(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
|
|
|
- Elżbieta Pietrzyk
- 8 lat temu
- Przeglądów:
Transkrypt
1 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: (97) O udzieleniu patentu europejskiego ogłoszono: Europejski Biuletyn Patentowy 2010/46 EP B1 (13) (51) T3 Int.Cl. C12N 15/86 ( ) C12N 15/861 ( ) A61K 48/00 ( ) (54) Tytuł wynalazku: Hybrydowy wektor adenowirusowy/alfawirusowy do skutecznego podawania i ekspresji genów terapeutycznych w komórkach nowotworowych (30) Pierwszeństwo: ES (43) Zgłoszenie ogłoszono: w Europejskim Biuletynie Patentowym nr 2007/10 (45) O złożeniu tłumaczenia patentu ogłoszono: Wiadomości Urzędu Patentowego 2011/04 (73) Uprawniony z patentu: Proyecto de Biomedicina Cima, S.L., Pamplona, ES (72) Twórca(y) wynalazku: PL/EP T3 CHENG QIAN, Pamplona, ES MIN GUAN, Pamplona, ES CRISTIAN SMERDOU PICAZO, Pamplona, ES JESUS PRIETO VALTUEÑA, Pamplona, ES (74) Pełnomocnik: rzecz. pat. Zofia Sulima SULIMA-GRABOWSKA-SIERZPUTOWSKA BIURO PATENTÓW I ZNAKÓW TOWAROWYCH SP.J. Skr. poczt Warszawa 10 Uwaga: W ciągu dziewięciu miesięcy od publikacji informacji o udzieleniu patentu europejskiego, każda osoba może wnieść do Europejskiego Urzędu Patentowego sprzeciw dotyczący udzielonego patentu europejskiego. Sprzeciw wnosi się w formie uzasadnionego na piśmie oświadczenia. Uważa się go za wniesiony dopiero z chwilą wniesienia opłaty za sprzeciw (Art. 99 (1) Konwencji o udzielaniu patentów europejskich).
 Zgłoszenie ogłoszono: 07.03.2007 w Europejskim Biuletynie Patentowym nr 2007/10 (45) O złożeniu tłumaczenia patentu ogłoszono: 29.04.2011 Wiadomości Urzędu Patentowego 2011/04 (73) Uprawniony z patentu: Proyecto de Biomedicina Cima, S.")
2 SGS-1411/VAL EP B1 Opis TECHNICZNA DZIEDZINA WYNALAZKU [0001] Niniejszy wynalazek dotyczy wektorów do ekspresji genów pochodzących od adenowirusów, do otrzymywania produktów terapeutycznych. STAN TECHNIKI DLA WYNALAZKU [0002] Jednym z głównych problemów napotykanych w standardowej terapii raka jest brak specyficzności względem nowotworu, która to sytuacja często prowadzi do powaŝnych działań ubocznych i ogranicza stosowalną dawkę terapeutyczną. Pomimo, Ŝe terapia genowa jest wysoce obiecująca w terapii raka, napotyka ona na specyficzny problem: skierowanie ekspresji transgenicznej do miejsca nowotworu. W kilku badaniach sugerowano, Ŝe gdy wektory wirusowe są podawane do miejsca zmienionego patologicznie, pomimo, Ŝe ekspresja transgenu jest zasadniczo ograniczona do regionu przylegającego do toru igły, taka ekspresja moŝe takŝe wystąpić w innych tkankach. W konsekwencji istotnym celem w terapii genowej stosowanej w raku jest skierowanie ekspresji genu terapeutycznego do nowotworów przez specyficzne podawanie do tkanki nowotworowej ( cel tkankowy ) i/lub specyficzną aktywację ( cel transkrypcyjny ) w tkance nowotworowej, bez wpływu na zdrowe komórki. Cel tkankowy moŝna osiągnąć przez wytworzenie wektora skierowanego przez modyfikacje oddziaływań receptor-ligand, umoŝliwiając w ten sposób zaka- Ŝanie komórek eksprymujących specyficzny receptor. Cel transkrypcyjny moŝna osiągnąć przez zastosowanie do kontrolowania ekspresji transgenu promotora specyficznego dla nowotworu. W dotychczasowych badaniach zastosowano róŝne promotory specyficzne dla nowotworu. JednakŜe mają one zasadnicze wady związane z tym, Ŝe nie pozwalają uzyskać wysokich poziomów ekspresji genu, tak Ŝe ich aktywność przeciwnowotworowa jest ograniczona. [0003] Pracę przeglądową na temat najnowszych osiągnięć w opracowywaniu wektorów wirusowych do terapii moŝna znaleźć w Lundstrom K. Latest development in viral vectors for gene therapy ; Trends in Biotechnology, 2003, 21: [0004] Obecnie stosowane wektory wirusowe obejmują alfawirusy. Alfawirusy są wirusami otoczkowymi zawierającymi jako genom prostą nić RNA o polarności dodatniej. Zaprojektowano i opracowano wektory ekspresyjne pochodzące od alfawirusów wirusa Sindbis (SIN), wirusa Semliki Forest (SFV) i wirusa wenezuelskiego końskiego zapalenia mó-

3 zgu (VEE). Wektory alfawirusowe są oparte na zastosowaniu replikujących się autonomicznie cząsteczek RNA pochodzących z genomów alfawirusowych, w których zachowano sekwencje 5' i 3' niezbędne do replikacji oraz gen replikazy (Rep), podczas gdy geny kodujące wirusowe białka strukturalne wydeletowano i zastąpiono transgenem. Po transfekcji komórek tymi wektorami Rep ulega translacji i wektor RNA jest kopiowany do nici RNA o polarności ujemnej, która będzie wykorzystywana jako matryca do amplifikacji wektora RNA. Rep moŝe takŝe rozpoznawać promotor subgenomowy w nici RNA polarności ujemnej, z którego z kolei syntetyzowany jest mniejszy subgenomowy segment RNA, który moŝe ulegać translacji z wytworzeniem istotnych poziomów heterologicznych białek. Wektory wirusowe moŝna stosować bezpośrednio jako RNA do transkrypcji in vitro z promotora prokariotycznego, takiego jak SP6 lub T7 lub jako DNA, gdy sekwencja replikonu jest umiejscowiona pod promotorem eukariotycznym, takim jak CMV. Wektor RNA moŝe zostać zapakowany w cząstkach wirusa przez jego kotransfekcję do komórek razem z jednym lub większą liczbą pomocniczych RNA, które kodują wirusowe białka strukturalne. Wektory alfawirusowe wykazują szereg właściwości, które czynią je atrakcyjnymi dla terapii genowej: bardzo rozległy tropizm, niską immunogenność i wysoki poziom ekspresji białek heterologicznych. Taka ekspresja jest jednakŝe przejściowa ze względu na wywoływanie apoptozy w komórkach pod wpływem wystąpienia replikacji. W dokumencie Rayner J.O., Dryga S.A., Kamrud K.I. Alphavirus vectors and vaccination ; Rev. Med. Virol. 2002; opisano opracowywanie wektorów ekspresyjnych opartych na alfawirusach do zastosowania w dziedzinie szczepionek. [0005] Inna seria wektorów wirusowych jest oparta na adenowirusach. Istnieje obszerna literatura dotycząca zastosowania adenowirusów, które zostały opracowane do przezwycię- Ŝenia niektórych niedogodności terapii genowej oraz jako źródło do wytworzenia wektorów ekspresyjnych. Dokument opisujący najnowsze postępy w dziedzinie wektorów adenowirusowych został opublikowany przez Volpers C, Kochanek S. Adenoviral vectors for gene transfer and therapy ; J Gene. Med. 2004; 6: S164-S171. Adenowirusy wykazują zaletę osiagania wysokiego poziomu wydajności transdukcji oraz zdolności do utrzymywania się w postaci episomalnej. JednakŜe ekspresja białek adenowirusowych wywołuje silne odpowiedzi immunologiczne, które ograniczają czas trwania ekspresji transgenicznej oraz wywołują toksyczność w komórkach zakaŝonych wektorem. W celu przezwycięŝenia tych problemów opracowano adenowirusy typu gutless. Te adenowirusy typu gutless pozbawiono wszystkich odpowiednich genów adenowirusowych (przy czym jedynymi zachowanymi sekwencjami były dwa odwrócone powtórzenia końcowe oraz sygnały pakowania), a w konsekwencji transdukowane komórki nie eksprymują produktu adenowirusowego i nie

4 wywołują odpowiedzi immunologicznej na wektor. W skrócie usunięcie wszystkich genów adenowirusowych pozostawia wystarczająco miejsca do przyjęcia duŝych kaset ekspresyjnych i z tego powodu adenowirusy typu gutless są takŝe nazywane wektorami adenowirusowymi o duŝej pojemności. Dokument opisujący konkretne aspekty wektorów adenowirusowych, związane z delecją wszystkich sekwencji kodujących białka wirusowe, został opublikowany przez Morsy MA i in. An adenoviral vector deleted for all viral coding sequences results in enhanced safety and extended expression of a leptin transgene ; Proc. Natl. Acad. Sci. USA 1998, 95: W dokumencie opublikowanym przez Schiedner G i in. Variables affecting in vivo performance of high-capacity adenovirus vectors"; J. Virol. 2002, 76: opisano zastosowanie wypychającego DNA w wektorach ekspresyjnych opartych na adenowirusach typu gutless wykazując, Ŝe obecność takiego wypychającego DNA jest zasadnicza dla uzyskania znaczącego wzrostu ekspresji genu, oraz Ŝe ogólnie zaprojektowanie wektorów opartych na wektorach adenowirusowych o du- Ŝej pojemności (typu gutless) moŝe zasadniczo zmodyfikować stopień i czas trwania ekspresji genu. [0006] Z drugiej strony w dokumencie US opisano wektor do transferu genów oparty na adenowirusach, który zawiera sekwencje odwróconego powtórzenia końcowego (ITR), co najmniej jedną sekwencję sygnałową pakowania oraz gen adenowirusowy VAI i/lub gen adenowirusowy VAII; a takŝe zawiera gen obcy dla adenowirusa funkcjonalnie związany z promotorem funkcjonalnym w komórkach docelowych adenowirusa. [0007] W okumencie US opisano wektor do transferu genów zawierający sekwencje odwróconego powtórzenia końcowego (ITR) adenowirusa oraz rekombinowane cząstki adenowirusa zawierające te sekwencje. [0008] W dokumencie US opisano wektory cdna pochodzące od alfawirusów, które zawierają DNA komplementarny do co najmniej części RNA alfawirusa istotnego dla replikacji alfawirusa oraz heterologiczny cdna, przykładowo cdna kodujący wymaganą substancję. Substancją tą moŝe być biologicznie aktywne białko lub polipeptyd, a takŝe immunogenne lub antygenowe białko lub polipeptyd, terapeutycznie aktywne białko lub polipeptyd lub terapeutycznie aktywny RNA. [0009] Celem niniejszego wynalazku jest poprawienie ekspresji transgenicznej oraz wywoływania apoptozy w komórkach nowotworowych przez wektory hybrydowe in vitro i in vivo. Dodatkowym celem jest poprawa skuteczności terapii przeciwnowotworowej w modelach zwierzęcych przez wektory hybrydowe. [0010] Dodatkowym celem jest takŝe opracowanie metody terapii genowej, w szczególności do leczenia raka przez zastosowanie wektorów hybrydowych.

5 4 [0011] Cele niniejszego wynalazku osiąga się przez połączenie poniŝszych cech w pojedynczym wektorze: 5 wysokiej zdolności zakaŝania, przez zastosowanie systemu uwalniania adenowirusa, wysokiej ekspresji transgenicznej oraz wywoływania apoptozy przez zastosowanie wektora pochodzącego od alfawirusa, takiego jak SFV, specyficzności względem nowotworu przez zastosowanie promotora specyficznego dla nowotworu. OPIS WYNALAZKU [0012] Po pierwsze wynalazek dotyczy adenowirusowego wektora hybrydowego do ekspresji genów, który zawiera co najmniej następujące elementy ustawione w kierunku 5' do 3': i. pierwszy łańcuch pochodzenia adenowirusowego, zawierający sekwencję pierwszego odwróconego powtórzenia końcowego (ITR) oraz sekwencję sygnałową pakowania adenowirusa; ii. pierwszą niekodującą sekwencję wypychającą; iii. sekwencję odpowiadającą promotorowi specyficznemu dla tkanki; iv. łańcuch cdna pochodzący od alfawirusa, którego sekwencja jest częściowo komplementarna do RNA alfawirusa, zawierający co najmniej sekwencję kodującą co najmniej jeden interesujący gen egzogenny, przy czym wspomniany łańcuch cdna zawiera: a. sekwencję 5' niezbędną do replikacji alfawirusa, b. sekwencję kodującą białka niestrukturalne wymagane do replikacji RNA alfawirusa, c. co najmniej jeden promotor subgenomowy alfawirusa, oraz d. sekwencję 3' niezbędną do replikacji alfawirusa; v. sekwencję poliadenylacji; oraz vi. drugą sekwencję odwróconego powtórzenia końcowego (ITR) adenowirusa. [0013] W szczególności wynalazek dotyczy konstrukcji adenowirusowego wektora hybrydowego zawierającego jako element iv łańcuch cdna pochodzący od alfawirusa, odpowiadający sekwencji rekombinowanego replikonu SFV, pod kontrolą transkrypcyjną promotora specyficznego dla nowotworu (element iii.), który jest promotorem alfafetoproteiny (AFP). W tym konstrukcie transgen moŝe być wstawiony do replikonu SFV, pod kontrolą subgenomowego promotora SFV. Po zakaŝeniu komórek nowotworowych tym wektorem hybrydowym mrna replikonu SFV ulega transkrypcji z promotora specy-

6 ficznego dla nowotworu oraz białka niestrukturalne, nsps, do których naleŝy gen replikazy SFV ulegają translacji ze wspomnianego mrna replikonu SFV. Te białka nsps replikaza wirusowa rozpoczynają replikację mrna replikonu SFV z wytworzeniem subgenomowego RNA SFV. W konsekwencji transgen moŝe być eksprymowany na wysokim poziomie z subgenomowego RNA SFV. Ten globalny proces replikacji wirusa spowoduje wytworzenie apoptozy w zakaŝonych komórkach. W sytuacji gdy ten wektor hybrydowy zakazi komórki nienowotworowe mrna replikonu SFV nie będzie transkrybowany z promotora specyficznego dla nowotworu, który nie będzie aktywny w tych komórkach. Zatem brak będzie ekspresji transgenu oraz apoptoza nie wystąpi w normalnych komórkach zakaŝonych wektorem hybrydowym. [0014] Niniejszy wynalazek dalej dotyczy sposobu uzyskiwania wspomnianego adenowirusowego wektora hybrydowego, obejmującego składanie zdefiniowanych powyŝej elementów od i. do vi. adenowirusowego wektora hybrydowego przez zastosowanie technik inŝynierii genetycznej. [0015] Niniejszy wynalazek dotyczy ponadto zastosowania wspomnianego wektora hybrydowego do przenoszenia materiału genetycznego do komórki, a w szczególności do wprowadzania i ekspresji obcych genów w komórkach eukariotycznych, które mogą być komórkami docelowymi dla adenowirusa. [0016] Przeniesienie materiału genetycznego korzystnie wywołuje indukcję odpowiedzi immunologicznej przeciw obcym antygenom we wspomnianej komórce. [0017] Niniejszy wynalazek takŝe dotyczy kompozycji farmaceutycznej zawierającej wspomniany adenowirusowo-alfawirusowy hybrydowy wektor oraz jego zastosowania w leczeniu terapeutycznym nowotworu, obejmującym podawanie pacjentowi wspomnianej kompozycji farmaceutycznej. [0018] Niniejszy wynalazek dotyczy takŝe sposobu leczenia nowotworu przez zastosowanie poprzednio zdefiniowanego wektora hybrydowego, obejmującego podawanie pacjentowi wspomnianego wektora hybrydowego. [0019] Niniejszy wynalazek dotyczy adenowirusowego wektora hybrydowego do ekspresji genów, który zawiera co najmniej następujące elementy, ustawione w kierunku 5' do 3': i. pierwszy łańcuch pochodzenia adenowirusowego zawierający sekwencję pierwszego odwróconego powtórzenia końcowego (ITR) oraz sekwencję sygnałową pakowania adenowirusa; ii. pierwszą niekodującą sekwencję wypychającą; iii. sekwencję odpowiadającą promotorowi specyficznemu dla tkanki; iv. łańcuch cdna pochodzący od alfawirusa, którego sekwencja jest częściowo

7 komplementarna do sekwencji RNA alfawirusa, zawierający co najmniej sekwencję kodującą co najmniej jeden interesujący gen egzogenny, przy czym wspomniany łańcuch cdna zawiera: a. sekwencję 5' niezbędną do replikacji alfawirusa, b. sekwencję kodującą białka niestrukturalne wymagane do replikacji RNA alfawirusa, c. co najmniej jeden promotor subgenomowy alfawirusa, oraz d. sekwencję 3' niezbędną do replikacji alfawirusa; v. sekwencję poliadenylacji; oraz vi. drugą sekwencję odwróconego powtórzenia końcowego (ITR) adenowirusa. [0020] Natura elementu i. w alfawirusowo-adenowirusowym wektorze hybrydowym według niniejszego wynalazku, tj. natura łańcucha pochodzenia adenowirusowego zawierającego sekwencję pierwszego odwróconego powtórzenia końcowego (ITR) oraz sekwencję sygnałową pakowania (ψ) adenowirusów nie jest krytycznym aspektem niniejszego wynalazku i moŝe on pochodzić z jakiegokolwiek serotypu adenowirusa. Wspomniane serotypy są dobrze znane w dziedzinie wynalazku obejmują, przykładowo, Ad12 (podrodzaj A), Ad3 i Ad7 (podrodzaj B), Ad2 i Ad5 (podrodzaj C), Ad8 (podrodzaj D), Ad4 (podrodzaj E), Ad40 (podrodzaj F) oraz inne znane nie-ludzkie adenowirusy, które mogą pochodzić od gatunków, takich jak świnie, owce, krowy i ptaki. Zatem ta sekwencja pierwszego odwróconego powtórzenia końcowego, która moŝe mieć długość w przybliŝeniu pomiędzy 100 a 500 pz, moŝe róŝnić się zaleŝnie od stosowanego serotypu adenowirusa. W ten sam sposób moŝe róŝnić się sekwencja sygnałowa pakowania adenowirusa zaleŝnie od zastosowanego serotypu adenowirusa. [0021] Według szczególnie korzystnej postaci wynalazku wspomniany wektor adenowirusowy do ekspresji genów zawiera element i. o sekwencji SEQ ID No 1 lub jakiejkolwiek innej sekwencji wykazującej homologię z SEQ ID No 1 wystarczającą do spełnienia tej funkcji. [0022] Natura elementu ii. w adenowiruowym wektorze hybrydowym według niniejszego wynalazku nie jest jego krytycznym aspektem. Wspomniany element ii., którego funkcją jest zwiększenie całkowitej wielkości konstruktu, moŝe być jakąkolwiek niekodująca sekwencją wypychającą. Wspomnianą sekwencją korzystnie jest ludzka sekwencja niekodująca. Jeszcze korzystniej tę niekodującą sekwencję wypychającą stanowi region intronu ludzkiej genomowej fosforybozylotransferazy hypoksantynowej (HPRT). [0023] Korzystnie, zdefiniowany powyŝej adenowirusowy wektor hybrydowy zawiera tak- Ŝe element vii., który stanowi druga niekodująca sekwencja wypychająca, umiejscowiona
![sekwencję 3' niezbędną do replikacji alfawirusa; v. sekwencję poliadenylacji; oraz vi. drugą sekwencję odwróconego powtórzenia końcowego (ITR) adenowirusa. [0020] Natura elementu i.](/docs-images/53/14208155/images/page_7.jpg "w alfawirusowo-adenowirusowym wektorze hybrydowym według niniejszego wynalazku, tj.")
8 pomiędzy elementem v. i elementem vi. zdefiniowanymi powyŝej. [0024] Natura elementu iii. w adenowirusowym wektorze hybrydowym według niniejszego wynalazku nie jest jego krytycznym aspektem. Tkankowo specyficzny promotor iii. jest korzystnie promotorem specyficznym dla nowotworu. Przykłady promotorów specyficznych dla nowotworu obejmują promotory AFP, telomerazy TERT, białko związane z trzustką PAP, E2F i HIF. [0025] Według korzystnej konkretnej postaci wynalazku promotor specyficzny dla nowotworu ma sekwencję SEQ ID No 7 odpowiadającą promotorowi/wzmacniaczowi AFP, (AFP p+e) lub sekwencję SEQ ID No 15 odpowiadającą telomerazie TERT lub jakąkolwiek inną sekwencję wykazującą wystarczającą homologię odpowiednio z sekwencją SEQ ID No 7 lub z sekwencją SEQ ID No 15 wystarczającą do spełnienia tej funkcji. [0026] Natura elementu iv. w adenowiruowym wektorze hybrydowym według niniejszego wynalazku nie jest jego krytycznym aspektem. Sekwencje alfawirusowe elementu iv. korzystnie pochodzą od wirusa Semliki Forest (SFV). JednakŜe moŝliwe jest zastosowanie innych sekwencji alfawirusowych pochodzących od gatunków naleŝących do rodziny Togaviridae, przykładowo SIN, RRV i VEE. [0027] Wspomniany łańcuch iv. cdna pochodzącego od alfawirusa, którego sekwencja jest częściowo komplementarna do RNA alfawirusa, korzystnie zawiera (oprócz sekwencji kodującej co najmniej jeden interesujący gen egzogenny): a) sekwencję 5' niezbędną do replikacji alfawirusa, b) sekwencję kodującą niestrukturalne białka wymagane do replikacji RNA alfawirusa, c) co najmniej jeden promotor subgenomowy alfawirusa, oraz d) sekwencję 3' niezbędną do replikacji alfawirusa; [0028] Element iv. korzystnie stanowi replikon funkcjonalnie kontrolowany przez promotor iii., w którym z kolei alfawirusowy promotor subgenomowy zawarty w iv.c) funkcjonalnie kontroluje ekspresję interesującego genu egzogennego. [0029] Według korzystnej konkretnej postaci wynalazku sekwencje a) do c) elementu iv. w całości zawierają sekwencję wybraną z SEQ ID No 3 (SFV 5'-rep-Psg) lub jakiejkolwiek innej sekwencji wykazującej homologię z SEQ ID No 3 wystarczającą do spełnienia tej funkcji, oraz SEQ ID No 4 (SFV 5'-rep-Psg-enh) lub jakiejkolwiek innej sekwencji wykazującej homologię z SEQ ID No 4 wystarczającą do spełnienia tej funkcji. [0030] Według szczególnie korzystnej konkretnej postaci wynalazku element iv.d) ma sekwencję SEQ ID No 5 (SFV3') lub jakąkolwiek inną sekwencji wykazującą homologię z
 lub sekwencję SEQ ID No 15")
9 SEQ ID No 5 wystarczającą do spełnienia tej funkcji. [0031] W elemencie iv. alfawirusowo-adenowirusowego wektora hybrydowego według niniejszego wynalazku interesującym egzogennym genem jest korzystnie gen terapeutyczny lub gen reporterowy albo połączenie obydwu z nich. Bez zamiaru ograniczania zakresu wynalazku gen terapeutyczny jest korzystnie wybrany spośród ssaczej interleukiny IL-12, czynnika stymulującego tworzenie kolonii GMCSF, interferonu alfa oraz kinazy tymidynowej (tk) wirusa opryszczki zwykłej. [0032] Interesujący egzogenny gen w elemencie iv. moŝe być ponadto genem reporterowym. Bez zamiaru ograniczania zakresu wynalazku gen reporterowy moŝe zostać wybrany spośród LacZ, Lucyferazy, tk i GFP. [0033] Genem terapeutycznym jest szczególnie korzystnie ssacza interleukina IL-12, a jeszcze korzystniej genem terapeutycznym jest ludzka interleukina, hil-12. [0034] Adenowirusowy wektor hybrydowy do ekspresji genów moŝe zawierać w elemencie iv serię jednego lub kilku podzestawów (promotor subgenomowy + interesujący gen egzogenny). [0035] Natura elementu v. w adenowirusowym wektorze hybrydowym według niniejszego wynalazku nie jest jego krytycznym aspektem. Elementem v. jest korzystnie sekwencja poliadenylacji SV40. Wspomnianą sekwencją poliadenylacji SV40 jest szczególnie korzystnie sekwencja SEQ ID No 6 lub jakakolwiek inna sekwencja wykazująca homologię z SEQ ID No 6 wystarczającą do spełnienia tej funkcji. [0036] Natura elementu vi. w adenowirusowym wektorze hybrydowym według niniejszego wynalazku nie jest jego krytycznym aspektem. Według korzystnej postaci wspomniany wektor adenowirusowy do ekspresji genów zawiera sekwencję odwróconego powtórzenia końcowego (ITR) jak element vi., który ma sekwencję SEQ ID No 2 lub jakąkolwiek inną sekwencją wykazującą homologię z SEQ ID No 2 wystarczającą do spełnienia tej funkcji. [0037] Natura elementu vii. w adenowirusowym wektorze hybrydowym nie jest jego krytycznym aspektem. Drugą niekodującą sekwencją wypychającą moŝe być jakaolwiek taka sekwencja. Korzystne jest to ludzka sekwencja niekodująca, a w szczególności sekwencja z ludzkiego kosmidu C346. [0038] Adenowirusowy wektor hybrydowy do ekspresji genów według niniejszego wynalazku moŝe mieć zmienną długość i korzystnie ma długość w zakresie pomiędzy 27 a 38 tysięcy par zasad. [0039] Według korzystnej konkretnej postaci adenowirusowy wektor hybrydowy zawiera ITR 5' jako pierwszą sekwencję odwróconego powtórzenia; HPRT, region intronu ludzkiej genomowej fosforybozylotransferazy hypoksantynowej jako pierwszą sekwencję wypy-
 wirusa opryszczki zwykłej. [0032] Interesujący egzogenny gen w elemencie iv. moŝe być ponadto genem reporterowym.")
10 chającą; AFP (p+e), promotor specyficzny dla nowotworu; sekwencję replikonu SFV zawierającą mil-12, mysiej interleukiny-12; SV40 PolyA, sekwencję poliadenylacji SV40; C346, sekwencję ludzkiego kosmidu genomowego C346 jako drugą sekwencję wypychającą oraz ITR 3' jako drugą sekwencję odwróconego powtórzenia końcowego. [0040] Według dodatkowej korzystnej konkretnej postaci adenowirusowy wektor hybrydowy zawiera ITR 5' jako pierwszą sekwencję odwróconego powtórzenia końcowego; HPRT, region intronu ludzkiej genomowej fosforybozylotransferazy hypoksantynowej jako pierwszą sekwencję wypychającą; AFP (p+e), promotor specyficzny dla nowotworu; sekwencję replikonu SFV zawierającą LacZ; SV40 PolyA, sekwencję poliadenylacji SV40; C346, sekwencję ludzkiego kosmidu genomowego C346 jako drugą sekwencję wypychającą oraz ITR 3' jako drugą sekwencję odwróconego powtórzenia końcowego. [0041] Według szczególnie korzystnej dodatkowej postaci adenowirusowy wektor hybrydowy zawiera ITR 5' jako pierwszą sekwencję odwróconego powtórzenia; HPRT, region intronu ludzkiej genomowej fosforybozylotransferazy hypoksantynowej jako pierwszą sekwencję wypychającą; AFP (p+e), promotor specyficzny dla nowotworu; sekwencję replikonu SFV zawierającą hil-12, ludzkiej interleukiny-12; SV40 PolyA, sekwencję poliadenylacji SV40; C346, sekwencję ludzkiego kosmidu genomowego C346 jako drugą sekwencję wypychającą oraz ITR 3' jako drugą sekwencję odwróconego powtórzenia. [0042] Według korzystnej konkretnej postaci według niniejszego wynalazku, adenowirusowy wektor hybrydowy do ekspresji genów ma sekwencję SEQ ID No 8 lub jakąkolwiek inną sekwencję wykazującą homologię z SEQ ID No 8 wystarczającą do spełnienia tej funkcji. [0043] Według dodatkowej korzystnej konkretnej postaci niniejszego wynalazku, adenowirusowy wektor hybrydowy do ekspresji genów ma sekwencję SEQ ID No 9 lub jakąkolwiek inną sekwencję wykazującą homologię z SEQ ID No 9 wystarczającą do spełnienia tej funkcji. [0044] Według dodatkowej korzystnej konkretnej postaci niniejszego wynalazku, adenowirusowy wektor hybrydowy do ekspresji genów ma sekwencję SEQ ID No 10 lub jakąkolwiek inną sekwencje wykazującą homologię z SEQ ID No 10 wystarczającą do spełnienia tej funkcji. [0045] Niniejszy wynalazek dotyczy ponadto sposobu otrzymywania wspomnianego adenowirusowego wektora hybrydowego, obejmującego skladanie zdefiniowanych powyŝej elementów i. do vi. lub i. do vii. adenowirusowego wektora hybrydowego przez zastosowanie technik inŝynierii genetycznej. [0046] Niniejszy wynalazek dotyczy takŝe zastosowania wspomnianego wektora hybry-
![[0040] Według dodatkowej korzystnej konkretnej postaci adenowirusowy wektor hybrydowy zawiera ITR 5' jako pierwszą sekwencję odwróconego powtórzenia końcowego; HPRT, region intronu ludzkiej genomowej](/docs-images/53/14208155/images/page_10.jpg "fosforybozylotransferazy hypoksantynowej jako pierwszą sekwencję wypychającą; AFP (p+e), promotor specyficzny dla nowotworu; sekwencję replikonu SFV zawierającą LacZ; SV40 PolyA, sekwencję")
11 dowego do przenoszenia materiału genetycznego do komórki, a w szczególności do wprowadzania i ekspresji obcych genów w komórkach eukariotycznych, które mogą być komórkami docelowymi dla adenowirusów. Wspomniane zastosowanie obejmuje podawanie pacjentowi wspomnianego wektora hybrydowego. [0047] ZakaŜenie komórek nowotworowych wspomnianym adenowirusowym wektorem hybrydowym według wynalazku powoduje transkrypcję mrna replikonu alfawirusa SFV z promotora specyficznego dla nowotworu, a tym samym gen Rep ulega translacji i RNA SFV ulega namnoŝeniu. Rep wytwarza takŝe subgenomowy RNA SFV, z którego gen terapeutyczny lub reporterowy będzie eksprymowany na wysokim poziomie. Produkt genu terapeutycznego wydzielany przez zakaŝone komórki będzie aktywować komórki układu immunologicznego w miejscu zakaŝenia. Replikacja SFV będzie ponadto wywoływać apoptozę zakaŝonych komórek, co doprowadzi do uwalniania antygenów nowotworowych z komórek apoptotycznych; które mogą zostać wychwycone przez komórki prezentujące antygen (APC), w ten sposób aktywując odpowiedź immunologiczną przeciw nowotworowi. JednakŜe, jeŝeli ten wektor hybrydowy zakazi komórki niebędące komórkami nowotworowymi mrna replikonu SFV nie będzie transkrybowany, a zatem nie wystąpi ekspresja transgenu ani teŝ apoptoza. [0048] Komórki nowotworowe są korzystnie zakaŝane adenowirusowym wektorem hybrydowym według wynalazku, tak Ŝe mrna replikonu SFV jest transkrybowany z promotora specyficznego dla nowotworu AFP, a zatem gen Rep ulegnie translacji oraz RNA SFV zostanie namnoŝony. Rep będzie takŝe wytwarzać subgenomowy RNA SFV, z którego mil- 12 lub hil-12 będą eksprymowane na wysokim poziomie. mil-12 lub hil-12 wydzielane przez zakaŝone komórki będą aktywować komórki układu immunologicznego w miejscu zakaŝenia. Replikacja SFV będzie ponadto indukować apoptozę zakaŝonych komórek, co doprowadzi do uwalniania antygenów nowotworowych przez komórki apoptotyczne; które mogą zostać wychwycone przez komórki prezentujące antygen (APC), aktywując w ten sposób odpowiedź immunologiczną przeciw nowotworowi. JednakŜe, jeŝeli ten wektor hybrydowy zakazi komórki niebędące komórkami nowotworowymi mrna replikonu SFV nie będzie transkrybowany, a zatem nie wystąpi ekspresja transgenu ani teŝ apoptoza. [0049] Dodatkowym przedmiotem niniejszego wynalazku jest zastosowanie zdefiniowanego powyŝej adenowirusowego wektora hybrydowego w procesie przenoszenia materiału genetycznego do komórki, korzystnie komórki nowotworowej, obejmujące podawanie wspomnianego wektora hybrydowego pacjentowi. Jeszcze korzystniej wspomnianą komórką jest komórka nowotworowa eksprymująca AFP. [0050] Dodatkowym przedmiotem niniejszego wynalazku jest zastosowanie zdefiniowa-
![[0047] ZakaŜenie komórek nowotworowych wspomnianym adenowirusowym wektorem hybrydowym według wynalazku powoduje transkrypcję mrna replikonu alfawirusa SFV z promotora specyficznego dla nowotworu, a](/docs-images/53/14208155/images/page_11.jpg "tym samym gen Rep ulega translacji i RNA SFV ulega namnoŝeniu. Rep wytwarza takŝe subgenomowy RNA SFV, z którego gen terapeutyczny lub reporterowy będzie eksprymowany na wysokim poziomie.")
12 nego powyŝej adenowirusowego wektora hybrydowego do wytwarzania skutecznego leku do leczenia nowotworów oraz jego zastosowanie do wywoływania odpowiedzi immunologicznej przeciw obcym antygenom. Wspomniane zastosowanie obejmuje podawanie wspomnianego leku pacjentowi. [0051] Dodatkowym przedmiotem niniejszego wynalazku jest kompozycja farmaceutyczna zawierająca co najmniej jeden adenowirusowy wektor hybrydowy zdefiniowany według niniejszego wynalazku oraz jego zastosowanie w procesie leczenia nowotworów lub wywoływania odpowiedzi immunologicznej na obce antygeny. [0052] Wspomniana kompozycja farmaceutyczna korzystnie zawiera adenowirusowy wektor hybrydowy według niniejszego wynalazku, w którym interesującym genem egzogennym jest ssacza interleukina, IL-12, korzystnie ludzka interleukina hil-12. Wspomniane zastosowanie obejmuje podawanie pacjentowi kompozycji farmaceutycznej zawierającej wspomniany wektor hybrydowy. [0053] Niniejszy wynalazek dotyczy takŝe sposobu leczenia raka przez zastosowanie wektora hybrydowego według niniejszego wynalazku, który to sposób obejmuje podawanie pacjentowi wspomnianego wektora hybrydowego. [0054] Zatem według korzystnych postaci niniejszego wynalazku, AFP (p+e) został wybrany jako promotor specyficzny dla nowotworu, skonstruowano dwa adenowirusowe wektory hybrydowe, w których replikon SFV jest pod kontrolą promotora AFP, oraz gen reporterowy LacZ i gen terapeutyczny IL-12 są wstawione pod kontrolą subgenomowego promotora SFV, odpowiednio - Ad/AFP-SFV-LacZ i Ad/AFP-SFV-mIL-12; a dwa wektory adenowirusowe typu gutless niosące Lacz i IL-12 z myszy bezpośrednio kontrolowane przez promotor AFP - Ad/AFP-LacZ i Ad/AFP-mIL-12- przygotowano jako wektory kontrolne. Wykazano, Ŝe wektor hybrydowy według niniejszego wynalazku działa skuteczniej niŝ dotychczas stosowane wektory kontrolne. [0055] Według niniejszego wynalazku wykazano, Ŝe wektor Ad/AFP-SFV-mIL-12 moŝe być wektorem przydatnym w terapii nowotworów HCC (raka wątroby) eksprymujących AFP. [0056] Wykazano, Ŝe zastosowanie innych promotorów nowotworowych, takich jak promotor telomerazy, TERT, który jest szeroko aktywowany w większości nowotworów złośliwych, do kontrolowania SFV moŝe przekształcić zastosowanie wektora hybrydowego, takiego jak ten według niniejszego wynalazku, w ogólną strategię do leczenia wszystkich typów raka. [0057] Ponadto, oraz korzystnie, wektor hybrydowy według niniejszego wynalazku specyficznie współdziała z komórkami nowotworowymi oraz niszczy komórki nowotworowe
![[0051] Dodatkowym przedmiotem niniejszego wynalazku jest kompozycja farmaceutyczna zawierająca co najmniej jeden adenowirusowy wektor hybrydowy zdefiniowany według niniejszego wynalazku oraz jego](/docs-images/53/14208155/images/page_12.jpg "zastosowanie w procesie leczenia nowotworów lub wywoływania odpowiedzi immunologicznej na obce antygeny.")
13 bez potrzeby wprowadzenia genu terapeutycznego. Ponadto wykazano, Ŝe wektor hybrydowy według niniejszego wynalazku korzystnie wywołuje silną aktywność przeciwnowotworową, gdy zawiera on gen terapeutyczny, taki jak IL-12. KRÓTKI OPIS FIGUR [0058] Figura 1 (A): przedstawia diagram wektora hybrydowego zgodnie z postacią według wynalazku, Ad-SFV, zawierającego sekwencję adenowirusową typu gutless, w której replikon SFV został wstawiony pod kontrolą promotora/wzmacniacza AFP (AFP (p+e)) oraz zawierającego heterologiczny gen mil-12, który jest wstawiony pod kontrolą promotora subgenomowego SFV (Pr sg). Figura 1 (B) przedstawia aktywność przeciwnowotworową wektora hybrydowego według wynalazku: po zakaŝeniu komórek nowotworowych HCC wektorem hybrydowym (strona prawa), ze względu na obecność w wektorze hybrydowym replikonu SFV zawierającego mil-12 następuje wysoka ekspresja mil-12, która będzie aktywować komórki układu immunologicznego w miejscu zakaŝenia. Replikacja SFV będzie ponadto wywoływać apoptozę zakaŝonych komórek. JednakŜe, jeŝeli ten wektor hybrydowy zakazi komórki nienowotworowe, mrna replikonu SFV nie ulegnie transkrypcji i w konsekwencji nie wystąpi ekspresja transgeniczna ani teŝ apoptoza. Na tej figurze: ITR, sekwencje adenowirusowych odwróconych powtórzeń końcowych; ψ, adenowirusowy sygnał pakowania; HPRT i C346, sekwencje wypychającego DNA odpowiednio z regionu intronu genomowej fosforybozylotransferazy hipoksantynowej lub z ludzkiego kosmidu C346; PolyA, sygnał poliadenylacji (przykładowo z SV40); APC, komórki prezentujące antygen. Figura 2 przedstawia strukturę adenowirusowych wektorów hybrydowych typu gutless oraz wektorów adenowirusowych typu gutless. AFP-SFV-LacZ i AFP-SFVmIL-12 są adenowirusowymi wektorami hybrydowymi w których sekwencja replikonu SFV jest pod kontrolą promotora/wzmacniacza AFP (AFP(p+e)) oraz heterologiczne geny odpowiednio LacZ lub mil-12 sklonowano pod kontrolą promotora subgenomowego SFV (Pr sg). AFP-lacZ i AFP-mIL-12 są wektorami adenowirusowymi zawierającymi LacZ lub mil-12 bezpośrednio kontrolowane przez AFP (p+e). SFV nsp1-4, niestrukturalne białka SFV. Figura 3 przedstawia specyficzną ekspresję mil-12 in vitro w komórkach raka wą-
![KRÓTKI OPIS FIGUR [0058] Figura 1 (A): przedstawia diagram wektora hybrydowego zgodnie z postacią według wynalazku, Ad-SFV, zawierającego sekwencję adenowirusową typu gutless, w której replikon SFV](/docs-images/53/14208155/images/page_13.jpg "został wstawiony pod kontrolą promotora/wzmacniacza AFP (AFP (p+e)) oraz zawierającego heterologiczny gen mil-12, który jest wstawiony pod kontrolą promotora subgenomowego SFV (Pr sg).")
14 troby eksprymujących AFP, HCC (A) oraz w komórkach niepochodzących od HCC (B) po zakaŝeniu wektorami hybrydowymi Ad-SFV: AFP-mIL-12 (AFP-12), AFP- SFV-mIL-12 (AFP-SFV-12) lub wektorem kontrolnym AdCMVmIL-12 (CMV-12). Zbadano róŝne wielokrotności zakaŝenia moi (10, 100 i 1000). Hep3B, Huh-7, HepG2 i PLC/PRF/5: linie komórek HCC; Hela, A549, MHC1, SK-Hep-1 oraz Clon 9: linie niepochodzących od HCC. Figura 3B przedstawia tylko ekspresję odpowiadającą moi Figura 4 przedstawia specyficzną ekspresję β-gal w 4 liniach komórek HCC (Hep3B, Huh-7, HepG2 i PLC/PRF/5) po zakaŝeniu in vitro wektorem hybrydowym AFP- SFV-lacZ oraz wektorem kontrolnym AFP-LacZ przy róŝnych moi (10, 100 lub 1000). Figura 5 przedstawia analizę ekspresji β-gal w liniach komórek HCC zakaŝonych AFP-LacZ i AFP-SFV-LacZ. Mikrofotografie komórek zakaŝonych AFP-LacZ (A-C) lub AFP-SFV-LacZ (D-F) oraz wybarwionych X-Gal. A i D, Hep3B; B i E, Huh7; C i F, HepG2. Figura 6 przedstawia kinetykę ekspresji IL-12 w liniach komórek HCC Hep3B (A) i Huh-7 (B), zakaŝonych in vitro wektorami adenowirusowymi AFP-mIL-12 (AFP-12) lub AFP-SFV-mIL-12 (AFP-SFV-12) przy moi Figura 7 przedstawia wywoływanie śmierci komórkowej po zakaŝeniu in vitro linii komórek HCC - Hep3B (A) i McA-RH7777 (B) - wektorami AFP-IL-12 (AFP-12), AFP-SFV-IL-12 (AFP-SFV-12), AFP-LacZ, AFP-SFV-LacZ lub wektorem kontrolnym Ad/CMVmIL-12 (CMV-12). PrzeŜycie komórek przedstawiono jako procent Ŝywych komórek w zakaŝonych studzienkach w porównaniu z Ŝywymi komórkami w niezakaŝonych kontrolnych studzienkach. Figura 8 przedstawia ekspresję SFV Rep w komórkach HCC - Hep3B (A-D) i Huh-7 (E i F) po zakaŝeniu wektorami AFP-mIL-12 (A i B) lub AFP-SFV-mIL-12 (C-F) przy moi Dwa dni po zakaŝeniu komórki utrwalono i analizowano metodą immunofluorescencji z przeciwciałem specyficznym względem Rep. Komórki eksprymujące Rep uwidoczniono za pomocą mikroskopu fluorescencyjnego z filtrem FITC (A, C i E), podczas gdy jądra komórkowe wybarwione DAPI we wszystkich komórkach uwidoczniono z zastosowaniem filtra UV (B, D i F). Figura 9 przedstawia przenoszenie genów z zastosowaniem wektorów hybrydowych typu gutless in vivo, (A-D), wydajność przenoszenia genów i wywoływania apoptozy w nowotworach Huh-7. Ludzkie nowotwory Huh-7 przyjęte w nagich myszach z niedoborem odporności traktowano przez donowotworowe wstrzyknięcie wektorów
 po zakaŝeniu in vitro wektorem hybrydowym AFP- SFV-lacZ oraz wektorem kontrolnym AFP-LacZ")
15 AFP-LacZ (n=4) lub AFP-SFV-LacZ (n=4) przy 1x10 10 cząstek wirusa/zwierzę. Trzy dni po podaniu wirusa myszy uśmiercono i skrawki guza analizowano w celu zbadania ekspresji transgenu przez wybarwienie X-Gal (A-B) lub w celu zbadania wywoływania apoptozy przez zastosowanie testu TUNEL (C-D). A, C; Mikrofotografie nowotworów, do których wstrzyknięto AFP-LacZ. B, D. Mikrofotografie nowotworów, do których wstrzyknięto AFP-SFV-LacZ. (E-G), Specyficzność ekspresji genów z wektorami hybrydowymi typu gutless. Zdrowym myszom Balb/c wstrzyknięto doŝylnie 1x10 10 cząstek wirusa wektorów AFP-LacZ (E), AFP-SFV-LacZ (F) lub Ad/CMV-LacZ (G). Przedstawiono mikrofotografie skrawków wątroby wykonanych trzy dni po zaszczepieniu i wybarwionych X-Gal. Figura 10 przedstawia traktowanie nowotworów HCC wektorami hybrydowymi. Ortotopowe nowotwory HCC załoŝono przez wszczepienie komórek McH-RH7777 do wątroby szczura. Gdy nowotwór osiągnął wielkość średnicy 7-10 mm, zwierzęta potraktowano (A-C) lub 2 x10 11 (D-G) cząstek wirusa AFP-mIL-12, AFP-SFVmIL-12 lub roztworem soli fizjologicznej jako kontrolą. Wielkość nowotworu zmierzono w dniach 15 i 30 po podaniu roztworu soli fizjologicznej (A i E), AFP-mIL-12 (B i F) lub AFP-SFV-mIL-12 (C i G). G; Ocena przeŝycia zwierząt. Figura 11 przedstawia badanie toksyczności na szczurach zaszczepionych wektorami eksprymującymi IL-12. Poziom transaminaz (GPT, GOLT i GGTL) (A) lub IL-12 (B) oznaczono w surowicy szczurów z nowotworami HCC w wątrobie oraz tych, które zaszczepiono donowotworowo wektorami adenowirusowymi AFP-SFV-IL-12, AFP-SFV-mIL-12, wektorem alfawirusowym SFV-IL-12 lub roztworem soli fizjologicznej. Pomiary wykonano w dniach 4 i 8 po potraktowaniu. Figura 12 przedstawia wybarwienie hematoksyliną/eozyną skrawków wątroby szczurów traktowanych adenowirusowymi wektorami hybrydowymi. Szczury niosące nowotwory HCC potraktowano przez donowotworowe wstrzyknięcia roztworu soli fizjologicznej (A), wektorów adenowirusowych AFP-IL-12 (B), AFP-SFV-IL-12 (C) lub cząstek wirusa SFV-IL-12 (D). Trzy dni po potraktowaniu zwierzęta uśmiercono, wątroby usunięto i utrwalono formolem, a uzyskane skrawki wybarwiono hematoksyliną/eozyną. Czarne strzałki wskazują obszary eozynochłonnych hepatocytów. Figury 13A i 13B przedstawiają mapy restrykcyjne plazmidów odpowiednio pgl3/afp i pbs/mil-12. Figury 14A i 14B przedstawiają mapy restrykcyjne plazmidów odpowiednio ptgc3001 i ptgc3011. Figury 15A i 15B przedstawiają mapy restrykcyjne plazmidów odpowiednio
, Specyficzność ekspresji genów z wektorami hybrydowymi typu gutless.")
16 ptgc3012 i ptgc3013. Figura 16 przedstawia mapę restrykcyjną plazmidu ptgc3014. POSTACIE WYNALAZKU Plazmidy [0059] pgem-t easy i pcmvb uzyskano z Promega, U.S.A., a pbs-sk+ z Stratagene, U.S.A. pstk120 uzyskano dzięki uprzejmości dr Kochanek (University of Ulm, Niemcy). pbk-sfv-1 i pbk-sfv-3 opisano w Berglund P. i in. Enhancing immune responses using suicidal DNA vaccines. Nature Biotechnology 1998, 16: pgl3/afp i pbs/mil-12 (Yonglian Sun, Cheng Qian, Dacheng Peng oraz Jesus Prieto Gene transfer to liver cancer cells of B7-1 in addition to IL-12 changes immunoeffector mechanisms and suppresses Th1 cytokine production induced by IL-12 alone. Human Gene Therapy 11: ) zostały wytworzone w naszym laboratorium. [0060] W celu skonstruowania pgl3/afp regiony promotora/wzmacniacza AFP (p+e) uzyskano przez amplifikację PCR ludzkiego DNA genomowego. Starterami zastosowanymi do amplifikacji promotora AFP (AFP pro) były CTCTAGATTTTCTGCCCCA- AAGAGCTC i CGGGATCCTGTTATTGGCAGTGGTGGAA. Starterami zastosowanymi do amplifikacji wzmacniacza AFP były CGGAATTCGCCTGTCATACAGCTAATAA i CTCTAGACTGTCAAATAAGTGG CCTGG. Sekwencje promotora (217 par zasad) i wzmacniacza (785 par zasad) wklonowano do plazmidów pgem-t. Następnie potwierdzenie zamplifikowanych fragmentów przeprowadzono przez sekwencjonowanie. Promotor AFP usunięto z plazmidu pgem-t/afp-p przez strawienie Xba I/BamHI i wstawiono przez ligację tępych końców do plazmidu pgl3-basic strawionego SmaI. W ten sposób uzyskano plazmid pgl3/afp-p. Wzmacniacz AFP usunięto z plazmidu pgem-t/afp-e przez strawienie XbaI/EcoRI i wstawiono przez ligację tępych końców do plazmidu pgl3/afp-p strawionego NheI, ostatecznie uzyskując plazmid pgl3/afp. Linie komórkowe i hodowle tkankowe 30 [0061] Linie ludzkich komórek HCC Hep3B, PLC/PRF/5, HepG2 i SK-Hep-1, ludzką linię komórek raka gruczołowego nabłonka szyjki macicy HeLa, ludzką linię komórek raka płuca A549, ludzką linię komórek embrionalnych pochodzenia nerkowego 293, szczurze komórki HCC McA-RH7777, MHC1 i Clone 9 prawidłowych szczurzych hepatocytów oraz szczurzą linię komórek HCC Hepa1-6 uzyskano z ATCC. Komórki 293 eksprymujące rekombinazę Cre (293Cre4) uzyskano z Merck Research Laboratories. Komórki Hep3B, PLC/PRF/5, HeLa, SK-Hep-1, Clone 9, Huh-7 i Hepa1-6 hodowano w poŝywce DMEM

17 16 5 uzupełnionej 10% inaktywowanej termicznie płodowej surowicy bydlęcej (FBS) oraz penicyliną/streptomycyną. Komórki HepG2 i A549 hodowano w poŝywce RPMI 1640 uzupełnionej 10% inaktywowanej termicznie FBS oraz penicyliną/streptomycyną. Komórki McH-RH7777 i MHC1 hodowano w poŝywce DMEM uzupełnionej 20% końskiej surowicy i 5% FBS. Komórki 293Cre4 hodowano w poŝywce DMEM uzupełnionej 10% FBS i 0,4 mg/ml G418. Zwierzęta [0062] Siedmiotygodniowe samice nagich myszy BALB/c z niedoborem odporności uzyskano z Charles Rivers Laboratories (Barcelona, Hiszpania). Samce szczurów Buffalo w wieku 4-6 tygodni uzyskano z CIFA (Animal Installations of the University of Navarra). Myszy i szczury hodowano w standardowych warunkach w CIFA. Myszy Nude otrzymywały napromieniowane poŝywienie z autoklawowaną wodą do picia. Manipulacje z nagimi myszami zawsze prowadzono w komorze z przepływem laminarnym. Wszystkie procedury stosowane w doświadczeniach na zwierzętach były zgodne ze standardowymi zaleceniami i protokołami dotyczącymi opieki nad zwierzętami laboratoryjnymi i ich stosowania. Konstrukcja wektorów Konstrukcja kaset ekspresyjnych AFP-SFV [0063] Sekwencję 5'-końca SFV (1-292 nt) namnoŝono z zastosowaniem PCR stosując plazmid pbk-sfv-1 (zawierający sekwencję replikonu SFV pełnej długości) jako matrycę. Starter 1 zawierał miejsce restrykcyjne SpeI na 5'-końcu (podkreślona), a następnie 50 nt sekwencji promotora AFP oraz pierwszych 20 nt sekwencji SFV (kursywa): 5'-ACT AGT TAA CAG GCA TTG CCT GAA AAG AGT ATA AAA GAA TTTCAG CAT GAT TTT CCA TGG CGG ATG TGT GAC ATA C-3'. Starter 2 zawierał miejsce restrykcyjne XhoI (podkreślone), a następnie 19 nt sekwencji SFV (kursywa): 5'-CTC GAG GAT ATC CAA GAT GAG TGT GT-3'. Z zastosowaniem PCR wytworzono fragment DNA wielkości 342 pz i wklonowano go bezpośrednio do plazmidu pgem-t-easy z wytworzeniem pgem-te-sfv-1. Brak błędów PCR w tym plazmidzie potwierdzono przez sekwencjonowanie. Fragment 342 pz wycięto z pgem-te-sfv-1 przez trawienie SpeI i XhoI i wklonowano do pgl3/afp strawionego tymi samymi enzymami uzyskując pgl3/afp-sfv-1, który zawierał kompletny promotor AFP (217 pz) i wzmacniacz (785 pz), a następnie sekwencję 5'-końca SFV (SFV-1, zawierającą 292 pz). Kasetę AFP-SFV-1 (1342 pz) uzyskano z pgl3/afp-sfv-1 przez trawienie MluI/XhoI, potraktowano polimerazą Klenowa i wklonowano do pbs-sk+ strawionego EcoRV, otrzymując pbs/afp-sfv-1. Późny sy-
.")
18 gnał PolyA SV40 (262 pz) usunięto z pgl3/afp przez trawienie XbaI/BamHI, końce przekształcono w końce tępe przez zastosowanie polimerazy Klenowa i wstawiono w miejsce SalI pbs/afp-sfv-1 takŝe z końcami przekształconymi w tępe końce przez zastosowanie polimerazy Klenowa, uzyskując pbs/afp-sfv-1-pa. Polilinker zawierający obydwa unikalne miejsca ApaI i NruI wstawiono pomiędzy miejsca BamHI i XmaI do pbs/afp-sfv-1-pa. Sekwencję 3'-końca SFV zawierającą 7985 pz usunięto przez trawienie SpeI/EcoRV z pbk-sfv-1, końce przekształcono w końce tępe przez zastosowanie polimerazy Klenowa i wstawiono w miejsce EcoRV pbs/afp-sfv-1-pa, otrzymując pbs/afp-sfv-pa. [0064] Gen reporterowy LacZ uzyskano z pcmvb przez strawienie NotI, potraktowano polimerazą Klenowa i wstawiono w miejsce BamHI pbs/afp-sfv-pa potraktowanego polimerazą Klenowa z wytworzeniem pbs/afp-sfv-lacz-pa. Kasetę mil-12 zawierającą geny kodujące podjednostki p35 i p40 połączone przez wewnętrzne miejsce wiązania rybosomu (IRES) oddzielono z pbs/mil-12 przez strawienie SpeI/XhoI, potraktowano polimerazą Klenowa i wstawiono w pozycję BamHI pbs/afp-sfv-mil-12-pa, takŝe z końcami doprowadzonymi do tępych końców z zastosowaniem polimerazy Klenowa, otrzymując plazmid pbs/afp-sfv-mil-12-pa. Konstrukcja adenowirusowych wektorów hybrydowych typu gutless [0065] Skonstruowano cztery wektory adenowirusowe typu gutless, jak przedstawiono na Figurze 2. AFP-SFV-lacZ i AFP-SFV-mIL-12 zawierają rekombinowaną sekwencję replikonu SFV pod kontrolą promotora i wzmacniacza AFP. W tych wektorach gen reporterowy LacZ lub gen terapeutyczny mil-12 sklonowano pod kontrolą odpowiednio promotora subgenomowego SFV. AFP-lacZ i AFP-mIL-12 są takŝe wektorami adenowirusowymi typu gutless, które zawierają geny LacZ i mil-12, bezpośrednio pod kontrola odpowiednio promotora/wzmacniacza AFP. Sposób zastosowany do skonstruowania tych wektorów opisano poniŝej. W celu wytworzenia wektora adenowirusowego z wystarczającą przestrzenią do klonowania, aby przyjął kasetę ekspresyjną AFP-SFV-IL-12, zmodyfikowano plazmid pstk120 zawierający sekwencję wektora adenowirusowego typu gutless. W tym celu fragment wielkości 9 kpz usunięto z plazmidu pstk120 przez trawienie ApaI. Ponadto, polilinker zawierający miejsca AscI i SbfI wstawiono do tego nowego plazmidu otrzymując ptgc3001. Plazmid ten zawierał lewy ITR, sygnał pakowania, wypychający DNA z HPRT i C346 oraz prawy ITR. Kasetę AFP-SFV-LacZ oddzielono przez trawienie ApaI z pbs/afp-sfv-lacz-pa i wstawiono w miejsce ApaI ptgc3001, otrzymując ptgc3011. Podobnie kasetę AFP-SFV-mIL-12 usunięto z pbs/afp-sfv-mil-12-pa przez trawienie BssHII, potraktowano polimerazą Klenowa i wstawiono w miejsce AscI

19 ptgc3001 podobnie potraktowanego polimerazą Klenowa, otrzymując ptgc3012. Konstrukcja wektorów kontrolnych Konstrukcja wektora adenowirusowego typu gutless AFP-LacZ [0066] Sekwencję wzmacniacza/promotora AFP (AFP p+e) oddzielono z pgl3/afp przez trawienie MluI/XhoI potraktowano polimerazą Klenowa i wstawiono do pcmvb, który uprzednio strawiono EcoRI/XhoI i potraktowano polimerazą Klenowa. W ten sposób usunięto najwcześniejszy promotor CMV z pcmvb i podstawiono AFP (p+e) z wytworzeniem pafpb. Następnie kasetę AFP-LacZ (5077 pz) usunięto z pafpb przez trawienie XbaI/NarI, potraktowano polimerazą Klenowa i wstawiono w miejsce SwaI pstk120 oraz takŝe potraktowanego polimerazą Klenowa w celu wytworzenia tępych końców, otrzymując ptgc3013. Konstrukcja wektora adenowirusowego typu gutless AFP-mIL [0067] Kasetę mil-12 usunięto z pbs/mil-12 przez trawienie XhoI/SpeI i wstawiono do pgl3/afp uprzednio strawionego XhoI/XbaI, co pozwoliło usunąć gen lucyferazy z drugiego wspomnianego plazmidu oraz wytworzyć pafp-mil-12. Kasetę AFP-mIL-12 (3760 pz) usunięto z pafp-mil-12 przez trawienie BamHI/ScaI, potraktowano polimerazą Klenowa oraz wstawiono do pstk120 strawionego SwaI oraz podobnie potraktowano polimerazą Klenowa otrzymując ptgc3014. Odzyskanie wektorów adenowirusowych typu gutless [0068] Po strawieniu PmeI, ekstrakcji fenolem/chloroformem oraz wytrąceniu etanolem, 2 µg ptgc3011, ptgc3012, ptgc3013 lub ptgc3014 DNA transfekowano odpowiednio do komórek 293Cre4. Po transfekcji komórki zakaŝono wirusem pomocniczym AdLC8cluc. Kolejne etapy amplifikacji na duŝą skalę i oczyszczania przeprowadzono w sposób poprzednio opisany (Philip Ng., Robin J. Parks i Frank L.Graham. Preparation of helper-dependent adenoviral vectors. Methods in Molecular Medicine, tom 69, Gene Therapy Protocols, wyd. 2 69, , 2002; H. Zhou, L. Pastore, A. L. Beaudet. Helperdependent adenoviral vectors. Methods in Enzymology, tom 346, , 2002; Hillgenberg M., i in. System for efficient helper-dependent minimal adenovirus constructions and rescue. Hum Gene Ther., 12; , 2001). Wszystkie preparaty wektorów oczyszczono dwukrotnie przez odwirowanie w gradiencie CsCl. Oczyszczone wektory DNA analizowano przez trawienie wektorami restrykcyjnymi i wykazano brak rearanŝacji sekwencji. Miana adenowirusa typu gutless oraz zanieczyszczenia wirusem pomocniczym oznaczono z zastosowaniem ilościowej PCR. Stosunek całkowitej liczby cząsteczek wirusa względem
 z wytworzeniem pafpb.")
20 19 jednostek infekcyjnych (iu) wynosiła 20:1. Zanieczyszczenie cząsteczkami wirusa pomocniczego wynosiło około 0,5-1%. Ilościowa PCR [0069] W celu oznaczenia stopnia zanieczyszczenia wirusem pomocniczym zaprojektowano sondę i startery do ilościowej PCR regionu Ad5 E4 z zastosowaniem programu TaqMan (TaqMan Probe #2) i zsyntetyzowano w Sigma-Genosys Ltd. (startery) i Applied Biosystems (sonda). W celu oznaczenia miana adenowirusów typu gutless sondy i startery do ilościowej reakcji PCR sekwencji LacZ i mil-12 z myszy zaprojektowano z zastosowaniem programu TaqMan (TaqMan Probe #2) i zsyntetyzowano w Sigma-Genosys Ltd (startery) i Applied Biosystems (sonda). W celu oznaczenia zanieczyszczenia Ad typu dzikiego, sondy i startery do ilościowej reakcji PCR regionu Ad5 E1 zaprojektowano z zastosowaniem programu TaqMan (TaqMan Probe #2) i zsyntetyzowano w Sigma-Genosys Ltd (startery) i Applied Biosystems (sonda). Doświadczenia in vitro Ekspresja transgeniczna w komórkach zakaŝonych wektorami adenowirusowymi typu gutless [0070] Linie komórkowe pochodzące od HCC (Hep3B, Huh7, HepG2 i PLC/PRF/5) oraz linie komórkowe niepochodzące od HCC (A549,HeLa, MHC1 i Clone 9) lub linie komórkowe pochodzące od HCC, ale nieeksprymujące AFP (SK-Hep-1) zakaŝono kaŝdym z czterech wektorów adenowirusowych typu gutless (AFP-LacZ, AFP-SFV-LacZ, AFPmIL-12 lub AFP-SFV-mIL-12) przy moi odpowiednio 1000, 100 lub 10 (cząstek/komórkę). Adenowirusy pierwszej generacji (Ad/CMV-mIL-12, Ad/CMV-LacZ Ad/AFP-LacZ) zastosowano jako kontrolę. Supernatanty z komórek zakaŝonych wektorami mil-12 oraz lizaty z komórek zakaŝonych wektorami LacZ zebrano w dwóch powtórzeniach z studzienek w celu oznaczenia odpowiednio mil-12 i β-galaktozydazy (β-gal). Komórki zakaŝone wektorami LacZ takŝe wybarwiono X-gal. Poziom mil-12 (p70) zmierzono z zastosowaniem zestawu ELISA (Pharmingen, San Diego, CA). Poziom β-gal zmierzono z zastosowaniem zestawu ELISA (Roche, Szwajcaria). Przebieg w czasie ekspresji mil-12 oszacowano w komórkach HCC (Hep3B, Huh7) po zakaŝeniu AFP-mIL-12, AFP- SFV-mIL-12 lub wektorem kontrolnym Ad/CMV-mIL-12 przy moi Supernatanty zbierano codziennie do 5 dni po zakaŝeniu. Analiza specyficzności ekspresji transgenu z zastosowaniem wektorów hybrydowych Ad- SFV in vitro [0071] W celu zbadania specyficzności ekspresji transgenu z poprzednio opisanymi re-
 i")
21 kombinowanymi wektorami cztery ludzkie linie komórek HCC (Hep3B, HepG2, Huh-7 i PLC/PRF/5) oraz dwie ludzkie linie niepochodzące od HCC (HeLa i A549) lub pochodzące od HCC, ale eksprymujące AFP (SK-Hep-1) zakaŝono AFP-mIL-12, AFP-SFV-mIL-12 lub Ad-CMV-mIL-12 jako kontrolą pozytywną przy róŝnych moi (10, 100 lub 1000). Dwa dni po zakaŝeniu zebrano supernatant i oznaczono w nim zawartość mil-12. Wyniki przedstawiono na Figurze 3 (A) i (B). Nie stwierdzono ekspresji mil-12 w ludzkich komórkach HCC zakaŝonych AFP-mIL-12 przy moi 10 lub 100, a tylko przy moi 1000 zaobserwowano bardzo niski poziom mil-12 w niektórych liniach komórkowych (Figura 3A). W odróŝnieniu od tego zakaŝenie tych komórek AFP-SFV-mIL-12 przy moi 10, 100 lub 1000 spowodowało ekspresję mil-12 w sposób zaleŝny od dawki (Figura 3A). Poziom ekspresji mil-12 w komórkach zakaŝonych przy moi 10 AFP-SFV-mIL-12 był porównywalny z poziomem uzyskanym w komórkach zakaŝonych AFP-mIL-12 przy moi Ponadto poziom mil-12 w komórkach HCC zakaŝonych AFP-SFV-mIL-12 przy róŝnych moi był porównywalny z tym uzyskanym z kontrolnym wektorem Ad-CMVmIL-12. JednakŜe zakaŝenie komórek nieeksprymujących AFP plazmidem AFP-mIL-12 lub AFP-SFV-mIL-12 nie wytworzyło dającego się wykryć poziomu mil-12, nawet przy najwyŝszej moi (1000) (Figura 3B). W tych komórkach tylko kontrolny wektor Ad- CMV-mII-12 był w stanie wytworzyć wysoki poziom ekspresji mil-12. [0072] Z drugiej strony cztery linie komórek HCC (Hep3B, Huh-7, HepG2 i PLC/PRF/5) zakaŝono wektorami hybrydowymi LacZ - AFP-lacZ lub AFP-SFV-lacZ przy róŝnych moi (10, 100 lub 1000) oraz oznaczono specyficzną ekspresję β-gal. W tym przypadku uzyskano równiez podobne dane, których wyniki przedstawiono na Figurze 4. [0073] Figura 5 przedstawia mikrofotografie komórek HCC zakaŝonych wektorami adenowirusowymi typu gutless AFP-lacZ i AFP-SFV-lacZ, a następnie wybarwionych X-gal. ZakaŜenie komórek HCC AFP-lacZ wytworzyło niski poziom ekspresji w zakaŝonych komórkach, które wybarwiły się tylko w słabym stopniu. W odróŝnieniu od tego zakaŝenie komórek HCC AFP-SFV-lacZ doprowadziło do wysokiego poziomu ekspresji β-gal, odzwierciedlonego przez intensywne wybarwienie X-gal. Dane te wskazują, Ŝe wektor hybrydowy Ad-SFV zawierający replikon SFV pod kontrolą promotora AFP moŝe wytworzyć wysoki poziom intensywności ekspresji transgenicznej w komórkach nowotworowych eksprymujących AFP. Przebieg w czasie ekspresji mil-12 w komórkach HCC in vitro [0074] W celu zbadania wytwarzania mil-12 w róŝnych punktach czasowych po zakaŝeniu wektorami hybrydowymi Ad-SFV dwie linie komórek HCC (Hep3B i Huh-7) zakaŝo-
22 21 5 no AFP-mIL-12 lub AFP-SFV-mIL-12 oraz supernatanty zbierano codziennie przez 5 dni od zakaŝenia. Figura 6 przedstawia wyniki ekspresji transgenicznej uzyskanej po zakaŝeniu wspomnianych komórek. Wyniki te odzwierciedlają stały wzrost ekspresji mil-12 od dnia 1 do dnia 4 po zakaŝeniu w komórkach zakaŝonych AFP-SFV-mIL-12 (Figura 6). JednakŜe w dniu 5 po zakaŝeniu, poziomy mil-12 nieznacznie się obniŝyły. W komórkach zakaŝonych AFP-mIL-12 poziomy ekspresji były bardzo niskie i z czasem zaobserwowano tylko niewielki wzrost wytwarzania mil-12. Testy cytotoksyczności ocena proliferacji komórek przez włączanie MTT [0075] Komórki HCC (Hep3B, Huh7, MCH-RH7777, Hep1-6) zakaŝono AFP-LacZ, AFP- SFV-LacZ, AFP-mIL-12, AFP-SFV-mIL-12 lub Ad/CMV-mIL-12 przy moi Pięć dni po zakaŝeniu oznaczono przeŝycie komórek w teście MTT (bromek 3-(4,5-dimetylotiazolilo)-2,5-difenylotetrazoliowy) Mosmann, T. (1983) J. Immunol. Meth. 65, 55-63; Tada, H. i in. (1986) J. Immunol. Meth. 93, W skrócie, komórki przemyto dwukrotnie PBS oraz do kaŝdej studzienki (w płytce 48-studzienkowej) dodano 200 µl świeŝo przygotowanego roztworu barwnika MTT. Komórki dalej hodowano przez 3-4 godziny, a następnie dodano 500 µl buforu rozpuszczającego. 100 µl kaŝdej próbki pobrano do pomiarów absorbancji w spektrofotometrze przy długości fali 570 nm. Wywoływanie śmierci komórkowej po zakaŝeniu komórek HCC wektorami hybrydowymi Ad-SFV in vitro [0076] Opisano, Ŝe replikacja wektorów SFV wywołuje śmierć komórkową na drodze apoptozy w większości komórek pochodzących od kręgowców. W celu ustalenia, czy proces ten występuje takŝe w komórkach HCC zakaŝonych wektorami hybrydowymi Ad- SFV, komórki Hep3B i Huh-7 zakaŝono tymi wektorami i oznaczono przeŝycie komórek w dniu 5 po zakaŝeniu. Jak moŝna zaobserwować na Figurze 7, przeŝycie w tym punkcie czasowym po zakaŝeniu było mniejsze niŝ 20% w komórkach zakaŝonych AFP-SFV-mIL- 12 lub AFP-SFV-lacZ. JednakŜe zakaŝenie tych samych komórek AFP-mIL-12 lub AFPlacZ lub wektorem kontrolnym AdCMVmIL-12 nie wpłynęło na przeŝycie komórek. Wyniki te wskazują, Ŝe SFV replikuje się w komórkach zakaŝonych wektorami AFP-SFV. Wykrywanie SFV Rep metodą immunofluorescencji 30 [0077] Komórki HCC (Hep3B, Huh7, MCH-RH7777) wysiano na szkiełka nakrywkowe w płytkach 6-studzienkowych (1x10 5 komórek/studzienkę) oraz zakaŝono AFP-mIL-12, AFP-SFV-mIL-12 lub Ad/CMV-mIL-12 przy moi Dwa dni po zakaŝeniu szkiełka nakrywkowe przemyto dwukrotnie PBS i komórki utrwalono w metanolu w -20 C przez 6
23 minut. Płytki ponownie przemyto trzykrotnie PBS i inkubowano w temperaturze pokojowej (RT) przez 30 min. z PBS zawierającym 0,5% Ŝelatyny i 0,25% BSA w celu wyblokowania niespecyficznego wiązania. Następnie bufor blokujący zastąpiono przeciwciałem pierwszorzędowym (MAb przeciw-replikazie) rozcieńczonym 1:10 w buforze blokujacym i inkubowano w RT przez 30 min. Komórki ponownie przemyto trzykrotnie PBS-0,25% BSA i inkubowano przez 30 min w RT z przeciwciałem drugorzędowym (królicza surowica przeciwmysia sprzęŝona z FITC, Sigma) rozcieńczonym 1:250 w buforze blokującym. Na koniec komórki przemyto trzykrotnie PBS-0,25% BSA, raz wodą i umieszczono na szkiełkach podstawowych z zastosowaniem Vecta shield z Dapi w celu wybarwienia jąder komórkowych. Ekspresja SFV Rep w komórkach HCC zakaŝonych wektorami hybrydowymi Ad-SFV in vitro [0078] Ekspresję SFV Rep analizowano w komórkach HCC zakaŝonych wektorami hybrydowymi Ad-SFV przez zastosowanie immunofluorescencji z przeciwciałem monoklonalnym specyficznym względem tego białka. Figura 8 pokazuje, Ŝe komórki HCC zakaŝone AFP-SFV-mIL-12 lub AFP-SFV-lacZ były intensywnie pozytywne cytoplazmatycznie względem Rep. W odróŝnieniu od tego komórki zakaŝone AFP-mIL-12 lub AFP-lacZ nie wykazały Ŝadnego wybarwienia. Doświadczenia in vivo Indukcja ksenoprzeszczepów HCC oraz badanie wydajności i specyficzności przenoszenia genów in vivo [0079] Komórki Huh-7 zebrano i przemyto dwukrotnie poŝywką wolną od surowicy. 2 x 10 6 komórek przeprowadzono w zawiesinę w 100 µl surowicy z solą fizjologiczną i wstrzyknięto podskórnie (s.c.) do prawego boku nagich myszy BALB/c. Cztery tygodnie po wszczepieniu komórek oraz gdy guzki nowotworowe osiągnęły średnicę 6-8 mm, wstrzyknięto donowotworowo 1x10 10 cząstek wirusa AFP-LacZ (n=4) lub AFP-SFV-LacZ (n=4) rozcieńczonych w 80 µl soli fizjologicznej. Zwierzętom kontrolnym (n=3) wstrzyknięto donowotworowo 80 µl soli fizjologicznej. Myszy uśmiercono w dniu 3 lub 6 po zaszczepieniu. W tych punktach czasowych od kaŝdego zwierzęcia pobrano nowotwory i wątroby, zatopiono je w O.C.T. (Sakura, Holandia) i zamroŝono w -80 C. ZamroŜone tkanki pocięto na skrawki histologiczne i umieszczono na szkiełkach podstawowych, które wybarwiono X-gal lub analizowano w teście TUNEL. W celu zbadania specyficzności zaka- Ŝenia wektorem hybrydowym in vivo, normalnym myszom Balb/c wstrzyknięto doŝylnie AFP-LacZ (n=4), AFP-SFV-LacZ (n=4) lub Ad/CMV-LacZ w dawce cząstek wiru-
24 23 sa/mysz. Zwierzęta uśmiercono trzeciego dnia po zakaŝeniu i zebrano główne narządy w celu zanalizowania ekspresji LacZ przez zastosowanie X-gal. Wydajność przenoszenia genów wektorów hybrydowych Ad-SFV w ludzkich ksenoprzeszczepach HCC w nagich myszach z niedoborem odporności [0080] W celu zbadania skuteczności transdukcji wektorami hybrydowymi typu gutless in vivo zastosowano model ludzkich komórek HCC oparty na komórkach Huh7 zdolnych do ekspresji AFP. Komórki Huh7 wszczepiono podskórnie nagim myszom Balb/c z niedoborem odporności i po wytworzeniu guzków nowotworowych po 30 dniach zwierzętom wstrzyknięto donowotworowo 1x10 10 cząstek wirusa AFP-SFV-LacZ lub AFP-LacZ jako kontroli. Myszy uśmiercono 3 lub 6 dni po wstrzyknięciu wirusa i usunięto zarówno nowotwór, jak i wątrobę i zbadano je przez wybarwienie X-gal. Jak widać na Figurze 9A, stwierdzono słabą ekspresję transgeniczna w skrawkach guza u zwierząt, które otrzymały AFP-lacZ. W odróŝnieniu od tego u zwierząt, które otrzymały AFP-SFV-LacZ, stwierdzono intensywną ekspresję LacZ w skrawkach guza (Figura 9B). Nie stwierdzono ekspresji transgenu w skrawkach wątroby pobranej od zwierząt, które otrzymały zarówno AFP- LacZ jak i AFP-SFV-LacZ, co wskazuje, Ŝe wektory były prawdopodobnie ograniczone do miejsca wszczepienia (dane nieprzedstawione). W celu zbadania, czy wektory hybrydowe Ad-SFV wywołują apoptozę w zakaŝonych komórkach nowotworowych, skrawki gózów analizowano z zastosowaniem techniki TUNEL. Nie zaobserwowano apoptozy w próbkach pobranych od myszy zaszczepionych AFP-LacZ (Figura 9C). JednakŜe w guzach zwierząt, które otrzymały AFP-SFV-LacZ, zaobserwowano znaczną liczbę komórek apoptotycznych (Figura 9D). Dane te wskazują, Ŝe wektory hybrydowe Ad-SFV nie tylko wywołują ekspresję genów specyficznie w komórkach nowotworowych, ale takŝe wybiórczo wywołują w tych samych komórkach śmierć komórkową na drodze apoptozy. Specyficzność wektorów hybrydowych Ad-SFV in vivo [0081] Aby wykazać specyficzność wektorów hybrydowych, cząstek wirusa AFP- LacZ, AFP-SFV-LacZ lub wektora kontrolnego Ad/CMV-LacZ podano doŝylnie myszom Balb/c. Trzy dni po podaniu wektorów analizowano ekspresję β-galaktozydazy w wątrobie. Jak przedstawiono na Figurze 9 (E-F), ani wektor AFP-LacZ ani AFP-SFV-LacZ nie był w stanie wywołać wykrywalnej ekspresji transgenu w wątrobie. JednakŜe wysoką proporcję komórek pozytywnych względem β-galaktozydazy w skrawkach tkanki wątroby zaobserwowano u tych zwierząt, które otrzymywały Ad/CMV-LacZ (Figura 9G). Dane te potwierdzają, Ŝe ekspresja za pośrednictwem wektorów hybrydowych jest specyficzna dla komórek nowotworowych.
25 24 Ortotopowe wywołanie HCC oraz terapia genowa in vivo 5 10 [0082] 5 x 10 5 McA-RH7777 komórek wszczepiono w lewy płat wątroby szczurów Buffalo. Dziesięć dni po wszczepieniu komórek nowotworowych u kaŝdego zwierzęcia obserwowano pojawienie się pojedynczego guzka nowotworowego o średnicy 7-10 mm. Nowotwory potraktowano lub 2x10 11 cząstek wirusa AFP-mIL-12, AFP-SFV-mIL-12 lub roztworem soli fizjologicznej jako kontrolą. Dwa i cztery tygodnie po leczeniu zwierzęta uśpiono i poddano laparotomii w celu obserwacji rozwoju nowotworu. Przeprowadzono takŝe analizę przeŝycia zwierząt. Wielkość guzów oszacowano przez pomiar długości i szerokości kaŝdego guzka stosując wzór: Objętość guza = (długość mm) X (szerokość mm) 2 x 0,5236 (Janik i in., 1975). Wydajność ortotopowego leczenia HCC u szczurów Buffalo [0083] W celu zbadania skuteczności przeciwnowotworowej wektora hybrydowego Ad- SFV niosącego IL-12 załoŝono ortotopowe nowotwory HCC u szczurów przez wszczepienie komórek McH-RH7777 do wątroby. Model ten wybrano, poniewaŝ uprzednio wykazano, Ŝe komórki McH-RH7777 eksprymują AFP. W pierwszym doświadczeniu zwierzęta potraktowano przez pojedyncze wstrzyknięcie donowotworowo 1x10 11 cząstek wirusa AFP-mIL-12, AFP-SFV-mIL-12 lub roztworu soli fizjologicznej jako kontroli (Figura 10A-C). Zwierzęta, które otrzymywały AFP-mIL-12, wykazywały zmniejszoną wielkość guza w porównaniu ze zwierzętami kontrolnymi, u który zaobserwowano stały wzrost wielkości guza w czasie przebiegu doświadczenia (Figura 10A-B). Natomiast traktowanie AFP-SFV-mIL-12 spowodowało całkowite cofnięcie się nowotworu u 1 z 4 leczonych szczurów, stabilizację choroby u dwóch szczurów oraz brak odpowiedzi u jednego zwierzęcia (Figura 10C). W celu potwierdzenia, czy większe dawki wektora hybrydowego mogą zwiększyć działanie przeciwnowotworowe, przeprowadzono drugie doświadczenie w którym zwierzęta potraktowano donowotworowo dawką 2 x cząstek wirusa AFP-mIL- 12, AFP-SFV-mIL-12 lub roztworem soli fizjologicznej jako kontrolą (Figura 10 D-G). Tak jak w poprzednim doświadczeniu zwierzęta, które otrzymywały wektor AFP/IL-12, wykazały tylko niewielką odpowiedź przeciwnowotworową, która oznaczała jedną całkowitą remisję choroby, guzy rosnące wolniej niŝ u kontroli u 4 zwierząt oraz brak odpowiedzi u 7 zwierząt, wśród całkowitej liczby 12 leczonych zwierząt (Figura 10E). Natomiast leczenie wektorem AFP-SFV-IL-12 wywołał duŝo silniejsze działanie, wywołując całkowite ustąpienie nowotworu u czterech zwierząt (33%), częściowe ustąpienie u 6 zwierząt (50%), opóźniony postęp wzrostu guza u 2 zwierząt (16%) oraz brak odpowiedzi u 2 innych zwierząt (16%), wśród całkowitej liczby 12 leczonych zwierząt (Figura 10F). W tym
26 25 drugim badaniu wektor AFP/SFV-IL-12 pozwolił na przeŝycie 50% leczonych zwierząt, w stosunku do 0% przeŝycia wśród zwierząt leczonych AFP-IL-12 lub roztworem soli fizjologicznej (Figura 10G) Badanie toksyczności in vivo: oznaczenie transaminaz i poziomów Il-12 w surowicy oraz ocena histologiczna wątroby [0084] Próbki krwi zebrano od szczurów leczonych donowotworowo wektorami adenowirusowymi AFP-SFV-IL-12 lub AFP-IL-12 w dawce 2 x lub roztworem soli fizjologicznej 4 i 8 po zaszczepieniu. Badanie to obejmowało takŝe szczury szczepione 10 8 cząstek alfawirusa SFV-IL-12. Surowicę oddzielono od krwi przez wirowanie przy 2000 obrotach/minutę przez 15 minut. Poziomy transaminaz oznaczono z zastosowaniem analizatora Hitachi 911 Automatic Analyzer (Boehringer Mannheim, Niemcy). Poziomy IL-12 oznaczono z zastosowaniem testu ELISA. Badanie histologiczne przeprowadzono przez pobranie wątroby od zwierząt zaszczepionych trzy dni po podaniu leczenia. Narządy utrwalono w formolu, zatopiono w parafinie i pokrojono na skrawki grubości 6 mikronów z zastosowaniem mikrotomu. Skrawki te wybarwiono hematoksyliną/eozyną. Badanie toksyczności wektorów hybrydowych Ad-SFV u szczurów [0085] W celu oceny toksyczności związanej z podawaniem wektora hybrydowego AFP- SFV-IL-12, oznaczono poziomy transaminaz (GOT, GPT i GGTL) w surowicy szczurów leczonych donowotworowo 2 x cząstek wirusa róŝnych wektorów (patrz powyŝsza część). Badanie to takŝe obejmowało grupę szczurów szczepionych równieŝ donowotworowo 10 8 cząstek wirusa wektora alfawirusowego SFV-IL-12 (Figura 11A). Szczury zaszczepione wektorami adenowirusowymi AFP-SFV-IL-12 lub AFP-IL-12 wykazywały bardzo niski poziom transaminaz, który był podobny do poziomu u zwierząt kontrolnych zaszczepionych roztworem soli fizjologicznej. JednakŜe poziomy transaminaz u zwierząt traktowanych cząstkami SFV-IL-12 były istotnie wyŝsze niŝ w innych grupach (p<0,05). W badaniu tym wykonano takŝe oznaczenia poziomu IL-12 obecnej w surowicy zwierząt w tych samych punktach czasowych. Nie wykryto IL-12 w surowicy zwierząt zaszczepionych wektorami Ad AFP-SFV-IL-12 lub AFP-IL-12 lub roztworem soli fizjologicznej (Figura 11B), co oznacza, Ŝe ekspresja transgenu w tych wektorach jest ograniczona do nowotworów oraz sugeruje, Ŝe toksyczność wektora hybrydowego Ad-SFV jest bardzo niska. JednakŜe leczenie cząstkami wirusa SFV-IL-12 wywołało wysoki poziom IL-12 w surowicy po krótkim okresie czasu, która to sytuacja moŝe spowodować toksyczność dla wątroby. Ostatecznie badanie toksyczności zakończono analizą histologiczną wybarwionych hematoksyliną/eozyną skrawków wątroby pochodzących od szczurów leczonych donowotwo-
27 rowo tymi samymi wektorami oraz w dawkach, takich samych jak juŝ opisano (Figura 12). Badanie to wykazało brak róŝnic histologicznych pomiędzy szczurami otrzymującymi roztwór soli fizjologicznej, a tymi, które otrzymywały wektory adenowirusowe AFP-SFV-IL- 12 lub AFP-IL-12. Natomiast obszary z hepatocytami eozynochłonnymi, a takŝe fuzję tych hepatocytów, obserwowano w skrawkach wątroby szczurów leczonych cząstkami wirusa SFV-IL-12, co wskazuje na pewien stopień toksyczności (czarne strzałki, Figura 12D). WYKAZ SEKWENCJI [0086] <110> Proyecto de Biomedicina CIMA S.L. <120> HYBRYDOWY WEKTOR ADENOWIRUSOWY/ALFAWIRUSOWY DO SKUTECZNEGO PODAWANA I EKSPRESJI GENÓW TERAPEUTYCZNYCH W KOMÓRKACH NOWOTWOROWYCH <160> 15 <170> PatentIn wersja <210> 1 <211> 438 <212> DNA <213> Adenowirus <400> <210> 2 <211> 161 <212> DNA <213> Adenowirus <400> 2
28 27 5 <210> 3 <211> 7412 <212> DNA <213> Wirus Semliki Forest (SFV) <400> 3
29 28
30 29
31 30
32 5 31
33 32 <210> 4 <211> 7521 <212> DNA <213> Wirus Semliki Forest (SFV) 5 <400> 4
34 33
35 34
36 35
37 36 5 <210> 5 <211> 841 <212> DNA <213> Wirus Semliki Forest (SFV) <400> 5 10 <210> 6 <211> 260 <212> DNA
38 37 <213> Wirus SV40 <400> 6 5 <210> 7 <211> 940 <212> DNA <213> Homo sapiens <400> <210> 8 <211> <212> DNA <213> Chimeryczny <220> <221> 5' ITR
39 38 <222> (1)..(438) <223> Pierwsza sekwencja odwróconego powtórzenia końcowego oraz sekwencja sygnałowa pakowania <220> <221> Sekwencja wypychająca <222> (439)..(10990) <223> Pierwsza niekodująca sekwencja wypychająca <220> <221> AFP(p+e) <222> (10991)..(11930) <223> Promotor alfafetoproteiny (AFP). Zawiera region promotora (p) oraz region wzmacniacza (e) <220> <221> SFV <222> (12257)..(19366) <223> Region sekwencji replikonu SFV <220> <221> mil-12 <222> (19389)..(21722) <223> Sekwencja genu mysiej interleukiny-12 (IL12) (gen egzogenny) <220> <221> PolyA <222> (22621)..(22880) <223> Sekwencja poliadenylacji pochodząca od wirusa SV <220> <221> Sekwencja wypychająca <222> (22881)..(28731) <223> Druga niekodująca sekwencja wypychająca <220> <221> 3' ITR <222> (28732)..(28892) <223> Sekwencja drugiego odwróconego powtórzenia końcowego <400> 8
40 39
41 40
42 41
43 42
44 43
45 44
46 45
47 46
48 47
49 48
50 49
51 50
52 51
53 52
54 53
55 54
56 <210> 9 <211> <212> DNA <213> Chimeryczny <220> <221> 5' ITR <222> (1)..(438) <223> Pierwsza sekwencja odwróconego powtórzenia końcowego oraz sekwencja sygnałowa pakowania <220> <221> Sekwencja wypychająca <222> (439)..(10905) <223> Pierwsza niekodująca sekwencja wypychająca <220>
57 <221> AFP(p+e) <222> (10906)..(11845) <223> Promotor alfafetoproteiny (AFP). Zawiera region promotora (p) oraz region wzmacniacza (e) <220> <221> SFV <222> (12175)..(19281) <223> Region sekwencji replikonu SFV <220> <221> LacZ <222> (19325)..(22397) <223> Sekwencja genu LacZ Escherichia coli (gen egzogenny jako gen reporterowy) <220> <221> PolyA <222> (23295)..(23554) <223> Sekwencja poliadenylacji pochodząca od wirusa SV40 <220> <221> Relleno <222> (23555)..(29350) <223> Druga niekodująca sekwencja wypychająca <220> <221> 3' ITR <222> (29351)..(29511) <223> Sekwencja drugiego odwróconego powtórzenia końcowego <400> 9
58 57
59 58
60 59
61 60
62 61
63 62
64 63
65 64
66 65
67 66
68 67
69 68
70 69
71 70
72 71
73 5 72
74 <210> 10 <211> <212> DNA <213> Chimeryczny <220> <221> 5' ITR <222> (1)..(438) <223> Pierwsza sekwencja odwróconego powtórzenia końcowego oraz sekwencja sygnałowa pakowania <220> <221> Sekwencja wypychająca <222> (439)..(10990) <223> Pierwsza niekodująca sekwencja wypychająca <220> <221> TERT <222> (10991)..(11285) <223> Promotor telomerazy (TERT) <220> <221> SFV <222> (11556)..(18665) <223> Region sekwencji replikonu SFV <220> <221> mil-12 <222> (18688)..(21021) <223> Sekwencja mysiego genu interleukiny-12 (IL12) <220> <221> PolyA <222> (21920)..(22179) <223> Sekwencja poliadenylacji pochodząca od wirusa SV40 <220> <221> Sekwencja wypychająca <222> (22180)..(28030) <223> Druga niekodująca sekwencja wypychająca
75 74 5 <220> <221> 3' ITR <222> (28031)..(28191) <223> Pierwsza sekwencja odwróconego powtórzenia końcowego <400> 10
76 75
77 76
78 77
79 78
80 79
81 80
82 81
83 82
84 83
85 84
86 85
87 86
88 87
89 88
90 89
91 90 5 <210> 11 <211> 5844 <212> DNA <213> Plazmid <220> <221> MCS <222> (1)..(25) <223> Miejsce wielokrotnego klonowania <220> <221> Wzmacniacz AFP <222> (26)..(820) <223> Wzmacniacz AFP <220> <221> Promotor AFP <222> (828)..(1054) <223> Promotor AFP <400> 11
92 91
93 92
94 5 93
95 94 5 <210> 12 <211> 2334 <212> DNA <213> Mus musculus <400> 12
96 95 5 <210> 13 <211> 2336 <212> DNA <213> Homo sapiens <400> 13
97 96
98 97 5 <210> 14 <211> 3057 <212> DNA <213> Escherichia coli <400> 14
99 98
100 99 5 <210> 15 <211> 336 <212> DNA <213> Homo sapiens <400> 15
101 100 ZastrzeŜenia patentowe Adenowirusowy wektor hybrydowy do ekspresji genów, zawierający co najmniej następujące elementy ustawione w kierunku 5' do 3': i. pierwszy łańcuch pochodzenia adenowirusowego zawierający sekwencję pierwszego odwróconego powtórzenia końcowego (ITR) oraz sekwencję sygnałową pakowania adenowirusa; ii. pierwszą niekodującą sekwencję wypychającą; iii. sekwencję odpowiadającą promotorowi specyficznemu dla tkanki; iv. łańcuch cdna pochodzący od alfawirusa, którego sekwencja jest częściowo komplementarna do sekwencji RNA alfawirusa, zawierający co najmniej sekwencję kodującą co najmniej jeden interesujący gen egzogenny, przy czym wspomniany łańcuch cdna zawiera: a) sekwencję 5' niezbędną do replikacji alfawirusa, b) sekwencję kodującą białka niestrukturalne wymagane do replikacji RNA alfawirusa, c) co najmniej jeden promotor subgenomowy alfawirusa, oraz d) sekwencję 3' niezbędną do replikacji alfawirusa; v. sekwencję poliadenylacji; oraz vi. drugą sekwencję odwróconego powtórzenia końcowego (ITR) adenowirusa. 2. Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 1, który ponadto zawiera element vii, który jest drugą niekodującą sekwencją wypychającą, pomiędzy elementem v i elementem vi. 3. Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 1, w którym elementem ii jest ludzka niekodująca sekwencja wypychająca. 4. Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 1 albo 3, w którym element ii stanowi region intronu ludzkiej genomowej fosforybozylo-
102 transferazy hypoksantynowej (HPRT). 5. Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z zastrzeŝeń 1 do 4, w którym element i ma sekwencję SEQ ID No Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 1, w którym element iii stanowi promotor specyficzny dla nowotworu. 7. Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 6, w którym element iii stanowi promotor specyficzny dla nowotworu, wybrany z AFP, telomerazy TERT, PAP, E2F i HIF. 8. Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 1, w którym element iii stanowi promotor specyficzny dla nowotworu, zawierający sekwencję wybraną z SEQ ID No 7, odpowiadającej AFP p+e, oraz SEQ ID No 15, odpowiadającej telomerazie TERT. 9. Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z zastrzeŝeń 1 do 8, w którym element iv zawiera sekwencję pochodzącą od wirusa Semliki Forest (SFV). 10. Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 1, w którym sekwencje a) do c) elementu iv w całości mają sekwencję wybraną z SEQ ID No 3 i SEQ ID No Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 1 albo 10, w którym element iv d) ma sekwencję SEQ ID No Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z poprzednich zastrzeŝeń, w którym interesujący gen egzogenny jest wybrany z jednego lub większej liczby genów terapeutycznych, jednego lub większej liczby genów reporterowych oraz ich połączeń. 13. Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 12, w którym interesującym genem egzogennym jest gen terapeutyczny ssaczej interleukiny IL Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 12, w którym interesującym genem egzogennym jest gen terapeutyczny ludzkiej interleukiny hil Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 12, w którym interesującym genem egzogennym jest gen terapeutyczny wybrany spośród genów czynnika stymulującego tworzenie kolonii (GMCSF), interferonu alfa oraz kinazy tymidynowej (HSV-TK) wirusa opryszczki zwykłej. 16. Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 10, w
103 którym element iv zawiera kolejno jeden lub kilka podzestawów (promotor subgenomowy + interesujący gen egzogenny). 17. Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 12, w którym interesującym genem egzogennym jest gen reporterowy wybrany z LacZ, Lucyferazy, kinazy tymidynowej HSV-TK wirusa opryszczki zwykłej i GFP. 18. Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z zastrzeŝeń 9 do 17, w którym element iv tworzy replikon funkcjonalnie kontrolowany przez promotor iii oraz w którym alfawirusowy promotor subgenomowy zawarty w iv.c) funkcjonalnie kontroluje ekspresję interesującego genu egzogennego. 19. Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z poprzednich zastrzeŝeń, w którym elementem v jest sekwencja poliadenylacji SV Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z poprzednich zastrzeŝeń, w którym element v ma sekwencję SEQ ID No Adenowirusowy wektor hybrydowy do ekspresji genów według zastrzeŝenia 2, w którym drugą niekodującą sekwencją wypychającą jest C Wektor adenowirusowy do ekspresji genów według któregokolwiek z poprzednich zastrzeŝeń, w którym element vi. ma sekwencję SEQ ID No Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z poprzednich zastrzeŝeń, który zawiera: i. pierwszy łańcuch pochodzenia adenowirusowego zawierający sekwencję pierwszego odwróconego powtórzenia końcowego (ITR) oraz sekwencję sygnałową pakowania adenowirusa; ii. pierwszą niekodującą sekwencję wypychającą, którą stanowi region intronu ludzkiej genomowej fosforybozylotransferazy hypoksantynowej (HPRT) iii. sekwencję odpowiadającą promotorowi specyficznemu dla tkanki, którą stanowi promotor AFP, iv. łańcuch cdna pochodzący od alfawirusa, którego sekwencja jest częściowo komplementarna do alfawirusowego RNA pochodzącego od wirusa SFV, który zawiera sekwencję kodującą interesujący gen egzogenny, który stanowi hil-12, przy czym wspomniany łańcuch cdna zawiera: a) sekwencję 5' niezbędną do replikacji alfawirusa, b) sekwencję kodującą białka niestrukturalne wymagane do replikacji RNA alfawirusa, c) co najmniej jeden promotor subgenomowy alfawirusa, oraz d) sekwencję 3' niezbędną do replikacji alfawirusa;
104 v. sekwencję poliadenylacji SV40, vi. drugą sekwencję odwróconego powtórzenia końcowego (ITR) adenowirusa, oraz vii. drugą niekodującą sekwencję wypychającą, którą jest ludzka sekwencja genomowa C346, umiejscowiona pomiędzy elementem v a elementem vi. 24. Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z poprzednich zastrzeŝeń, który zawiera: i. pierwszy łańcuch pochodzenia adenowirusowego zawierający sekwencję pierwszego odwróconego powtórzenia końcowego (ITR) oraz sekwencję sygnałową pakowania adenowirusa; ii. pierwszą niekodującą sekwencję wypychającą, którą stanowi region intronu ludzkiej genomowej fosforybozylotransferazy hypoksantynowej (HPRT); iii. sekwencję odpowiadającą promotorowi specyficznemu dla tkanki, którą stanowi promotor AFP, iv. łańcuch cdna pochodzący od alfawirusa, którego sekwencja jest częściowo komplementarna do alfawirusowego RNA pochodzącego od wirusa SFV, który zawiera sekwencję kodującą interesujący gen egzogenny, który stanowi mil-12 i LacZ, przy czym wspomniany łańcuch cdna zawiera: a) sekwencję 5' niezbędną do replikacji alfawirusa, b) sekwencję kodującą białka niestrukturalne wymagane do replikacji RNA alfawirusa, c) co najmniej jeden promotor subgenomowy alfawirusa, oraz d) sekwencję 3' niezbędną do replikacji alfawirusa; v. sekwencję poliadenylacji SV40, vi. drugą sekwencję odwróconego powtórzenia końcowego (ITR) adenowirusa, oraz vii. drugą niekodującą sekwencję wypychającą, którą jest ludzka sekwencja genomowa C346, umiejscowiona pomiędzy elementem v a elementem vi. 25. Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z poprzednich zastrzeŝeń, w którym wspomniany wektor ma długość w zakresie pomiędzy 27 a 38 tysięcy par zasad. 26. Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z poprzednich zastrzeŝeń, w którym wspomniany wektor ma sekwencję SEQ ID No Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z zastrzeŝeń 1 do 25, w którym wspomniany wektor ma sekwencję SEQ ID No 9.
105 Adenowirusowy wektor hybrydowy do ekspresji genów według któregokolwiek z zastrzeŝeń 1 do 25, w którym wspomniany wektor ma sekwencję SEQ ID No Zastosowanie adenowirusowego wektora hybrydowego według któregokolwiek z powyŝszych zastrzeŝeń 1 do 28 w sposobie przenoszenia materiału genetycznego do komórki in vitro, w którym wspomnianą komórką nie jest komórka ludzkiej linii zarodkowej. 30. Zastosowanie według zastrzeŝenia 29, w którym wspomnianą komórkę stanowi komórka nowotworowa. 31. Zastosowanie według zastrzeŝenia 30, w którym wspomnianą komórkę stanowi komórka nowotworowa eksprymującą AFP. 32. Zastosowanie adenowirusowego wektora hybrydowego zdefiniowanego w którymkolwiek z zastrzeŝeń 1 do 28 do wytwarzania leku do leczenia nowotworów. 33. Adenowirusowy wektor hybrydowy zdefiniowany w którymkolwiek z zastrzeŝeń 1 do 28 do stosowania w leczeniu nowotworów. 34. Zastosowanie adenowirusowego wektora hybrydowego zdefiniowanego w którymkolwiek z zastrzeŝeń 1 do 28 do wytwarzania leku do wywoływania odpowiedzi immunologicznej przeciw obcemu antygenowi. 35. Adenowirusowy wektor hybrydowy zdefiniowany w którymkolwiek z zastrzeŝeń 1 do 28, do stosowania w wywoływaniu odpowiedzi immunologicznej przeciw obcemu antygenowi. 36. Kompozycja farmaceutyczna, która zawiera co najmniej jeden adenowirusowy wektor hybrydowy zdefiniowany w którymkolwiek z zastrzeŝeń 1 do Kompozycja farmaceutyczna, która zawiera co najmniej jeden adenowirusowy wektor hybrydowy zdefiniowany w którymkolwiek z zastrzeŝeń 1 do 28, w którym interesującym genem egzogennym jest hil-12. Uprawniony: Pełnomocnik: Proyecto de Biomedicina Cima, S.L. mgr inŝ. Zofia Sulima Rzecznik patentowy
106 105
107 106
108 107
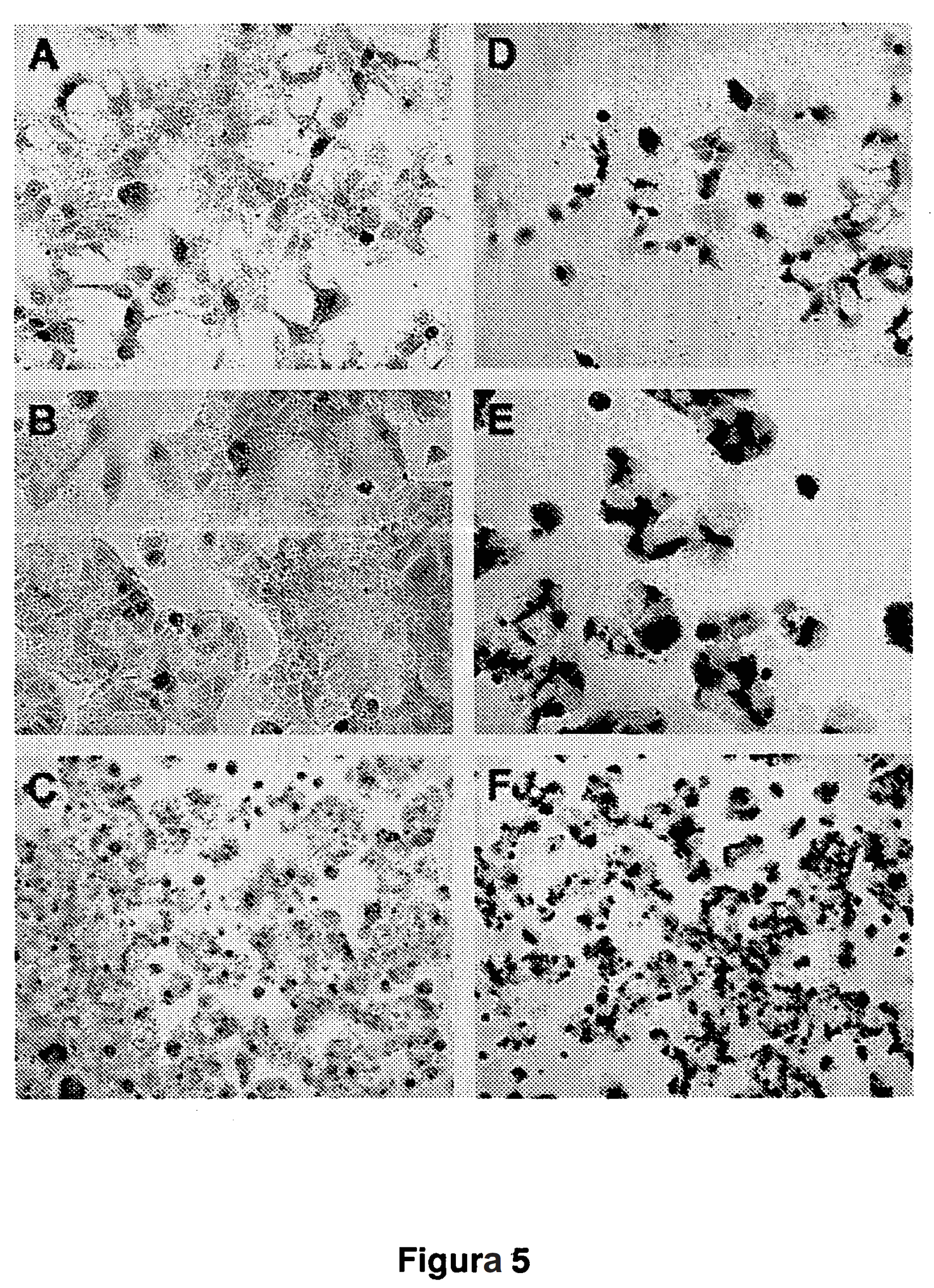
109 108
110 109
111 110
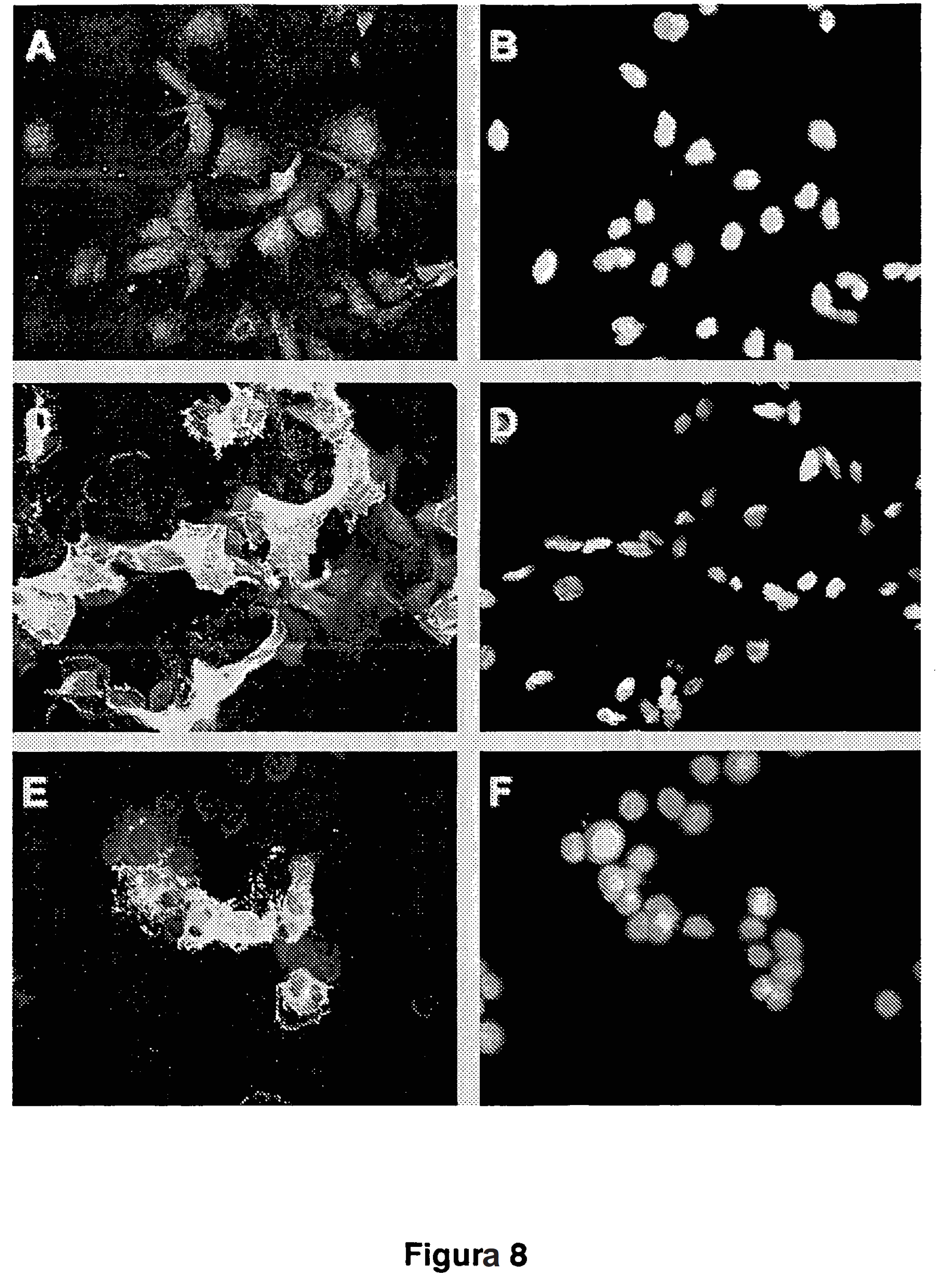
112 111
Nowoczesne systemy ekspresji genów
 Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE. Ewa Waszkowska ekspert UPRP
 WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
Klonowanie molekularne Kurs doskonalący. Zakład Geriatrii i Gerontologii CMKP
 Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Dr. habil. Anna Salek International Bio-Consulting 1 Germany
 1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
Ekspresja białek w komórkach ssaczych
 Ekspresja białek w komórkach ssaczych Ekspresja białek w komórkach ssaczych Używane powszechnie w laboratoriach ssacze linie komórkowe są zmodyfikowane w celu pełnienia roli gospodarza ekspresji białek
Ekspresja białek w komórkach ssaczych Ekspresja białek w komórkach ssaczych Używane powszechnie w laboratoriach ssacze linie komórkowe są zmodyfikowane w celu pełnienia roli gospodarza ekspresji białek
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680075 (13) T3 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2004
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680075 (13) T3 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2004
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1767642 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 19.11.2004 04797352.4
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1767642 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 19.11.2004 04797352.4
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1747298 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.7 (51) Int. Cl. C22C14/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1747298 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.7 (51) Int. Cl. C22C14/00 (2006.01)
Zawartość. Wstęp 1. Historia wirusologii. 2. Klasyfikacja wirusów
 Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
Ćwiczenia 1 Wirtualne Klonowanie Prowadzący: mgr inż. Joanna Tymeck-Mulik i mgr Lidia Gaffke. Część teoretyczna:
 Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
Inżynieria genetyczna- 6 ECTS. Inżynieria genetyczna. Podstawowe pojęcia Część II Klonowanie ekspresyjne Od genu do białka
 Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2120618. (96) Data i numer zgłoszenia patentu europejskiego: 28.02.2008 08719309.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2120618. (96) Data i numer zgłoszenia patentu europejskiego: 28.02.2008 08719309.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2120618 (96) Data i numer zgłoszenia patentu europejskiego: 28.02.2008 08719309.0 (13) (1) T3 Int.Cl. A41B 11/02 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2120618 (96) Data i numer zgłoszenia patentu europejskiego: 28.02.2008 08719309.0 (13) (1) T3 Int.Cl. A41B 11/02 (2006.01)
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii
 Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych aspekty praktyczne dr Małgorzata Kozłowska Ekspert
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych aspekty praktyczne dr Małgorzata Kozłowska Ekspert
TERAPIA GENOWA. dr Marta Żebrowska
 TERAPIA GENOWA dr Marta Żebrowska Pracownia Diagnostyki Molekularnej i Farmakogenomiki, Zakładu Biochemii Farmaceutycznej, Uniwersytetu Medycznego w Łodzi Źródło zdjęcia: httpblog.ebdna.plindex.phpjednoznajwiekszychzagrozenludzkosciwciazniepokonane
TERAPIA GENOWA dr Marta Żebrowska Pracownia Diagnostyki Molekularnej i Farmakogenomiki, Zakładu Biochemii Farmaceutycznej, Uniwersytetu Medycznego w Łodzi Źródło zdjęcia: httpblog.ebdna.plindex.phpjednoznajwiekszychzagrozenludzkosciwciazniepokonane
S YL AB US MODUŁ U ( PRZEDMIOTU) I nforma c j e ogólne
 I nforma c j e ogólne") Załącznik Nr 3 do Uchwały Senatu PUM 14/2012 S YL AB US MODUŁ U ( PRZEDMIOTU) I nforma c j e ogólne Kod modułu Rodzaj modułu Wydział PUM Kierunek studiów Specjalność Poziom studiów Nazwa modułu INŻYNIERIA
Załącznik Nr 3 do Uchwały Senatu PUM 14/2012 S YL AB US MODUŁ U ( PRZEDMIOTU) I nforma c j e ogólne Kod modułu Rodzaj modułu Wydział PUM Kierunek studiów Specjalność Poziom studiów Nazwa modułu INŻYNIERIA
Dane mikromacierzowe. Mateusz Markowicz Marta Stańska
 Dane mikromacierzowe Mateusz Markowicz Marta Stańska Mikromacierz Mikromacierz DNA (ang. DNA microarray) to szklana lub plastikowa płytka (o maksymalnych wymiarach 2,5 cm x 7,5 cm) z naniesionymi w regularnych
Dane mikromacierzowe Mateusz Markowicz Marta Stańska Mikromacierz Mikromacierz DNA (ang. DNA microarray) to szklana lub plastikowa płytka (o maksymalnych wymiarach 2,5 cm x 7,5 cm) z naniesionymi w regularnych
Wektory DNA - klonowanie molekularne
 Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609. (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609. (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609 (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.2 (13) (51) T3 Int.Cl. A61K 38/17 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609 (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.2 (13) (51) T3 Int.Cl. A61K 38/17 (2006.01)
Klonowanie i transgeneza. dr n.med. Katarzyna Wicher
 Klonowanie i transgeneza dr n.med. Katarzyna Wicher Klonowanie proces tworzenia idealnej kopii z oryginału oznacza powstawanie lub otrzymywanie genetycznie identycznych: cząsteczek DNA komórek organizmów
Klonowanie i transgeneza dr n.med. Katarzyna Wicher Klonowanie proces tworzenia idealnej kopii z oryginału oznacza powstawanie lub otrzymywanie genetycznie identycznych: cząsteczek DNA komórek organizmów
KLONOWANIE DNA REKOMBINACJA DNA WEKTORY
 KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2300459. (96) Data i numer zgłoszenia patentu europejskiego: 22.06.2009 09772333.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2300459. (96) Data i numer zgłoszenia patentu europejskiego: 22.06.2009 09772333.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2049 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.06.09 09772333.2 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2049 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.06.09 09772333.2 (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 161679 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 24.06.0 064.7 (1) Int. Cl. B60R21/01 (06.01) (97) O udzieleniu
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 161679 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 24.06.0 064.7 (1) Int. Cl. B60R21/01 (06.01) (97) O udzieleniu
TATA box. Enhancery. CGCG ekson intron ekson intron ekson CZĘŚĆ KODUJĄCA GENU TERMINATOR. Elementy regulatorowe
 Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 71811 (96) Data i numer zgłoszenia patentu europejskiego: 29.09.06 06791167.7 (13) (1) T3 Int.Cl. H04Q 11/00 (06.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 71811 (96) Data i numer zgłoszenia patentu europejskiego: 29.09.06 06791167.7 (13) (1) T3 Int.Cl. H04Q 11/00 (06.01) Urząd
mikrosatelitarne, minisatelitarne i polimorfizm liczby kopii
 Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1711158 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 16.11.2004 04806793.8
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1711158 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 16.11.2004 04806793.8
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2044552. (96) Data i numer zgłoszenia patentu europejskiego: 11.05.2007 07719230.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2044552. (96) Data i numer zgłoszenia patentu europejskiego: 11.05.2007 07719230.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2044552 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.05.2007 07719230.0
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2044552 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.05.2007 07719230.0
TECHNIKI ANALIZY RNA TECHNIKI ANALIZY RNA TECHNIKI ANALIZY RNA
 DNA 28SRNA 18/16S RNA 5SRNA mrna Ilościowa analiza mrna aktywność genów w zależności od wybranych czynników: o rodzaju tkanki o rodzaju czynnika zewnętrznego o rodzaju upośledzenia szlaku metabolicznego
DNA 28SRNA 18/16S RNA 5SRNA mrna Ilościowa analiza mrna aktywność genów w zależności od wybranych czynników: o rodzaju tkanki o rodzaju czynnika zewnętrznego o rodzaju upośledzenia szlaku metabolicznego
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ 1. Gen to odcinek DNA odpowiedzialny
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ 1. Gen to odcinek DNA odpowiedzialny
Wykład 14 Biosynteza białek
 BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
października 2013: Elementarz biologii molekularnej. Wykład nr 2 BIOINFORMATYKA rok II
 10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1477128 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 14.05.2004 04076445.8 (51) Int. Cl. A61D1/02 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1477128 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 14.05.2004 04076445.8 (51) Int. Cl. A61D1/02 (2006.01)
Inżynieria genetyczna
 Inżynieria genetyczna i technologia rekombinowanego DNA Dr n. biol. Urszula Wasik Zakład Biologii Medycznej Inżynieria genetyczna świadoma, celowa, kontrolowana ingerencja w materiał genetyczny organizmów
Inżynieria genetyczna i technologia rekombinowanego DNA Dr n. biol. Urszula Wasik Zakład Biologii Medycznej Inżynieria genetyczna świadoma, celowa, kontrolowana ingerencja w materiał genetyczny organizmów
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii
 Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
Badanie funkcji genu
 Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886669 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 02.08.2007 07113670.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886669 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 02.08.2007 07113670.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2190940 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.09.2008 08802024.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2190940 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.09.2008 08802024.3
TRANSKRYPCJA - I etap ekspresji genów
 Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
Agenda: Wynalazek jako własnośd intelektualna. Co może byd wynalazkiem. Wynalazek biotechnologiczny. Ochrona patentowa
 dr Bartosz Walter Agenda: Wynalazek jako własnośd intelektualna Co może byd wynalazkiem Wynalazek biotechnologiczny Ochrona patentowa Procedura zgłoszenia patentowego Gdzie, kiedy i jak warto patentowad
dr Bartosz Walter Agenda: Wynalazek jako własnośd intelektualna Co może byd wynalazkiem Wynalazek biotechnologiczny Ochrona patentowa Procedura zgłoszenia patentowego Gdzie, kiedy i jak warto patentowad
Biologia Molekularna z Biotechnologią ===============================================================================================
 Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
Inżynieria Genetyczna ćw. 3
 Materiały do ćwiczeń z przedmiotu Genetyka z inżynierią genetyczną D - blok Inżynieria Genetyczna ćw. 3 Instytut Genetyki i Biotechnologii, Wydział Biologii, Uniwersytet Warszawski, rok akad. 2018/2019
Materiały do ćwiczeń z przedmiotu Genetyka z inżynierią genetyczną D - blok Inżynieria Genetyczna ćw. 3 Instytut Genetyki i Biotechnologii, Wydział Biologii, Uniwersytet Warszawski, rok akad. 2018/2019
Znaczenie genetyki. Opracował A. Podgórski
 Znaczenie genetyki Opracował A. Podgórski InŜynieria genetyczna InŜynieria genetyczna ingerencja w materiał genetyczny organizmów, w celu zmiany ich właściwości dziedzicznych. Istota inŝynierii genetycznej
Znaczenie genetyki Opracował A. Podgórski InŜynieria genetyczna InŜynieria genetyczna ingerencja w materiał genetyczny organizmów, w celu zmiany ich właściwości dziedzicznych. Istota inŝynierii genetycznej
Gdzie chirurg nie może - - tam wirusy pośle. czyli o przeciwnowotworowych terapiach wirusowych
 Gdzie chirurg nie może - - tam wirusy pośle czyli o przeciwnowotworowych terapiach wirusowych Wirusy zwiększające ryzyko rozwoju nowotworów HBV EBV HCV HTLV HPV HHV-8 Zmniejszenie liczby zgonów spowodowanych
Gdzie chirurg nie może - - tam wirusy pośle czyli o przeciwnowotworowych terapiach wirusowych Wirusy zwiększające ryzyko rozwoju nowotworów HBV EBV HCV HTLV HPV HHV-8 Zmniejszenie liczby zgonów spowodowanych
Wektory DNA - klonowanie molekularne
 Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
Wektory DNA - klonowanie molekularne
 Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2086467 (96) Data i numer zgłoszenia patentu europejskiego: 26.11.2007 07824706.1 (13) (51) T3 Int.Cl. A61F 2/16 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2086467 (96) Data i numer zgłoszenia patentu europejskiego: 26.11.2007 07824706.1 (13) (51) T3 Int.Cl. A61F 2/16 (2006.01)
Biologia Molekularna Podstawy
 Biologia Molekularna Podstawy Budowa DNA Budowa DNA Zasady: Purynowe: adenina i guanina Pirymidynowe: cytozyna i tymina 2 -deoksyryboza Grupy fosforanowe Budowa RNA Budowa RNA Zasady: purynowe: adenina
Biologia Molekularna Podstawy Budowa DNA Budowa DNA Zasady: Purynowe: adenina i guanina Pirymidynowe: cytozyna i tymina 2 -deoksyryboza Grupy fosforanowe Budowa RNA Budowa RNA Zasady: purynowe: adenina
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1801209 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.08.2005 05781421.2
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1801209 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.08.2005 05781421.2
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1732433 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.01.2005 05702820.1
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1732433 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.01.2005 05702820.1
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1671552 (96) Data i numer zgłoszenia patentu europejskiego: 02.12.2005 05026319.3 (13) T3 (51) Int. Cl. A23L1/305 A23J3/16
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1671552 (96) Data i numer zgłoszenia patentu europejskiego: 02.12.2005 05026319.3 (13) T3 (51) Int. Cl. A23L1/305 A23J3/16
PL B1. AKADEMIA MEDYCZNA, Warszawa, PL UNIWERSYTET MEDYCZNY IM. KAROLA MARCINKOWSKIEGO, Poznań, PL BUP 02/09
 RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 207902 (21) Numer zgłoszenia: 382945 (22) Data zgłoszenia: 18.07.2007 (13) B1 (51) Int.Cl. A61K 39/00 (2006.01)
RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 207902 (21) Numer zgłoszenia: 382945 (22) Data zgłoszenia: 18.07.2007 (13) B1 (51) Int.Cl. A61K 39/00 (2006.01)
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii
 Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
(86) Data i numer zgłoszenia międzynarodowego: , PCT/US99/21081 (87) Data i numer publikacji zgłoszenia międzynarodowego:
 Data i numer zgłoszenia międzynarodowego: , PCT/US99/21081 (87) Data i numer publikacji zgłoszenia międzynarodowego:") RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 201738 (21) Numer zgłoszenia: 355963 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 14.09.1999 (86) Data i numer zgłoszenia
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 201738 (21) Numer zgłoszenia: 355963 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 14.09.1999 (86) Data i numer zgłoszenia
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1968711 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 05.01.2007 07712641.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1968711 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 05.01.2007 07712641.5
Badanie funkcji genu
 Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Powodzenie reakcji PCR wymaga właściwego doboru szeregu parametrów:
 Powodzenie reakcji PCR wymaga właściwego doboru szeregu parametrów: dobór warunków samej reakcji PCR (temperatury, czas trwania cykli, ilości cykli itp.) dobór odpowiednich starterów do reakcji amplifikacji
Powodzenie reakcji PCR wymaga właściwego doboru szeregu parametrów: dobór warunków samej reakcji PCR (temperatury, czas trwania cykli, ilości cykli itp.) dobór odpowiednich starterów do reakcji amplifikacji
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 240040 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 09.07. 007077.0 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 240040 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 09.07. 007077.0 (97)
Wektory DNA - klonowanie molekularne
 Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1586320 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.02.2005 05472001.6 (51) Int. Cl. A61K31/435 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1586320 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.02.2005 05472001.6 (51) Int. Cl. A61K31/435 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1706498 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 26.09.2004 04775181.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1706498 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 26.09.2004 04775181.3
starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg
 STRESZCZENIE Przewlekła białaczka limfocytowa (PBL) jest najczęstszą białaczką ludzi starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg kliniczny, zróżnicowane rokowanie. Etiologia
STRESZCZENIE Przewlekła białaczka limfocytowa (PBL) jest najczęstszą białaczką ludzi starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg kliniczny, zróżnicowane rokowanie. Etiologia
Wektory DNA - klonowanie molekularne
 Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1755549 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.04.2005 05780098.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1755549 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.04.2005 05780098.9
Sylabus Biologia molekularna
 Sylabus Biologia molekularna 1. Metryczka Nazwa Wydziału Program kształcenia Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Analityka Medyczna, studia jednolite magisterskie, studia stacjonarne
Sylabus Biologia molekularna 1. Metryczka Nazwa Wydziału Program kształcenia Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Analityka Medyczna, studia jednolite magisterskie, studia stacjonarne
FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach
 FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach FOCUS Plus to dodatek dostępny dla standardowych pasz tuczowych BioMaru, dostosowany specjalnie do potrzeb ryb narażonych na trudne
FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach FOCUS Plus to dodatek dostępny dla standardowych pasz tuczowych BioMaru, dostosowany specjalnie do potrzeb ryb narażonych na trudne
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2152872. (96) Data i numer zgłoszenia patentu europejskiego: 21.05.2008 08748815.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2152872. (96) Data i numer zgłoszenia patentu europejskiego: 21.05.2008 08748815.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 212872 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.0.08 0874881.1 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 212872 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.0.08 0874881.1 (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2321564 (96) Data i numer zgłoszenia patentu europejskiego: 10.08.2008 08785479.0 (13) (51) T3 Int.Cl. F16L 21/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2321564 (96) Data i numer zgłoszenia patentu europejskiego: 10.08.2008 08785479.0 (13) (51) T3 Int.Cl. F16L 21/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2334863. (96) Data i numer zgłoszenia patentu europejskiego: 31.08.2009 09782381.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2334863. (96) Data i numer zgłoszenia patentu europejskiego: 31.08.2009 09782381.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2334863 (96) Data i numer zgłoszenia patentu europejskiego: 31.08.2009 09782381.9 (13) (51) T3 Int.Cl. D06F 39/08 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2334863 (96) Data i numer zgłoszenia patentu europejskiego: 31.08.2009 09782381.9 (13) (51) T3 Int.Cl. D06F 39/08 (2006.01)
Ćwiczenia 1 Wirtualne Klonowanie. Prowadzący: mgr Anna Pawlik i mgr Maciej Dylewski. Część teoretyczna:
 Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią ===============================================================================================
Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią ===============================================================================================
DYREKTYWA KOMISJI 2009/120/WE
 15.9.2009 Dziennik Urzędowy Unii Europejskiej L 242/3 DYREKTYWY DYREKTYWA KOMISJI 2009/120/WE z dnia 14 września 2009 r. zmieniająca dyrektywę 2001/83/WE Parlamentu Europejskiego i Rady w sprawie wspólnotowego
15.9.2009 Dziennik Urzędowy Unii Europejskiej L 242/3 DYREKTYWY DYREKTYWA KOMISJI 2009/120/WE z dnia 14 września 2009 r. zmieniająca dyrektywę 2001/83/WE Parlamentu Europejskiego i Rady w sprawie wspólnotowego
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445326 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.10.2011 11186353.6
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445326 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.10.2011 11186353.6
Biotechnologia i inżynieria genetyczna
 Wersja A Test podsumowujący rozdział II i inżynieria genetyczna..................................... Imię i nazwisko.............................. Data Klasa oniższy test składa się z 16 zadań. rzy każdym
Wersja A Test podsumowujący rozdział II i inżynieria genetyczna..................................... Imię i nazwisko.............................. Data Klasa oniższy test składa się z 16 zadań. rzy każdym
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 21737 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2010 10790844.4 (13) (1) T3 Int.Cl. A47L 1/42 (2006.01) A47L
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 21737 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2010 10790844.4 (13) (1) T3 Int.Cl. A47L 1/42 (2006.01) A47L
Rok akademicki: 2014/2015 Kod: EIB-2-206-BN-s Punkty ECTS: 3. Kierunek: Inżynieria Biomedyczna Specjalność: Bionanotechnologie
 Nazwa modułu: Genetyka molekularna Rok akademicki: 2014/2015 Kod: EIB-2-206-BN-s Punkty ECTS: 3 Wydział: Elektrotechniki, Automatyki, Informatyki i Inżynierii Biomedycznej Kierunek: Inżynieria Biomedyczna
Nazwa modułu: Genetyka molekularna Rok akademicki: 2014/2015 Kod: EIB-2-206-BN-s Punkty ECTS: 3 Wydział: Elektrotechniki, Automatyki, Informatyki i Inżynierii Biomedycznej Kierunek: Inżynieria Biomedyczna
Metody odczytu kolejności nukleotydów - sekwencjonowania DNA
 Metody odczytu kolejności nukleotydów - sekwencjonowania DNA 1. Metoda chemicznej degradacji DNA (metoda Maxama i Gilberta 1977) 2. Metoda terminacji syntezy łańcucha DNA - klasyczna metoda Sangera (Sanger
Metody odczytu kolejności nukleotydów - sekwencjonowania DNA 1. Metoda chemicznej degradacji DNA (metoda Maxama i Gilberta 1977) 2. Metoda terminacji syntezy łańcucha DNA - klasyczna metoda Sangera (Sanger
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2259949 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2009 09727379.1 (13) (51) T3 Int.Cl. B60L 11/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2259949 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2009 09727379.1 (13) (51) T3 Int.Cl. B60L 11/00 (2006.01)
Wektory DNA - klonowanie molekularne
 Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
Wektory DNA - klonowanie molekularne Fragment DNA (np. pojedynczy gen) można trwale wprowadzić do komórek gospodarza (tzn. zmusić go do powielania w tym gospodarzu) tylko wtedy, gdy zostanie on wbudowany
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 21 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.07.09 098000.7 (97) O
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 21 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.07.09 098000.7 (97) O
Ocena ekspresji genów proangiogennych w komórkach nowotworowych OVP-10 oraz transfektantach OVP-10/SHH i OVP-10/VEGF
 Agnieszka Gładysz Ocena ekspresji genów proangiogennych w komórkach nowotworowych OVP-10 oraz transfektantach OVP-10/SHH i OVP-10/VEGF Katedra i Zakład Biochemii i Chemii Klinicznej Akademia Medyczna Prof.
Agnieszka Gładysz Ocena ekspresji genów proangiogennych w komórkach nowotworowych OVP-10 oraz transfektantach OVP-10/SHH i OVP-10/VEGF Katedra i Zakład Biochemii i Chemii Klinicznej Akademia Medyczna Prof.
Przeciwciała poliklonalne przeciwko białkom Helicobacter pylori oraz sposób ich wytwarzania. (74) Pełnomocnik:
 Pełnomocnik:") RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 201982 (21) Numer zgłoszenia: 361069 (22) Data zgłoszenia: 03.07.2003 (13) B1 (51) Int.Cl. A61K 39/40 (2006.01)
RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 201982 (21) Numer zgłoszenia: 361069 (22) Data zgłoszenia: 03.07.2003 (13) B1 (51) Int.Cl. A61K 39/40 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 213136 (96) Data i numer zgłoszenia patentu europejskiego: 14.03.2008 08723469.6 (13) (1) T3 Int.Cl. F24D 19/ (2006.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 213136 (96) Data i numer zgłoszenia patentu europejskiego: 14.03.2008 08723469.6 (13) (1) T3 Int.Cl. F24D 19/ (2006.01) Urząd
(96) Data i numer zgłoszenia patentu europejskiego:
 Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1690978 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2005 05101042.9 (13) T3 (51) Int. Cl. D06F81/08 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1690978 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2005 05101042.9 (13) T3 (51) Int. Cl. D06F81/08 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2131847. (96) Data i numer zgłoszenia patentu europejskiego: 27.02.2008 08716068.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2131847. (96) Data i numer zgłoszenia patentu europejskiego: 27.02.2008 08716068.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2131847 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.02.2008 08716068.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2131847 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.02.2008 08716068.5
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886585 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.07.2006 06291197.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886585 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.07.2006 06291197.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1854925 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2005 05826699.0 (13) (51) T3 Int.Cl. E03D 1/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1854925 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2005 05826699.0 (13) (51) T3 Int.Cl. E03D 1/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1799953 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.08.2005 05770398.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1799953 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.08.2005 05770398.5
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2480370 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 08.09.2010 10773557.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2480370 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 08.09.2010 10773557.3
Patentowanie wynalazków z dziedziny biotechnologii
 chroń swoje pomysły 3 Kongres Świata Przemysłu Farmaceutycznego Poznań, 16-17 czerwca 2011 Patentowanie wynalazków z dziedziny biotechnologii dr Izabela Milczarek Czym jest biotechnologia? Biotechnologia,
chroń swoje pomysły 3 Kongres Świata Przemysłu Farmaceutycznego Poznań, 16-17 czerwca 2011 Patentowanie wynalazków z dziedziny biotechnologii dr Izabela Milczarek Czym jest biotechnologia? Biotechnologia,
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1505553. (96) Data i numer zgłoszenia patentu europejskiego: 05.08.2004 04018511.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1505553. (96) Data i numer zgłoszenia patentu europejskiego: 05.08.2004 04018511.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 3 (96) Data i numer zgłoszenia patentu europejskiego: 0.08.04 0401811.8 (13) (1) T3 Int.Cl. G08C 17/00 (06.01) Urząd Patentowy
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 3 (96) Data i numer zgłoszenia patentu europejskiego: 0.08.04 0401811.8 (13) (1) T3 Int.Cl. G08C 17/00 (06.01) Urząd Patentowy
LEKI CHEMICZNE A LEKI BIOLOGICZNE
 LEKI CHEMICZNE A LEKI BIOLOGICZNE PRODUKT LECZNICZY - DEFINICJA Art. 2 pkt.32 Ustawy - Prawo farmaceutyczne Substancja lub mieszanina substancji, przedstawiana jako posiadająca właściwości: zapobiegania
LEKI CHEMICZNE A LEKI BIOLOGICZNE PRODUKT LECZNICZY - DEFINICJA Art. 2 pkt.32 Ustawy - Prawo farmaceutyczne Substancja lub mieszanina substancji, przedstawiana jako posiadająca właściwości: zapobiegania
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2528702 (96) Data i numer zgłoszenia patentu europejskiego: 03.12.2010 10796315.9 (13) (51) T3 Int.Cl. B21D 53/36 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2528702 (96) Data i numer zgłoszenia patentu europejskiego: 03.12.2010 10796315.9 (13) (51) T3 Int.Cl. B21D 53/36 (2006.01)
Najważniejsze z nich to: enzymy restrykcyjne wektory DNA inne enzymy np. ligazy, fosfatazy, polimerazy, nukleazy
 Aby manipulować genami niezbędne są odpowiednie narzędzia molekularne, które pozwalają uzyskać tzw. zrekombinowane DNA (umożliwiają rekombinację materiału genetycznego in vitro czyli w próbówce) Najważniejsze
Aby manipulować genami niezbędne są odpowiednie narzędzia molekularne, które pozwalają uzyskać tzw. zrekombinowane DNA (umożliwiają rekombinację materiału genetycznego in vitro czyli w próbówce) Najważniejsze
GENOMIKA FUNKCJONALNA. Jak działają geny i genomy? Poziom I: Analizy transkryptomu
 GENOMIKA FUNKCJONALNA Jak działają geny i genomy? Poziom I: Analizy transkryptomu Adnotacja (ang. annotation) pierwszy etap po uzyskaniu kompletnej sekwencji nukleotydyowej genomu analiza bioinformatyczna
GENOMIKA FUNKCJONALNA Jak działają geny i genomy? Poziom I: Analizy transkryptomu Adnotacja (ang. annotation) pierwszy etap po uzyskaniu kompletnej sekwencji nukleotydyowej genomu analiza bioinformatyczna
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1730054 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 22.03.2005 05731932.9 (51) Int. Cl. B65G17/06 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1730054 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 22.03.2005 05731932.9 (51) Int. Cl. B65G17/06 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 223771 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.08 0886773.1 (13) (1) T3 Int.Cl. A47L 1/42 (06.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 223771 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.08 0886773.1 (13) (1) T3 Int.Cl. A47L 1/42 (06.01) Urząd
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1659297 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 12.10.2005 05354036.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1659297 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 12.10.2005 05354036.5