,
 i 6 szalek na linię komórkową.")
10 to jest PFU/komórkę). Po zainfekowaniu komórki zliczano codziennie przez 6 dni. Krzywa wzrostu przedstawia wyniki uzyskane w czterech oddzielnych badaniach. Figura 7B przedstawia krzywą wzrostu ludzkich komórek raka płuc z linii komórek H322 zainfekowanych wirusem. Komórki wysiano w ilości 105 komórek na szalkę (60 mm) i 6 szalek na linię komórkową. Po 24 godzinach komórki infekowano Ad5CMV-p53 lub Ad5/RSV/GL2 przy 10 PFU/komórkę. Po zainfekowaniu komórki zliczano codziennie przez 6 dni. Krzywe wzrostu prezentują dane uzyskane z 4 oddzielnych badań. Figura 7C przedstawia krzywą wzrostu ludzkich komórek raka płuc z linii komórek H460 zainfekowanych wirusem. Komórki wysiano w ilości 105komórek na szalkę (60 mm) i 6 szalek na linię komórkową. Po 24 godzinach komórki infekowano Ad5CMV-p53 lub Ad5/RSV/GL2 przy 10 PFU/komórkę. Po zainfekowaniu komórki zliczano codziennie przez 6 dni. Krzywe wzrostu prezentują dane uzyskane z 4 oddzielnych badań. Figura 8 przedstawia schemat badania Ad5CMV-p53 na modelu ortotopicznego raka płuc. Dawki i program leczenia myszy, którym wszczepiono komórki H226Br i wirusy podsumowano na schemacie. Figura 9A, 9B, 9C i 9D są obrazami próbek z rozciętych płuc i śródpiersia leczonych i kontrolnych myszy. Myszy zabito pod koniec 6 tygodniowego okresu kuracji. Tkanki płuc i śródpiersia wycięto dla oceny rozwoju nowotworu. Figura 9A przedstawia śródpiersie zdrowej myszy; fig. 9B przedstawia próbkę śródpiersia z myszy leczonej zaróbką (PBS); fig. 9C przedstawia śródpiersie z myszy leczonej Ad5CMV-p53; fig. 9D przedstawia śródpiersie myszy leczonej Ad5/RSV/GL2. Strzałki pokazują narosłe guzy. Wynalazcy w swoich badaniach zidentyfikowali istotne zjawiska komórkowe, które prowadzą do powstania i rozwoju raka. To umożliwiło otrzymanie nowych sposobów przywracania określonych, normalnych funkcji białka, tak aby powstrzymać in vivo rozwój złośliwego fenotypu. Najczęstsze histologie raka płuc (80%) określane są mianem raka małokomórkowego (SCLC). SCLC może zostać odróżniony od NSCLC dzięki neuroendokrynowym właściwościom komórek. SCLC jest bardzo wrażliwy na chemioterapię i powtarza się szybko po leczeniu. Zgromadzono liczne dowody, że proces złośliwej transformacji następuje za pośrednictwem kodu genetycznego. Główne zmiany chorobowe wykryte w komórkach rakowych występują w dominujących onkogenach i genach powstrzymujących rozwój nowotworu. Dominujące onkogeny mają zmiany w klasie genów zwanych protoonkogenami, które biorą udział w normalnych funkcjach komórki, włącznie z przekazywaniem sygnałów i transkrypcją. Do modyfikacji struktury pierwszorzędowej dominujących onkogenów, które nadają onkogenom zdolność transformowania zalicza się mutacje punktowe, translokacje, przestawienia i powtórzenia. Wydaje się, że geny hamujące rozwój nowotworu potrzebują homozygotycznej utraty funkcji, w wyniku mutacji, delecji lub ich kombinacji aby doszło do transformacji. Niektóre geny hamujące rozwój nowotworu wydają się odgrywać istotną rolę w zarządzaniu proliferacją poprzez regulowanie transkrypcji. Modyfikacja ekspresji dominujących oraz powstrzymujących rozwój nowotworu onkogenów może wpłynąć na pewne cechy komórek, które składają się na złośliwy fenotyp. Pomimo rosnącej wiedzy na temat mechanizmów prowadzących do transformacji w wyniku działania onkogenów, niewielki postęp dokonał się w rozwoju metod leczniczych wykorzystujących onkogeny i ich produkty. Początkowo badania w tej dziedzinie skupiały się na onkogenach dominujących, jako że zostały one scharakteryzowane jako pierwsze. Badania prowadzone nad transferem genów z DNA wykazały uzyskiwanie złośliwego fenotypu przez normalne komórki w wyniku przeniesieniu DNA z komórek złośliwych nowotworów ludzkich. P53 uznawany jest obecnie za gen powstrzymujący rozwój nowotworu (Montenarh 1992). Wysoki poziom p53 stwierdzono w wielu komórkach przekształconych przez chemikalia kancerogenne, naświetlanie ultrafioletem i kilka wirusów włącznie z SV40. Gen p53 jest często unieczynniany w wyniku mutacji, w wielu nowotworach ludzkich i udokumentowano, iż jest najczęściej mutowanym genem w typowych odmianach raka u ludzi (Mercer 1992).

11 W genie tym wykryto mutacje w 50% przypadków ludzkiego NSCLC (Hollestein i inni 1991) i wielu innych nowotworach. Gen p53 koduje fosfoproteinę składającą się z 375 reszt aminokwasowych, która może tworzyć kompleksy z białkami gospodarza, takimi jak duży antygen-t oraz El B. Białko to obecne jest w normalnych tkankach i komórkach, lecz w minimalnym stężeniu w porównaniu do komórek i tkanek przekształconych. Co ciekawe, białko p53 typu dzikiego okazuje się ważne w regulacji wzrostu i podziału komórek. Wykazano, że nadekspresja p53 typu dzikiego w niektórych przypadkach ma skutki przeciwdziałające rozrostowi w ludzkich liniach komórkowych nowotworu. Tak więc p53 może działać po wstrzymująco do rozwoju komórek (Weinberg 1991) i może bezpośrednio powstrzymywać niekontrolowany wzrost komórki. Z czego wynika, że brak lub dezaktywacja p53 typu dzikiego może być niezbędna do całkowitej ekspresji transformowanego potencjału genu. Pomimo, że p53 typu dzikiego uznawany jest za istotny regulator wzrostu w wielu rodzajach komórek, jego charakterystyka genetyczna i biochemiczna wydają się posiadać także znaczenie. Mutacje mis-sens często występują w genie p53 i są konieczne do uzyskania zdolności transformacyjnej onkogenu. Pojedyncza zmiana genetyczna wywołana mutacjami punktowymi może spowodować utworzenie rakotwórczego p53. Inaczej jednak niż w innych onkogenach, mutacje punktowe w p53 zachodzą co najmniej w 30 różnych kodonach, zwykle tworząc dominujące allele, powodujące przesunięcia w fenotypie komórek bez obniżenia homzygotyczności. Dodatkowo wiele z tych dominujących, negatywnych alleli wydaje się tolerowanych w organizmie i przekazywanych w linii zarodkowej. Występuje zróżnicowanie zmutowanych aleli od minimalnie dysfunkcjonalnych do znacznie zmienionych negatywnych alleli dominujących (Weinberg 1991). Casey i jego współpracownicy stwierdzili, że transfekcja kodującego DNA p53 typu dzikiego do komórek dwóch ludzkich linii raka piersi, przywraca w tych komórkach kontrolę powstrzymywania wzrostu (Casey i inni 1991). Podobny efekt wykazano także w przypadku transfekcji, nie zmutowanego, p53 typu dzikiego, do komórek ludzkich linii raka płuc (Takahasi i inni 1992). Wydaje się, że po transfekcji do komórek mających zmutowany gen, p53 dominuje nad zmutowanym genem i przeciwdziała rozrostowi tych komórek. Normalna ekspresja p53 po transfekcji nie wpływa na rozwój komórek z endogennym p53. W związku z tym takie konstrukty mogą być pobrane przez normalne komórki bez efektów ubocznych. Możliwe jest zatem, że leczenie związanych z p53 nowotworów przy użyciu p53 typu dzikiego może zmniejszyć liczbę komórek nowotworowych. Pomimo to badania takie jak opisane powyżej dalekie są od osiągnięcia celu, nie tylko z powodu niemożności użycia transfekcji DNA do wprowadzenia DNA do komórek rakowych wewnątrz ciała pacjenta. Istnieje wiele proponowanych podejść eksperymentalnych do terapii genowej, jednak każde posiada swoje szczególne wady (Mulligan 1993). Jak wspomniano wcześniej istnieją podstawowe metody transfekcji, za pomocą których DNA zawierający interesujący nas gen wprowadzany jest do komórek w sposób niebiologiczny, na przykład poprzez zwiększanie przepuszczalności błony komórkowej w wyniku działania fizycznego lub chemicznego. Naturalnie podejście to ogranicza się do komórek, które mogą być usunięte z ciała na pewien czas oraz tolerują cytotoksyczność leczenia tj. limfocytów. Do transfekcji można zastosować liposomy lub koniugaty białkowe utworzone z określonych lipidów i amfipatycznych peptydów, lecz wydajność integracji genów pozostaje bardzo niska - w granicach jednego przyłączenia na 1000 do komórek, a ekspresja genów po transfekcji jest często ograniczona do dni w wypadku komórek proliferujących i tygodni w przypadku komórek nie proliferujących. Transfekcja DNA nie jest więc najwyraźniej właściwą metodą leczenia raka. Drugie podejście wykorzystuje naturalne zdolności wirusów do wnikania z ich własnym materiałem genetycznym do komórek. Retrowirusy są obiecującymi nośnikami genów, ze względu na zdolność retrowirusów do wprowadzania swoich genów do genomu gospodarza, przenoszenia dużej ilości obcego materiału genetycznego, zarażania wielu różnych gatunków i typów komórek oraz łatwość w umieszczaniu dużych ich ilości w określanych liniach komórek. Jednak trzy główne problemy podważają możliwość użycia wektorów retrowirusowych. Po pierwsze efektywność retrowirusowa zależy od dostępności receptorów wirusowych na

12 docelowej powierzchni. Po drugie, retrowirusy włączają się skutecznie jedynie do komórek ulegających replikacji. Po trzecie retrowirusy trudno jest zatężyć i oczyścić. Wynalazek przedstawia genową terapię raka przy użyciu nowego i bardziej efektywnego wektora powstrzymującego rozwój nowotworu. Ten produkt rekombinacji wirusa wykorzystuje zaletę wektorów adenowirusowych, takich jak wysoka pojemność, szeroki zasięg komórek docelowych, wydajna transdukcja oraz nieprzyłączanie się do komórek docelowych. W tym zestawieniu według wynalazku otrzymywany jest niezależny adenowirus o obniżonej zdolności replikacyjnej, który powoduje ekspresję p53 typu dzikiego (Ad5CMV-p53) pod kontrolą promotora wirusa cytomegalii. Szczególne zalety sytemu adenowirusowego dla dostarczania obcych białek do komórki są to: zdolność do substytucji względnie dużych fragmentów DNA wirusowego poprzez obce DNA, stabilność strukturalna zrekombinowanych adenowirusów, bezpieczeństwo podawania adenowirusów ludziom, brak znanego powiązania infekcji adenowirusowej z rakiem, zdolność otrzymywania dużych miar zrekombinowanych wirusów, wysoka infekcyjność. Kolejną zaletą adenowirusów w stosunku do retro wirusów jest wysoki poziom ekspresji genów. Dodatkowo replikacja adenowirusów jest niezależna od replikacji genu, w którym on się znajduje, w odróżnieniu od sekwencji retorowirusowych. Ponieważ geny transformujące adenowirusy w obszarze E 1 mogą być usunięte i wciąż pozostają efektywnymi wektorami ekspresyjnymi onkogenowe ryzyko uważane jest za pomijalne (Grunhaus & Horwitz 1992). Funkcje kontrolne wektorów ekspresyjnych, jeśli ekspresja ma zachodzić w komórkach ssaków, często pochodzą z wirusów. Na przykład powszechnie używane promotory otrzymywane są z wirusa poliomy, adenowirusa 2 i małpiego wirusa 40 (SV40). Wczesne i późne promotory wirusa SV 40 są szczególnie przydatne, ponieważ oba są łatwe do otrzymania z wirusa jako fragment, który zawiera także wirusowy początek replikacji SV40. Mniejsze i większe fragmenty SV40 mogą być również użyte, pod warunkiem że zawierają się w sekwencji o około 250 par zasad od miejsca cięcia Hindlll do Bgll umieszczonej w wirusowym początku replikacji. Dodatkowo możliwe jest, a często również pożądane, użycie promotora lub sekwencji kontrolującej normalnie związanych z włączoną sekwencją genową o ile takie sekwencje kontrolujące współpracują z komórkami gospodarza. Początek replikacji można zapewnić przez konstrukcję wektora, może zawierać egzogenny początek, taki jak można uzyskać z SV40 lub innych źródeł wirusowych (np. wirus polimy, adenowirus, VSV, BPV) lub może być wykorzystany mechanizm replikacji chromosomowej komórki gospodarza. Jeśli wektor jest włączony do chromosomu komórki wystarczy jedynie mechanizm startu replikacji komórki. Zaprojektowanie i namnażanie korzystnego adenowirusa p53 przedstawia fig. 1. W powiązaniu z tym opracowano ulepszony protokół rozmnażania i identyfikacji zrekombinowanych adenowirusów (dyskutowane poniżej). Po identyfikacji sprawdzono strukturę zrekombinowanego adenowirusa p53 przy użyciu analizy PCR, jak przedstawiono na fig. 2. Po wyizolowaniu i potwierdzeniu jego struktury adenowirus p53 użyto do zainfekowania ludzkiego raka płuc, linii komórkowej H358, która posiada homozygotyczną delecję genu p53. Analiza Western blot wskazywała, że poziom ekspresji egzogennego białka p53 był wysoki (fig. 4 i fig. 5) i najwyższy 3 dnia po infekcji (fig. 6). Wykazano również, że w przypadku mutacji punktowej w p53 linii komórkowej H322, zmutowany p53 podlegało regulacji negatywnej pod wpływem ekspresji egzogennego p53. Jako kontrolę doświadczenia użyto wirionu (Ad5/RSV/GL2) mającego strukturalne podobieństwo do Ad5CMV-p53. Wirion ten zawierał gen lucyferazy CDNA wprowadzony przez promotor LTR wirusa sarkomy Rousa w kasecie ekspresyjnej wirionu. W komórkach zainfekowanych wirionem Ad5/RSV/GL2 nie zauważono ani ekspresji p53, ani zmiany w ekspresji aktyny. Rozwój komórek H358 zainfekowanych Ad5CMV-p53 został znacząco powstrzymany w przeciwieństwie do niezanifekowanych komórek oraz komórek zainfekowanych wirionem kontrolnym (fig. 7A). Rozwój komórek H322 został również powstrzymany w dużym stopniu przez wirion p53 (fig. 7B), podczas gdy wpływ na wzrost komórek H460 płuc ludzkich zawierających p53 typu dzikiego był znacznie mniejszy (fig. 7C).

13 Ad5CMV-p53 pośredniczył w silnym efekcie hamującym rozwój komórek rakowych płuc in vitro. Hamowanie rozwoju nie było tak widoczne jak gdy komórki zainfekowano Ad5CMV-p53 przy wielokrotności infekcji niższej niż 1 PFU/komórkę, podczas kiedy przy wielokrotności infekcji wyższym niż loopfu/komórkę można było zaobserwować cytotoksyczność nawet z kontrolnym wirusem Ad5/RSV/GL2. W naszych badaniach optymalna dawka dla badania stopnia rozwoju wynosiła PFU/komórkę. W tym obszarze dawki, hamowanie rozwoju komórek można było przypisać ulegającej ekspresji proteinie p53. Testy na nagich myszach pokazały, że rakotwórczość komórek leczonych Ad5CMVp53 została w dużym stopniu zahamowana. W opracowanym na myszy, modelu ortotopowego raka płuc u ludzi, rakotwórcze komórki H226Br z mutacją punktową w p53 wszczepiono do oskrzeli na 3 dni przed leczeniem wirusowym. Zakropienie do oskrzeli Ad5CMV-p53 zapobiegło utworzeniu się nowotworu w tym układzie modelowym, co sugeruje, że zmodyfikowane adenowirusy są efektywnym wektorem do pośredniczenia w transferze i ekspresji genów powstrzymujących rozwój nowotworu w komórkach ludzkich oraz że wirus Ad5CMV-p53 może być w ulepszony aby stanowić czynnik terapeutyczny do użycia w terapii genowej. Wywołany Ad5CMV-p53 wysoki poziom ekspresji genu p53 w raku płuc u ludzi wykazano analizą Western blot. Egzogennego białka p53 było około czternastokrotnie więcej niż endogennego p53 typu dzikiego, w komórkach H460 oraz około dwa do czterech razy więcej niż w kontroli wewnętrznej β-aktyny w komórkach H358. Wysoki poziom ekspresji może być przypisany: wysoce efektywnemu transferowi genów, silnemu promotorowi CMV kontrolującemu cdna p53 lub adenowirusowemu enchanserowi E 1 wzmacniającemu transkrypcję cdna p53. Czas trwania ekspresji p53 po infekcji wyniósł ponad 15 dni w komórkach H358. Jednak nastąpił nagły spadek w poziomie ekspresji po pierwszych 5 dniach. Analizy PCR próbek DNA z zainfekowanych komórek H358 pokazały spadek poziomu wirusowego DNA wraz ze zmniejszającym się poziomem białka, co wskazywało na utratę wirusowego DNA podczas ciągłego rozwoju in vitro komórek rakowych. Spadek ekspresji p53 mógł także być rezultatem osłabienia komórkowego promotora CMV, który kontroluje ekspresję p53, ponieważ wcześniej zarejestrowano zjawisko wyłączania promotora CMV za pośrednictwem komórki, w której on się znajduje (Dai i inni 1992). Wektory adenowirusowe są nieintegrowalnymi wektorami przenoszącymi geny, więc czas trwania ekspresji genów zależy od kilku czynników: komórek w których proces się odbywa, przenoszonych genów i odpowiedniego promotora. Crystal i współpracownicy pokazali ekspresję na niskim poziomie genu regulującego przewodnictwo transmembranowe zwłóknienia torbielowatego w komórkach nabłonkowych szczurów, która była wykrywalna 6 tygodni po zainfekowaniu (Rosenfeld i inni 1992). Laboratoriu Perricaudeta zademonstrowało, iż minimalna ekspresja genu minidystrofiny w mięśniu myszy mdx może trwać dłużej niż 3 miesiące po zainfekowaniu. Krótkoterminowa ekspresja na wysokim poziomie białka p53 typu dzikiego stwierdzona w niniejszym wynalazku może mieć korzystny wpływ na normalne komórki w wyniku leczenia in vivo przy użyciu Ad5CMV-p53. Badania opisane wykazują, że zrekombinowane adenowirusy posiadają właściwości powstrzymujące rozwój nowotworu, co wydaje się funkcjonować poprzez przywracanie funkcji p53 w komórkach nowotworowych. Te rezultaty popierają użycie wirionów Ad5CMV-p53 jako środka leczniczego w leczeniu raka. Zrekombinowane adenowirusy jako nowe systemy dostarczania genów mają wiele potencjalnych zastosowań w terapii genowej i otrzymywaniu szczepionek. Reprodukcja rekombinowanych adenowirusów jest w związku z tym istotnym narzędziem molekularnym. Istniejące metody namnażania rekombinowanych adenowirusów wykorzystują transfekcję do komórek 293 za pośrednictwem fosforanu wapnia, a następnie próby łysinkowe po transfekcji. Zarówno wydajność transfekcji związana z tą metodą powinna być poprawiona, jak i sama procedura uproszczona. W znanym stanie techniki, rozmnażanie zrekombinowanych adenowirusów prowadzono zazwyczaj w wyniku transfekcji z fosforanem wapnia. Ta procedura polega na poddawaniu komórek działaniu przez kilka godzin, wektora lub plazmidu DNA w fosforanie wapnia, a następnie stosowaniu krótkiego wstrząsu np. minuta w 15% glicerolu. Ta metoda posiada

14 wiele istotnych wad co powoduje, że niewielka ilość DNA jest włączana do komórki. Jest to bardzo mało wydajna metoda transfekcji. Obecność wirusów wykrywana była także zazwyczaj dzięki łysinkom widocznym jako wyraźne okrągłe obszary wokół rozpadających się komórek, co wskazuje na lizę komórek spowodowaną replikacją wirusa. Wynalazcy opracowali nową procedurę otrzymywania adenowirusów, która w znacznym stopniu poprawia wydajność transfekcji a także upraszcza selekcję. Niespodziewanie odkryto, że kombinacja transfekcji za pośrednictwem liposomów, np DOTAP, z obserwacją efektu cytopatycznego (CPE) prowadzi zarówno do ulepszenia wydajności jak i szybkiej i uproszczonej detekcji. W nowej procedurze transfer genów za pośrednictwem lipozomu DOTAP stosuje się do przeprowadzenia transdukcji wektora ekspresyjnego i zrekombinowanego plazmidu do komórek 293. Komórki po transfekcji pozostają w pożywce MEM w celu obserwacji efektu cytopatycznego (CPE). W dwóch badaniach przy użyciu tej nowej metody 2 dołki na płytce o 24 dołkach i 3 płytki na pięć w sześćdziesięciomilimetrowych płytek wykazywały CPE odpowiednio w 10 i 12 dniu po kotransfekcji. Tymczasem przy użyciu metody transfekcji z fosforanu wapnia nie uzyskano żadnego zrekombinowanego wirusa w trzech próbach, po kotransfekcji z dwudziestoma 60 mm płytkami w każdym eksperymencie. Używając prób CAT w liniach komórkowych Hep G2 i HeLa, transfekcja za pośrednictwem DOTAP dała pięciokrotnie wyższą aktywność CAT niż transfekcja z fosforanem wapnia. Wyeliminowanie potrzeby obłożenia agarozą po kotransfekcji również uprościło procedurę. Końcowy punkt badań - replikacja wirusa staje się znacznie prostsza i jasna przy bezpośredniej obserwacji CPE zamiast identyfikacji łysinek, co zazwyczaj jest niejasne i trudne do zdeterminowania po dniach inkubacji. Figura 3 przedstawia kulturę komórek z CPE w porównaniu z kulturą bez CPE. Opracowano również szybką technikę określania miana przy użyciu PCR. Bezpośrednia analiza PCR próbek DNA z nadsączu kultur komórek z CPE zazwyczaj wykorzystuje dwie pary starterów - jedną do amplifikacji sekwencji specyficznych dla wstawek, a drugą do amplifikacji sekwencji specyficznych dla genomów wirusowych. Wynalazcy pokazali, że adenowirusy wprowadzone do pożywki stosowanej w hodowli komórek są wykrywalne przez PCR, pozwalając na użycie nawet do 50 μ nadsączu do przygotowania matryc DNA. Identyfikacja sekwencji DNA specyficznych dla wstawek i specyficznych dla genomów wirusowych wynikające z amplifikacji PCR może być określona poprzez na przykład analizę produktów PCR po amplifikacji w żelach agarozowych. Prążki odpowiadające DNA specyficznemu dla wstawek i specyficznemu dla genomu wirusowego, a także startery mogą być zidentyfikowane przez porównanie z odpowiednimi standardowymi markerami. Jeśli do amplifikacji produktów genowych specyficznego dla wstawek i genomów wirusowych używa się PCR, należy najpierw przygotować starter specyficzny dla sekwencji, która ma być amplifikowana. Wydajną i selektywną amplifikację uzyskuje się stosując dwie pary starterów: jedną do amplifikacji określonej sekcji produktu specyficznego dla wstawki i innego do odszukania fragmentu produktu specyficznego dla genomu wirusowego. Na przykład kaseta ekspresyjna p53 została skonstruowana z wirusa cytomegalii (CMV) promotora i wczesnego sygnału poliadenylacji SV40. Pierwszy zestaw starterów zawierać będzie starter umieszczony w pierwszym intronie w dół od ludzkiego CMV głównego promotora genu EE, podczas gdy drugi starter pierwszego zestawu starterów znajduje się w obszarze wczesnego sygnału poliadenylacji SV40. Idealnie oba te startery są 15 do 20 par zasad od wstawki cd- NA, którym w tym przykładzie jest cdna p53. Następnie powinien zostać wybrany zdefiniowany produkt PCR - na przykład cdna p53 o 1,40 kb. Przykładowo drugi zestaw starterów może znajdować się w 11 M.U. i 13,4 M.U. genomu Ad5 aby zdefiniować 0,86 specyficzny dla genomu wirusowego produkt PCR. Dobór starterów jest dobrze znany dla osób obeznanych ze stanem techniki. Można konstruować startery do amplifikacji wybranych fragmentów sekwencji DNA, sekwencja par zasad jest znana. Startery hybrydyzują z DNA i działają jako miejsca początku syntezy fragmentu genu. Startery nukleotydowe są opracowywane tak aby wiązały się w oddzielnych miejscach na przeciwległych niciach definiując w ten sposób sekwencje będące przedmiotem

15 amplifikacji. Cząsteczki kwasu nukleinowego mające być użyte jako startery zawierają sekwencję o co najmniej 10 parach zasad, która może być komplementarna do fragmentu DNA poddanego amplifikacji. 10 par zasad zasadniczo uważa się za dolną granicę, ponieważ sekwencja o mniej niż 10 zasadach może nie hybrydyzować wydajnie i prowadzić do braku stabilności kompleksu. Korzystnie stosuje się sekwencje o zasad, korzystnie według wynalazku stosowane są pary starterów pokazane na fig. 1 o 19 lub 20 parach. Wynalazcy uważają, że lokalne dostarczanie adeno wirusowych konstruktów genu p53 do komórek raka płuc u pacjentów z rakiem związanym z p53, np. rakiem wewnątrzoskrzelowym, będzie bardzo wydajną metodą na dostarczanie leczniczo skutecznych genów i przeciwdziałanie klinicznemu rozwojowi choroby. Sugeruje się, że to podejście jest znacznym ulepszeniem w stosunku do obecnych metod leczenia raka, które opierają się na próbach zniszczenia lub usunięcia wszystkich komórek rakowych. Ponieważ uśpienie komórek rakowych jest częstym zjawiskiem, powoduje to, że skuteczne zabicie komórek rakowych jest wysoce nieprawdopodobne. Przewidywane jest, że pobieranie konstruktów adenowirusowych przez komórki NSCLC zmniejszy poziom poliferacji tych komórek. To zwiększyłoby okres czasu, przez który dane płuco mogłoby się rozprężać, przeciwdziałałoby ponownemu rozwojowi uśpionego nowotworu i przedłużyło okres życia pacjenta. Pacjenci z nienadajacym się do wycięcia uśpionym nowotworem nawracającym się, który częściowo lub całkowicie blokuje wlot powietrza i u których nie powiodła się, lub nie możliwa jest radioterapia zewnętrzną wiązką promieniowania, mogą być poddani tej metodzie leczenia. Istniejące metody leczenia w takich stanach oferują jedynie krótkoterminowe łagodzenie objawów. U wielu pacjentów następuje nawrót pomimo radioterapii. Istnieje możliwość zastosowania wewnętrznej radioterapii. Pacjenci poddani tej formie leczenia przeciętnie przeżywają 6 miesięcy. Pacjenci u których ta metoda się nie powiodła także mogli by być leczeni terapią genową. Nowotwór może zostać usunięty z wlotu powietrza przy użyciu lasera lub kleszczyków do biopsji. To może zostać połączone z wstrzyknięciem konstruktów adenowirusowych, co zmniejszyłoby objętość zastrzyku. Podanie konstruktów wirusowych nie wyklucza otrzymywania przez pacjenta innych form terapii uśmierzającej, jeśli nowotwór rozwija się. Dołączono następujące przykłady aby zademonstrować korzystne odmiany wynalazku. Techniki opisane w przykładach zostały opracowane przez wynalazcę do poprawnego funkcjonowania przy stosowaniu wynalazku i mogą być uważane za korzystniejsze sposoby jego stosowania. Jednak obeznani ze stanem techniki powinni zdawać sobie sprawę, że można dokonać wielu zmian w przedstawionych zestawieniach, wciąż uzyskując takie same rezultaty i nie wychodząc poza ideę oraz zasięg wynalazku. Przykład I: Otrzymywanie wektora ekspresyjnego p53 Przykład opisuje otrzymywanie wektora ekspresyjnego p53. Wektor ten jest konstruowany według opisu i przeznaczony do zastąpienia regionu El (1,3-9,2 m.u.) genomu adenowirusa szczepu Ad5 oraz stosowany do konstruowania wiriona adenowirusowego opisanego w przykładzie II. Kaseta ekspresyjna p53 przedstawiona na fig. 1, która zawiera promotor ludzkiego wirusa cytomegalii (CMV) (Boshart i inni 1985), cdna p53 oraz wczesny sygnał poliadenylacji SV40, wstawiona została do pxcjll w miejsca Xba I i Cla (dostarczone przez Dr. Frank L. Graham, McMaster University, Canada). Wielkość genomu wynoszący około 35,4 kb, podzielono na 100 jednostek mapowych (1 m. u. = 0,35 kb). Kaseta ekspresyjna p53 zastąpiła region El (1,3-9,2 m.u.) genomu Ad5. Starter 1 ma sekwencję 5 -GGCCCACCCCCTTGGCTTC-3 (Seq ID Nr 1) i jest umieszczony w pierwszym intronie w dół od ludzkiego CMV głównego promotora genu IE (Boshart i inni 1985). Starter 2 ma sekwencję 5'-TTGTAACCATTATAAGCTGC-3' (Seq ID Nr 2) i jest umieszczony we wczesnym sygnale poliadenylacji SV40. Oba startery par zasad od wstawki cdna p53 z obu końców, definiują 1,40 kb produkt PCR. Starter 3 ma sekwencję 5'-TCGTTTCTCAGCAGCTGTTG-'3 (Seq ID Nr 3) a starter 4 ma sekwencję

16 '- CATCTGAACTCAAAGCGTGG-3' (Seq ID Nr 4) i znajdują się odpowiednio w 11 m.u. i 13,4 m.u. genou Ad5, co definiuje produkt PCR o 0,86 kb specyficzny dla genomu wirusowego. Przykład II: Otrzymywanie i namnażanie zrekombinowanych adenowirusów p53 Przykład opisuje jeden ze sposobów odpowiednich do otrzymywania niezależnych zrekombinowanych adenowirusów wywołujących ekspresję p53. Sposób otrzymywania zrekombinowanego adenowirusa oparty jest na fakcie, iż ze względu na ograniczenia pojemności adenowirusa pjm17 nie może on tworzyć własnych wirusów. Z tego powodu homologiczna rekombinacja plazmidu wektora ekspresyjnego i pjm17 wewnątrz komórki poddanej transfekcji prowadzi do uzyskania aktywnego wirusa, który może być wprowadzony jedynie do komórek, w których zachodzi ekspresja odpowiedniego białka adenowirusowego. W obecnym sposobie używa się komórek 293 jako komórek gospodarza do namnażania wirusa, zawierającego substytucje kaset ekspresyjnych obcego DNA w obszarze El lub E3. Ten proces wymaga kotransfekcji DNA do komórek 293. Transfekcja w dużym stopniu determinuje wydajność namnażania wirusów. Sposób transfekcji DNA do komórek 293 przed odkryciem tego wynalazku polegał zwykle na współstrącaniu fosforanu wapnia i DNA (Graham i van der Eb Jednak ta metoda połączona z analizą łysinkową jest stosunkowo trudna i zwykle daje niską wydajność reprodukcji wirusa. Jak pokazano w tym przykładzie transfekcja i następująca po niej identyfikacja zainfekowanych komórek znacznie się poprawiła gdy użyto do transfekcji liposomów oraz identyfikacji komórek po transfekcji poprzez efekt cytopatyczny (CPE). Hodowlę Unię komórek 293 prowadzono w zmodyfikowanej pożywce Dulbecco'a z dodatkiem 10% serum końskiego dezaktywowalnego pod wpływem ciepła. Wektor ekspresyjny p53 i plazmid pjm17 (McGrory i inni 1988) do rekombinacji homologicznej zostały poddane kotransfekcji do komórek 293 za pośrednictwem DOTAP (Boehringer Mannheim Biochemicals 1992). Jest to schematycznie przedstawione na fig. 1. Komórki 293 (pasaż 35, 60% konfluencji) zaszczepiono 24 godziny przed transfekcją na 60 mm szalkach lub na płytkach titracyjnych z 24 zagłębieniami. Komórki w każdym zagłębieniu poddano transfekcji z 30 μl DOTAP, 2 μl wektora ekspresyjnego p53 i 3 μl plazmidu pjm17. Po transfekcji do komórek dostarczano pożywkę MEM co 2-3 dni aż do pojawienia się CPE. Przykład III: Potwierdzanie tożsamości zrekombinowanych adenowirusa. Ten przykład ilustruje nowy sposób potwierdzenia, metodą reakcji łańcuchowej polimerazy (PCR), tożsamości zrekombinowanych wirionów po kotransfekcji do odpowiedniej linii komórek. Z płytek zebrano jednakową ilość nadsączu kultury komórkowej (50 do 370 μ ), dodano proteinazę K (50 μl/ml z 0,5% SDS i 20 mm EDTA), pozostawiono w temperaturze 56 C na 1 godzinę, ekstrahowano mieszaniną fenolu i chloroformu, a następnie etanolem wytrącano kwasy nukleinowe. Osadzone DNA ponownie zawieszono w 20 μl dh2o i użyto jako matrycę do amplifikacji PCR. Odpowiednie położenia starterów PCR i ich sekwencje pokazane są na fig. 1, są to odpowiednio Seq ID Nr: 1, 2, 3 i 4. Startery specyficzne dla wstawek określają produkt PCR o wielkości 1,4 kb, a startery specyficzne dla genomu wirusowego określają produkt PCR 0,86 kb. Reakcje PCR przeprowadzano w objętości 50 μ l, co zawierało 4 mm MgCl2, 50 mm KCl, 0,1% tryton X-100, 200 μl każdy z dntp'ow, 10 mm Tris-Cl (ph 9,0), 2 μ l każdy starter i 1,0 jednostkę polimerazy Taq (Promega). Reakcje przeprowadzano w temp. 94 C, 0,5 min, 56 C, 0,5 min oraz 72 C, 1min w 30 cyklach. W celu uproszczenia procedury identyfikacji nowo otrzymanych zrekombinowanych wirusów opracowano bezpośrednią próbę PCR na próbkach DNA z kultury komórek. Do równej objętości (50 lub 370 μm) nadsączu pożywki komórek z CPE dodano proteinazę K i ekstrahowano fenolem z chloroformem. Po wytrąceniu etanolem, próbki DNA zanalizowano metodą PCR, używając dwóch par starterów, w celu amplifikacji sekwencji specyficznych dla wstawki i genomu wirusowego. Miejsca odpowiadające starterom PCR i ich sekwencje są przedstawione na fig. 1. Startery 1, 2, 3 i 4 są to odpowiednio sekwencje nr 1, 2, 3 i 4. W rezultacie otrzymano wstawkę cdna o 1,4 kb i fragment genomu wirusowego o 0,86 kb z wektora ekspresyjnego (kontrola pozytywna) i próbki DNA z pozytywnej kultury
.")
17 komórek (fig. 2B, ścieżki odpowiednio 1 i 4). Jedynie fragment 0,86 kb amplifikowano z próbki DNA wirusa Ad4/RSV/GL2 (kontrola negatywna, ścieżka 2). Żadne prążki nie pojawiły się po reakcjach PCR, w których wykorzystano jeden z pozytywnych nadsączy pożywki kultury komórek (ścieżka 3). Te wyniki wskazują, że adenowirusy wprowadzone do pożywki kultury komórkowej są możliwe do wykrycia przez PCR, używając do przygotowania matryc DNA jedynie 50 μ nadsączu. Rezultaty te pozwolą na opracowanie ilościowych sposobów określania ilości adenowirusą, tradycyjnie dokonywanych analizą łysinkową. Sekwencja cdna p53 typu dzikiego w wirusie Ad5CMV-p53 została potwierdzona przez sekwencjonowanie dideoksy DNA na wirusowym DNA oczyszczonym z gradientu CsCl. Kontrolny wirus Ad5/RSV/GL2 otrzymywany w podobny sposób ma strukturę zbliżoną do Ad5CMV-p53 z wyjątkiem promotora wirusa mięśniaka Rousa i cdna luciferazy użytych w jego kasecie ekspresyjnej. Zrekombinowany adenowirus przenoszący gen b-galaktocydazy z E. coli (LacZO, Ad5CMV-LacZ, również ma strukturę podobną do Ad5CMV-p53 i uzyskany został od Dr Franka L. Grahama. Pojedyncze klony wirusów Ad5CMV-p53, Ad5/RSV/GL2 i Ad5CMV-LacZ uzyskano poprzez oczyszczanie kolonii zgodnie ze sposobem Grahama i Preveca (1991). Pojedyncze klony wirusowe namnażano w komórkach 293. Pożywka kultury komórek 293 wykazujących całkowity efekt cytopatyczny została zebrana i odwirowana przy 1000 x g przez 10 min. Z połączonych nadsączy pobrano próbki równe objętościowo i składowano w temp -20 C jako zapas wirusów. Miano wirusów określono badając kolonie (Graham i Prevec 1991). Infekcje linii komórek przeprowadzono dodając zawiesiny wirusów (0,5 ml na 60 mm szalkę) do komórek w jednej warstwie i inkubując w temperaturze pokojowej przez 30 min przy lekkim mieszaniu co 5 min. Następnie dodano pożywkę i ponownie wstawiono zainfekowane komórki do inkubatora o temp. 37 C. Wydajność przeniesienia genów zrekombinowanych adenowirusów wyznaczano także używając Ad5CMV-LacZ w różnych liniach komórkowych: H226Br, H3222, H460, HeLa, Hep G2, LM2 i Vero. W obecności barwnika X-gal po zainfekowaniu Ad5CMV-LacZ, przy wielokrotności infekcji 30 PFU/komórkę, wszystkie linie komórek % były wybarwione na niebiesko. Przykład IV: Ekspresja genu p53 ukierunkowana na Ad5CMV-p53 w ludzkich komórkach raka płuc. Ten przykład opisuje użycie zrekombinowanego adenowirusa p53 do zainfekowania ludzkich komórek reakcji płuc homozygotyczną delecją genu p53. Rezultatay pokazują, że rozwój tych komórek i ekspresja zmutowanych p53 została powstrzymana, co wskazuje na potencjał wirionu Ad5CNV-p53 jako przydatnego czynnika kontroli komórek metastatycznych. Wykonano analizę immunohistochemiczną zainfekowanych komórek w jednej warstwie, które zostały utrwalone 3,8% formaliną i poddane działaniu 3% H20 2 w metanolu przez 5 min. Analizy immunohistochemiczne przeprowadzono używając zestawu Vactastain Elite (Vector Burlingame, CA). Pierwszym przeciwciałem użytym było przeciwciało Pab 1801 przeciw-p53 (Oncogene Science, Manhasset, NY); MOPC-21 (Organon Teknika Corp., West Chester, PA) użyte jako kontrola negatywna. Drugim przeciwciałem było przeciwciało znakowane awidyną przeciw mysiemu IgG (Vector). Do wykrycia kompleksu antygenprzeciwciało użyto odczynnika kompleksującego peroksydazy biotyny z chrzanu. Na koniec komórki wybarwiono hematoksyliną Harris'a (Sigma) i osadzono na szkiełku przy pomocy Cytoseal 60 (Stephens Scientific, Riverdale N J). Aby zbadać in situ ekspresję p53 prowadzoną z promotorem CMY wirusa Ad5CMVp53, przeprowadzono analizę immunohistochemiczną zainfekowanych linii komórek. W linii komórek H358, które posiadają delecje homozygotyczną p53, gen p53 został przeniesiony z wydajnością %, co wykazała analiza imunohistochemiczna. Komórki zainfekowano Ad5CMV-p53 z wielokrotnością infekcji jednostek łysinkowych wirusa (PFU) na komórkę.

18 Wysoka wydajność transferu przez zrekombinowane adenowirusy potwierdzono stosując Ad5CMV-LacZ, wirus który przenosi gen LacZ ulegający transkrypcji pod ludzkim promotorem CMV IE. Przy wielokrotności infekcji PFU na komórkę, wszystkie badane komórki włącznie z HeLa, Hep G2, LM2 oraz ludzką linią komórkową NSCLC, wykazywały w % aktywność b-galaktozydazy w obecności barwika X-gal. Te rezultaty wskazują, że wektory adenowirusowe są skutecznym nośnikiem genów do transferu genów do ludzkich komórek rakowych. Przeprowadzono analizy Westerna na wszystkich lizatach komórek otrzymanych w wyniku lizy pojedynczych warstw komórek na szalkach z buforem SDS-PAGE (0,5 ml na 60 mm płytkę) po przemyciu komórek wodnym roztworem soli buforowanym fosforanem (PBS). Stosując metodę SDS-PAGE do ścieżek wprowadzono lizaty komórek odpowiadające 5x104 komórek (10-15 ml). Białka z żelu przenoszono na membranę Hybrond -ECL (Amersham, Arlington Heights, IL). Membrany zablokowano rozpuszczonym 0,5% mlekiem w proszku w PBS i znakowano pierwszorzędowymi przeciwciałami mysie przeciwciało monoklonalne i przeciw ludzkiemu p53 Pab 1801, mysie przeciwciało monoklonalne przeciw ludzkiej β-aktynie (Amersham), przemyto i sondowano drugorzędowym przeciwciałem: sprzężone z peroksydazą królicze przeciwciało przeciwko mysiemu IgG (Pierce Chemical Co., Rockford, IL). Membrany wywoływano zgodnie z rozszerzonym protokołem chemioluminescencji. Względne ilości egzogennego p53 otrzymanego w wyniku ekspresji określono densytometrycznie (Molecular Dynamics Inc. sunnyvale, CA). Analiza Westerna pokazała, że poziom ekspresji egzogennych białek p53 był wysoki (fig 5A ścieżki 2,3 oraz 5,6). Najwyższą ekspresję białek obserwowano 3 dni po zainfekowaniu (fig. 6 wstawka, 0,5 dni do 3 dni). Jako materiał kontrolny skonstruowano wirion o strukturze podobnej do rekombinowanego Ad5MCV-p53 według przykładu 1. Wirion ten zawiera cdna lucyferazy pod promotorem wirusa mięsaka Rousa LTR w kasecie ekspresyjnej wirionu. Nie wykryto ekspresji p53 ani zmiany ekspresji aktyny w komórkach zainfekowanych wirionem Ad5/RSV/GL2. Zrekombinowany adenowirus użyto do zainfekowania trzech ludzkich linii komórek płuc NSCLC: linii komórek H358, która ma homozygotyczną delecję genu p53, linii komórkowej H322, która posiada mutację punktową genu p53 w kodone 248 (G do T) oraz linii komórkowej H460, która posiada gen p53 typu dzikiego. Stopień rozwoju ludzkich komórek płuc NSCLC określono po zaszczepieniu H322 i H460 (1x105) lub (2xl05) na płytkach 60- mm w 24 godz. przed infekcją wirusową. Komórki zainfekowano wirusami z wielokrotnością infekcji (MOI ) 10 PFU na komórkę. Jako próbnej infekcji kontrolnej użyto pożywki. Trzy powtórzenia każdej z linii komórkowej poddanych działaniu różnych kuracji zliczano przez 1-6 dni po infekcji. Wzrost hodowli komórek H358 zainfekowanych Ad5CMV-p53 został ogromnie zahamowany w porównaniu do niezainfekowanych komórek i komórek zainfekowanych wirionem kontrolnym (fig. 7A). Wzrost hodowli komórek H322 również został znacznie zahamowany wirionem p53 (fig. 7B), podczas gdy rozwój komórek H460 zawierających p53 typu dzikiego został zmieniony w mniejszym stopniu (fig. 7C). Wzrost hodowli komórek H358 zainfekowanych wirusem Ad5CMV-p53 został zahamowany w 79%, podczas gdy niezainfekowanych komórek lub zainfekowanych wirusem kontrolnym nie został zahamowany. Rozwój linii komórkowej H322 posiadającej mutację punktową w p53 został przez Ad5CMV-p53 zahamowany w 72%, podczas kiedy rozwój linii komórkowej H460 zawierającej p53 typu dzikiego został zahamowany o 28%. Rezultaty wskazują, że zrekombinowane adenowirusy p53 posiadają właściwości powstrzymujące rozwój nowotworu, w wyniku przywracania funkcji białka p53 w komórkach nowotworowych. Przykład V: Leczenie za pomocą Ad5CMV-p53 komórek o brakującym p53. Przykład dotyczy użycia zrekombinowanych adenowirusa p53 do przywracania in vitro właściwości powstrzymujących rozwój komórek rakowych, a więc leczenia złośliwego lub metastatycznego rozwoju komórek. Przykład ten przedstawia niektóre sposoby leczenia raka przy wykorzystaniu terapii genowej za pośrednictwem adenowirusów.
.")
19 Komórki H358 zainfekowano wirusami Ad5CMV-p53 i Ad5/RSV/GL2 z wielokrotnością infekcji 10 PFU na komórkę. Równa ilość komórek leczona była pożywką jako infekcja próbna. 24 godziny po zainfekowaniu leczone komórki zebrano i przemyto dwukrotnie PBS. W każdym przypadku wstrzyknięto trzy miliony komórek w objętości 0,1 ml do każdej myszy (Harlan Co., Houston TX). Każdy sposób leczenia prowadzono na pięciu myszach. Myszy naświetlano (300 cgy, 60Co) przed zastrzykiem i badano tydzień po zastrzyku. Oceniono stopień tworzenia się guza na końcu 6-tygodniowego okresu i obliczono objętość guza uznając jego kształt za sferyczny o średnicy równej średniej średnic w różnych kierunkach. Aby określić efekt hamujący rakotwórczość, komórki H358 (ludzkie komórki NSCLC) wstrzyknięto myszom, w celu wywołania rozwoju nowotworu. Każda mysz otrzymała jeden zastrzyk komórek zainfekowanych AdCMV-p53 lub Ad5/RSV/GL2 przy 10 PFU na komórkę przez 24 godz. Jako próbnie zainfekowane kontrolne komórki użyto komórki H358 leczone jedynie pożywką. Guzy początkowo wyczuwalne palcami w 14 dniu po infekcji wywołane były jedynie próbnymi lub kontrolnymi komórkami zainfekowanych wirusem jak przedstawiono w tabeli I: Tabela I: Wpływ Ad5CMV-p53 na rakotwórczość H358 u nagiej myszy1 Sposób leczenia Liczba guzów Liczba myszy (%) Średnia objętość (mm3 ± SD) Pożywka 4/5(80) 37 ±12 Ad5/RSV/GL2 3/4(75) 30 ±14 Ad5CMV-p53 0/4 (0) - 'Leczone komórki H358 wstrzyknięto podskórnie w ilości 2 x 106 komórek na mysz. Rozmiary guzów Jak przedstawiono w Tabeli I u myszy, które otrzymały komórki leczone Ad5CMV-p53 nie rozwinęły się nowotwory. Guzy na koniec 6-tygodniowego okresu miały 4-10 mm średnicy. Badania te były rozpoczęte z pięcioma myszami na grupę; jedna z myszy w grupie Ad5CMV-p53 lub Ad5/RSV/GL2 nie dotrwała do końca badania. Wczesna śmierć wystąpiła prawdopodobnie z powodu infekcji szpitalnej. Przykład VI: Ad5CMV-p53 w leczeniu raka płuc Przykład dotyczy zastosowania produktów rekombinacji adenowirusa p53 do przywrócenia hamowania rozwoju komórek rakowych in vivo, a więc leczenia raka u zwierząt. Opisuje kilka sposobów w jaki wynalazek może być zastosowany do leczenia raka drogą terapii genowej za pośrednictwem adenowirusów. Wydajność Ad5CMV-p53 w hamowaniu rakotwórczości oceniono na mysim modelu ludzkiego ortotopicznego raka płuc. Ponieważ komórki H358 i H322 nie tworzyły nowotworów w tym modelu, użyto linii komórkowej H226Br. Ta linia komórkowa pochodzi od komórek raka płuc. H226Br posiada mutację punktową (ATC na GTC) w egzonie 7, kodon 254 genu p53 i jest rakotwórcza u myszy. Opisano już (Georges i inni 1993) procedurę do badania na modelu mysim ludzkiego ortotopicznego raka płuc została już opisana. W skrócie, nagim myszom naświetlanym (300 cgy, Co) zaszczepiono w oskrzelach komórki H226Br. Każda mysz dostała 2 x 106 komórek w objętości 0,1 ml PBS. Trzy dni po zaszczepieniu, 10 myszy z każdej grupy leczono 0,1 ml wirusów lub zarobki (PBS) przez zaszczepienie w oskrzelach raz dziennie przez dwa dni. Dawka wirusa wynosiła 5 x 107 Ad5CMV-p53 lub Ad5/RSV/GL2 na mysz. Myszy następnie poddano eutanazji po 6 tygodniach. Określono stopień uformowania się guza dokonując sekcji płuc i śródpiersia i mierząc rozmiary guzów. Guzy potwierdzono analizą histologiczną fragmentów masy guza. Naświetlonym nagim myszom zaszczepiono 2 x 106 komórek H226Br na mysz poprzez i zaszczepienie w oskrzelach. Trzy dni po zaszczepieniu, każdą z myszy leczono 0,1 ml Ad5CMV-p53 lub Ad5/RSV/GL2 lub zaróbką (PBS) poprzez zaszczepienie w oskrzelach raz dziennie przez dwa dni. Użyta dawka wirusa wynosiła 5 x 107 PFU na mysz. Stopień ufor-

20 mowania się guza oceniono po 6 tygodniach, poprzez sekcję tkanek płuc i śródpiersia oraz zmierzenie wielkości nowotworu. Schemat procedury przedstawia fig. 7, a reprezentatywne próbki z sekcji przedstawione są na fig. 8. Wykryte guzy potwierdzono przez analizy histologiczne. Dane dotyczące wielkości guzów zebrano w tabeli II: Tabela II. Wpływ Ad5CMV-p53 na rakotwórczość H226Br w modelu mysim ortotopicznego raka płuc u ludzi2 Sposób leczenia Liczba myszy z guzami Całkowita liczba myszy Objętość guza (mm3 ± SD) Zaróbką 7/10(70) 30 ±8,4 Ad5/RSV/GL2 8/10(80) 25 ± 6,9 Ad5CMV-p53 2/8(25) 8±3,33 1określono na końcu okresu 6-tygodniowego. 2 Myszom w oskrzelach zaszczepiono 2 x 1006 komórek H226Br na mysz. Na 3 dzień po zaszczepieniu myszy otrzymały zaróbkę lub wirusy (5 x 107 każda w 0,1 ml) do oskrzeli raz dziennie przez 2 dni. Formowanie się guza określono po 6 tygodniach. 3p<0,05 poprzez dwustronną analizę wariancji w porównaniu z grupami otrzymującymi zaróbkę (PBS) lub wirus kontrolny Jedynie u 25% myszy leczonych Ad5CMV-p53 utworzyły się guzy, podczas gdy w przypadku zarobki lub wirusa kontrolnego u 70-80% leczonych myszy rozwinęły się nowotwory. Przeciętna wielkość guza grupy Ad5CMV-p53 była znacznie mniejsza niż w przypadku grup kontrolnych. Rezultaty te wskazują, że Ad5CMV-p53 może zapobiegać tworzeniu nowotworów H226Br w mysim modelu ludzkiego ortotopicznego raka płuc. Przykład VII: Sposób leczenia Ad5CMV-p53 Naturalnie zwierzęce modele będą stosowane jako część prób przedklinicznych, jak opisano w przykładach V i VI. Następnie jednak pacjenci, dla których ustalono wskazanie medyczne do leczenia genowego, mogą być przebadani na obecność przeciwciał skierowanych przeciwko adenowirusom. Jeśli przeciwciała występują, a u pacjenta występowały już alergie na substancje farmakologiczne lub występujące w przyrodzie, dawkę testową 103 do 106 zrekombinowane adenowirusy podaje się pod ścisłą obserwacją kliniczną. Do leczenia raka przy użyciu Ad5CMV-p53, mogą być przygotowane i oczyszczone zrekombinowane adenowirusy wywołujące ekspresję p53 pod kontrolą odpowiednich promotorów/enchancerów, takich jak promotor CMV, zgodnie ze sposobem akceptowanym przez Ministerstwo Opieki Zdrowotnej (Food and Drug Administration, FDA) do podawania ludziom. Metody takie to między innymi wirowanie w gradiencie gęstości chlorku cezu, po którym następuje bada się wydajność i czystość preparatu. Dwa podstawowe sposoby uważane są za najodpowiedniejsze sposoby leczenia adenowirusem p53, tj. podawanie bezpośrednie lub miejscowe oraz podawanie bardziej ogólne. Sposoby te są odpowiednie do leczenia wielu odmian raka, o których wiadomo, że są związane z mutacjami p53. W przypadku podawania ogólnego wykazano, że zwykły zastrzyk dożylny adenowirusów jest wystarczający aby spowodować infekcję wirusową tkanek w miejscach oddalonych od zastrzyku (Stratford-Perricaudet i inni 1991 b), jest więc właściwy do leczenia nowotworów związanych z p53. Wirus może być podany pacjentom dożylnie w roztworze akceptowalnym farmakologicznie lub jako wlew przez pewien okres. Mówiąc ogólnie, uważa się, że wydajna ilość funkcjonalnych wirusów, którą można podać pacjentowi wynosi od 1 x1012do 5 x1012. Również, szczególnie przy raku płuc, może być zastosowane w razie potrzeby, bardziej bezpośrednie podawanie, analogicznie do dooskrzelowego podawania regulatora przewodnictwa membranowego w mukowiscydozie (Rosenfeld i inni 1992). To spowodowałoby dostarczenie adenowirusa p53 bliżej do miejsca komórek docelowych. Dokładniej rzecz biorąc korzystne protokoły leczenia mogą być opracowane według następujących zasad. Pacjenci mogą najpierw przejść bronchoskopię aby określić poziom niedrożności. Jak największa część nowotworu powinna zostać usunięta endoskopowo. Pacjenci powinni przechodzić korzystnie bronchoskopię pod miejscowym lub ogólnym znieczuleniem.

21 Igła Sticofor powinna zostać przełożona przez kanał biopsji bronchoskopu. Do pozostającej części nowotworu należy następnie wstrzyknąć adenowirus p53 w małej objętości takiej jak 10 ml lub mniejszej. W każdym przypadku, ze względu na to, że użyty adenowirus będzie wybrakowany replikacyjnie, nie oczekuje się żadnego niekorzystnego efektu wpływu wirusa na zdrowie leczonego. Jednak pacjenci powinni być hospitalizowani podczas leczenia przez co najmniej 48 godzin aby obserwować niepożądane reakcje. Sprawy związane z bezpieczeństwem użycia wybrakowanych replikacyjnie adenowirusów jako środka przenoszenia genów do komórek ludzkich poruszane już były w przeszłości (Rosenfeld i inni 1992; Jaffe i inni 1992), lecz dawka adenowirusa do podania powinna być odpowiednio obserwowana aby zminimalizować prawdopodobieństwo efektów ubocznych. Istnieją różne kryteria jakie należy wziąć pod uwagę przy określaniu toksyczności. Aby określić istnienie toksyczności podłoże guza powinno być sfotografowane przed rozpoczęciem terapii. Zmierzyć należy najdłuższą średnicę. Na podstawie średnic należy określić wielkość. Z tych danych wyliczyć można stopień ponownego rozwoju guza. Okres postępu leczenia może być również mierzony od pierwszej obserwacji obniżenia masy guza do czasu uzyskania dowodów na postępowanie choroby. Progresja choroby definiowana jest jako wzrost o ponad 25% w sumie średnic. Pacjenci powinni otrzymać dwie tury leczenia przed zadecydowaniem o progresywności. Poziom przeżywania pacjentów mierzony będzie wpisami do protokołu. Następujące potem badania zawierałyby wszystkie sposoby stosowane rutynowo w leczeniu raka, między innymi: obserwowanie znaków klinicznych i pobieranie biopsji do standardowych i molekularnych analiz, w których ocenić można ekspresję różnych genów p53. Dałoby to informacje na temat ilości komórek, które przejęły niesiony gen i o względnej mocy promotora in vivo. Opierając się na tych danych można wprowadzić ulepszenia do metody leczenia. Usprawnienia te mogą obejmować konstrukcje adenowirusowe z różnymi promotorami lub zmianę ilości wstrzykiwanych jednostek łysinkowych wirusa, aby zapewnić zainfekowanie większej ilości lub wszystkich komórek rakowych bez niefizjologicznej nadekspresji zrekombinowanych genów. Uważa się, że ekspresja egzogennych genów przenoszonych in vivo poprzez adenowirus może trwać przez przedłużony okres czasu. Terapeutycznie wydajna długoterminowa ekspresja egzogennych genów przeniesionych wirusowo będzie musiała być w każdym przypadku specyficzna. Użyteczność genów markerowych do oceniania terapeutycznie skutecznego czasu trwania ekspresji genów jest ograniczona, jako ze poziomy ekspresji wymagane do poprawy każdego błędu genetycznego mogą znacznie różnić się od poziomu wymaganego do całkowitego wyleczenia innej choroby. Pomimo, że kompozycje i sposoby według wynalazku opisano poprzez korzystne zestawienia, jasne będzie dla biegłych w stanie techniki, iż można zastosować różne odmiany kompozycji, sposobów i w różnych etapach czy kolejności etapów w sposobach, bez odchodzenia od idei i zakresu wynalazku. Mówiąc szczegółowo oczywista jest możliwość zastąpienia pewnych czynników przez inne czynniki chemicznie i fizjologicznie powiązane jeśli wyniki byłyby takie same. Wszystkie takie zastąpienia oczywiste dla biegłych w stanie techniki zawsze pozostaną w zakresie i idei wynalazku, jak zdefiniowano w zastrzeżeniach. (0) LISTINGI SEKWENCJI (1) INFORMACJE OGÓLNE (i) ZGŁASZAJĄCY: (A) NAZWA: BOARD OF REGENTS, THE UNIVERSITY OF TEXAS SYSTEM (B) ULICA: 201 West 7th Street (C) STAN: Austin (D) KRAJ: Stany Zjednoczone Ameryki Północnej (E) KOD POCZTOWY: (F) TELEFON: (512) , (G) TELEFAX: (512)
22 (ii) WYNALAZCY: ZHANG, Wei-Wei; ROTH, Jack A. (iii) TYTUŁ WYNALAZKU: Produkty rekombinacji adenowirusów, kompozycje zawierające je, sposoby ich użycia i zastosowania (iv) ILOŚĆ SEKWENCJI: 4 (v) ADRES KORESPONDENCYJNY: (A) ADRESAT: Arnold, White & Durkee (B) STREET: P.O. BOK 4433 (C) MIASTO: Houston (D) STAN:Texas (E) KRAJ: USA (F) KOD: (vi) FORMA KOMPUTEROWA: (A) NOŚNIK: Dyskietka (B) KOMPUTER: IBM PC (C) SYSTEM OPERACYJNY: PC-DOS/MS-DOS, ASCII (D) OPROGRAMOWANIE: PATENTIN RELEASE #1.0, WERSJA #1,25 (vii) OBECNE DANE O ZGŁOSZENIU: (A) NUMER ZGŁOSZENIA: PCT/US94/12401 (B) DATA ZGŁOSZENIA: 28 PAŹDZIERNIKA 1994 (C) KLASYFIKACJA: NIEZNANA (viii) WCZEŚNIEJSZE DANE O ZGŁOSZENIU: (A) NUMER ZGŁOSZENIA: USSN 08/145,826 (B) DATA ZGŁOSZENIA: 29 PAŹDZIERNIKA 1993 (C) KLASYFIKACJA: NIEZNANA (ix) INFORMACJA O RZECZNIKU: (A) IMIĘ I NAZWISKO: BARBARA S. KTCHELL (B) NUMER REJESTRACJI: 33, 928 (C) NUMER ODNIESIENIA: UTFC350P (x) INFORMACJA TELEKOMUNIKACYJNA: (A) TELEFON: (512) (B) TELEFAX: (713) (C) TELEX: (2) INFORMACJA O SEKWENCJI NR 1 (i) CHARAKTERYSTYKA SEKWENCJI: (A) DŁUGOŚĆ: 19 par bazowych (B) TYP: kwas nukleinowy (C) ŁAŃCUCH: pojedynczy (D) TOPOLOGIA: linerara (ii) TYP CZĄSTECZKI: inny kwas nukleinowy (A) OPIS: /desc = DNA (iii) OPIS SEKWENCJI: SEKWENCJA NR 1: GGCCCACCCC CTTGGCTTC 19 (3) INFORMACJA O SEKWENCJI NR 2 (i) CHARAKTERYSTYKA SEKWENCJI: (A) DŁUGOŚĆ: 20 par bazowych (B) TYP: kwas nukleinowy (C) ŁAŃCUCH: pojedynczy (D) TOPOLOGIA: linerama (ii) TYP CZĄSTECZKI: inny kwas nukleinowy (A) OPIS: /desc = DNA (iii) OPIS SEKWENCJI: SEKWENCJA NR 2: TTGTAACCAT TATAAGCTGC 20
23 (4) INFORMACJA O SEKWENCJI NR 3: (i) CHARAKTERYSTYKA SEKWENCJI: (A) DŁUGOŚĆ: 20 par bazowych (B) TYP: kwas nukleinowy (C) ŁAŃCUCH: pojedynczy (D) TOPOLOGIA: linerama (ii) TYP CZĄSTECZKI: inny kwas nukleinowy (A) OPIS: /desc = DNA (iii) OPIS SEKWENCJI: SEKWENCJA NR 3: TCGTTTCTCAGCAGCTGTTG 20 (5) INFORMACJA O SEKWENCJI NR 4 (i) CHARAKTERYSTYKA SEKWENCJI: (A) DŁUGOŚĆ: 20 par bazowych (B) TYP: kwas nukleinowy (C) ŁAŃCUCH: pojedynczy (D) TOPOLOGIA: linerama (ii) TYP CZĄSTECZKI: inny kwas nukleinowy (A) OPIS: /desc = DNA (iii) OPIS SEKWENCJI: SEKWENCJA NR 4: CATCTGAACT CAAAGCGTGG
24
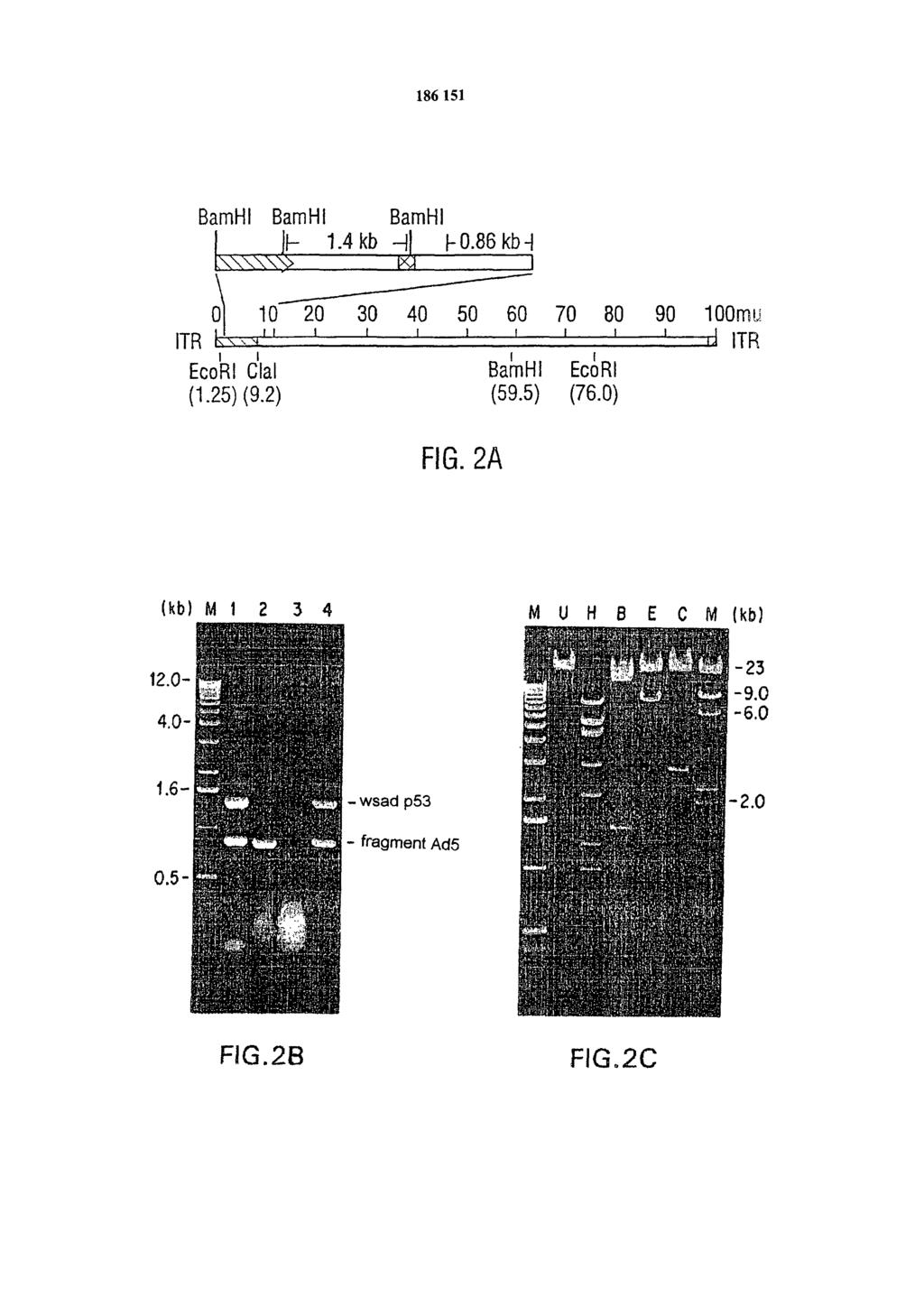
25

26

27
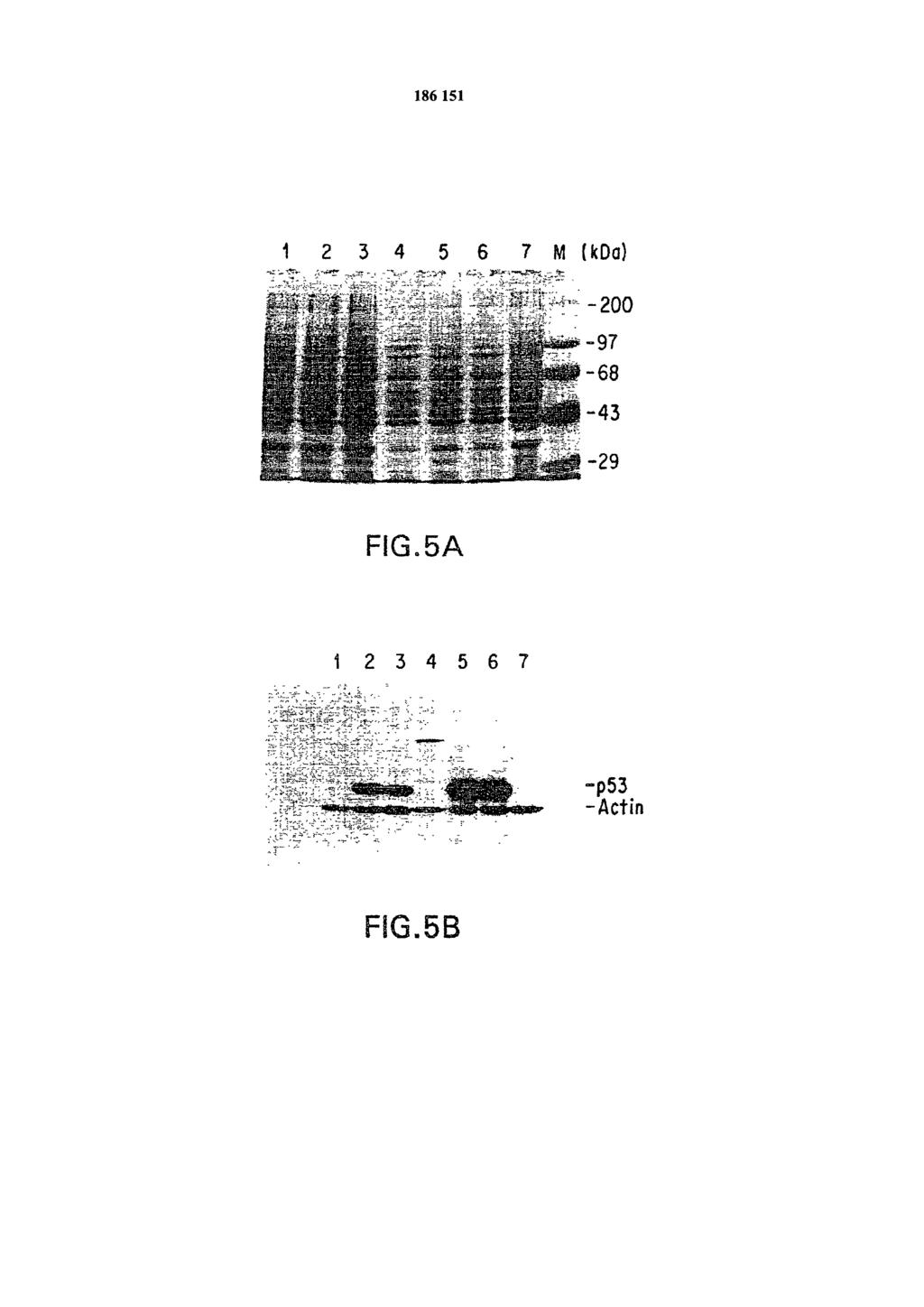
28
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE. Ewa Waszkowska ekspert UPRP
 WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
Rozmnażanie i wzrost komórek sąściśle kontrolowane. Genetyczne podłoże nowotworzenia
 Rozmnażanie i wzrost komórek sąściśle kontrolowane Genetyczne podłoże nowotworzenia Rozmnażanie i wzrost komórek sąściśle kontrolowane Rozmnażanie i wzrost komórek sąściśle kontrolowane Połączenia komórek
Rozmnażanie i wzrost komórek sąściśle kontrolowane Genetyczne podłoże nowotworzenia Rozmnażanie i wzrost komórek sąściśle kontrolowane Rozmnażanie i wzrost komórek sąściśle kontrolowane Połączenia komórek
Nowoczesne systemy ekspresji genów
 Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Inżynieria genetyczna- 6 ECTS. Inżynieria genetyczna. Podstawowe pojęcia Część II Klonowanie ekspresyjne Od genu do białka
 Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
CHOROBY NOWOTWOROWE. Twór składający się z patologicznych komórek
 CHOROBY NOWOTWOROWE Twór składający się z patologicznych komórek Powstały w wyniku wielostopniowej przemiany zwanej onkogenezą lub karcinogenezą Morfologicznie ma strukturę zbliżoną do tkanki prawidłowej,
CHOROBY NOWOTWOROWE Twór składający się z patologicznych komórek Powstały w wyniku wielostopniowej przemiany zwanej onkogenezą lub karcinogenezą Morfologicznie ma strukturę zbliżoną do tkanki prawidłowej,
Klonowanie molekularne Kurs doskonalący. Zakład Geriatrii i Gerontologii CMKP
 Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych
 Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
ROZPORZĄDZENIE KOMISJI (UE) / z dnia r.
 / z dnia r.") KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
TERAPIA GENOWA. dr Marta Żebrowska
 TERAPIA GENOWA dr Marta Żebrowska Pracownia Diagnostyki Molekularnej i Farmakogenomiki, Zakładu Biochemii Farmaceutycznej, Uniwersytetu Medycznego w Łodzi Źródło zdjęcia: httpblog.ebdna.plindex.phpjednoznajwiekszychzagrozenludzkosciwciazniepokonane
TERAPIA GENOWA dr Marta Żebrowska Pracownia Diagnostyki Molekularnej i Farmakogenomiki, Zakładu Biochemii Farmaceutycznej, Uniwersytetu Medycznego w Łodzi Źródło zdjęcia: httpblog.ebdna.plindex.phpjednoznajwiekszychzagrozenludzkosciwciazniepokonane
Zawartość. Wstęp 1. Historia wirusologii. 2. Klasyfikacja wirusów
 Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
Inżynieria Genetyczna ćw. 3
 Materiały do ćwiczeń z przedmiotu Genetyka z inżynierią genetyczną D - blok Inżynieria Genetyczna ćw. 3 Instytut Genetyki i Biotechnologii, Wydział Biologii, Uniwersytet Warszawski, rok akad. 2018/2019
Materiały do ćwiczeń z przedmiotu Genetyka z inżynierią genetyczną D - blok Inżynieria Genetyczna ćw. 3 Instytut Genetyki i Biotechnologii, Wydział Biologii, Uniwersytet Warszawski, rok akad. 2018/2019
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny
 Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny Zadanie 1 1 pkt. za prawidłowe podanie typów dla obydwu zwierząt oznaczonych literami A oraz B. A. ramienionogi, B. mięczaki A.
Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny Zadanie 1 1 pkt. za prawidłowe podanie typów dla obydwu zwierząt oznaczonych literami A oraz B. A. ramienionogi, B. mięczaki A.
Dane mikromacierzowe. Mateusz Markowicz Marta Stańska
 Dane mikromacierzowe Mateusz Markowicz Marta Stańska Mikromacierz Mikromacierz DNA (ang. DNA microarray) to szklana lub plastikowa płytka (o maksymalnych wymiarach 2,5 cm x 7,5 cm) z naniesionymi w regularnych
Dane mikromacierzowe Mateusz Markowicz Marta Stańska Mikromacierz Mikromacierz DNA (ang. DNA microarray) to szklana lub plastikowa płytka (o maksymalnych wymiarach 2,5 cm x 7,5 cm) z naniesionymi w regularnych
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Dr. habil. Anna Salek International Bio-Consulting 1 Germany
 1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii
 Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Gdzie chirurg nie może - - tam wirusy pośle. czyli o przeciwnowotworowych terapiach wirusowych
 Gdzie chirurg nie może - - tam wirusy pośle czyli o przeciwnowotworowych terapiach wirusowych Wirusy zwiększające ryzyko rozwoju nowotworów HBV EBV HCV HTLV HPV HHV-8 Zmniejszenie liczby zgonów spowodowanych
Gdzie chirurg nie może - - tam wirusy pośle czyli o przeciwnowotworowych terapiach wirusowych Wirusy zwiększające ryzyko rozwoju nowotworów HBV EBV HCV HTLV HPV HHV-8 Zmniejszenie liczby zgonów spowodowanych
KLONOWANIE DNA REKOMBINACJA DNA WEKTORY
 KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
października 2013: Elementarz biologii molekularnej. Wykład nr 2 BIOINFORMATYKA rok II
 10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
Agenda: Wynalazek jako własnośd intelektualna. Co może byd wynalazkiem. Wynalazek biotechnologiczny. Ochrona patentowa
 dr Bartosz Walter Agenda: Wynalazek jako własnośd intelektualna Co może byd wynalazkiem Wynalazek biotechnologiczny Ochrona patentowa Procedura zgłoszenia patentowego Gdzie, kiedy i jak warto patentowad
dr Bartosz Walter Agenda: Wynalazek jako własnośd intelektualna Co może byd wynalazkiem Wynalazek biotechnologiczny Ochrona patentowa Procedura zgłoszenia patentowego Gdzie, kiedy i jak warto patentowad
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
Warszawa, dnia 3 sierpnia 2016 r. Poz. 1173
 Warszawa, dnia 3 sierpnia 2016 r. Poz. 1173 ROZPORZĄDZENIE MINISTRA ŚRODOWISKA 1) z dnia 18 lipca 2016 r. w sprawie określenia wzorów wniosków oraz zgłoszeń związanych z zamkniętym użyciem mikroorganizmów
Warszawa, dnia 3 sierpnia 2016 r. Poz. 1173 ROZPORZĄDZENIE MINISTRA ŚRODOWISKA 1) z dnia 18 lipca 2016 r. w sprawie określenia wzorów wniosków oraz zgłoszeń związanych z zamkniętym użyciem mikroorganizmów
(86) Data i numer zgłoszenia międzynarodowego: 13.12.1999,PCT/EP99/09864 (87) Data i numer publikacji zgłoszenia międzynarodowego:
 Data i numer zgłoszenia międzynarodowego: 13.12.1999,PCT/EP99/09864 (87) Data i numer publikacji zgłoszenia międzynarodowego:") RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 202483 (21) Numer zgłoszenia: 349335 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 13.12.1999 (86) Data i numer zgłoszenia
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 202483 (21) Numer zgłoszenia: 349335 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 13.12.1999 (86) Data i numer zgłoszenia
Co to jest transkryptom? A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH 2
 ALEKSANDRA ŚWIERCZ Co to jest transkryptom? A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH 2 Ekspresja genów http://genome.wellcome.ac.uk/doc_wtd020757.html A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH
ALEKSANDRA ŚWIERCZ Co to jest transkryptom? A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH 2 Ekspresja genów http://genome.wellcome.ac.uk/doc_wtd020757.html A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH
SPOSÓB PRZEDSTAWIANIA DOKUMENTACJI DOŁĄCZANEJ DO WNIOSKU O DOPUSZCZENIE DO OBROTU PRODUKTU LECZNICZEGO WETERYNARYJNEGO IMMUNOLOGICZNEGO
 Załącznik nr 2 SPOSÓB PRZEDSTAWIANIA DOKUMENTACJI DOŁĄCZANEJ DO WNIOSKU O DOPUSZCZENIE DO OBROTU PRODUKTU LECZNICZEGO WETERYNARYJNEGO IMMUNOLOGICZNEGO CZĘŚĆ I STRESZCZENIE DOKUMENTACJI I A Wniosek o dopuszczenie
Załącznik nr 2 SPOSÓB PRZEDSTAWIANIA DOKUMENTACJI DOŁĄCZANEJ DO WNIOSKU O DOPUSZCZENIE DO OBROTU PRODUKTU LECZNICZEGO WETERYNARYJNEGO IMMUNOLOGICZNEGO CZĘŚĆ I STRESZCZENIE DOKUMENTACJI I A Wniosek o dopuszczenie
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU
 SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Czy priony zawsze są szkodliwe? SPIS TREŚCI: Wprowadzenie. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. Karty pracy. 1.
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Czy priony zawsze są szkodliwe? SPIS TREŚCI: Wprowadzenie. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. Karty pracy. 1.
Materiał i metody. Wyniki
 Abstract in Polish Wprowadzenie Selen jest pierwiastkiem śladowym niezbędnym do prawidłowego funkcjonowania organizmu. Selen jest wbudowywany do białek w postaci selenocysteiny tworząc selenobiałka (selenoproteiny).
Abstract in Polish Wprowadzenie Selen jest pierwiastkiem śladowym niezbędnym do prawidłowego funkcjonowania organizmu. Selen jest wbudowywany do białek w postaci selenocysteiny tworząc selenobiałka (selenoproteiny).
Making the impossible possible: the metamorphosis of Polish Biology Olympiad
 Making the impossible possible: the metamorphosis of Polish Biology Olympiad Takao Ishikawa Faculty of Biology, University of Warsaw, Poland Performance of Polish students at IBO Gold Silver Bronze Merit
Making the impossible possible: the metamorphosis of Polish Biology Olympiad Takao Ishikawa Faculty of Biology, University of Warsaw, Poland Performance of Polish students at IBO Gold Silver Bronze Merit
Tematyka zajęć z biologii
 Tematyka zajęć z biologii klasy: I Lp. Temat zajęć Zakres treści 1 Zapoznanie z przedmiotowym systemem oceniania, wymaganiami edukacyjnymi i podstawą programową Podstawowe zagadnienia materiału nauczania
Tematyka zajęć z biologii klasy: I Lp. Temat zajęć Zakres treści 1 Zapoznanie z przedmiotowym systemem oceniania, wymaganiami edukacyjnymi i podstawą programową Podstawowe zagadnienia materiału nauczania
Transformacja pośrednia składa się z trzech etapów:
 Transformacja pośrednia składa się z trzech etapów: 1. Otrzymanie pożądanego odcinka DNA z materiału genetycznego dawcy 2. Wprowadzenie obcego DNA do wektora 3. Wprowadzenie wektora, niosącego w sobie
Transformacja pośrednia składa się z trzech etapów: 1. Otrzymanie pożądanego odcinka DNA z materiału genetycznego dawcy 2. Wprowadzenie obcego DNA do wektora 3. Wprowadzenie wektora, niosącego w sobie
Oznaczenie polimorfizmu genetycznego cytochromu CYP2D6: wykrywanie liczby kopii genu
 Ćwiczenie 4 Oznaczenie polimorfizmu genetycznego cytochromu CYP2D6: wykrywanie liczby kopii genu Wstęp CYP2D6 kodowany przez gen występujący w co najmniej w 78 allelicznych formach związanych ze zmniejszoną
Ćwiczenie 4 Oznaczenie polimorfizmu genetycznego cytochromu CYP2D6: wykrywanie liczby kopii genu Wstęp CYP2D6 kodowany przez gen występujący w co najmniej w 78 allelicznych formach związanych ze zmniejszoną
Wymagania edukacyjne
 Rok szkolny 2018/2019 Wymagania edukacyjne Przedmiot Klasa Nauczyciel uczący Poziom biologia 1t Edyta Nowak podstawowy Ocena dopuszczająca Ocenę dopuszczającą otrzymuje uczeń, który: przyswoił treści konieczne,
Rok szkolny 2018/2019 Wymagania edukacyjne Przedmiot Klasa Nauczyciel uczący Poziom biologia 1t Edyta Nowak podstawowy Ocena dopuszczająca Ocenę dopuszczającą otrzymuje uczeń, który: przyswoił treści konieczne,
STRESZCZENIE PRACY DOKTORSKIEJ
 mgr Bartłomiej Rospond POSZUKIWANIE NEUROBIOLOGICZNEGO MECHANIZMU UZALEŻNIENIA OD POKARMU - WPŁYW CUKRÓW I TŁUSZCZÓW NA EKSPRESJĘ RECEPTORÓW DOPAMINOWYCH D 2 W GRZBIETOWYM PRĄŻKOWIU U SZCZURÓW STRESZCZENIE
mgr Bartłomiej Rospond POSZUKIWANIE NEUROBIOLOGICZNEGO MECHANIZMU UZALEŻNIENIA OD POKARMU - WPŁYW CUKRÓW I TŁUSZCZÓW NA EKSPRESJĘ RECEPTORÓW DOPAMINOWYCH D 2 W GRZBIETOWYM PRĄŻKOWIU U SZCZURÓW STRESZCZENIE
Raport z badania Działanie wirusobójcze środka dezynfekującego wobec Feline calicivirus. 25 października 2006
 Raport z badania Działanie wirusobójcze środka dezynfekującego wobec Feline calicivirus 25 października 2006 Dr Tobias J. Tuthill Wydział Nauk Biologicznych Uniwersytet Leeds Leeds LS2 9JT www.fbs.leeds.ac.uk
Raport z badania Działanie wirusobójcze środka dezynfekującego wobec Feline calicivirus 25 października 2006 Dr Tobias J. Tuthill Wydział Nauk Biologicznych Uniwersytet Leeds Leeds LS2 9JT www.fbs.leeds.ac.uk
Pamiętając o komplementarności zasad azotowych, dopisz sekwencję nukleotydów brakującej nici DNA. A C C G T G C C A A T C G A...
 1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
Biologia komórki i biotechnologia w terapii schorzeń narządu ruchu
 Biologia komórki i biotechnologia w terapii schorzeń Ilość godzin: 40h seminaria Ilość grup: 2 Forma zaliczenia: zaliczenie z oceną Kierunek: Fizjoterapia ścieżka neurologiczna Rok: II - Lic Tryb: stacjonarne
Biologia komórki i biotechnologia w terapii schorzeń Ilość godzin: 40h seminaria Ilość grup: 2 Forma zaliczenia: zaliczenie z oceną Kierunek: Fizjoterapia ścieżka neurologiczna Rok: II - Lic Tryb: stacjonarne
Konspekt do zajęć z przedmiotu Genetyka dla kierunku Położnictwo dr Anna Skorczyk-Werner Katedra i Zakład Genetyki Medycznej
 Seminarium 1 część 1 Konspekt do zajęć z przedmiotu Genetyka dla kierunku Położnictwo dr Anna Skorczyk-Werner Katedra i Zakład Genetyki Medycznej Genom człowieka Genomem nazywamy całkowitą ilość DNA jaka
Seminarium 1 część 1 Konspekt do zajęć z przedmiotu Genetyka dla kierunku Położnictwo dr Anna Skorczyk-Werner Katedra i Zakład Genetyki Medycznej Genom człowieka Genomem nazywamy całkowitą ilość DNA jaka
Biologia molekularna
 Biologia molekularna 1. Metryczka Nazwa Wydziału Program kształcenia Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Analityka Medyczna, studia jednolite magisterskie, studia stacjonarne i niestacjonarne
Biologia molekularna 1. Metryczka Nazwa Wydziału Program kształcenia Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Analityka Medyczna, studia jednolite magisterskie, studia stacjonarne i niestacjonarne
Promotor: prof. dr hab. Katarzyna Bogunia-Kubik Promotor pomocniczy: dr inż. Agnieszka Chrobak
 INSTYTUT IMMUNOLOGII I TERAPII DOŚWIADCZALNEJ IM. LUDWIKA HIRSZFELDA WE WROCŁAWIU POLSKA AKADEMIA NAUK mgr Milena Iwaszko Rola polimorfizmu receptorów z rodziny CD94/NKG2 oraz cząsteczki HLA-E w patogenezie
INSTYTUT IMMUNOLOGII I TERAPII DOŚWIADCZALNEJ IM. LUDWIKA HIRSZFELDA WE WROCŁAWIU POLSKA AKADEMIA NAUK mgr Milena Iwaszko Rola polimorfizmu receptorów z rodziny CD94/NKG2 oraz cząsteczki HLA-E w patogenezie
SPOSÓB PRZEDSTAWIANIA DOKUMENTACJI DOŁĄCZANEJ DO WNIOSKU O DOPUSZCZENIE DO OBROTU PRODUKTU LECZNICZEGO WETERYNARYJNEGO IMMUNOLOGICZNEGO
 Załącznik nr 2 SPOSÓB PRZEDSTAWIANIA DOKUMENTACJI DOŁĄCZANEJ DO WNIOSKU O DOPUSZCZENIE DO OBROTU PRODUKTU LECZNICZEGO WETERYNARYJNEGO IMMUNOLOGICZNEGO CZĘŚĆ I - STRESZCZENIE DOKUMENTACJI I A IB I B 1 I
Załącznik nr 2 SPOSÓB PRZEDSTAWIANIA DOKUMENTACJI DOŁĄCZANEJ DO WNIOSKU O DOPUSZCZENIE DO OBROTU PRODUKTU LECZNICZEGO WETERYNARYJNEGO IMMUNOLOGICZNEGO CZĘŚĆ I - STRESZCZENIE DOKUMENTACJI I A IB I B 1 I
mikrosatelitarne, minisatelitarne i polimorfizm liczby kopii
 Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
LEKI CHEMICZNE A LEKI BIOLOGICZNE
 LEKI CHEMICZNE A LEKI BIOLOGICZNE PRODUKT LECZNICZY - DEFINICJA Art. 2 pkt.32 Ustawy - Prawo farmaceutyczne Substancja lub mieszanina substancji, przedstawiana jako posiadająca właściwości: zapobiegania
LEKI CHEMICZNE A LEKI BIOLOGICZNE PRODUKT LECZNICZY - DEFINICJA Art. 2 pkt.32 Ustawy - Prawo farmaceutyczne Substancja lub mieszanina substancji, przedstawiana jako posiadająca właściwości: zapobiegania
Sylabus Biologia molekularna
 Sylabus Biologia molekularna 1. Metryczka Nazwa Wydziału Program kształcenia Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Analityka Medyczna, studia jednolite magisterskie, studia stacjonarne
Sylabus Biologia molekularna 1. Metryczka Nazwa Wydziału Program kształcenia Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Analityka Medyczna, studia jednolite magisterskie, studia stacjonarne
Prezentuje: Magdalena Jasińska
 Prezentuje: Magdalena Jasińska W którym momencie w rozwoju embrionalnym myszy rozpoczyna się endogenna transkrypcja? Hipoteza I: Endogenna transkrypcja rozpoczyna się w embrionach będących w stadium 2-komórkowym
Prezentuje: Magdalena Jasińska W którym momencie w rozwoju embrionalnym myszy rozpoczyna się endogenna transkrypcja? Hipoteza I: Endogenna transkrypcja rozpoczyna się w embrionach będących w stadium 2-komórkowym
Biologia Molekularna z Biotechnologią ===============================================================================================
 Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
Zagrożenia i ochrona przyrody
 Wymagania podstawowe Uczeń: Wymagania ponadpodstawowe Uczeń: Zagrożenia i ochrona przyrody wskazuje zagrożenia atmosfery powstałe w wyniku działalności człowieka, omawia wpływ zanieczyszczeń atmosfery
Wymagania podstawowe Uczeń: Wymagania ponadpodstawowe Uczeń: Zagrożenia i ochrona przyrody wskazuje zagrożenia atmosfery powstałe w wyniku działalności człowieka, omawia wpływ zanieczyszczeń atmosfery
Scenariusz lekcji z biologii w szkole ponadgimnazjalnej
 Scenariusz lekcji z biologii w szkole ponadgimnazjalnej Temat lekcji: Planowanie doświadczeń biologicznych jak prawidłowo zaplanować próbę kontrolną? Cele kształcenia IV etap edukacyjny: 1. Wymagania ogólne:
Scenariusz lekcji z biologii w szkole ponadgimnazjalnej Temat lekcji: Planowanie doświadczeń biologicznych jak prawidłowo zaplanować próbę kontrolną? Cele kształcenia IV etap edukacyjny: 1. Wymagania ogólne:
WNIOSEK O WYDANIE ZGODY NA ZAMKNIĘTE UŻYCIE GMO
 WNIOSEK O WYDANIE ZGODY NA ZAMKNIĘTE UŻYCIE GMO 1. Informacje o użytkowniku GMO i osobach odpowiedzialnych za realizację planowanego zamkniętego użycia GMO 1.1 (*) Nazwa i siedziba użytkownika lub imię,
WNIOSEK O WYDANIE ZGODY NA ZAMKNIĘTE UŻYCIE GMO 1. Informacje o użytkowniku GMO i osobach odpowiedzialnych za realizację planowanego zamkniętego użycia GMO 1.1 (*) Nazwa i siedziba użytkownika lub imię,
Metody analizy genomu
 Metody analizy genomu 1. Mapowanie restrykcyjne. 2. Sondy do rozpoznawania DNA 3. FISH 4. Odczytanie sekwencji DNA 5. Interpretacja sekwencji DNA genomu 6. Transkryptom 7. Proteom 1. Mapy restrykcyjne
Metody analizy genomu 1. Mapowanie restrykcyjne. 2. Sondy do rozpoznawania DNA 3. FISH 4. Odczytanie sekwencji DNA 5. Interpretacja sekwencji DNA genomu 6. Transkryptom 7. Proteom 1. Mapy restrykcyjne
S YL AB US MODUŁ U ( PRZEDMIOTU) I nforma c j e ogólne
 I nforma c j e ogólne") Załącznik Nr 3 do Uchwały Senatu PUM 14/2012 S YL AB US MODUŁ U ( PRZEDMIOTU) I nforma c j e ogólne Kod modułu Rodzaj modułu Wydział PUM Kierunek studiów Specjalność Poziom studiów Nazwa modułu INŻYNIERIA
Załącznik Nr 3 do Uchwały Senatu PUM 14/2012 S YL AB US MODUŁ U ( PRZEDMIOTU) I nforma c j e ogólne Kod modułu Rodzaj modułu Wydział PUM Kierunek studiów Specjalność Poziom studiów Nazwa modułu INŻYNIERIA
Patentowanie wynalazków z dziedziny biotechnologii
 chroń swoje pomysły 3 Kongres Świata Przemysłu Farmaceutycznego Poznań, 16-17 czerwca 2011 Patentowanie wynalazków z dziedziny biotechnologii dr Izabela Milczarek Czym jest biotechnologia? Biotechnologia,
chroń swoje pomysły 3 Kongres Świata Przemysłu Farmaceutycznego Poznań, 16-17 czerwca 2011 Patentowanie wynalazków z dziedziny biotechnologii dr Izabela Milczarek Czym jest biotechnologia? Biotechnologia,
Pytania Egzamin magisterski
 Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
Uczeń potrafi. Dział Rozdział Temat lekcji
 Plan wynikowy z biologii- zakres podstawowy, dla klasy III LO i III i IV Technikum LO im.ks. Jerzego Popiełuszki oraz Technikum w Suchowoli Nauczyciel: Katarzyna Kotiuk Nr programu: DKOS-4015-5/02 Dział
Plan wynikowy z biologii- zakres podstawowy, dla klasy III LO i III i IV Technikum LO im.ks. Jerzego Popiełuszki oraz Technikum w Suchowoli Nauczyciel: Katarzyna Kotiuk Nr programu: DKOS-4015-5/02 Dział
TRANSKRYPCJA - I etap ekspresji genów
 Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
PL 204536 B1. Szczepanik Marian,Kraków,PL Selmaj Krzysztof,Łódź,PL 29.12.2003 BUP 26/03 29.01.2010 WUP 01/10
 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 204536 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 354698 (22) Data zgłoszenia: 24.06.2002 (51) Int.Cl. A61K 38/38 (2006.01)
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 204536 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 354698 (22) Data zgłoszenia: 24.06.2002 (51) Int.Cl. A61K 38/38 (2006.01)
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ 1. Gen to odcinek DNA odpowiedzialny
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ 1. Gen to odcinek DNA odpowiedzialny
Dr hab. Anna Bębenek Warszawa,
 Dr hab. Anna Bębenek Warszawa, 14.01. 2018 Instytut Biochemii i Biofizyki PAN Ul. Pawińskiego 5a 02-106 Warszawa Recenzja pracy doktorskiej Pana mgr Michała Płachty Pod Tytułem Regulacja funkcjonowania
Dr hab. Anna Bębenek Warszawa, 14.01. 2018 Instytut Biochemii i Biofizyki PAN Ul. Pawińskiego 5a 02-106 Warszawa Recenzja pracy doktorskiej Pana mgr Michała Płachty Pod Tytułem Regulacja funkcjonowania
Ćwiczenia 1 Wirtualne Klonowanie Prowadzący: mgr inż. Joanna Tymeck-Mulik i mgr Lidia Gaffke. Część teoretyczna:
 Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
Sylabus Biologia molekularna
 Sylabus Biologia molekularna 1. Metryczka Nazwa Wydziału Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Program kształcenia Farmacja, jednolite studia magisterskie, forma studiów: stacjonarne
Sylabus Biologia molekularna 1. Metryczka Nazwa Wydziału Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Program kształcenia Farmacja, jednolite studia magisterskie, forma studiów: stacjonarne
TATA box. Enhancery. CGCG ekson intron ekson intron ekson CZĘŚĆ KODUJĄCA GENU TERMINATOR. Elementy regulatorowe
 Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Hybrydyzacja kwasów nukleinowych
 Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg
 STRESZCZENIE Przewlekła białaczka limfocytowa (PBL) jest najczęstszą białaczką ludzi starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg kliniczny, zróżnicowane rokowanie. Etiologia
STRESZCZENIE Przewlekła białaczka limfocytowa (PBL) jest najczęstszą białaczką ludzi starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg kliniczny, zróżnicowane rokowanie. Etiologia
DNA musi współdziałać z białkami!
 DNA musi współdziałać z białkami! Specyficzność oddziaływań między DNA a białkami wiążącymi DNA zależy od: zmian konformacyjnych wzdłuż cząsteczki DNA zróżnicowania struktury DNA wynikającego z sekwencji
DNA musi współdziałać z białkami! Specyficzność oddziaływań między DNA a białkami wiążącymi DNA zależy od: zmian konformacyjnych wzdłuż cząsteczki DNA zróżnicowania struktury DNA wynikającego z sekwencji
TaqNova-RED. Polimeraza DNA RP20R, RP100R
 TaqNova-RED Polimeraza DNA RP20R, RP100R RP20R, RP100R TaqNova-RED Polimeraza DNA Rekombinowana termostabilna polimeraza DNA Taq zawierająca czerwony barwnik, izolowana z Thermus aquaticus, o przybliżonej
TaqNova-RED Polimeraza DNA RP20R, RP100R RP20R, RP100R TaqNova-RED Polimeraza DNA Rekombinowana termostabilna polimeraza DNA Taq zawierająca czerwony barwnik, izolowana z Thermus aquaticus, o przybliżonej
Mechanizm działania terapii fotodynamicznej w diagnozowaniu i leczeniu nowotworów. Anna Szczypka Aleksandra Tyrawska
 Mechanizm działania terapii fotodynamicznej w diagnozowaniu i leczeniu nowotworów Anna Szczypka Aleksandra Tyrawska Metody fotodynamiczne PDT Technika diagnostyczna i terapeutyczna zaliczana do form fotochemioterapii
Mechanizm działania terapii fotodynamicznej w diagnozowaniu i leczeniu nowotworów Anna Szczypka Aleksandra Tyrawska Metody fotodynamiczne PDT Technika diagnostyczna i terapeutyczna zaliczana do form fotochemioterapii
PL 196881 B1. Trójfazowy licznik indukcyjny do pomiaru nadwyżki energii biernej powyżej zadanego tg ϕ
 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 196881 (13) B1 (21) Numer zgłoszenia: 340516 (51) Int.Cl. G01R 11/40 (2006.01) G01R 21/00 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22)
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 196881 (13) B1 (21) Numer zgłoszenia: 340516 (51) Int.Cl. G01R 11/40 (2006.01) G01R 21/00 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22)
Informacje dotyczące pracy kontrolnej
 Informacje dotyczące pracy kontrolnej Słuchacze, którzy z przyczyn usprawiedliwionych nie przystąpili do pracy kontrolnej lub otrzymali z niej ocenę negatywną zobowiązani są do dnia 06 grudnia 2015 r.
Informacje dotyczące pracy kontrolnej Słuchacze, którzy z przyczyn usprawiedliwionych nie przystąpili do pracy kontrolnej lub otrzymali z niej ocenę negatywną zobowiązani są do dnia 06 grudnia 2015 r.
Składniki diety a stabilność struktury DNA
 Składniki diety a stabilność struktury DNA 1 DNA jedyna makrocząsteczka, której synteza jest ściśle kontrolowana, a powstałe błędy są naprawiane DNA jedyna makrocząsteczka naprawiana in vivo Replikacja
Składniki diety a stabilność struktury DNA 1 DNA jedyna makrocząsteczka, której synteza jest ściśle kontrolowana, a powstałe błędy są naprawiane DNA jedyna makrocząsteczka naprawiana in vivo Replikacja
[2ZPK/KII] Inżynieria genetyczna w kosmetologii
![[2ZPK/KII] Inżynieria genetyczna w kosmetologii](/thumbs/98/136385472.jpg "[2ZPK/KII] Inżynieria genetyczna w kosmetologii") [2ZPK/KII] Inżynieria genetyczna w kosmetologii 1. Ogólne informacje o module Nazwa modułu Kod modułu Nazwa jednostki prowadzącej modułu Nazwa kierunku studiów Forma studiów Profil kształcenia Semestr
[2ZPK/KII] Inżynieria genetyczna w kosmetologii 1. Ogólne informacje o module Nazwa modułu Kod modułu Nazwa jednostki prowadzącej modułu Nazwa kierunku studiów Forma studiów Profil kształcenia Semestr
Jaki koń jest nie każdy widzi - genomika populacji polskich ras koni
 Jaki koń jest nie każdy widzi - genomika populacji polskich ras koni Gurgul A., Jasielczuk I., Semik-Gurgul E., Pawlina-Tyszko K., Szmatoła T., Bugno-Poniewierska M. Instytut Zootechniki PIB Zakład Biologii
Jaki koń jest nie każdy widzi - genomika populacji polskich ras koni Gurgul A., Jasielczuk I., Semik-Gurgul E., Pawlina-Tyszko K., Szmatoła T., Bugno-Poniewierska M. Instytut Zootechniki PIB Zakład Biologii
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
Wybrane zastosowania metod inżynierii genetycznej
 Wybrane zastosowania metod inżynierii genetycznej Inżynieria genetyczna to inaczej ingerencja w materiał genetyczny organizmów, w celu zmiany ich właściwości dziedzicznych. Polega ona na wprowadzaniu do
Wybrane zastosowania metod inżynierii genetycznej Inżynieria genetyczna to inaczej ingerencja w materiał genetyczny organizmów, w celu zmiany ich właściwości dziedzicznych. Polega ona na wprowadzaniu do
Projektowanie reakcji multiplex- PCR
 Projektowanie reakcji multiplex- PCR Dzięki nowoczesnym, wydajnym zestawom do PCR, amplifikacja DNA nie jest już takim wyzwaniem, jak w latach 80-tych i 90-tych. Wraz z poprawą metodyki, pojawiły się ciekawe
Projektowanie reakcji multiplex- PCR Dzięki nowoczesnym, wydajnym zestawom do PCR, amplifikacja DNA nie jest już takim wyzwaniem, jak w latach 80-tych i 90-tych. Wraz z poprawą metodyki, pojawiły się ciekawe
Do moich badań wybrałam przede wszystkim linię kostniakomięsaka 143B ze względu na jej wysoki potencjał przerzutowania. Do wykonania pracy
 Streszczenie Choroby nowotworowe stanowią bardzo ważny problem zdrowotny na świecie. Dlatego, medycyna dąży do znalezienia nowych skutecznych leków, ale również rozwiązań do walki z nowotworami. Głównym
Streszczenie Choroby nowotworowe stanowią bardzo ważny problem zdrowotny na świecie. Dlatego, medycyna dąży do znalezienia nowych skutecznych leków, ale również rozwiązań do walki z nowotworami. Głównym
Aneks II. Wnioski naukowe
 Aneks II Wnioski naukowe 8 Wnioski naukowe Solu-Medrol 40 mg, proszek i rozpuszczalnik do sporządzania roztworu do wstrzykiwań (zwany dalej Solu-Medrol ) zawiera metyloprednisolon i (jako substancję pomocniczą)
Aneks II Wnioski naukowe 8 Wnioski naukowe Solu-Medrol 40 mg, proszek i rozpuszczalnik do sporządzania roztworu do wstrzykiwań (zwany dalej Solu-Medrol ) zawiera metyloprednisolon i (jako substancję pomocniczą)
Indukowane pluripotencjalne komórki macierzyste
 Indukowane pluripotencjalne komórki macierzyste Nagroda Nogla w dziedzinie medycyny i fizjologii z roku 2012 dla Brytyjczyka John B.Gurdon oraz Japooczyka Shinya Yamanaka Wykonały: Katarzyna Białek Katarzyna
Indukowane pluripotencjalne komórki macierzyste Nagroda Nogla w dziedzinie medycyny i fizjologii z roku 2012 dla Brytyjczyka John B.Gurdon oraz Japooczyka Shinya Yamanaka Wykonały: Katarzyna Białek Katarzyna
Geny, a funkcjonowanie organizmu
 Geny, a funkcjonowanie organizmu Wprowadzenie do genów letalnych Geny kodują Białka Kwasy rybonukleinowe 1 Geny Występują zwykle w 2 kopiach Kopia pochodząca od matki Kopia pochodząca od ojca Ekspresji
Geny, a funkcjonowanie organizmu Wprowadzenie do genów letalnych Geny kodują Białka Kwasy rybonukleinowe 1 Geny Występują zwykle w 2 kopiach Kopia pochodząca od matki Kopia pochodząca od ojca Ekspresji
WYBRANE SKŁADNIKI POKARMOWE A GENY
 WYBRANE SKŁADNIKI POKARMOWE A GENY d r i n ż. Magdalena Górnicka Zakład Oceny Żywienia Katedra Żywienia Człowieka WitaminyA, E i C oraz karotenoidy Selen Flawonoidy AKRYLOAMID Powstaje podczas przetwarzania
WYBRANE SKŁADNIKI POKARMOWE A GENY d r i n ż. Magdalena Górnicka Zakład Oceny Żywienia Katedra Żywienia Człowieka WitaminyA, E i C oraz karotenoidy Selen Flawonoidy AKRYLOAMID Powstaje podczas przetwarzania
(12) OPIS PATENTOWY (19) PL (11)
 OPIS PATENTOWY (19) PL (11)") RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 188998 (21 ) Numer zgłoszenia: 333174 (22) Data zgłoszenia: 23.10.1997 (86) Data i numer zgłoszenia międzynarodowego:
RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 188998 (21 ) Numer zgłoszenia: 333174 (22) Data zgłoszenia: 23.10.1997 (86) Data i numer zgłoszenia międzynarodowego:
Potrzeby instytutów badawczych w zakresie wsparcia procesu rozwoju nowych leków, terapii, urządzeń medycznych... OŚRODEK TRANSFERU TECHNOLOGII
 Potrzeby instytutów badawczych w zakresie wsparcia procesu rozwoju nowych leków, terapii, urządzeń medycznych... DR ADAM SOBCZAK OŚRODEK TRANSFERU TECHNOLOGII BIO&TECHNOLOGY INNOVATIONS PLATFORM POLCRO
Potrzeby instytutów badawczych w zakresie wsparcia procesu rozwoju nowych leków, terapii, urządzeń medycznych... DR ADAM SOBCZAK OŚRODEK TRANSFERU TECHNOLOGII BIO&TECHNOLOGY INNOVATIONS PLATFORM POLCRO
Ćwiczenie 3. Amplifikacja genu ccr5 Homo sapiens wykrywanie delecji Δ32pz warunkującej oporność na wirusa HIV
 Ćwiczenie 3. Amplifikacja genu ccr5 Homo sapiens wykrywanie delecji Δ32pz warunkującej oporność na wirusa HIV Cel ćwiczenia Określenie podatności na zakażenie wirusem HIV poprzez detekcję homo lub heterozygotyczności
Ćwiczenie 3. Amplifikacja genu ccr5 Homo sapiens wykrywanie delecji Δ32pz warunkującej oporność na wirusa HIV Cel ćwiczenia Określenie podatności na zakażenie wirusem HIV poprzez detekcję homo lub heterozygotyczności
Rozkład materiału z biologii dla klasy III AD. 7 godz / tyg rok szkolny 2016/17
 Rozkład materiału z biologii dla klasy III AD zakres rozszerzony LO 7 godz / tyg rok szkolny 2016/17 Biologia na czasie 2 zakres rozszerzony nr dopuszczenia 564/2/2012 Biologia na czasie 3 zakres rozszerzony
Rozkład materiału z biologii dla klasy III AD zakres rozszerzony LO 7 godz / tyg rok szkolny 2016/17 Biologia na czasie 2 zakres rozszerzony nr dopuszczenia 564/2/2012 Biologia na czasie 3 zakres rozszerzony
Przykładowe zadania. przygotowujące do egzaminu maturalnego
 Przykładowe zadania z BIOLOGii przygotowujące do egzaminu maturalnego Prezentowane zadania są zgodne z podstawą programową kształcenia ogólnego w zakresie rozszerzonym i podstawowym. Prócz zadań, które
Przykładowe zadania z BIOLOGii przygotowujące do egzaminu maturalnego Prezentowane zadania są zgodne z podstawą programową kształcenia ogólnego w zakresie rozszerzonym i podstawowym. Prócz zadań, które
ANEKS I. Strona 1 z 5
 ANEKS I WYKAZ NAZW, POSTAĆ FARMACEUTYCZNA, MOC WETERYNARYJNYCH PRODUKTÓW LECZNICZYCH, GATUNKI ZWIERZĄT, DROGA PODANIA, PODMIOT ODPOWIEDZIALNY POSIADAJĄCY POZWOLENIE NA DOPUSZCZENIE DO OBROTU W PAŃSTWACH
ANEKS I WYKAZ NAZW, POSTAĆ FARMACEUTYCZNA, MOC WETERYNARYJNYCH PRODUKTÓW LECZNICZYCH, GATUNKI ZWIERZĄT, DROGA PODANIA, PODMIOT ODPOWIEDZIALNY POSIADAJĄCY POZWOLENIE NA DOPUSZCZENIE DO OBROTU W PAŃSTWACH
Stosowanie metod alternatywnych dla badań na zwierzętach do celów rozporządzenia REACH
 Nr ref.: ECHA-11-FS-06-PL ISBN-13: 978-92-9217-610-5 Stosowanie metod alternatywnych dla badań na zwierzętach do celów rozporządzenia Jednym z głównych powodów opracowania i przyjęcia rozporządzenia był
Nr ref.: ECHA-11-FS-06-PL ISBN-13: 978-92-9217-610-5 Stosowanie metod alternatywnych dla badań na zwierzętach do celów rozporządzenia Jednym z głównych powodów opracowania i przyjęcia rozporządzenia był
Inżynieria genetyczna
 Inżynieria genetyczna i technologia rekombinowanego DNA Dr n. biol. Urszula Wasik Zakład Biologii Medycznej Inżynieria genetyczna świadoma, celowa, kontrolowana ingerencja w materiał genetyczny organizmów
Inżynieria genetyczna i technologia rekombinowanego DNA Dr n. biol. Urszula Wasik Zakład Biologii Medycznej Inżynieria genetyczna świadoma, celowa, kontrolowana ingerencja w materiał genetyczny organizmów
2. Enzymy pozwalające na manipulację DNA a. Polimerazy DNA b. Nukleazy c. Ligazy
 Metody analizy DNA 1. Budowa DNA. 2. Enzymy pozwalające na manipulację DNA a. Polimerazy DNA b. Nukleazy c. Ligazy 3. Klonowanie in vivo a. w bakteriach, wektory plazmidowe b. w fagach, kosmidy c. w drożdżach,
Metody analizy DNA 1. Budowa DNA. 2. Enzymy pozwalające na manipulację DNA a. Polimerazy DNA b. Nukleazy c. Ligazy 3. Klonowanie in vivo a. w bakteriach, wektory plazmidowe b. w fagach, kosmidy c. w drożdżach,
GIMNAZJUM SPRAWDZIANY SUKCES W NAUCE
 GIMNAZJUM SPRAWDZIANY BIOLOGIA klasa III SUKCES W NAUCE II GENETYKA CZŁOWIEKA Zadanie 1. Cechy organizmu są warunkowane przez allele dominujące i recesywne. Uzupełnij tabelę, wykorzystując poniższe określenia,
GIMNAZJUM SPRAWDZIANY BIOLOGIA klasa III SUKCES W NAUCE II GENETYKA CZŁOWIEKA Zadanie 1. Cechy organizmu są warunkowane przez allele dominujące i recesywne. Uzupełnij tabelę, wykorzystując poniższe określenia,
Ocena ekspresji genu ABCG2 i białka oporności raka piersi (BCRP) jako potencjalnych czynników prognostycznych w raku jelita grubego
 jako potencjalnych czynników prognostycznych w raku jelita grubego") Aleksandra Sałagacka Ocena ekspresji genu ABCG2 i białka oporności raka piersi (BCRP) jako potencjalnych czynników prognostycznych w raku jelita grubego Pracownia Biologii Molekularnej i Farmakogenomiki
Aleksandra Sałagacka Ocena ekspresji genu ABCG2 i białka oporności raka piersi (BCRP) jako potencjalnych czynników prognostycznych w raku jelita grubego Pracownia Biologii Molekularnej i Farmakogenomiki
Konkurs szkolny Mistrz genetyki etap II
 onkurs szkolny istrz genetyki etap II 1.W D pewnego pierwotniaka tymina stanowi 28 % wszystkich zasad azotowych. blicz i zapisz, jaka jest zawartość procentowa każdej z pozostałych zasad w D tego pierwotniaka.
onkurs szkolny istrz genetyki etap II 1.W D pewnego pierwotniaka tymina stanowi 28 % wszystkich zasad azotowych. blicz i zapisz, jaka jest zawartość procentowa każdej z pozostałych zasad w D tego pierwotniaka.
FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach
 FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach FOCUS Plus to dodatek dostępny dla standardowych pasz tuczowych BioMaru, dostosowany specjalnie do potrzeb ryb narażonych na trudne
FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach FOCUS Plus to dodatek dostępny dla standardowych pasz tuczowych BioMaru, dostosowany specjalnie do potrzeb ryb narażonych na trudne
Bloki licencjackie i studia magisterskie na Kierunkach: Biotechnologia, specjalność Biotechnologia roślinna oraz Genetyka
 Bloki licencjackie i studia magisterskie na Kierunkach: Biotechnologia, specjalność Biotechnologia roślinna oraz Genetyka INSTYTUT BIOLOGII EKSPERYMENTALNEJ W Katedrze Genetyki Ogólnej, Biologii Molekularnej
Bloki licencjackie i studia magisterskie na Kierunkach: Biotechnologia, specjalność Biotechnologia roślinna oraz Genetyka INSTYTUT BIOLOGII EKSPERYMENTALNEJ W Katedrze Genetyki Ogólnej, Biologii Molekularnej
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii
 Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
Ocena. wykonanej pod kierunkiem prof. dr hab. med. Małgorzaty Polz-Docewicz
 UNIWERSYTET MEDYCZNY IM. KAROLA MARCINKOWSKIEGO W POZNANIU KATEDRA I ZAKŁAD MIKROBIOLOGII LEKARSKIEJ Kierownik: prof. dr hab. Andrzej Szkaradkiewicz ul. Wieniawskiego 3 tel. 61 8546 138 61-712 Poznań fax
UNIWERSYTET MEDYCZNY IM. KAROLA MARCINKOWSKIEGO W POZNANIU KATEDRA I ZAKŁAD MIKROBIOLOGII LEKARSKIEJ Kierownik: prof. dr hab. Andrzej Szkaradkiewicz ul. Wieniawskiego 3 tel. 61 8546 138 61-712 Poznań fax
PL B1 (12) O P I S P A T E N T O W Y (19) P L (11) (13) B 1 A61K 9/20. (22) Data zgłoszenia:
 O P I S P A T E N T O W Y (19) P L (11) (13) B 1 A61K 9/20. (22) Data zgłoszenia:") R Z E C Z PO SPO L IT A PO LSK A (12) O P I S P A T E N T O W Y (19) P L (11) 1 7 7 6 0 7 (21) Numer zgłoszenia: 316196 (13) B 1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 13.03.1995
R Z E C Z PO SPO L IT A PO LSK A (12) O P I S P A T E N T O W Y (19) P L (11) 1 7 7 6 0 7 (21) Numer zgłoszenia: 316196 (13) B 1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 13.03.1995
Rośliny modyfikowane genetycznie (GMO)
") Rośliny modyfikowane genetycznie (GMO) Organizmy modyfikowane genetycznie Organizm zmodyfikowany genetycznie (międzynarodowy skrót: GMO Genetically Modified Organizm) to organizm o zmienionych cechach,
Rośliny modyfikowane genetycznie (GMO) Organizmy modyfikowane genetycznie Organizm zmodyfikowany genetycznie (międzynarodowy skrót: GMO Genetically Modified Organizm) to organizm o zmienionych cechach,
Model Marczuka przebiegu infekcji.
 Model Marczuka przebiegu infekcji. Karolina Szymaniuk 27 maja 2013 Karolina Szymaniuk () Model Marczuka przebiegu infekcji. 27 maja 2013 1 / 17 Substrat Związek chemiczny, który ulega przemianie w wyniku
Model Marczuka przebiegu infekcji. Karolina Szymaniuk 27 maja 2013 Karolina Szymaniuk () Model Marczuka przebiegu infekcji. 27 maja 2013 1 / 17 Substrat Związek chemiczny, który ulega przemianie w wyniku
Lek od pomysłu do wdrożenia
 Lek od pomysłu do wdrożenia Lek od pomysłu do wdrożenia KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU
Lek od pomysłu do wdrożenia Lek od pomysłu do wdrożenia KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU
SYLABUS. Wydział Biologiczno-Rolniczy. Katedra Biochemii i Biologii Komórki
 SYLABUS 1.1. PODSTAWOWE INFORMACJE O PRZEDMIOCIE/MODULE Nazwa przedmiotu/ modułu Biologia molekularna z elementami inżynierii genetycznej Kod przedmiotu/ modułu* Wydział (nazwa jednostki prowadzącej kierunek)
SYLABUS 1.1. PODSTAWOWE INFORMACJE O PRZEDMIOCIE/MODULE Nazwa przedmiotu/ modułu Biologia molekularna z elementami inżynierii genetycznej Kod przedmiotu/ modułu* Wydział (nazwa jednostki prowadzącej kierunek)