(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
|
|
|
- Gabriel Żurawski
- 7 lat temu
- Przeglądów:
Transkrypt
 O udzieleniu patentu europejskiego ogłoszono: 23.02.2011 Europejski Biuletyn Patentowy 2011/08 EP 1801209 B1 (13) (51) T3 Int.Cl. C12N 15/09 (2006.01) A61K 39/29 (2006.")
1 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: (97) O udzieleniu patentu europejskiego ogłoszono: Europejski Biuletyn Patentowy 2011/08 EP B1 (13) (51) T3 Int.Cl. C12N 15/09 ( ) A61K 39/29 ( ) A61K 48/00 ( ) A61P 1/16 ( ) A61P 31/14 ( ) C12N 5/10 ( ) C12N 7/00 ( ) C12Q 1/68 ( ) C12Q 1/70 ( ) (54) Tytuł wynalazku: Zmodyfikowany genomowy RNA ludzkiego wirusa zapalenia wątroby typu C posiadający zdolność replikacji autonomicznej (30) Pierwszeństwo: JP JP JP JP (43) Zgłoszenie ogłoszono: w Europejskim Biuletynie Patentowym nr 2007/26 (45) O złożeniu tłumaczenia patentu ogłoszono: Wiadomości Urzędu Patentowego 2011/07 (73) Uprawniony z patentu: Tokyo Metropolitan Organization for Medical Research, Tokyo, JP TORAY INDUSTRIES, INC., Tokyo, JP (72) Twórca(y) wynalazku: PL/EP T3 TAKAJI WAKITA, Tokyo, JP TAKANOBU KATO, Nagoya, JP TOMOKO DATE, Kawasaki, JP MICHIKO MIYAMOTO, Fuchu, JP RALF BARTENSCHLAGER, Schriesheim, DE JUN-ICHI TANABE, Fujisawa, JP SABURO SONE, Yokohama, JP (74) Pełnomocnik: rzecz. pat. Urszula Bartnik POLSERVICE KANCELARIA RZECZNIKÓW PATENTOWYCH SP. Z O.O. ul. Bluszczańska Warszawa Uwaga: W ciągu dziewięciu miesięcy od publikacji informacji o udzieleniu patentu europejskiego, każda osoba może wnieść do Europejskiego Urzędu Patentowego sprzeciw dotyczący udzielonego patentu europejskiego. Sprzeciw wnosi się w formie uzasadnionego na piśmie oświadczenia. Uważa się go za wniesiony dopiero z chwilą wniesienia opłaty za sprzeciw (Art. 99 (1) Konwencji o udzielaniu patentów europejskich).
2 3 Opis patentowy Dziedzina wynalazku [0001] Niniejszy wynalazek dotyczy sposobu autonomicznej replikacji wirusów ludzkiego zapalenia wątroby typu C (HCV) o różnych genotypach w układzie hodowli komórkowej, użytego do tego celu modyfikowanego genomowego RNA HCV, oraz komórki, w której przebiega replikacja powyższego genomowego RNA HCV. Tło wynalazku [0002] W wyniku badań prowadzonych w ostatnich latach stało się jasne, że wirus zapalenia wątroby typu C obejmuje ogromną ilość typów, zależnie od genotypu bądź serotypu. Zgodnie z metodą filoanalizy Simmondsa i wsp. z użyciem sekwencji nukleotydowych szczepów HCV, która to metoda jest ostatnio stosowana jako główna metoda klasyfikacji genotypów HCV, HCV klasyfikuje się na 6 następujących typów: genotyp 1a, genotyp 1b, genotyp 2a, genotyp 2b, genotyp 3a i genotyp 3b (dokument niepatentowy 1). Te typy klasyfikuje się dalej na szereg podtypów. Oznaczono również sekwencje nukleotydowe genomów o pełnej długości wielu różnych genotypów HCV (dokument patentowy 1 i dokumenty niepatentowe od 2 do 4). [0003] HCV powoduje przewlekłe zapalenie wątroby w wyniku długotrwale utrzymującej się infekcji. Główną przyczyną przewlekłego zapalenia wątroby, które jest schorzeniem występującym w skali globalnej, jest uporczywie utrzymująca się infekcja wirusem HCV. W rzeczywistości, u około 50% trwale zakażonych pacjentów rozwija się przewlekłe zapalenie wątroby a u około 20% pacjentów przeradza się ono w marskość wątroby w
3 4 okresie 10 do 20 lat. Co więcej, u niektórych pacjentów rozwijają się zagrażające życiu stany patologiczne, takie jak rak wątroby. [0004] obecnie, główne sposoby leczenia zapalenia wątroby typu C obejmują stosowanie -interferonu lub - interferonu oraz łączne stosowanie -interferonu z rybawiryną będącą pochodną nukleozydu purynowego. Pomimo tego, że takie leczenie stosuje się u pacjentów to jego skutki lecznicze obserwuje się jedynie u około 60% tych pacjentów. Jeśli po osiągnięciu efektów leczniczych zaprzestaje się leczenia, to u ponad połowy tych pacjentów następuje nawrót choroby. Wiadomo, że działanie lecznicze interferonu zależy od genotypu HCV. To znaczy, uważa się, że działanie interferonu jest słabe wobec genotypu 1b, natomiast jest ono silne wobec genotypu 2a (nie-patentowy dokument 5). Poza tym, swoistość wobec substratu proteazy HCV jest różna, zależnie od genotypu. Aktywność hamująca inhibitora opracowanego z użyciem proteazy NS3 z genotypu 1b jest 50 razy lub więcej słabsza od aktywności inhibitora opracowanego z użyciem proteaz NS3 z innych genotypów (nie-patentowy dokument 6). Tak więc, w celu opracowania skutecznego środka leczniczego przeciw HCV, należy opracować ten środek potwierdzając jednocześnie jego reaktywność wobec każdego z genotypów HCV. [0005] Ostatnio wytworzono subgenomowy replikon RNA HCV jako RNA pochodzący z HCV, który ma zdolność replikacji autonomicznej (dokumenty patentowe 2 i 3 i niepatentowe dokumenty 7 do 9). Dzięki temu stała się możliwa analiza mechanizmów replikacji HCV z użyciem hodowli komórkowych. Ten subgenomowy replikon RNA HCV wytworzono przez podstawienie strukturalnego białka znajdującego się w dół od wewnętrznego miejsca
4 5 wiązania rybosomy (sekwencji IRES HCV), w nie podlegającym translacji regionie 5 genomowego RNA HCV, genem oporności na neomycynę i EMCV-IRES, które wprowadzono przez ligację, od niego w dół. Ten replikon RNA wprowadzono do ludzkich komórek raka wątroby Huh7 i następnie prowadzono hodowlę tych komórek w obecności neomycyny. W wyniku tego doświadczenia wykazano, że ten replikon RNA podlega autonomicznej replikacji w komórkach Huh7. Wykazano ponadto, że kilka replikonów subgenomowego RNA HCV podlega autonomicznej replikacji w komórkach innych niż Huh7, takich jak ludzkie komórki raka szyjki macicy HeLa albo ludzkie komórki raka wątroby HepG2 (dokument patentowy 3). [0006] Jednakże, takie wewnątrzkomórkowe układy replikonu RNA HCV wytworzono dla ograniczonej liczby genotypów albo raczej, takie układy wytworzono stosując jedynie genomowe RNA z ograniczonej ilości szczepów HCV. A zatem, w odniesieniu do HCV posiadającego ogromną ilość genotypów, jest wyjątkowo trudno zanalizować różnice w działaniach leczniczych opracowywanych środków leczniczych przeciw HCV wynikające z różnic w genotypach tych środków. Taki replikon RNA jest układem eksperymentalnym, który jest użyteczny tylko do oceny replikacji wirusowego RNA podczas procesu wzrostu i replikacji wirusa HCV. Jest zatem niemożliwe, aby za pomocą takiego replikonu RNA dało się oceniać procesy takie jak powstawanie cząsteczek wirusa HCV w zakażonej komórce, uwalnianie ich z komórki bądź zakażanie nowej komórki. [0007] Obecnie, użycie metody oceny takich procesów, jak tworzenie się cząsteczek wirusa HCV, uwalnianie ich z komórki bądź zakażanie nowej komórki, jest ograniczone do układu eksperymentalnego, w którym
5 6 angażuje się zwierzęta, takie jak szympansy (dokument 10 niepatentowy). Jednakże taki układ eksperymentalny, w którym stosuje się bezpośrednio organizmy żywych zwierząt wymagają skomplikowanych operacji, a zatem w takim układzie eksperymentalnym jest wyjątkowo trudno prowadzić analizy. W związku z tym, w celu analizowania takich procesów, jak powstawanie cząsteczek wirusa HCV, ich uwalnianie z komórki i zakażanie nowej komórki albo w celu opracowania środka przeciw HCV z wykorzystaniem hamowania takich procesów jako mechanizmu działania, istnieje potrzeba skonstruowania jak najbardziej uproszczonego układu eksperymentalnego, w którym można powielać te procesy, a konkretnie, układu replikacji cząsteczki wirusa HCV z użyciem układu hodowli komórkowej. [0008] Jeśli istniałaby możliwość stałego dostarczania cząsteczek wirusa HCV z takiego układu hodowli komórkowej, wówczas można byłoby wirusa poddawać atenuacji albo można byłoby wytworzyć wirusa nieinfekcyjnego metodami opartymi na biologii molekularnej, i dzięki temu stosować takie wirusy jako szczepionki. Ponieważ jednak sekwencje białkowe HCV różnią się w zależności od genotypu, antygeniczność HCV również jest różna, zależnie genotypu. W rzeczywistości, występowanie różnych genotypów stanowi znaczącą przeszkodę w wytwarzaniu szczepionek przeciw HCV (dokument 11 niepatentowy). W związku z tym, w celu wydajnego wytwarzania szczepionek przeciw HCV cząstki wirusa HCV o różnych genotypach powinny być w trwały sposób wytwarzane w układzie hodowli komórkowej. [0009] Wiadomo, że HCV jest cząsteczką kolistą o wielkości między 55 a 65 nm, znajdującą się we krwi pacjenta zakażonego wirusem HCV. Jako metoda
6 7 oczyszczania HCV występującego w surowicy ludzkiej znana jest chromatografia powinowactwa z użyciem lektyny (dokument 12 nie patentowy) i chromatografia z użyciem heparyny (dokument 13 niepatentowy). Tymi metodami można jednak oczyścić mniej niż 1 ml wirusa o stężeniu około 1 M kopii/ml. Metody te nie mają zatem zastosowania przemysłowego. [0010] Dotychczas opracowano kilka sposobów oczyszczania cząstek wirusa innych niż HCV (np. dokumenty patentowe 4, 5 i 6). Z publikacji tych wynika jednak, że cząsteczki wirusów mają różne właściwości, a zatem nie dają one żadnej użytecznej informacji odnośnie optymalnego sposobu oczyszczania ludzkiego wirusa zapalenia wątroby typu C. W dokumencie patentowym 7 ujawniono, że ludzki wirus zapalenia wątroby typu A, który jest również wirusem zapalenia wątroby, można oczyścić przez usunięcie DNA metodą chromatografii anionowymiennej. Jednakże, jakkolwiek wirus zapalenia wątroby typu A jest również wirusem zapalenia wątroby, jest on wirusem posiadającym DNA jako gen. Wiadomo, że wirus zapalenia wątroby typu C posiada RNA jako gen, a więc nie występują odnośne podobieństwa pomiędzy wirusem zapalenia wątroby typu A a wirusem zapalenia wątroby typu C, nie ma tu zatem żadnej informacji dotyczącej odnośnych sposobów oczyszczania. W celu zastosowania cząsteczek ludzkiego wirusa zapalenia wątroby typu C jako szczepionek lub produktów podobnych, w przemyśle w przyszłości, należy wysoce oczyszczać te cząstki na dużą skalę. W związku z tym, oczekuje się opracowania sposobu oczyszczania. [Dokument patentowy 1] Publikacja patentowa Japonii (Kokai), JP A
7 8 [Dokument patentowy 2] Publikacja patentowa Japonii (Kokai), JP A [Dokument patentowy 3] Publikacja Zgłoszenia Międzynarodowego, WO 2004/ A1 [Dokument patentowy 4] Patent Japonii nr [Dokument patentowy 5] Publikacja patentowa Japonii (Kohyo), JP A [Dokument patentowy 6] Publikacja patentowa Japonii (Kohyo), JP A [Dokument patentowy 7] Publikacja patentowa Japonii (Kokoku), nr B (1994) [Dokument niepatentowy 1] Simmonds P. i wsp., Hepatology, 10 (1994), str [Dokument niepatentowy 2] Choo Q. L. i wsp., Science, 244 (1989), str [Dokument niepatentowy 3] Okamoto H. i wsp., J. Gen. Virol., 73 (1992), str [Dokument niepatentowy 4] Mori S. i wsp., Biochem. Biophis. Res. Commun. 183 (1992), str [Dokument niepatentowy 5] Yoshioka K. i wsp., Hepatology, 16 (1992), str [Dokument niepatentowy 6] Thibeault D. i wsp., J. Virol., 78 (2004), str [Dokument niepatentowy 7] Blight i wsp., Science, 290 (2000), str [Dokument niepatentowy 8]
8 9 Friebe i wsp., J. Virol., 75 (2001), str [Dokument niepatentowy 9] Kato T. i wsp., Gastroenterology, 125 (2003) str [Dokument niepatentowy 10] Kolykhalov i wsp., Science, 277 (1997), str [Dokument niepatentowy 11] Farci P. i wsp., Semin Liver Dis 20 (2000), str [Dokument niepatentowy 12] Virology, 196 (1993), str [Dokument niepatentowy 13] Journal of General Virology 86 (2005), str Ujawnienie wynalazku [0011] Przedmiotem wynalazku jest dostarczenie sposobu replikacji i amplifikacji wirusów zapalenia wątroby typu C o różnych genotypach w układzie hodowli komórkowej. [0012] W wyniku intensywnych badań ukierunkowanych na osiągnięcie powyższego celu, twórcy niniejszego wynalazku wytworzyli zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C przez połączenie genomowego RNA ze szczepu JFH1 HCV, który ma zdolność replikacji autonomicznej, z genomowym RNA szczepu HCV, który nie ma zdolności autonomicznej replikacji in vitro. Twórcy wynalazku wykryli, że tak wytworzony genomowy RNA ma zdolność autonomicznej replikacji w układzie hodowli komórkowej. Szczególnie, w odniesieniu do niniejszego wynalazku, okazało się, że wprowadzenie odcinka genomu rozciągającego się od sekwencji kodującej białko NS3 szczepu JFH1 do jej końca 3 umożliwia modyfikację
9 10 genomowego RNA wirusa HCV 1b, który nie ulega autonomicznej replikacji in vitro, otrzymując RNA, który może być autonomicznie replikowany w układzie hodowli komórkowej. [0013] Oznacza to, że przedmiotem wynalazku jest zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C, zawierający sekwencje nukleotydowe odcinków genomowego RNA z dwóch lub większej liczby typów wirusów zapalenia wątroby typu C, który to zmodyfikowany genom RNA zawiera nie podlegający translacji region 5, sekwencję kodującą białko rdzeniowe, sekwencję kodującą białko E1, sekwencję kodującą białko E2, sekwencję kodującą białko p7, sekwencję kodującą białko NS2, sekwencje kodujące białka NS3, NS4A, NS4B, NS5A i NS5B ze szczepu JFH1 oraz nie podlegający translacji region 3 i który to RNA może się autonomicznie replikować, przy czym jednym ze szczepów wirusa zapalenia wątroby typu C jest wirus o genotypie 1b. [0014] Szczególnie, w jednej z postaci, niniejszy wynalazek dostarcza zmodyfikowany genomowy RNA z wirusa zapalenia wątroby typu C, który wytwarza się przez podstawienie odcinka genomowego RNA wirusa zapalenia wątroby typu C o genotypie 1b, rozciągającgo się od sekwencji kodującej białko NS3 do sekwencji kodującej białko NS5B, będącej sekwencją genomową końca 3, przez częściową sekwencję RNA kodującą białka NS3, NS4, NS5A i NS5B ze szczepu JFH1, przedstawioną jako SEQ ID Nr: 1 (sekwencja otrzymana przez podstawienie T przez U w sekwencji odpowiadającej odcinkowi sekwencji DNA zdeponowanej w bibliotece GenBank pod numerem dostępu AB047639) i mającą zdolność autonomicznej replikacji.
10 11 [0015] W innym rozwiązaniu, przedmiotem wynalazku jest zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C, który wytwarza się przez podstawienie sekwencji kodującej białko NS5B w genomowym RNA wirusa zapalenia wątroby typu C przez sekwencję kodującą białko NS5B ze szczepu JFH1, przedstawioną jako SEQ ID Nr: 2 i który to zmodyfikowany genomowy RNA ma zdolność autonomicznej replikacji, przy czym jednym ze szczepów wirusa zapalenia wątroby typu C jest wirus o genotypie 1b. [0016] Szczepy dwóch lub większej ilości typów wirusów zapalenia wątroby typu C stosowanych w wynalazku obejmują wirusa zapalenia wątroby typu C o genotypie 1b i wirusa zapalenia wątroby typu C o genotypie 2a. Przykłady szczepu wirusa o genotypie 1b mogą obejmować szczep HCV-con1, szczep HCV-TH, szczep HCV-J, szczep HCV-JT i szczep HCV-BK. Szczepem wirusa o genotypie 2a jest szczep HCV-JFH1. [0017] Ten zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według wynalazku może poza tym zawierać co najmniej jeden selekcyjny gen markerowy i/lub co najmniej jeden gen reporterowy i co najmniej jedną sekwencję IRES. W tym przypadku ten zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C zawiera opisany powyżej, nie podlegający translacji region 5, co najmniej jeden selekcyjny gen markerowy, i/lub co najmniej jeden gen reporterowy, co najmniej jedną sekwencję IRES, sekwencję kodującą białko rdzeniowe, sekwencję kodującą białko E1, sekwencję kodującą białko E2, sekwencję kodującą białko p7, sekwencję kodującą białko NS2, sekwencję kodującą białko NS3, sekwencję kodującą białko NS4A, sekwencję kodującą białko NS4B, sekwencję kodującą białko NS5A, sekwencję kodującą białko NS5B i
11 12 nie podlegający translacji region 3, w tej kolejności, w kierunku od końca 5 do końca 3. [0019] Jako przykład opisanego powyżej zmodyfikowanego genomowego RNA wirusa zapalenia wątroby typu C, w niniejszym opisie patentowym opisany jest zmodyfikowany genomowy RNA zapalenia wątroby typu C, który zawiera: (a) RNA posiadający sekwencję nukleotydową przedstawioną jako SEQ ID Nr. 11; albo (b) RNA posiadający sekwencję nukleotydową zawierającą delecję, substytucję lub addycję jednego lub większej ilości, korzystnie 100, bardziej korzystnie 50, a jeszcze bardziej korzystnie 10 nukleotydów w odniesieniu do sekwencji nukleotydowej przedstawionej jako SEQ ID Nr: 11 i który może być autonomicznie replikowany i może generować cząsteczki wirusa zapalenia wątroby typu C. [0020] Ponadto, przedmiotem wynalazku jest również komórka, do której jest wprowadzony ten zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według niniejszego wynalazku i w której przebiega replikacja wyżej opisanego genomowego RNA wirusa zapalenia wątroby typu C i która może generować cząsteczki wirusa. W wynalazku, jako komórkę gospodarza stosuje się komórkę proliferującą. Szczególnie korzystne przykłady takiej komórki gospodarza obejmują komórki eukariotyczne, w tym komórki pochodzące z wątroby ludzkiej, takie jak komórki Huh7, komórki HepG2, komórki IMY-N9, komórki HeLa albo komórki 293, komórki ludzkiej szyjki macicy i komórki pochodzące z nerki ludzkiego płodu. [0021] Ponadto, przedmiotem wynalazku jest również sposób wytwarzania cząsteczek wirusa zapalenia wątroby typu C, znamienny tym, że obejmuje prowadzenie hodowli
12 13 wyżej opisanej komórki i odzyskiwanie cząsteczek wirusa z tej hodowli; oraz cząsteczki wirusa zapalenia wątroby typu C wytworzone powyższym sposobem. [0022] Ponadto, niniejszy wynalazek dostarcza także sposobu wytwarzania komórki zakażonej wirusem zapalenia wątroby typu C, znamiennego tym, że sposób obejmuje hodowlę wyżej opisanej komórki i zakażanie innych komórek cząsteczkami wirusa znajdującymi się w tej hodowli, a także komórki zakażonej wirusem zapalenia wątroby typu C, wytworzonej tym sposobem. W niniejszym wynalazku te cząsteczki HCV oczyszcza się metodą chromatografii kolumnowej i/lub przez wirowanie w gradiencie gęstości, tak, aby otrzymać cząsteczki HCV o czystości pozwalającej na ich przemysłowe zastosowanie do wytwarzania produktów farmaceutycznych. W niniejszym sposobie, jako chromatografię, stosuje się jeden lub większą ilość typów chromatografii, wybranych spośród chromatografii jonowymiennej, chromatografii żelowej i chromatografii powinowactwa. Wirowanie w gradiencie gęstości prowadzi się z użyciem jednego lub większej ilości substancji rozpuszczonych wybranych spośród chlorku cezu, sacharozy i polimerów cukrowych, tak, aby oczyścić HCV. [0023] Dalszym przedmiotem wynalazku jest również sposób przesiewowego badania substancji na działanie przeciw wirusowi zapalenia wątroby typu C z użyciem komórki według wynalazku albo komórki zakażonej wirusem zapalenia wątroby typu C. Sposób ten jest znamienny tym, że obejmuje hodowlę komórki według wynalazku albo komórki zakażonej wirusem zapalenia wątroby typu C w obecności badanej substancji i wykrywanie RNA wirusa zapalenia wątroby typu C albo cząsteczek wirusa w hodowli, i w ten sposób ocenę skutków działania przeciw
13 14 wirusowi zapalenia wątroby typu C powyższej badanej substancji. [0024] Przedmiotem wynalazku jest też sposób wytwarzania szczepionki przeciw zapaleniu wątroby typu C z użyciem cząsteczek wirusa zapalenia wątroby typu C według wynalazku albo ich części jako antygenu. [0025] Ponadto, przedmiotem wynalazku jest: sposób replikacji i/lub ekspresji obcego genu w komórce, charakteryzujący się tym, że sposób ten obejmuje insercję RNA kodującego ten obcy gen do zmodyfikowanego genomowego RNA wirusa zapalenia wątroby typu C według wynalazku i wprowadzenie genomowego RNA do komórki będącej przedmiotem zainteresowania, tak, aby przebiegała w niej replikacja lub ekspresja tego obcego genu, oraz wektor wirusowy ukierunkowany na komórkę wątroby, zawierający zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według wynalazku. [0026] Zgodnie z niniejszym wynalazkiem cząsteczki wirusa HCV mające zdolność zakażania mogą być wytwarzane w układzie hodowli komórkowej. Co więcej, nawet w przypadku szczepu HCV, który nie może być autonomicznie replikowany i który jest wyizolowany od pacjentów, podstawia się jego region odpowiadający regionowi od regionu NS3 do końca 3 genomowym RNA wirusa JFH1 albo region NS5B podstawia się regionem NS5B z JFH1, tak aby powyższy szczep HCV mógł ulegać autonomicznej replikacji in vitro. Tak więc, cząsteczki wirusa HCV o różnych genotypach można wytwarzać w układzie hodowli komórkowej i te cząsteczki wirusa wydajnie się stosuje do badań procesu zakażenia wirusem HCV albo do wytwarzania układu do badania przesiewowego dla różnych substancji, które mają wpływ na proces
14 15 zakażenia wirusem HCV, a także do wytwarzania szczepionki przeciw HCV. Krótki opis rysunków Na fig. 1 jest schematycznym przedstawieniem procedury konstruowania matrycowego DNA stosowanego do wytworzenia genomowego RNA HCV według wynalazku. Na figurze jest przedstawiona struktura plazmidowego klonu pjfh1 wytworzonego przez insercję genomu HCV o pełnej długości w dół od promotora T7. Symbole podane na figurze mają następujące znaczenie: T7: promotor RNA T7; 5 -UTR: nie podlegający translacji region 5 ; C: białko rdzeniowe; E1, E2: białka otoczki; NS2, NS3, NS4A, NS4B, NS5A, NS5B: białka niestrukturalne; 3 -UTR: nie podlegający translacji region 3 ; AgeI, PmeI, XbaI: miejsca cięcia przez enzymy restrykcyjne AgeI, PmeI i Xba;: GDD: pozycja motywu aminokwasowego GDD odpowiadająca centrum aktywnemu białka NS5B; Fig. 2 przedstawia fotografię obrazującą wyniki analizy techniką hybrydyzacji northern, wskazujące na replikację rjfh1 w komórkach Huh7, do których został wprowadzony rjfh1, to znaczy genomowy RNA z HCV; Fig. 3 przedstawia wyniki dotyczące detekcji rdzeniowego białka HCV, białka NS3, białka NS5A i białka E2 w pożywce; Fig. 4 przedstawia wyniki odnośnie czasu trwania zmian w uwalnianiu do pożywki białka rdzeniowego z komórek, do których został wprowadzony genomowy RNA HCV; Na fig. 5 zawiera wykresy obrazujące ilość białka rdzeniowego HCV i ilość genomowego RNA HCV w każdej frakcji uzyskanej przez frakcjonowanie w gradiencie gęstości sacharozy supernatantu z hodowli komórek Huh7, do których wprowadzono rjfh1. Czarne kółka oznaczają
15 16 białko rdzeniowe HCV a białe kółka oznaczają genomowy RNA HCV. Fig. 5A przedstawia wyniki dla nietraktowanych komórek Huh7 z wprowadzonym rjfh1. Fig. 5B przedstawia wyniki dla traktowanych RNazą komórek Huh7 z wprowadzonym rjfh1. Fig. 5C przedstawia wyniki dla traktowanych przez NP40 komórek Huh7 z wprowadzonym rjfh1. Fig. 5D przedstawia wyniki dla traktowanych NP40 + RNazą komórek Huh7 z wprowadzonym rjfh1; Fig. 6 przedstawia infekcyjność wydzielonych cząsteczek wirusa w roztworze hodowli komórek Huh7 z wprowadzonym rjfh1. Na fig. 6A są podane fotografie przedstawiające wyniki barwienia immunologicznego przeciwciałem przeciw rdzeniowi (po lewej stronie) i przeciwciałem przeciw NS5A (po prawej stronie). Fig. 6B jest graficznym przedstawieniem ilości komórek dodatnich, które ulegają barwieniu przeciwciałem przeciw rdzeniowi. Na fig. 6C są podane wykresy przedstawiające zmianę w czasie poziomu RNA HCV w tych komórkach (po lewej stronie) i w supernatancie (po prawej stronie); Fig. 7 przedstawia infekcyjność wydzielonych cząsteczek wirusa w roztworze hodowli komórek Huh7 z wprowadzonym rjch1/ns5b(jfh1). Fig. 7A jest graficznym przedstawieniem amplifikacji cząsteczek RNA wirusa HCV wydzielonych w roztworze hodowli komórek Huh7 z wprowadzonym rjch1/ns5b(jfh1), w naturalnych (nienaruszonych) komórkach Huh7. Fig. 7B jest graficznym przedstawieniem ilości komórek dodatnich, które ulegają barwieniu przeciwciałem przeciw rdzeniowi; Na fig. 8 jest przedstawiona struktura chimerycznego replikonu TH/JFH1;
16 17 Na fig. 9 są przedstawione wyniki dotyczące tworzenia kolonii przez transfekcję chimerycznym replikonem RNA, rth/jfh1; Na fig. 10 są przedstawione wyniki dotyczące tworzenia kolonii przez zakażenie supernatantu hodowli chimerycznym replikonem TH/JFH1; Fig. 11 przedstawia profile elucji w chromatografii żelowej. Na osi pionowej odłożona jest absorbancja przy długości fali 490 nm. S-300, S-400 i S-500 oznaczają odpowiednio żel Sephacryl S-300, S-400 i S500. Oś pozioma przedstawia ilości eluatu wypływającego z kolumny; Na fig. 12 są przedstawione profile elucji w chromatografii jonowymiennej. Na osi pionowej są odłożone ilości białka rdzeniowego cząsteczek HCV; Na fig. 13 są przedstawione profile elucji w chromatografii powinowactwa na lektynie. Na osi pionowej są odłożone ilości białka rdzeniowego cząsteczek HCV; Na fig. 14 są przedstawione profile elucji z dwóch typów chromatografii powinowactwa, z użyciem heparyny i siarczanu celulofiny. Oś pionowa oznacza absorbancję przy długości fali 490 nm. Fig. 15 przedstawia profil elucji w chromatografii powinowactwa z barwnikiem niebieskim. Na osi pionowej są odłożone ilości białka rdzeniowego cząsteczek HCV; i Na fig. 16 są przedstawione profile oczyszczania z łącznym użyciem chromatografii kolumnowej i wirowania w gradiencie gęstości sacharozy. Na osi pionowej są odłożone ilości białka rdzeniowego cząsteczek HCV. W przypadku wirowania w gradiencie gęstości sacharozy, gęstość roztworu każdej frakcji oraz ilość białka rdzeniowego HCV są przedstawione na osi pionowej.
17 18 [0028] Niniejszy opis patentowy obejmuje treść ujawnioną w tym opisie i/lub rysunki ze zgłoszeń patentowych Japonii o numerach: , , i , które są dokumentami pierwszeństwa dla niniejszego zgłoszenia patentowego. Najlepszy sposób realizacji wynalazku [0029] Wynalazek jest opisany szczegółowo poniżej. 1. Zmodyfikowany chimeryczny genomowy RNA wirusa zapalenia wątroby typu C [0030] Genom wirusa zapalenia wątroby typu C (HCV) jest jednoniciowym RNA, będącym nicią (+) złożoną z około 9600 nukleotydów. Ten genomowy RNA zawiera nie podlegający translacji region 5 (oznaczany również symbolami 5 -NTR albo 5 -UTR), region podlegający translacji złożony z regionu strukturalnego i regionu niestrukturalnego i nie podlegający translacji region 3 (oznaczany również symbolami 3 -NTR albo 3 -UTR). Region strukturalny koduje białka strukturalne wirusa HCV a region niestrukturalny koduje wiele białek niestrukturalnych. [0031] Te strukturalne białka HCV (rdzeniowe, E1 i E2) oraz niestrukturalne białka HCV (NS2, NS3, NS4A, NS4B, NS5A i NS5B) są produktami translacji występującymi jako jedna ciągła poliproteina z regionu translacyjnego. Następnie, ta poliproteina podlega ograniczonemu trawieniu proteazą i w ten sposób proteiny mogą być uwalniane i generowane. Wśród tych białek strukturalnych i niestrukturalnych (białek wirusa HCV) rdzeń jest białkiem rdzeniowym a E1 i E2 są białkami otoczki. Białko niestrukturalne jest białkiem związanym z replikacją wirusa jako taką. Wiadomo, że NS2 ma aktywność metaloproteazy zaś NS3 ma aktywność
18 19 proteazy serynowej (jedna trzecia od strony końca N) oraz aktywność helikazy (dwie trzecie od strony końca C). Donoszono również, że NS4A jest kofaktorem dla aktywności NS3 jako proteazy i że NS5B wykazuje RNAzależną aktywność polimerazy RNA. [0032] Obecnie wiadomo, że genotypy HCV klasyfikuje się na co najmniej typy od 1 do 6. HCV klasyfikuje się, zależnie od jego sekwencji, na różne genotypy (HCV1a, HCV1b, HCV2a, HCV2b i tak dalej), zgodnie z międzynarodową klasyfikacją Simmonds a i wsp. (Simmonds P. i wsp., Hepatology (1994) 10, str ). W niniejszym wynalazku, genomowy RNA HCV, który nie ma zdolności autonomicznej replikacji nie ogranicza się do powyższych, znanych typów wirusów lecz obejmuje wszystkie typy genomowego RNA HCV, które nie mają zdolności autonomicznej replikacji, czyli zdolności do uwalniania zakaźnych cząsteczek poza komórkę. W niniejszym wynalazku, wyrażenie: RNA ma zdolność autonomicznej replikacji albo jest autonomicznie replikowany jest stosowane w tym znaczeniu, że jeśli genomowy RNA HCV zostanie wprowadzony do komórki, wówczas ten genomowy RNA HCV replikuje się autonomicznie, czyli ma zdolność uwalniania zakaźnych cząsteczek poza tę komórkę. [0033] W niniejszym opisie wynalazku RNA, w tym wyżej wspomniany genomowy RNA HCV, który może ulegać autonomicznej replikacji w układzie hodowli komórkowej jest nazwany replikonem RNA albo RNA-replikonem. W niniejszym opisie wynalazku, replikon RNA według wynalazku będący replikonem RNA o pełnej długości nazwany jest replikonem RNA HCV o pełnej długości. Replikon RNA HCV o pełnej długości według wynalazku ma zdolność generowania cząsteczek wirusa. Ponadto,
19 20 zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według wynalazku jest replikonem RNA HCV o pełnej długości. [0034] Zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według wynalazku obejmuje zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C, który posiada sekwencje nukleotydowe genomowych odcinków RNA z dwóch lub z większej ilości typów wirusów zapalenia wątroby typu C, obejmujące nie podlegający translacji region 5, sekwencję kodującą białko rdzeniowe, sekwencję kodującą białko E1, sekwencję kodującą białko E2, sekwencję kodującą białko p7, sekwencję kodującą białko NS2, sekwencje kodujące dla każdego z białek NS3, NS4A, NS4B, NS5A i NS5B ze szczepu JFH1 oraz nie podlegający translacji region 3, i który to RNA ma zdolność autonomicznej replikacji. Szczególnie w jednym rozwiązaniu, wynalazek obejmuje zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C, który wytwarza się przez podstawienie odcinka genomowego RNA wirusa zapalenia wątroby typu C rozciągającego się od sekwencji kodującej białko NS3 do sekwencji kodującej białko NS5B, czyli sekwencji genomowej przy końcu 3, przez częściową sekwencję RNA kodującą białka NS3, NS4, NS5A i NS5B ze szczepu JFH1, przedstawioną jako SEQ ID nr 1 (sekwencja RNA otrzymana przez podstawienie T przez U w sekwencji odpowiadającej części sekwencji DNA zdeponowanej w bibliotece GenBank pod numerem dostępu AB047639) i który to RNA ma zdolność autonomicznej replikacji. [0035] W innym rozwiązaniu, wynalazek dostarcza zmodyfikowanego, genomowego RNA wirusa zapalenia wątroby typu C, który wytwarza się przez podstawienie sekwencji kodującej białko NS5B z genomowego RNA wirusa
20 21 zapalenia wątroby typu C sekwencją kodującą białko NS5B ze szczepu JFH1, przedstawioną jako SEQ ID Nr: 2, i który ma zdolność autonomicznej replikacji. [0036] Korzystnie, wynalazek obejmuje zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C, otrzymany z użyciem wirusów zapalenia wątroby typu C o genotypach 1b i 2a, który to RNA ma sekwencję nukleotydową obejmującą nie podlegający translacji region 5, sekwencję kodującą białko rdzeniowe, sekwencję kodującą białko E1, sekwencję kodującą białko E2, sekwencję kodującą białko p7, sekwencję kodującą białko NS2, sekwencje kodujące dla każdego z białek NS3, NS4A, NS4B, NS5A i NS5B ze szczepu JFH1 oraz nie podlegający translacji region-3, i który to RNA ma zdolność autonomicznej replikacji. [0037] Wyżej opisany zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C może ponadto zawierać przynajmniej jeden markerowy gen selekcyjny i/lub przynajmniej jeden gen reporterowy oraz co najmniej jedną sekwencję IRES. [0038] W niniejszym wynalazku, stosując szczep HCV, który ma zdolność autonomicznej replikacji w układzie hodowli komórkowej w kombinacji ze szczepem HCV, który nie może być autonomicznie replikowany w tym układzie hodowli komórkowej, jako dwa lub większą ilość typów wirusów zapalenia wątroby typu C, można modyfikować szczep HCV, który nie ma zdolności autonomicznej replikacji tak, aby stał się on zdolny do autonomicznej replikacji. Innymi słowy, szczep wirusa, który podlega autonomicznej replikacji wydajnie można tak modyfikować, aby replikował się autonomicznie bardzo wydajnie.
21 22 [0039] Konkretne przykłady znanego szczepu wirusa HCV o genotypie 1a mogą obejmować szczep HCV-1, szczep HCV-H i szczep HCV-J1. Konkretne przykłady znanego szczepu wirusa HCV o genotypie 1b mogą obejmować szczep HCV-con1, szczep HCV-TH, szczep HCV-J, szczep HCV-JT i szczep HCV-BK. Konkretne przykłady znanego szczepu wirusa HCV o genotypie 2a mogą obejmować szczep HCV-J6, szczep JFH-1 i szczep JCH1. Przykładem znanego szczepu HCV o typie 2b może być szczep HC-J8. Przykładem znanego szczepu HCV o typie 3a może być szczep E-b1. Struktura tych wirusów jest w zasadzie oparta na 5 - UTR, rdzeniu, E1, E2, p7, NS2, NS3, NS4a, NS4b, NS5a, NS5b i 3 -UTR (jak opisano powyżej). Oznaczono sekwencję nukleotydową każdego regionu każdego z powyższych szczepów HCV. Dla przykładu, sekwencje nukleotydowe regionów odpowiadających rdzeniowi, E1, E2, p7 i NS2 oznaczono na sekwencji o pełnej długości szczepu TH. Ponadto, na sekwencji szczepu HCV-JT oznaczono regiony odpowiadające rdzeniowi, E1, E2, p7 i NS2. Przykładem replikonu RNA według wynalazku może być chimeryczny replikon RNA HCV, który otrzymuje się stosując szczep JFH1 z HCV typu 2a i szczepy inne niż szczep JFH1 z HCV typu 2a, takie jak szczep HCV-1, szczep HCV-H, szczep HCV-J1, szczep HCV-con1, szczep HCV-TH [Wakita i wsp., J. Biol. Chem. (1994) 269, str ; Moradpour i wsp., Biochem. Biophys. Res. Commun., (1998) 246, str ], szczep HCV-J, szczep HCV-JT, szczep HCV-BK, szczep HCV-J6, szczep JCH1, szczep HC-J8 albo szczep E-b1. [0040] Poza tym korzystnym przykładem zmodyfikowanego genomowego RNA HCV według wynalazku może być genomowy RNA HCV otrzymany przez podstawienie regionu odpowiadającego regionowi rozciągającemu się od
22 23 regionu NS3 do końca 3 w genomowym RNA HCV szczepu JFH1 wirusa zapalenia wątroby typu C, wirusowym genomowym RNA z JFH1 albo przez podstawienie sekwencji kodującej białko NS5B sekwencją kodującą białko NS5B z genomowego RNA innego HCV albo przez insercję do niego powyższej sekwencji. Na przykład, w przypadku genomowego RNA wirusa HCV JCH1 (ref), o którym wiadomo, że nie ma zdolności replikacji in vitro, podstawia się region odpowiadający jego regionowi od regionu NS3 do miejsca końca 3 genomowym wirusowym RNA szczepu JFH1, i w ten sposób, genomowy RNA HCV można modyfikować uzyskując genomowy RNA HCV, który ma zdolność autonomicznej replikacji. [0041] Ponadto, w przypadku klonu genomowego RNA HCV Con-1 (ref) o genotypie HCV 1b (EMBL, nr dostępu AJ238799), podstawia się część jego sekwencji RNA kodującą białka NS3, NS4, NS5A i NS5B sekwencją RNA ze szczepu JFH1 kodującą białka NS3, NS4, NS5A i NS5B albo podstawia się tylko część sekwencji RNA kodującą białko NS5B z klonu Con-1 (ref) o genotypie HCV b1, sekwencją RNA, która koduje białko NS5B ze szczepu JFH1, i w ten sposób, genomowy RNA HCV można modyfikować, uzyskując genomowy RNA HCV, który ma zdolność autonomicznej replikacji. [0042] Replikon o pełnej długości, uzyskany z użyciem genu z klonu Con-1 może się autonomicznie replikować, ale nie tworzy cząsteczek HCV [Pietschmann i wsp., Journal of Virology, (2002), 76, str ]. Jednakże, jak opisano w przykładzie w opisie niniejszego wynalazku, takie cząsteczki HCV można tworzyć przez podstawienie części sekwencji RNA kodującej białka NS3, NS4, NS5A i NS5B sekwencją RNA, która koduje białka NS3, NS4, NS5A i NS5B ze szczepu
23 24 JFH1. Innymi słowy, sposobem według wynalazku, genomowy RNA wirusa zapalenia wątroby typu C, który ma zdolność autonomicznej replikacji ale nie może tworzyć cząsteczek HCV można przekształcić w zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C, który ma zdolność tworzenia cząsteczek. [0043] Poza tym, nawet w przypadku HCV, który nie może wytworzyć replikonu mającego zdolność autonomicznej replikacji, takiego jak szczep TH albo szczep JCH, tworzy się cząsteczki HCV przez wytworzenie jego chimerycznego genu ze szczepem JFH-1, jak to opisano w przykładzie niniejszego wynalazku. Tak więc, wynalazek umożliwia konwersję genomowego RNA HCV, który nie ma zdolności autonomicznej replikacji do modyfikowanego genomowego RNA wirusa zapalenia wątroby typu C, który może tworzyć cząsteczki HCV. [0044] Co więcej, przez wprowadzenie mutacji do NS5B w części sekwencji RNA szczepu JFH1, zatrzymuje się wzrost genomowego RNA HCV i również zatrzymuje się generowanie cząsteczek HCV. Tak więc, najwyraźniej, NS5B pełni ważną rolę w umożliwianiu autonomicznej replikacji i tworzenia cząsteczek przez genomowy RNA HCV. [0045] Obecnie, HCV klasyfikuje się na różne genotypy (HCV1a, HCV1b, HCV2a, HCV2b i tak dalej) zależnie od jego sekwencji, zgodnie z międzynarodową klasyfikacją Simmondsa i wsp. [Simmonds P. i wsp., Hepatology, (1994) 10, str ]. W niniejszym wynalazku, genomowy RNA HCV, który nie ma zdolności autonomicznej replikacji nie ogranicza się do powyższych, znanych typów wirusów lecz obejmuje wszystkie typy genomowego RNA HCV, które nie mają zdolności autonomicznej replikacji.
24 25 [0046] W niniejszym opisie, sekwencją kodującą białko NS5B jest sekwencja kodująca białko NS5B pochodząca ze szczepu JFH1 (SEQ ID nr: 3) i ma ona sekwencję nukleotydową przedstawioną na SEQ ID nr: 2. Sekwencja kodująca białko NS5B według wynalazku obejmuje jednak również sekwencje nukleotydowe, które mogą hybrydyzować z sekwencją nukleotydową przedstawioną jako SEQ ID nr: 2 w warunkach ścisłych, tak długo, jak długo te sekwencje nukleotydowe kodują aminokwasy posiadające funkcję białka NS5B (na przykład białko NS5B zawierające substytucję konserwatywną). [0047] Termin warunki ścisłe stosuje się w znaczeniu na przykład warunków, w których stężenie sodu wynosi od 300 do 2000 mm a temperatura wynosi od 40 o C do 75 o C, korzystniej, stężenie sodu wynosi od 600 do 900 mm a temperatura wynosi 65 o C. Specjaliści z tej dziedziny z łatwością otrzymają powyższy homolog NS5B, wykorzystując odniesienia do Molecular Cloning (Sambrook J. i wsp., Molecular Cloning: A Laboratory Manual, wyd. II, Cold Spring Harbor Laboratory Press, 10 Skyline Drive Plainview, NY (1989). [0048] Genomowy RNA HCV według wynalazku posiada część sekwencji RNA, która koduje białka NS3, NS4, NS5A i NS5B w genomowym RNA HCV JFH1 albo sekwencję kodującą białko NS5B. [0049] W jednym z rozwiązań, genomowym RNA HCV według wynalazku jest RNA, który posiada sekwencję nukleotydową obejmującą nie podlegający translacji 5 - region, sekwencję kodującą białko rdzeniowe, sekwencję kodującą białko E1, sekwencję kodującą białko E2, sekwencję kodującą białko NS2, sekwencję kodującą białko NS3, sekwencję kodującą białko NS4A, sekwencję kodującą białko NS4B, sekwencję kodującą białko NS5A,
25 26 sekwencję kodującą białko NS5B oraz nie podlegający translacji region 3 z genomowego RNA szczepu wirusa zapalenia wątroby typu C. Poza tym, w powyższym RNA, wspomniana część sekwencji kodująca białka NS3, NS4, NS5A i NS5B jest częścią sekwencji RNA kodującą białka NS3, NS4, NS5A i NS5B, która pochodzi z zewnątrz wprowadzonego genomowego RNA HCV szczepu JFH1. Korzystnie, jest to RNA, w którym jego sekwencja kodująca białko NS5B jest sekwencją kodującą białko NS5B pochodzącą z zewnątrz wprowadzonego genomowego RNA HCV szczepu JFH1. [0050] W opisie niniejszego wynalazku nie podlegający translacji 5 region (5 -NTR albo 5 -UTR), sekwencja kodująca białko rdzeniowe (region rdzeniowy albo region C), sekwencja kodująca białko E1 (region E1), sekwencja kodująca białko E2 (region E2), sekwencja kodująca białko NS2 (region NS2), sekwencja kodująca białko NS3 (region NS3), sekwencja kodująca białko NS4A (region NS4A), sekwencja kodująca białko NS4B (region NS4B), sekwencja kodująca białko NS5A (region NS5A), sekwencja kodująca białko NS5B (region NS5B), nie podlegający translacji region 3 (3 -NTR albo 3 - UTR) i inne specyficzne regiony lub miejsca, zostały już poznane w różnych genotypach. Powyższe regiony lub miejsca w nieznanym szczepie HCV można łatwo oznaczyć przez przyrównanie genomowej sekwencji RNA o pełnej długości znanego HCV z sekwencją z powyższego szczepu HCV. [0051] Termin selekcyjny gen markerowy jest w niniejszym opisie wynalazku stosowany w znaczeniu genu, który nadaje komórkom cechę selektywności, umożliwiając selekcję tylko tych komórek, w których przebiega ekspresja tego genu. Powszechnym przykładem takiego
26 27 selekcyjnego genu markerowego może być gen oporności na antybiotyk. Przykłady takiego selekcyjnego genu markerowego, który może być korzystnie użyty w wynalazku obejmują gen oporności na neomycynę, gen kinazy tymidynowej, gen oporności na kanamycynę, gen oporności na pirytiaminę, gen adenylylotransferazy, gen oporności na zeocynę i gen oporności na puromycynę. Spośród nich, korzystne są gen oporności na neomycynę i gen kinazy tymidynowej, przy czym gen oporności na neomycynę jest bardziej korzystny. Selekcyjne geny markerowe stosowane w niniejszym wynalazku nie ograniczają się jednak do powyższych genów. [0052] Stosowany w wynalazku termin gen reporterowy oznacza gen markerowy, który koduje produkt tego genu, będący wskaźnikiem ekspresji tego genu. Powszechnym przykładem takiego genu reporterowego może być strukturalny gen dla enzymu katalizującego reakcję luminescencyjną albo reakcję barwną. Przykłady genu reporterowego, który może być korzystnie użyty w niniejszym wynalazku obejmują gen acetylotransferazy chloramfenikolu pochodzący z transposonu Tn9, gen - glukuronidazy albo gen -galaktozydazy pochodzący z Escherichia coli, gen lucyferazy, gen zielonego białka fluorescencyjnego, gen ekworyny pochodzący z meduzy i sekrecyjna forma genu ludzkiej alkalicznej fosfatazy z łożyska (SEAP). Geny reporterowe stosowane w niniejszym wynalazku nie ograniczają się jednak do powyższych genów. [0053] Dowolny z powyższych selekcyjnych genów markerowych i genów reporterowych może być zawarty w replikonie RNA albo mogą być w nim zawarte obydwa takie geny. W odniesieniu do takiego selekcyjnego genu markerowego albo genu reporterowego, jeden gen może być
27 28 zawarty w zmodyfikowanym genomowym RNA wirusa zapalenia wątroby typu C albo mogą być w nim zawarte dwa albo większa ilość tych genów. [0054] Genomowy RNA HCV według wynalazku może ponadto zawierać RNA kodujący dowolny obcy gen przeznaczony do ekspresji w komórkach, do których wprowadzono genomowy RNA HCV o pełnej długości. Taki RNA kodujący obcy gen można wprowadzić przez ligację w dół od nie podlegającego translacji regionu 5 albo w górę od nie podlegającego translacji regionu 3. Taki RNA może być również wprowadzony przez insercję do dowolnej przestrzeni pomiędzy sekwencją kodującą białko rdzeniowe, sekwencją kodującą białko E1, sekwencją kodującą białko E2, sekwencją kodującą białko NS2, sekwencją kodującą białko NS3, sekwencją kodującą białko NS4A, sekwencją kodującą białko NS4B, sekwencją kodującą białko NS5A i sekwencją kodującą białko NS5B. [0055] Jeśli w komórkach, do których wprowadzono ten RNA będzie przebiegać translacja genomowego RNA HCV zawierającego RNA kodujący obcy gen, wówczas będzie możliwa ekspresja produktu kodowanego przez ten obcy gen. Zgodnie z tym, taki genomowy RNA HCV zawierający RNA kodujący obcy gen może być korzystnie wykorzystany również do celów generowania produktu tego obcego genu w komórkach. [0056] W genomowym RNA HCV według wynalazku, powyższe sekwencje kodujące białka wirusowe, obcy gen i inne poddaje się ligacji jedna do drugiej w taki sposób, aby mogły one podlegać translacji z tego genomowego RNA HCV w prawidłowej fazie odczytu. Białka kodowane przez ten genomowy RNA HCV korzystnie poddaje się ligacji jedno z drugim, wykorzystując miejsca rozszczepienia przez proteazę lub podobne, aby w wyniku translacji, białka
28 29 te powstawały w formie ciągłego polipeptydu i aby była możliwa ich ekspresja i tak, aby polipeptyd mógł być następnie rozszczepiony proteazą na każde z białek i następnie uwolniony. [0057] Tak wytworzony genomowy RNA HCV zawierający część sekwencji RNA kodującą białka NS3, NS4, NS5A i NS5B ze szczepu JFH1 wprowadza się następnie do odpowiednich komórek gospodarza, tak, aby uzyskać komórki rekombinacyjne, w których może przebiegać autonomiczna replikacja genomowego RNA HCV, a korzystnie, może trwale przebiegać autonomiczna replikacja genomowego RNA HCV (to znaczy, że może się replikować genomowy RNA HCV). W dalszej części niniejszego opisu wynalazku, takie rekombinacyjne komórki, w których może przebiegać replikacja genomowego RNA HCV zawierającego część sekwencji kodującą białka NS3, NS4, NS5A i NS5B ze szczepu JFH1 są nazwane komórkami replikującymi genomowy RNA HCV. [0058] Typ komórek gospodarza stosowanych do takich komórek replikujących genomowy RNA HCV nie jest w jakiś szczególny sposób limitowany, tak długo, jak długo można prowadzić subkultury. Korzystne są komórki eukariotyczne. Bardziej korzystne są komórki ludzkie a jeszcze bardziej korzystne są ludzkie komórki pochodzące z wątroby, ludzkie komórki szyjki macicy i ludzkie komórki nerki płodu. Ponadto, korzystne są komórki proliferacyjne, w tym szczepy komórek rakowych lub szczepy komórek macierzystych. Szczególnie korzystne są między innymi komórki Huh7, komórki HepG2, komórki IMY-N9, komórki HeLa, komórki 293 i podobne. Jako takie komórki można użyć komórki dostępne w obrocie albo takie komórki można również uzyskać z instytucji deponujących komórki. Poza tym, można
29 30 również użyć komórki wyhodowane z dowolnych komórek (np. z komórek rakowych albo komórek macierzystych). [0059] Genomowy RNA HCV można wprowadzić do komórek gospodarza dowolną znaną techniką. Przykładami takich metod wprowadzania mogą być elektroporacja, bombardowanie cząsteczkami, metoda lipofekcji, metoda z fosforanem wapnia, metoda mikroiniekcji i metoda na sefarozie DEAE. Spośród tych metod szczególnie korzystna jest metoda z użyciem elektroporacji. [0060] Genomowy RNA HCV można wprowadzać pojedynczo albo można go mieszać z innym kwasem nukleinowym i następnie wprowadzać. Aby zmienić ilość genomowego RNA HCV wprowadzonego przy stałej ilości wprowadzonego RNA, można zmieszać pewną ilość genomowego RNA HCV z całkowitym RNA komórkowym wyekstrahowanym z komórek, do których ma być wprowadzony genomowy RNA HCV, tak, aby przygotować pewną całkowitą ilość RNA i następnie, tę całkowitą ilość RNA można wprowadzić do komórek. Ilość genomowego RNA HCV wprowadzonego do komórek można oznaczyć zależnie od użytej metody wprowadzenia. Ilość takiego wprowadzonego genomowego RNA HCV wynosi korzystnie od 1 pikograma do 100 mikrogramów a jeszcze korzystniej, od 10 pikogramów do 10 mikrogramów. [0061] Replikację genomowego RNA HCV w komórkach replikujących genomowy RNA HCV można potwierdzić dowolną znaną metodą detekcji RNA. Przykładowo, całkowity RNA wyekstrahowany z komórek poddaje się hybrydyzacji typu northern z użyciem fragmentu DNA swoistego dla wprowadzonego genomowego RNA HCV jako sondy albo prowadzi się reakcję RT-PCR, stosując startery swoiste dla wprowadzonego genomowego RNA HCV. [0062] Poza tym, jeśli wśród białek ekstrahowanych z komórek replikujących genomowy RNA HCV wykrywa się
30 31 białko HCV, wówczas można przyjąć, że w tych komórkach przebiega replikacja genomowego RNA HCV. Takie białko HCV można wykryć dowolną metodą znaną dla detekcji białek. Dla przykładu, takie białko HCV można wykryć w reakcji przeciwciała reagującego z białkiem HCV, które musi być wynikiem ekspresji tego wprowadzonego genomowego RNA HCV, z białkiem ekstrahowanym z tych komórek. Konkretniej, próbkę białka wyekstrahowanego z tych komórek nanosi się na filtr nitrocelulozowy i następnie umożliwia się jego reakcję z przeciwciałem przeciw białku HCV (np. z przeciwciałem swoistym wobec NS3 albo z antysurowicą zebraną od pacjenta z zapaleniem wątroby typu C), po czym wykrywa się to przeciwciało, np. przeciw białku HCV. [0063] Fakt, że genomowy RNA HCV ma zdolność autonomicznej replikacji można potwierdzić na przykład przez transfekcję komórek Huh7, stosując RNA jako cel, hodowlę tych komórek Huh7 i poddanie RNA wyekstrahowanego z tych komórek w otrzymanej hodowli hybrydyzacji typu northern, stosując sondę swoiście wykrywającą wprowadzony RNA, jednak ta metoda nie jest jedyną metodą detekcji. Szczegółowe postępowanie dla potwierdzenia, że RNA ma zdolność autonomicznej replikacji są podane w opisach odnoszących się do potwierdzenia ekspresji białka HCV albo detekcji genomowego RNA HCV w przykładzie niniejszego opisu patentowego. 2. Wytwarzanie cząsteczek HCV [0064] Komórki replikujące genomowy RNA HCV wytworzone w sposób opisany powyżej mają zdolność generowania cząsteczek wirusa HCV in vitro. Innymi słowy, prowadzi się hodowlę komórek replikujących genomowy RNA HCV
31 32 według wynalazku w odpowiedniej pożywce i następnie zbiera się wytworzone cząsteczki wirusa z hodowli (korzystnie z roztworu hodowli), uzyskując z łatwością cząsteczki HCV. [0065] Zdolność generowania cząsteczek wirusa przez komórki replikujące genomowy RNA HCV można potwierdzić dowolną znaną metodą detekcji wirusa. Przykładowo, roztwór hodowli zawierający komórki, które prawdopodobnie generują cząsteczki wirusa, frakcjonuje się w gradiencie gęstości sacharozy i dokonuje się pomiaru gęstości, stężenia białka rdzeniowego HCV i ilości genomowego RNA HCV w każdej frakcji. W wyniku, jeśli pik dla białka rdzeniowego HCV odpowiada pikowi genomowego RNA HCV i jeśli gęstość frakcji, w której wykrywa się ten pik jest mniejsza niż gęstość tej samej frakcji, którą uzyskuje się z frakcjonowania po traktowaniu supernatantu hodowli 0,25% NP40 (eterem polioksyetyleno(9)oktylo-fenylowym)(wynosi na przykład od 1,15 mg do 1,22 mg), to można potwierdzić, że komórki mają zdolność generowania cząsteczek wirusa. [0066] Cząsteczki wirusa HCV uwalniane do roztworu hodowli można również wykryć stosując przeciwciało reagujące z białkiem rdzeniowym, z białkiem E1 albo z białkiem E2. Poza tym, jest również możliwa pośrednia detekcja obecności cząsteczek wirusa HCV przez amplifikację genomowego RNA HCV zawartego w cząsteczkach wirusa w roztworze hodowli i następna detekcja amplifikowanego produktu metodą RT-PCR z użyciem swoistych starterów. 3. Zakażenie innych komórek cząsteczkami HCV według wynalazku
32 33 [0067] Cząsteczki wirusa HCV generowane sposobem według wynalazku mają zdolność zakażania komórek (korzystnie komórek wrażliwych na HCV). Wynalazek dostarcza również sposobu wytwarzania komórki zakażonej wirusem zapalenia wątroby typu C, który to sposób polega na hodowaniu komórek replikujących genomowy RNA HCV i następnym zakażaniu innych komórek (korzystnie komórek wrażliwych na HCV) cząsteczkami wirusa zawartymi w uzyskanej hodowli (korzystnie w roztworze hodowli). Termin komórki wrażliwe na HCV w niniejszym opisie oznacza komórki mające cechę zakażania się wirusem HCV. Takimi komórkami wrażliwymi na HCV są korzystnie komórki wątroby albo limfocyty ale przykłady nie ograniczają się do nich. Szczególne przykłady takich komórek wątroby obejmują pierwotne hepatocyty, komórki Huh7, komórki HepG2, komórki IMY-N9, komórki HeLa i komórki 293. Szczególne przykłady limfocytów obejmują komórki Molt4, komórki HPB-Ma i komórki Daudi ego. Ich przykłady nie ograniczają się jednak do tych komórek. [0068] Po zakażeniu komórek (na przykład komórek wrażliwych na HCV) cząsteczkami HCV generowanymi w komórkach replikujących genomowy RNA HCV według wynalazku, genomowy RNA HCV jest replikowany w zakażonych komórkach i następnie powstają cząsteczki wirusa. Następnie, przez stworzenie możliwości aby komórki uległy zakażeniu cząsteczkami wirusa generowanymi w komórkach replikujących genomowy RNA HCV według wynalazku, w tych komórkach może zachodzić replikacja genomowego RNA HCV i w dalszym ciągu mogą powstawać cząsteczki wirusa. [0069] Jeśli zwierzęta, które mogą być zakażone wirusem HCV, takie jak szympansy, ulegną zakażeniu cząsteczkami wirusa HCV generowanymi w komórkach replikujących
33 34 genomowy RNA HCV według wynalazku, wówczas cząsteczki te mogą powodować u tych zwierząt zapalenie wątroby wywołane przez HCV. 4. Oczyszczanie cząsteczek HCV [0070] Roztwór zawierający wirusy HCV przeznaczony do oczyszczania cząsteczek HCV może pochodzić z jednego lub większej ilości źródeł wybranych spośród krwi pobranej od pacjenta zakażonego HCV, zakażonej HCV hodowli komórek, pożywki hodowli komórkowej zawierającej komórki generujące cząsteczki HCV w wyniku rekombinacji genetycznej i roztworu uzyskanego z homogenatu tych komórek. [0071] Roztwór zawierający wirusy HCV poddaje się wirowaniu i/lub filtracji przez filtr w celu usunięcia komórek i pozostałości komórkowych. Roztwór otrzymany po usunięciu tych pozostałości zatęża się od 10 do 100-krotnie na membranie do ultrafiltracji wycinającej masy cząsteczkowe od do [0072] Zawierający HCV roztwór, z którego usunięto pozostałości, oczyszcza się metodą chromatografii albo przez wirowanie w gradiencie gęstości, jak opisano poniżej, albo przez zastosowanie kombinacji metod chromatografii i wirowania w gradiencie gęstości, w dowolnej kolejności. Reprezentatywne metody chromatografii i wirowania w gradiencie gęstości są opisane poniżej ale wynalazek nie ogranicza się do tych metod. [0073] Do oczyszczania cząsteczek HCV można użyć chromatografię z filtracją żelową, korzystnie stosując nośnik chromatograficzny posiadający jako matrycą żelową sieciowany polimer składający się z allilodekstranu i N,N -metylenobisakryloamidu a
34 35 korzystniej, stosując Sephacryl S-300, S-400 albo S [0074] Do oczyszczania cząsteczek HCV można też zastosować chromatografię jono-wymienną, korzystnie na Q-Sepharose jako żywicy aniono-wymiennej i korzystnie, na SP Sepharose jako żywicy kationo-wymiennej. [0075] Do oczyszczania cząsteczek HCV można wykorzystać chromatografię powinowactwa, korzystnie stosując jako nośnik żywicę, jako ligand, z którym wiąże się substrat wybrany spośród heparyny, siarczanu cellulofiny, lektyny i różnych barwników. Taką chromatografię powinowactwa można zastosować do oczyszczania cząsteczek HCV, korzystniej stosując HiTrap Heparyn HP, HiTrap Blue HP, HiTrap benzamidynę FF, siarczan celulofiny albo nośniki, z którymi wiążą się LCA, ConA, RCA-120 i WGA. Taką chromatografię powinowactwa można użyć do oczyszczania cząsteczek HCV, najkorzystniej stosując siarczan celulofiny jako nośnik. Nieoczekiwanie, cząsteczki HCV oczyszczono 30-krotnie w odniesieniu do stosunku całkowitej masy białka w roztworze do ilości kopii RNA HCV przed i po oczyszczaniu. [0076] W oczyszczaniu przez wirowanie w gradiencie gęstości, jako substancję rozpuszczoną dającą gradient gęstości korzystnie stosuje się chlorek cezu, sacharozę, Nycodenz albo polimer cukrowy, taki jak Ficoll albo Percoll. Bardziej korzystnie, stosuje się sacharozę. Poza tym, jako rozpuszczalnik stosuje się wodę albo roztwór buforu, taki jak bufor fosforanowy, bufor Tris, bufor octanowy albo bufor glicynowy. [0077] Oczyszczanie prowadzi się w temperaturze korzystnie od 0 o C do 40 o C, bardziej korzystnie od 0 o C do 25 o C, a najkorzystniej od 0 o C do 10 o C.
35 36 [0078] W sposobie oczyszczania z wirowaniem w gradiencie gęstości, siła odśrodkowa użyta w procesie oczyszczania wynosi korzystnie od 1 x 10 4 do 1 x 10 9 g, jeszcze korzystniej od 5 x 10 4 do 1 x 10 7 g a najkorzystniej od 5 x 10 4 do 5 x 10 5 g. [0079] W odniesieniu do łącznego użycia metod oczyszczania, wirowanie w gradiencie gęstości i chromatografię kolumnową można łączyć w dowolnej kolejności. Korzystnie, po oczyszczeniu cząsteczek HCV wieloma typami chromatografii kolumnowej, otrzymany preparat poddaje się wirowaniu w gradiencie gęstości. Bardziej korzystnie, prowadzi się chromatografię kolumnową aniono-wymienną i następnie chromatografię powinowactwa, uzyskując frakcję zawierającą cząsteczki HCV i tę frakcję oczyszcza się następnie przez wirowanie w gradiencie gęstości. Najkorzystniej, frakcję zawierającą cząsteczki HCV otrzymaną po chromatografii kolumnowej z użyciem Q-Sepharose oczyszcza się dalej na kolumnie wypełnionej siarczanem cellulofiny i następnie, otrzymaną frakcję zawierającą cząsteczki HCV oczyszcza się przez wirowanie w gradiencie gęstości. Poza tym, pomiędzy procesem chromatografii kolumnowej a procesem wirowania w gradiencie gęstości można przeprowadzić dializę albo ultrafiltrację, tak aby przeprowadzić wymianę substancji rozpuszczonej w roztworze zawierającym cząsteczki HCV i/lub zatężenie cząsteczek HCV. 5. Inne rozwiązania według wynalazku [0080 Genomowy RNA HCV replikuje się z wysoką wydajnością w komórkach replikujących genomowy RNA HCV według wynalazku. Zgodnie z tym, stosując komórki replikujące genomowy RNA HCV według wynalazku,
36 37 genomowy RNA HCV może być wytwarzany z wysoką wydajnością. [0081] W niniejszym wynalazku prowadzi się hodowlę komórek replikujących genomowy RNA HCV i następnie prowadzi się ekstrakcję RNA z tej hodowli (z komórek hodowli i/lub z pożywki hodowli) Wyekstrahowany RNA poddaje się elektroforezie aby wyizolować i oczyścić wydzielony genomowy RNA HCV, wytwarzając w ten sposób genomowy RNA HCV. Tak wytworzony RNA zawiera sekwencję genomu HCV. Dzięki zapewnieniu sposobu wytwarzania RNA zawierającego sekwencję genomu HCV stało się możliwe bardziej szczegółowe zanalizowanie genomu HCV. [0082] Poza tym, komórki replikujące genomowy RNA HCV według wynalazku można korzystnie użyć do wytworzenia białka HCV. Takie białko HCV można wytworzyć w znany sposób. Przykładowo, do komórek wprowadza się genomowy RNA HCV, wytwarzając komórki rekombinantowe. Następnie prowadzi się hodowlę tych rekombinantowych komórek i z uzyskanej hodowli (z komórek hodowli i/lub z pożywki hodowli) odzyskuje sie białko w znanymi sposobami. [0083] Cząsteczki wirusa HCV mogą wykazywać ukierunkowanie na komórki wątroby. Można zatem skonstruować ukierunkowany na komórki wątroby wektor wirusowy wykorzystując genomowy RNA HCV według wynalazku. Taki wektor wirusowy korzystnie stosuje się w terapii genowej. W niniejszym wynalazku, RNA kodujący obcy gen wprowadza się do genomowego RNA HCV i taki RNA wprowadza się następnie do komórek, tak, aby wprowadzić powyższy obcy gen do tych komórek. Następnie, ten obcy gen można replikować i prowadzić jego ekspresję w komórkach. [0084] Ponadto, RNA wytwarza się przez wymianę sekwencji kodującej białko E1 i/lub sekwencji kodującej
37 38 białko E2 w genomowym RNA HCV na białko otoczki wirusa pochodzącego z innych żywych organizmów. Wytworzony RNA wprowadza się następnie do komórek, tak aby wytworzyć cząsteczki wirusa. Staje się więc możliwe zakażenie komórek różnych żywych organizmów przez ten RNA. Również i w tym przypadku, obcy gen wprowadza się do genomowego RNA HCV i uzyskany RNA stosuje się jako ukierunkowany na komórkę wektor wirusowy w celu umożliwienia ekspresji tego obcego genu w różnych typach komórek, zależnie od kierunkowości białka otoczki rekombinowanego wirusa. [0085] Wynalazek odnosi się również do sposobu wytwarzania wektora wirusowego zawierającego obcy gen, który to sposób polega na wprowadzeniu przez insercję RNA kodującego ten obcy gen do genomowego RNA HCV, wprowadzeniu genomowego RNA do komórek i na hodowli tych komórek, umożliwiając generowanie cząsteczek wirusa w tych komórkach. [0086] Wynalazek dostarcza również sposobu wytwarzania szczepionki przeciw zapaleniu wątroby typu C albo szczepionki przeciw wirusowi użytemu do rekombinacji genetycznej białka otoczki, wykorzystując cząsteczki HCV według wynalazku albo ich część jako antygen albo wykorzystując cząsteczki wytworzone przez rekombinację genetyczną białka otoczki wirusa albo ich część dla zmiany ukierunkowania na dany typ komórek, jako antygen. Poza tym, można również wytworzyć neutralizujące przeciwciało przeciw zakażeniu HCV, wykorzystując cząsteczki HCV według wynalazku lub ich część jako antygen albo wykorzystując cząsteczki wytworzone przez genetyczną rekombinację białka otoczki wirusa albo ich część, w celu zmiany jego ukierunkowania na komórki, jako antygen.
38 39 [0087] Komórki replikujące genomowy RNA HCV według wynalazku albo komórki zakażone wirusem HCV, które zostały zakażone cząsteczkami wirusa generowanymi w komórkach replikujących genomowy RNA HCV, można na przykład użyć do replikacji HCV albo rekonstrukcji cząsteczek wirusa albo jako układ testowy do prób przesiewowych na substancję, która pobudza bądź hamuje uwalnianie cząsteczek wirusa (substancję przeciw wirusowi zapalenia wątroby typu C). Szczególnie, przykładowo prowadzi się hodowlę takich komórek w obecności testowanej substancji i wykrywa się genomowy RNA HCV albo cząsteczki wirusa zawarte w otrzymanej hodowli. Następnie, określa się, czy powyższa testowana substancja pobudza czy hamuje replikację replikonu RNA albo genomowego RNA HCV, powstawanie tych cząsteczek wirusa albo ich uwalnianie, prowadząc badania przesiewowe na substancję, która pobudza bądź hamuje wzrost wirusów zapalenia wątroby typu C. W tym przypadku, genomowy RNA HCV zawarty w hodowli można wykryć przez pomiar ilości genomowego RNA HCV w RNA ekstrahowanym z powyższych komórek, przez jego udział albo jego obecność lub brak. Cząsteczki wirusa zawarte w hodowli (głównie w roztworze hodowli) można wykryć przez pomiar ilości białka HCV zawartego w roztworze hodowli, przez pomiar jego udziału albo stwierdzenie jego obecności lub braku. [0088] Cząsteczki HCV generowane w komórkach replikujących genomowy RNA HCV według wynalazku i komórki wrażliwe na HCV można stosować jako układy testowe do prób przesiewowych na substancję, która pobudza lub hamuje wiązanie HCV z komórkami. Szczególnie, prowadzi się hodowlę komórek wrażliwych na HCV razem z cząsteczkami HCV generowanymi w komórkach
39 40 replikujących genomowy RNA HCV według wynalazku w obecności testowanej substancji. Następnie wykrywa się genomowy RNA HCV albo cząsteczki wirusa w otrzymanej hodowli. Określa się, czy powyższa testowana substancja pobudza czy hamuje replikację genomowego RNA HCV albo powstawanie cząsteczek wirusa, prowadząc próby przesiewowe na substancję, która pobudza bądź hamuje wzrost wirusów zapalenia wątroby typu C. [0089] Taki genomowy RNA HCV albo cząsteczki wirusa można wykrywać wyżej opisanymi sposobami albo sposobami podanymi w poniższych przykładach. Wyżej opisany układ testowy można użyć do wytworzenia lub oceny środka zapobiegawczego, środka leczniczego albo środka diagnostycznego na zakażenie wirusem zapalenia wątroby typu C. [0090] Szczególne przykłady użycia powyższego układu testowego według wynalazku są podane poniżej. (1) Próby przesiewowe na substancję, która hamuje wzrost HCV i zakażenie tym wirusem [0091] Przykłady substancji, która hamuje wzrost HCV i zakażenie tym wirusem mogą obejmować: związek organiczny, który bezpośrednio bądź pośrednio wpływa na wzrost HCV i zakażenie tym wirusem; oligonukleotyd antysensowny, który hybrydyzuje z docelową sekwencją w genomie HCV albo z nicią komplementarną do tej sekwencji, wpływając bezpośrednio lub pośrednio na wzrost HCV albo na translację białka HCV. (2) Ocena różnych substancji wykazujących aktywność przeciwwirusową w hodowli komórkowej [0092] Przykładem powyższych różnych substancji może być substancja uzyskana metodą racjonalnego
40 41 projektowania leku albo wysoko-wydajnego skriningu (na przykład wyizolowany i oczyszczony enzym). (3) Identyfikacja nowego celu, który należy zaatakować i użyć do leczenia pacjentów zakażonych wirusem HCV [0093] Komórki replikujące genomowy RNA HCV według wynalazku można na przykład użyć do identyfikacji białka komórki gospodarza pełniącego ważną rolę w replikacji wirusa HCV. (4) Ocena zdolności wirusa HCV do nabywania oporności na środki lub inne czynniki i identyfikacja mutacji związanej z tą opornością (5) Produkcja białka wirusowego stosowanego jako antygen, które to białko może być wykorzystane do opracowania, wytwarzania i oceny środka diagnostycznego albo środka leczniczego przeciw zakażeniu wirusem zapalenia wątroby typu C. (6) Wytwarzanie białka wirusowego i atenuowanego HCV, stosowanych jako antygeny, które mogą być wykorzystane do opracowania, produkcji i oceny szczepionki przeciw zakażeniu wirusem zapalenia wątroby typu C. PRZYKŁADY [0094] Wynalazek jest bardziej szczegółowo przedstawiony w poniższych przykładach i na rysunkach. Przykłady te jednak nie mają na celu ograniczenia zakresu technicznego niniejszego wynalazku. [Przykład 1] Wytwarzanie genomowego RNA HCV 1. Konstruowanie wektora ekspresyjnego
41 42 [0095] DNA odpowiadający całkowitemu regionowi genomu wirusa zapalenia wątroby typu C, szczepu JFH1 (genotyp 2a) wyizolowany od pacjentów cierpiących na piorunujące zapalenie wątroby, otrzymano z klonu JFH1 zawierającego genomowy cdna o pełnej długości z powyższego szczepu wirusa [Kato T. i wsp., J. Med. Virol. 64 (2001), str ]. Następnie, otrzymany DNA wbudowano przez insercję w dół od sekwencji promotora RNA T7, która została wprowadzona do plazmidu puc19. Konkretnie, fragment z reakcji RT-PCR otrzymany przez amplifikację wirusowego RNA ze szczepu JFH1 klonowano do wektora pgem-t EASY (Promega), aby uzyskać różne plazmidowe DNA, takie jak pgem1-258, pgem44-486, pgem , pgem , pgem , pgem , pgem , pgem , pgem , pgem , pgem , pgem , pgem i pgem [Kato T. i wsp., Gastroenterology, 125 (2003), str ]. Genomowy cdna wirusa zawarty w każdym plazmidzie poddano ligacji, jeden z drugim, w reakcji PCR i stosując enzymy restrykcyjne i następnie sklonowano ten genomowy cdna o pełnej długości. W górę od niego dokonano insercji sekwencji promotora RNA T7, uzyskując klon JFH1 (pjfh1) (fig. 1). Zaznacza się, że ta sekwencja cdna o pełnej długości z pjfh1 jest zarejestrowana w Międzynarodowej Bibliotece DNA (DDBJ/EMBL/GenBank) pod numerem dostępum AB [0096] Następnie, w odniesieniu do regionu NS5B w pjfh1 (sekwencja nukleotydowa: SEQ ID Nr. 2; sekwencja aminokwasowa: SEQ ID Nr. 3), motyw aminokwasowy GDD odpowiadający centrum aktywnemu polimerazy RNA kodowanej przez powyższy region, mutuje się do GND, wytwarzając zmutowany klon plazmidowy pjfh1/gnd. Ponieważ sekwencja aminokwasowa centrum aktywnego
42 43 białka NS5B kodowanego przez ten zmutowany klon plazmidowy pjfh1/gnd jest zmutowana to w tym klonie nie może przebiegać ekspresja aktywnego białka NS5B potrzebnego do replikacji RNA HCV. [0097] Następnie wycina się region E1 i region E2 z pjfh1, otrzymując pjfh1/ E1-E2. Poza tym, wbudowuje się cdna HCV o pełnej długości ze szczepu J6CF (GenBank, numer dostępu AF177036), który różni się od szczepu JFH1 i od szczepu JCH1 (Kato T. i wsp., J. Med. Virol. 64 (2001), str ), w dół od sekwencji promotora RNA T7, która została wbudowana przez insercję do plazmidu puc19, wytwarzając odpowiednio pj6cf i pjch1. Poza tym, podstawia się region kodujący NS5B w pjch1 przez NS5B z JFH1, wytwarzając pjch1/ns5b(jfh1). 2. Wytwarzanie genomowego RNA HCV [0098] W celu wytworzenia matrycowego DNA użytego następnie do syntezy RNA, każdy z plazmidów pjfh1, pjfh1/gnd, pjfh1/ E1-E2, pj6cf, pjch1 i pjch1/ns5b(jfh1) trawiono enzymem restrykcyjnym XbaI. Następnie, ilości 10 do 20 µg każdego z fragmentów powstałych po trawieniu enzymem XbaI inkubowano z 20 jednostkami enzymu, nukleazy z fasoli mung (Mung Bean) (w całkowitej objętości roztworu reakcyjnego 50 µl) w temperaturze 30 o C przez 30 minut. Nukleaza Mung Bean jest enzymem katalizującym reakcję selektywnego trawienia jednoniciowej części dwuniciowego DNA. Na ogół, jeśli RNA syntetyzuje się bezpośrednio, stosując jako matrycę powyższy fragment z trawienia enzymem XbaI, wówczas jest syntetyzowany replikon RNA, do którego, na jego końcu 3 dodaje się w nadmiarze 4 nukleotydy CUGA, stanowiące część sekwencji rozpoznawanej przez XbaI. Tak więc, w niniejszym
43 44 przykładzie, taki fragment z rozszczepienia enzymem XbaI traktowano nukleazą z fasoli mung, eliminując w ten sposób 4 nukleotydy CUGA z tego fragmentu trawienia XbaI. Ten traktowany nukleazą Mung Bean roztwór zawierający fragment z rozszczepienia XbaI, poddano traktowaniu polegającemu na eliminacji białka, postępując w znany sposób, tak aby można było oczyścić fragment z trawienia XbaI, z którego usunięto 4 nukleotydy CUGA. Ten oczyszczony fragment użyto jako matrycę DNA. [0099] Na powyższej matrycy DNA syntetyzowano następnie in vitro RNA. Ten RNA syntetyzowano w reakcji 20 µl roztworu reakcyjnego zawierającego 0,5 do 1,0 µg matrycowego DNA w temperaturze 37 o C w czasie 3 do 16 godzin, stosując MEGAscript wytwarzany przez firmę Ambion. [0100] Po zakończeniu syntezy RNA do roztworu reakcyjnego dodano DNA-zę (2 jednostki) i mieszaninę utrzymywano w temperaturze 37 o C przez 15 minut. Następnie dalej ekstrahowano RNA kwaśnym fenolem i usunięto matrycowy DNA. W ten sposób kilka typów RNA HCV zsyntetyzowanych na powyższej matrycy DNA, pochodzących z pjfh1 i pjfh1/gnd nazwano symbolami rjfh1, rjfh1/gnd, rjfh1/ E-E2, rj6cf, rjch1 i rjch1/ns5b(jfh1). [0101] W odniesieniu do tak otrzymanych RNA HCV, rjfh1 stanowi RNA wytworzony z użyciem DNA skatalogowanego w GenBanku pod numerem dostępu AB jako matrycy; JFH1/GND jest to RNA wytworzony z użyciem jako matrycy, DNA otrzymanego przez podstawienie G przy nukleotydzie 8618 przez A, w DNA skatalogowanym w GenBanku pod numerem dostępu AB047639; rjfh1/ E-E2 jest to RNA wytworzony z użyciem jako matrycy, DNA z delecją w
44 45 części sekwencji w DNA skatalogowanym w GenBanku pod numerem dostępu AB047639; rj6cf jest to RNA wytworzony z użyciem jako matrycy, DNA skatalogowanego w GenBanku pod numerem dostępu AF177036; rjch1 jest to RNA wytworzony z użyciem jako matrycy, DNA skatalogowanego w GenBanku pod numerem dostępu AB047640; i rjch1/ns5b(jfh1) jest to RNA wytworzony z użyciem jako matrycy, DNA uzyskanego przez ligację części sekwencji DNA z DNA skatalogowanego w GenBanku pod numerem dostępu AB do części sekwencji DNA z DNA skatalogowanego w GenBanku pod numerem dostępu AB047639, wykorzystując miejsce dla enzymu restrykcyjnego AvrII. Sekwencje nukleotydowe tych RNA można potwierdzić. Ten ostatni zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C jest rozwiązaniem według niniejszego wynalazku. [Przykład 2] Generowanie komórek replikujących genomowy RNA HCV i cząsteczek wirusa w komórkach 1. Replikacja genomu HCV i generowanie cząsteczek wirusa w komórkach [0102] Ilości każdego z wyżej zsyntetyzowanych genomowych RNA HCV o pełnej długości (rjfh1 i rjfh1/gnd) tak dobrano, aby poziom całkowitego RNA wynosił 10 µg. Następnie ten zmieszany RNA wprowadzano do komórek Huh7 metodą elektroporacji. Komórki Huh7 po elektroporacji posiewano na płytce do hodowli i następnie prowadzono ich hodowlę przez 12 godzin, 24 godziny, 48 godzin i 72 godziny. Następnie komórki zbierano i z tych komórek ekstrahowano RNA. Wyekstrahowany RNA analizowano techniką hybrydyzacji northern. Analizę Northern prowadzono w sposób opisany w Molecular Cloning, A Laboratory Manual, II wydanie,
45 46 J. Sambrook, E.F. Fritsch, T. Maniatis,; Cold Spring Harbor Laboratory Press (1989). RNA wyekstrahowany z komórek poddano denaturującej elektroforezie na agarozie. Po zakończeniu elektroforezy transkrypcję tego RNA prowadzono na naładowanej dodatnio membranie nylonowej. Znakowaną 32 P sondę DNA albo sondę RNA wytworzoną z pjfh1 poddawano hybrydyzacji z RNA transkrybowanym na tej membranie w sposób opisany powyżej. Następnie, membranę płukano i naświetlano nią film, wykrywając prążek RNA specyficzny dla genomu HCV. [0103] Jak widać na fig. 2, jeśli komórki uległy transfekcji JFH1/GND, wprowadzony prążek RNA był potwierdzony jako słaby sygnał po 4 godzinach od transfekcji. Ten sygnał jednak słabł zależne od czasu i po 24 godzinach prążki sygnałów były prawie niewykrywalne. [104] Z drugiej strony, gdy komórki poddawano transfekcji rjfh1, po 4 do 12 godzinach od transfekcji, moc sygnału wprowadzonego prążka RNA była prawie taka sama jak w przypadku wprowadzenia JFH1/GND. Następnie, sygnał słabnął raz ale wyraźny sygnał prążka RNA można było potwierdzić po 24 godzinach. Ten sygnał był specyficzny dla HCV. Innymi słowy, uznano, że porcja wprowadzonego RNA rjfh1 replikowała się i rosła. Takiej replikacji nie obserwowano w przypadku rjfh1/gnd otrzymanego przez mutację aktywnego motywu NS5B, który jest enzymem replikującym RNA. Tak więc stwierdzono, że aktywność NS5B jest ważna dla replikacji RNA pełnej długości wirusa HCV. Przeprowadzono takie samo doświadczenie, stosując szczep JCH1 [Kato T. i wsp., J. Med. Virol. 69 (2001), str ] wyizolowany przez twórców niniejszego wynalazku od pacjentów chorych na
46 47 przewlekłe zapalenie wątroby. W przypadku tego szczepu w ogóle nie stwierdzono replikacji RNA HCV. 2. Detekcja białka HCV [0105] Z komórek transfekowanych RNA rjfh1 albo rjfh1/gnd ekstrahowano białko w sposób zależny od czasu, postępując zgodnie ze znanymi metodami i białko to analizowano metodą elektroforezy SDS-PAGE i techniką Western. W celu przeprowadzenia tej analizy, transfekowano przejściowo komórki Huh7 plazmidem ekspresyjnym DNA zawierającym gen dla NS3, NS5A, rdzeniowy lub E2 i uzyskany ekstrakt komórkowy użyto jako kontrolę dodatnią (białko NS3). Poza tym, białko wyekstrahowane z nietransfekowanych komórek Huh7 użyto jako kontrolę negatywną. Próbkę białka wyekstrahowaną z każdego klonu komórek nanoszono na filtr PVDF (Immobilon-P, wytwarzany przez Millipore). Do detekcji białek NS3, NS5A, białka rdzeniowego i białek E2 kodowanych przez RNA JFH1, użyto przeciwciała swoistego przeciw NS3 (dostarczone przez dr Moradpour; Wolk B. i wsp., J. Virology, 2000; 74: ), przeciwciało swoiste przeciw NS5A (wytworzone przez insercję regionu NS5A w JFH1 do wektora ekspresyjnego i zastosowanie go u myszy zgodnie z procedurami immunizacji DNA), przeciwciało swoiste przeciw białku rdzeniowemu (przeciwciało przeciw klonowi 2H9) i przeciwciało swoiste przeciw białku E2 (wytworzone przez zsyntetyzowanie peptydu o sekwencji GTTTVGGAVARSTN (SEQ ID Nr. 4) w regionie E2 JFH1 i o sekwencji CDLEDRDRSQLSPL (SEQ ID Nr. 5) w tym samym regionie i następnie immunizację królika tymi dwoma syntetycznymi peptydami). Poza tym, jako kontrolę wewnętrzną
47 48 wykrywano białko aktyny stosując przeciwciało przeciw aktynie. [0106] Jak przedstawiono na fig. 3 w komórkach transfekowanych przez rjfh1 od 24 godzin po transfekcji wykrywano białka NS3, NS5A, rdzeniowe i E2, i potwierdzono, że wzrost poziomu ekspresji był zależny od upływu czasu. Odwrotnie, w komórkach transfekowanych przez rjfh1/gnd oraz w nietransfekowanych komórkach Huh7 nie wykrywano żadnego z białek NS3, NS5A, rdzeniowego ani E2. Okazało się, że ekspresja tych białek w komórkach jest wynikiem autonomicznej replikacji transfekowanego rjfh1. [0107] Na podstawie wyników otrzymanych w powyższych punktach 1 i 2 potwierdzono, że rjfh1 replikuje się w komórkach ustanowionych przez transfekcję za pomocą rjfh1. 3. Detekcja białka rdzeniowego HCV w pożywce hodowli transfekowanych komórek. [0108] Komórki Huh7, do których przez elektroporację wprowadzono rjfh1, rjfh1/gnd, rjfh1/ E1-E2, rj6cf i rjch1 posiano na płytki hodowlane. Hodowlę komórek prowadzono przez 2 godziny, 12 godzin, 24 godziny, 48 godzin i 72 godziny. Następnie oznaczono zawartość białka rdzeniowego HCV w pożywce hodowli. Pomiary prowadzono testem IRMA z wykorzystaniem antygenu HCV Ortho [Aoyagi i wsp., J. Clin. Microbiol., 37 (1999) str ]. [0109] Jak to jest uwidocznione na fig. 4, białko rdzeniowe wykrywano w pożywce hodowli po 48 do 72 godzin od transfekcji za pomocą rjfh1. Z drugiej strony, w pożywce hodowli komórek transfekowanych rjfh1/gnd, rj6cf i rjch1 nie wykrywano białek
48 49 rdzeniowych HCV. W pożywce hodowli komórek transfekowanych rjfh1/ E1-E2 wykrywano niewielkie ilości białka rdzeniowego HCV. Tak więc rjfh1/gnd, rj6cf i rjch1 nie mają zdolności autonomicznej replikacji w komórkach Huh7, podczas gdy rjfh1 i rjfh1/ E1-E2 mogą być w nich autonomicznie replikowane. Tak wiec okazało się, że autonomiczna replikacja wprowadzonego RNA HCV odgrywa zasadniczą rolę w uwalnianiu tego białka rdzeniowego i ponadto, że E1 i E2 są niezbędne do stabilnego uwalniania dużych ilości białka rdzeniowego z tych komórek. 4. Detekcja cząsteczek HCV w pożywce hodowli transfekowanych komórek [0110] W celu zanalizowania, czy białko rdzeniowe uwalniane do pożywki hodowli w powyższym przykładzie jest wydzielane w formie cząsteczek wirusa, czy też nie, pożywkę hodowli otrzymaną po 6 dniach od transfekcji prze rjfh1 frakcjonowano przez wirowanie w gradiencie gęstości sacharozy. Mianowicie, 2 ml 60% (wagowo) roztworu sacharozy (rozpuszczonej w 50 mm Tris, ph 7,5/ 0,1 M NaCl/ 1 mm EDTA), 1 ml 50% roztworu sacharozy, 1 ml 40% roztworu sacharozy, 1 ml 30% roztworu sacharozy, 1 ml 20% roztworu sacharozy i 1 ml 10% roztworu sacharozy nanoszono warstwami w probówce do wirowania i następnie od góry naniesiono 4 ml supernatantu hodowli. Probówkę wirowano z prędkością obrotów/minutę w temperaturze 4 o C przez 16 godzin w wirówce Beckmann SW41Ti. Po zakończeniu wirowania od dołu probówki do wirowania pobierano po 0,5 ml każdej frakcji. W każdej frakcji oznaczano gęstość, stężenie białka rdzeniowego HCV i ilość kopii RNA HCV. Przeprowadzono detekcję replikonu RNA metodą
49 50 ilościowej analizy RT-PCR przez detekcję RNA w nie podlegającym translacji regionie 5 RNA HCV, postępując zgodnie z metodą Takeuchi i wsp. [Takeuchi T. i wsp., Gastroenterology 116: (1999)]. Konkretnie, replikon RNA zawarty w RNA wyekstrahowanym z komórek amplifikowano techniką PCR, stosując podane poniżej syntetyczne startery i zestaw EZ rtth RNA PCR (Applied Biosystems). Detekcję prowadzono w układzie ABI Prism 7700 sequence detektor system (Applied Biosystems). R6-130-S17: 5 CGGGAGAGCCATAGTGG-3 (SEQ ID Nr 6) R6-290-R19: 5 -AGTACCACAAGGCCTTTCG-3 (SEQ ID Nr 7) Sonda TaqMan, R6-148-S21FT: 5 -CTGCGGAACCGGTGAGTACAC-3 (SEQ ID Nr 8). [0111] Jak to jest przedstawione na fig. 5A, pik białka rdzeniowego odpowiadał RNA HCV we frakcji o gęstości 1,17 mg/ml. Gęstość tej frakcji wynosiła około 1,17 mg/ml. Jest to gęstość właściwa mniejsza od gęstości właściwej produktu związanego, złożonego z białka rdzeniowego i kwasu nukleinowego, opisanego poprzednio. Jeśli białko rdzeniowe HCV i RNA HCV występujące we frakcji o gęstości 1,17 mg/ml tworzy strukturę cząsteczek HCV, należy przyjąć, że ta frakcja jest oporna na nukleazę. A więc, na roztwór hodowli uzyskany po 6 dniach od transfekcji JFH1 działano stężeniem 10 µg/ml RNA-zy, w czasie 20 minut i następnie frakcjonowano go w gradiencie gęstości sacharozy. [0112] Jak przedstawiono na fig. 5B, w rezultacie, RNA HCV uległ rozłożeniu i pik dla białka rdzeniowego i dla RNA HCV wykryto we frakcji 1,17 mg/ml, tak, jak w przypadku próby nietraktowanej RNA-zą A. W ten sposób potwierdzono, że białko rdzeniowe i RNA HCV występujące we frakcji 1,17 mg/ml tworzą strukturę podobną do cząsteczek HCV.
50 51 [0113] Następnie roztwór hodowli poddano takiemu samemu frakcjonowaniu jak opisano powyżej, po traktowaniu 0,25 % NP40. W rezultacie, pik odpowiadający białku rdzeniowemu i RNA HCV przesunął się do gęstości 1,28 mg/ml (fig. 5C). Jeśli na roztwór hodowli jednocześnie działano 0,25% NP40 i RNA-zą A, pik dla RNA HCV zanikał (fig. 5D). Uznano więc, że błona powierzchniowa zawierająca lipidy o niskim ciężarze właściwym uległa odwarstwieniu z cząsteczek wirusa w wyniku traktowania przez NP40 i cząsteczki wirusa stały się cząsteczkami jedynie rdzeniowymi, złożonymi tylko z kwasu nukleinowego i białka rdzeniowego, nie mającymi struktury podobnej do struktury wirusa, co spowodowało wzrost gęstości właściwej. [0114] Na podstawie powyższych wyników potwierdzono, że wirusowy RNA replikuje się po transfekcji komórek Huh7 przez rjfh1 i że dzięki temu powstają cząsteczki wirusa i uwalniają się one do roztworu hodowli. 5. Doświadczenie dotyczące infekcyjności cząsteczek wirusa w pożywce hodowli [0115] Komórki Huh7 transfekowano rjfh1 i oznaczano infekcyjność cząsteczek HCV wydzielonych do pożywki hodowlanej. Po 3 dniach od transfekcji komórek Huh7 przez rjfh1 albo rjfh1/ E1-E2 odzyskiwano supernatant hodowli. Odzyskaną pożywkę hodowlaną wirowano i supernatant po wirowaniu zebrano po czym przefiltrowano go przez filtr 0,47 µm. W obecności tej pożywki hodowlanej prowadzono hodowlę komórek Huh7, które nie były uprzednio transfekowane przez RNA. Po 48 godzinach komórki poddawano fluorescencyjnemu barwieniu immunologicznemu przeciwciałem przeciw rdzeniowi albo przeciwciałem anty-ns5a. Jak pokazano na fig. 6A, w
51 52 przypadku komórek hodowanych w obecności pożywki hodowli uzyskanej z transfekcji komórek Huh7 przez rjfh1, w komórkach tych obserwowano ekspresję białka rdzeniowego i białka NS5A. Z drugiej strony, w przypadku komórek hodowanych w obecności pożywki hodowlanej uzyskanej przez transfekcję komórek Huh7 przez rjfh1/ E1-E2, nie obserwowano w tych komórkach takiej ekspresji białka rdzeniowego i białka NS5A (dane nie są przedstawione). [0116] Następnie, zebrano supernatant hodowli po 3 dniach od transfekcji komórek rhuh7 przez JFH1, zatężono go 30-krotnie stosując filtr do ultrafiltracji o wartości odcięcia 1 x 10 5 Da. Komórki Huh7, które nie były transfekowane przez RNA hodowano w 100 µl pożywki hodowli zawierającej zatężone cząsteczki HCV na szkiełku nakrywkowym o średnicy 15 mm. Po 48 godzinach komórki poddawano barwieniu immunologicznemu przeciwciałem przeciw rdzeniowi i zliczano ilość komórek immunododatnich barwionych przeciw białku rdzeniowemu, czyli komórek zakażonych. W wyniku, jak przedstawiono na fig. 6B, potwierdzono obecność 394,0 ± 26,5 zakażonych komórek (co stanowi około 0,51% całkowitej ilości komórek). Następnie zbadano, czy to zakażenie jest spowodowane przez cząsteczki HCV, które zostały wydzielone do pożywki hodowlanej w wyniku transfekcji komórek Huh7 przez rjfh1. Stosując pożywkę hodowli przygotowaną przez poddanie roztworu hodowli użytego do zakażenia działaniu promieniowania UV i inną pożywkę hodowli przygotowaną z pominięciem etapu transfekcji przez RNA, prowadzono hodowlę komórek Huh7, które nie były poddane transfekcji przez RNA na szkiełku nakrywkowym o średnicy 15 mm. Po 48 godzinach komórki poddawano barwieniu immunologicznemu
52 53 przeciwciałem przeciw rdzeniowi i zliczano ilość zakażonych komórek. W przypadku, gdy komórki były napromieniane UV, ilość zakażonych komórek gwałtownie spadła. W przypadku pożywki hodowli uzyskanej z pominięciem etapu transfekcji RNA, nie obserwowano żadnych zakażonych komórek. [0117] Poza tym sprawdzono, czy zakażające cząsteczki HCV amplifikują RNA w tych komórkach i następnie uwalniają nowe cząsteczki HCV do pożywki hodowli. Komórki Huh7 nie poddawane transfekcji RNA hodowano w 100 µl pożywki hodowli zawierającej cząsteczki HCV przygotowanej przez zatężenie pożywki hodowli otrzymanej po 48 godzinach od transfekcji komórek Huh7 przez rjfh1. Następnie, w odstępach jednodniowych pobierano komórki i pożywkę hodowli i odzyskiwano z nich RNA. Ilość RNA HCV oznaczano wyżej opisaną metodą. Jak to jest przedstawione na fig. 6C, RNA HCV ulegał amplifikacji w komórkach do pewnej ilości i w supernatancie ilość RNA HCV wzrastała w sposób zależny od czasu. To samo badanie przeprowadzono z użyciem roztworu hodowli uzyskanego po transfekcji komórek Huh7 przez rjfh1/ E1-E2. Nie wykryto jednakże RNA HCV ani w komórkach ani w roztworze hodowli. [0118] Na podstawie tych wyników potwierdzono, że cząsteczki HCV wydzielane do pożywki hodowli mają cechę infekcyjności będącą wynikiem transfekcji komórek Huh7 przez rjfh1 i mają również zdolność amplifikacji RNA HCV w zakażonych komórkach i wytwarzania cząsteczek HCV. 6. Wytwarzanie cząsteczek wirusa HCV z użyciem rjch1/ns5b(jfh1)
53 54 [0119] Badano, czy cząsteczki HCV są wydzielane do pożywki hodowli w wyniku transfekcji komórek Huh7 przez rjch1/ns5b(jfh1) i czy wydzielane cząsteczki HCV posiadają cechę infekcyjności. Roztwór hodowli otrzymanej po 6 dniach od transfekcji komórek Huh7 przez rjch1/ns5b(jfh1) zatężono sposobem opisanym w powyższej sekcji 5. W obecności tej pożywki hodowli, hodowano komórki Huh7 które nie były poddawane transfekcji przez RNA i śledzono zależne od czasu zmiany w ilości RNA HCV w tych komórkach. Począwszy od 12 godzin po założeniu hodowli, ilość RNA HCV w tych komórkach wzrastała w sposób zależny od czasu (fig. 7A). Poza tym, komórki Huh7 które nie były poddawane transfekcji przez RNA hodowano na szkiełku przykrywkowym o średnicy 15 mm i następnie komórki te hodowano w obecności zatężonej pożywki hodowli. Po 48 godzinach, komórki barwiono immunologicznie przeciwciałem przeciw rdzeniowi i zliczano ilość komórek dodatnich w reakcji z przeciwciałem przeciw rdzeniowi, czyli komórek zakażonych. Jak przedstawiono na fig. 7B, obserwowano komórki zakażone. Na podstawie tych wyników okazało się, że cząsteczki HCV wydzielane do pożywki hodowli nabywają cechę infekcyjności w wyniku transfekcji komórek Huh7 przez rjch1/ns5b(jfh1) i mają również zdolność amplifikacji RNA HCV w zakażonych komórkach i wytwarzania nowych cząsteczek HCV. [0120] Zgodnie z powyższym, nawet w przypadku szczepu, który nie ma zdolności autonomicznej replikacji in vitro, takiego jak szczep HCV wyizolowany od pacjentów, substytucja jego regionu HS5B przez NS5B z rjfh1 umożliwia jego autonomiczną replikację w układzie hodowli komórkowej i generowanie cząsteczek HCV.
54 55 [Przykład 3] 1. Wytwarzanie cząsteczek wirusa HCV z użyciem Con1/C-NS2/JFH-1 [0121] Komórki Huh7 poddawano transfekcji chimerycznym RNA HCV obejmującym część NS5B ze szczepu Con-1 o genotypie 1b HCV i ze szczepu JFH-1, i następnie badano, czy cząsteczki HCV są wydzielane do roztworu hodowli i czy wydzielane cząsteczki HCV mają cechę infekcyjności. [0122] Sekwencję ze szczepu Con-1 o genotypie b1 HCV odpowiadającą regionowi od 1 do 1026 (regiony rdzenia, E1, E2, p7 i NS2 w szczepie Con1) poddano ligacji ze szczepem JFH-1, w dół od 5 -UTR. Następnie w dół od niego dołączono przez ligację region ze szczepu JFH-1 (od NS3 do NS5B). Następnie, w dół od niego dołączono przez ligację 3 -UTR ze szczepu JFH-1, wytwarzając konstrukt. Z użyciem tego konstruktu wytworzono chimerowy RNA HCV, rcon1/c-ns2/jfh-1, sposobem opisanym w powyższym przykładzie 1-2. Powyższym RNA transfekowano komórki Huh7 sposobem opisanym w powyższym przykładzie 2-1. Komórki Huh7 transfekowano RNA HCV i oznaczano białko rdzeniowe zawarte w supernatancie względem czasu. Począwszy od czasu 48 godzin, białko rdzenia wykrywano w supernatancie, a zatem, można było stwierdzić, że cząsteczki HCV są generowane w supernatancie komórkowym. Ten supernatant zatężono 20-krotnie metodą ultrafiltracji i koncentrat dodano do komórek Huh7. Po 48-godzinnej hodowli komórki barwiono króliczym przeciwciałem anty-ns3. [0123] Nie wykryto obecności komórek dodatnich, reagujących na przeciwciało anty-ns3 w ślepych próbkach
55 56 i w rjfh-1/ EE1-E2 ale takie komórki reagujące z przeciwciałem anty-ns3 wykryto w rjfh-1 i w rcon1/c- NS2/JFH-1. Na podstawie powyższych wyników można stwierdzić, że rcon1/c-ns2/jfh-1 ma zdolność generowania infekcyjnych cząsteczek HCV, tak jak JFH-1. [Przykład 4] Wytwarzanie chimerycznego replikonu RNA HCV o pełnej długości, wyprowadzonego z genomowego RNA chimerycznego HCV o pełnej długości (1) Konstruowanie wektora ekspresyjnego [0124] DNA (klon JFH-1: SEQ ID Nr 9) zawierający genomowy cdna o pełnej długości ze szczepu JFH-1 (genotyp 2a), będącego wirusem zapalenia wątroby typu C wyizolowanym od pacjentów z piorunującym zapaleniem wątroby, wprowadzono przez insercję do plazmidu puc19, w dół od sekwencji promotora RNA T7, wytwarzając plazmidowy DNA. [0125] Konkretnie, fragment z RT-PCR otrzymany przez amplifikację wirusowego RNA ze szczepu JFH-1 klonowano do wektora pgem-t EASY (Promega), otrzymując różne plazmidowe DNA, takie jak pgem1-258, pgem44-486, pgem , pgem , pgem , pgem , pgem , pgem , pgem , pgem , pgem , pgem , pgem i pgem [Kato i wsp., Gastroenterology, (2003) 125: ]. cdna wyprowadzony z wirusowego genomowego RNA zawarty w każdym z tych plazmidów poddawano ligacji jeden do drugiego, metodą PCR i stosując enzymy restrykcyjne, po czym taki genomowy cdna o pełnej długości klonowano. W górę od genomu wirusowego o pełnej długości wprowadzono przez insercję sekwencję promotora T7 RNA. Tak skonstruowany
56 57 plazmidowy DNA jest w dalszej części opisu oznaczony symbolem pjfh1. Należy zauważyć, że wytwarzanie powyższego klonu JFH-1 opisano w publikacji patentowej Japonii (Kokai), nr A i w publikacji Kato i wsp. [Kato i wsp., J. Med. Virol. (2001) 64(3): ]. Ponadto sekwencję nukleotydową cdna o pełnej długości z klonu JFH-1 zarejestrowano w Międzynarodowej Bibliotece DNA (DDBJ/EMBL/GenBank) pod numerem dostępu AB [0126] Następnie, pomiędzy nie podlegający translacji region 5 i region rdzeniowy pjfh1, który jest plazmidowym DNA, wprowadzono przez insercję EMCV-IRES (wewnętrzne miejsce wejścia dla rybosomu dla wirusa zapalenia mózgu i mięśnia sercowego) i gen oporności na neomycynę (neo; nazwany również genem fosfotransferazy neomycynowej), konstruując pfgrep- JFH1, który jest plazmidowym DNA. Ten konstrukt przeprowadzono w sposób opisany przez Ikeda i wsp. [Ikeda i wsp., J. Virol. (2002) 76(6): ]. (2) Konstruowanie chimerycznego wektora ekspresyjnego [0127] Szczep JFH1 jest HCV wyprowadzonym z HCV o typie 2a. Szczep TH pochodzący ze szczepu HCV typu 1b [Wakita i wsp., J. Biol. Chem. (1994) 269: ; i Moradpour i wsp., Biochem. Biophys. Res. Commun. (1998) 246: ] użyto do wytworzenia chimerycznego wektora HCV. Części rdzeniową, E1, E2 i p7 w pfgrep-jfh1 wytworzonym powyżej zastąpiono odpowiednimi częściami ze szczepu TH, wytwarzając chimeryczny HCV, pfgrep- TH/JFH1. [0128] W niniejszym opisie sekwencja RNA o pełnej długości z powyższego szczepu JFH1 (wyprowadzonego z
57 58 klonu JFH-1) i częściowa sekwencja RNA szczepu TH użyta do wytworzenia powyższego ciałka chimerycznego (częściowy genomowy RNA (1-3748) zawierający część odpowiadającą regionowi z niepodlegającego translacji regionu 5 w szczepie TH HCV do jego regionu NS3) są przedstawione odpowiednio jako sekwencje o SEQ ID nr 9 i 10. W powyższej genomowej sekwencji RNA o pełnej długości ze szczepu JFH-1 (SEQ ID Nr 9), nie podlegający translacji region 5 odpowiada pozycjom 1-340, sekwencja kodująca białko rdzenia odpowiada pozycjom , sekwencja kodująca białko E1 odpowiada pozycjom , sekwencja kodująca białko E2 odpowiada pozycjom , sekwencja kodująca białko NS2 odpowiada pozycjom , sekwencja kodująca białko NS3 odpowiada pozycjom , sekwencja kodująca białko NS4A odpowiada pozycjom , sekwencja kodująca białko NS4B odpowiada pozycjom , sekwencja kodująca białko NS5A odpowiada pozycjom i sekwencja kodująca białko NS5B odpowiada pozycjom (3) Wytwarzanie replikonu RNA chimerycznego HCV o pełnej długości [0129] W celu wytworzenia matrycowego DNA stosowanego do syntezy replikonu RNA chimerycznego HCV o pełnej długości, skonstruowany powyżej wektor ekspresyjny pfgrep-th/jfh1 trawiono enzymem restrykcyjnym XbaI. Następnie ilości 10 µg do 20 µg fragmentu po trawieniu XbaI zmieszano z 50µl roztworu reakcyjnego i otrzymaną mieszaninę inkubowano z 20 jednostkami nukleazy fasolo mung, w temperaturze 30 o, w czasie 30 minut. Nukleaza Mung Bean jest enzymem katalizującym reakcję selektywnego trawienia jednoniciowego odcinka
58 59 dwuniciowego DNA. W zasadzie, jeśli RNA syntetyzuje się bezpośrednio, stosując wyżej wymieniony fragment trawienia XbaI jako matrycę, wówczas powstaje replikon RNA, do którego przy końcu 3 dodaje się w nadmiarze 4 nukleotydy CUGA stanowiące część sekwencji rozpoznawanej przez XbaI. Tak więc w niniejszym przykładzie, taki fragment po trawieniu XbaI traktowano nukleazą Mung Bean w celu usunięcia z tego fragmentu XbaI 4 nukleotydów CUGA. Następnie roztwór traktowany nukleazą z fasoli mung, zawierający fragment rozszczepienia XbaI poddano traktowaniu usuwającemu białko, postępując w znany sposób, i w ten sposób oczyszczono fragment po trawieniu XbaI, z którego usunięto 4 nukleotydy CUGA. Oczyszczony fragment użyto jako matrycowy DNA. [0130] Następnie na matrycy DNA syntetyzowano RNA in vitro, stosując polimerazę RNA T7. Do tej syntezy RNA użyto MEGAscript wytwarzany przez firmę Ambion. Ilość 20 µl roztworu reakcyjnego zawierającego 0,5 do 1,0 µg matrycowego DNA poddawano reakcji, postępując według instrukcji dostarczonej przez producenta. [0131] Po zakończeniu syntezy RNA do roztworu reakcyjnego dodano 2 jednostki DNA-zy i otrzymaną mieszaninę utrzymywano w 37 o C przez 15 minut. Następnie ekstrahowano RNA kwaśnym fenolem i usunięto matrycowy DNA. RNA zsyntetyzowany z powyższego matrycowego DNA wyprowadzonego z pfgrep-th/jfh1 oznaczono symbolem rfgrep-th/jfh1. Sekwencję nukleotydową chimerycznego genomowego RNA HCV w tym rfgrep-th/jfh1 przedstawiono jako SEQ ID Nr 11. Ten rfgrep-th/jfh1 jest przykładem replikonu RNA chimerycznego HCV o pełnej długości według wynalazku.
59 60 [Przykład 5] Wytwarzanie komórek replikujących replikon RNA chimerycznego HCV o pełnej długości i założenie klonu komórkowego (1) Wprowadzenie genomowego RNA chimerycznego HCV o pełnej długości do komórek [0132] Różne ilości genomowego RNA z chimerycznego HCV o pełnej długości (rfgrep-th/jfh1) zsyntetyzowanego w sposób podany powyżej zmieszano z całkowitym komórkowym RNA ekstrahowanym z komórek Huh7, otrzymując całkowitą ilość RNA wynoszącą 10 µg. Ten zmieszany RNA wprowadzano następnie do komórek Huh7 metodą elektroporacji. Hodowlę komórek prowadzono w czasie 16 do 24 godzin i następnie dodano do niej G418 w różnych ilościach. Hodowlę kontynuowano, zmieniając roztwór hodowli na świeży dwa razy w tygodniu. Po zakończeniu hodowli po 21 dniach, komórki, które przeżyły, barwiono fioletem krystalicznym. Zliczano ilość kolonii, które się zabarwiły i wyliczano ilość kolonii otrzymanych w stosunku do masy RNA stosowanego do transfekcji. Ponadto, na kilku płytkach z hodowlą sklonowano kolonie żywych komórek i hodowlę kontynuowano. Ze sklonowanych komórek ekstrahowano RNA, genomowy DNA i białko i następnie wykonywano detekcję repliko nu RNA chimerycznego HCV o pełnej długości, sprawdzano obecność lub brak wprowadzenia genu oporności na neomycynę do genomowego DNA i ekspresję białka HCV. Wyniki są szczegółowo przedstawione poniżej. (2) Zdolność tworzenia kolonii [0133] Po powyższej transfekcji obserwowano tworzenie kolonii przez komórki, nawet w przypadku, gdy stężenie G418 wynosiło 1,0 mg/ml. Okazało się, że replikon RNA rfgrep-th/jfh1 ulegał autonomicznej replikacji w
60 61 komórkach Huh7 transfekowanych replikonem RNA rfgrep- TH/JFH1 i że gen oporności na neomycynę podlegał stałej ekspresji, i w ten sposób była utrzymywana oporność na G418. Tak więc komórki były zdolne do wzrostu i komórki Huh7 nabywały zdolności tworzenia kolonii. [Przykład 6] Infekcyjność chimerycznego wirusa HCV w supernatancie hodowli. Doświadczenie dotyczące infekcyjności chimerycznych cząsteczek wirusa HCV w supernatancie hodowli [0134] Komórki Huh7 poddawano transfekcji rfgrep- TH/JFH1, po czym odzyskiwano supernatant hodowli zawierający ustalone klony komórek replikujących replikonowy RNA chimerycznego HCV o pełnej długości. Ten supernatant hodowli dodano do komórek Huh7, które nie były zakażone, a więc komórki Huh7 ulegały zakażeniu cząsteczkami wirusa obecnymi w supernatancie hodowli. Następnego dnia po zakażeniu do pożywki hodowlanej zawierającej zakażone komórki Huh7 dodano 0,3 mg/ml G418 i dalej prowadzono hodowlę mieszaniny przez 21 dni. Po zakończeniu hodowli komórki utrwalano i barwiono fioletem krystalicznym. W wyniku tego obserwowano tworzenie się kolonii w komórkach zakażonych supernatantem hodowli zawierającym klony komórek replikujących replikonowy RNA chimerycznego HCV o pełnej długości, uzyskanych przez transfekcję przez rfgrep-th/jfh1. Oznacza to, że klony komórek replikujących replikonowy RNA chimerycznego HCV o pełnej długości, uzyskanych przez transfekcję rfgrep- TH/JFH1, generują infekcyjny HCV, a także, że ten HCV jest infekcyjny w stosunku do nowych komórek.
61 62 [Przykład 7] Oczyszczanie cząsteczek HCV (1) Filtracja żelowa [0135] Na fig. 11 przedstawiono rozkład cząsteczek HCV w każdej frakcji otrzymanej z filtracji żelowej. Jako nośniki żelowe stosowano Sephacryl S300, S400 i S500. Roztwór zawierający cząsteczki HCV użyty do chromatografii kolumnowej oczyszczano metodą chromatografii kolumnowej na każdym z powyższych nośników żelowych. Bufor stosowany do oczyszczania miał skład: 10 mm Tris.HCl, 1 mm kwas etylenodiaminotetraoctowy i 100 mm chlorek sodu (ph = 8,0). W rezultacie, w przypadku użycia żelu Sephacryl S-300, uzyskano cząsteczki HCV w wypływającej frakcji zwanej frakcją pustą. Tak więc, stosując Sephacryl S- 300 oddzielono cząsteczki HCV od białek o niskiej masie cząsteczkowej, a więc powinno się zmienić stężenie soli w roztworze. Stosunek białka rdzeniowego HCV do masy białka całkowitego wynosił 3,78 w porównaniu do cząsteczek HCV przed oczyszczaniem kolumnowym, a więc stosunek cząsteczek HCV do białka całkowitego wzrósł. Z drugiej strony, w przypadku użycia żelu Sephacryl S- 400 i S-500, jako, że cząsteczki HCV otrzymano jako frakcję eluującą zależnie od masy cząsteczkowej, cząsteczki te można oddzielić od innych białek o różniących się od nich masach cząsteczkowych. (2) Chromatografia jonowymienna [0136] Na fig. 12 przedstawiono rozmieszczenie cząsteczek HCV w każdej frakcji uzyskanej z chromatografii jonowymiennej. Jako nośniki żelowe stosowano: SP Sepharose HP i Q-Sepharose HP.
62 63 [0137] W przypadku kolumny wypełnionej żelem SP Sepharose HP, kolumnę równoważono buforem z 50 mm kwasem cytrynowym (ph 6,2). Roztwór zawierający cząsteczki HCV uprzednio zatężony na filtrze do ultrafiltracji o wartości odcięcia masy cząsteczkowej od do i następnie rozcieńczony buforem 50 mm kwasu cytrynowego (ph 6,2), wprowadzono na kolumnę. Następnie przez kolumnę przepuszczano bufor 50 mm kwasu cytrynowego (ph 6,2) w objętości stanowiącej około 10 objętości kolumny. Następnie przez kolumnę przepuszczano kolejno bufory z 50 mm kwasem cytrynowym (ph 6,2), do których dodano 0,1 M NaCl, 0,3 M NaCl albo 1 M NaCl, w objętościach stanowiących około 3-krotną objętość kolumny. Następnie, przez kolumnę przepuszczono bufor 50 mm kwasu cytrynowego (ph 6,2), do którego dodano 1 M NaCl, w objętości stanowiącej około 5-krotną objętość kolumny (frakcja 1 M NaClW). Cząsteczki HCV były eluowane z kolumny we frakcji 50 mm kwasu cytrynowego (ph 6,2) z dodatkiem 0,1 M NaCl. [0138] W przypadku kolumny wypełnionej żelem Q Sepharose HP kolumnę równoważono buforem 50 mm Tris.HCl (ph 8,0). Roztwór zawierający cząsteczki HCV, uprzednio zatężony na filtrze do ultrafiltracji o wartości odcięcia masy cząsteczkowej od do i następnie rozcieńczony buforem z 50 mm Tris.HCl (ph 8,0) wprowadzono na kolumnę. Następnie przez kolumnę przepuszczano bufor 50 mm Tris.HCl (ph 8,0) w objętości stanowiącej około 10 objętości kolumny. Następnie przez kolumnę przepuszczano kolejno bufory 50 mm Tris.HCl (ph 8,0), do których dodano 0,1 M NaCl, 0,3 M NaCl albo 1 M NaCl, w objętościach stanowiących około 3-krotną objętość kolumny. Następnie przez kolumnę przepuszczono bufor z 50 mm Tris.HCl (ph
63 64 8,0), do którego dodano 1 M NaCl, w objętości stanowiącej około 5-krotną objętość kolumny (frakcja 1 M NaClW). Cząsteczki HCV były eluowane z kolumny we frakcji 50 mm Tris.HCl (ph 8,0) z dodatkiem 0,3 M NaCl. Stosunek białka rdzeniowego HCV do masy całkowitego białka wyniósł 2,32 w porównaniu do cząsteczek HCV przed oczyszczaniem kolumnowym, a więc, stosunek cząsteczek HCV do całkowitego białka wzrósł. (3) Chromatografia powinowactwa [0139] Na fig. 13 przedstawiono dystrybucję cząsteczek HCV w każdej frakcji uzyskanej z chromatografii powinowactwa na lektynie. W tej chromatografii powinowactwa użyto nośniki, z którymi wiąże się każdy z RCA-120, ConA, LCA i WGA. [0140] W przypadku chromatografii powinowactwa ConA, LCA i WGA, kolumnę równoważono solą fizjologiczną buforowaną fosforanem. Roztwór zawierający cząsteczki HCV uprzednio zatężony na filtrze do ultrafiltracji o wartości odcięcia masy cząsteczkowej od do i następnie rozcieńczony buforowaną fosforanem solą fizjologiczną wprowadzono na kolumnę. Następnie przepuszczano przez kolumnę buforowaną fosforanem sól fizjologiczną w objętości stanowiącej około 10 objętości kolumny. Następnie przez kolumnę przepuszczano buforowaną fosforanem sól fizjologiczną, do której dodano 0,35 M laktozy, w objętości stanowiącej około 3-krotną objętość kolumny. Następnie przez kolumnę przepuszczano buforowaną fosforanem sól fizjologiczną, do której dodano 0,5 M laktozy, w objętości stanowiącej około 5-krotną objętość kolumny. W rezultacie, w przypadku chromatografii powinowactwa LCA i ConA, nie obserwowano żadnego swoistego wiązania
64 65 z nośnikiem. W przypadku chromatografii powinowactwa WGA, cząsteczki HCV eluowały we frakcji buforowanej fosforanem soli fizjologicznej, do której dodano 0,35 M laktozę. [0141] W przypadku chromatografii powinowactwa RCA-120, kolumnę równoważono solą fizjologiczną buforowaną fosforanem. Roztwór zawierający cząsteczki HCV uprzednio zatężony na filtrze do ultrafiltracji o wartości odcięcia masy cząsteczkowej od do i następnie rozcieńczony buforowaną fosforanem solą fizjologiczną wprowadzono na kolumnę. Następnie przepuszczano przez kolumnę buforowaną fosforanem sól fizjologiczną w objętości stanowiącej około 10 objętości kolumny. Następnie przez kolumnę przepuszczano buforowaną fosforanem sól fizjologiczną, do której dodano 0,38 M laktozy, w objętości stanowiącej około 3-krotną objętość kolumny. Następnie przez kolumnę przepuszczano buforowaną fosforanem sól fizjologiczną, do której dodano 0,38 M laktozy, w objętości stanowiącej około 5-krotną objętość kolumny. W rezultacie, w przypadku chromatografii powinowactwa RCA-120, cząsteczki HCV eluowały we frakcji buforowanej fosforanem soli fizjologicznej, do której dodano 0,38 M laktozę. [0142] Na fig. 14 jest przedstawiono dystrybucję cząsteczek HCV w każdej frakcji uzyskanej z chromatografii powinowactwa z użyciem heparyny i siarczanu celulofiny. [0143] W każdej chromatografii powinowactwa, kolumnę równoważono 20 mm buforem fosforanowym (ph 7,0). Na kolumnę wprowadzano roztwór zawierający cząsteczki HCV, uprzednio zatężony na filtrze do ultrafiltracji o wartości odcięcia masy cząsteczkowej od do
65 i następnie rozcieńczony 20 mm buforem fosforanowym (ph 7,0). Następnie, przez kolumnę przepuszczano bufor fosforanowy (ph 7,0) w objętości stanowiącej około 10 objętości kolumny. Następnie przez kolumnę przepuszczano kolejno bufory fosforanowe (ph 7,0), do których dodano 0,1 M NaCl, 0,3 m NaCl, 0,5 M NaCl i 1 M NaCl w objętościach stanowiących około 3- krotną objętość kolumny. Następnie przez kolumnę przepuszczano 20 mm bufor fosforanowy (ph 7,0), do którego dodano 1 M NaCl w objętości stanowiącej około 5-krotną objętość kolumny. W rezultacie, w przypadku chromatografii powinowactwa z użyciem heparyny, cząsteczki HCV eluowały we frakcji 20 mm buforu fosforanowego (ph 7,0), do którego dodano 0,3 M NaCl. Stosunek białka rdzeniowego do masy białka całkowitego wyniósł 0,36 w porównaniu do cząsteczek HCV przed oczyszczaniem kolumnowym, a więc, stosunek cząsteczek HCV do całkowitego białka spadł. W przypadku chromatografii powinowactwa na siarczanie celulofiny, cząsteczki HCV eluowały we frakcji 20 mm buforu fosforanowego (ph 7,0), do którego dodano 0,1 M NaCl. [0144] Na fig. 15 przedstawiono dystrybucję cząsteczek HCV w każdej frakcji uzyskanej z chromatografii powinowactwa z użyciem barwnika niebieskiego. [0145] W chromatografii powinowactwa z barwnikiem niebieskim jako wypełnienie kolumny użyto nośnika otrzymanego przez związanie barwnika Cibacron Blue F3G- A z cząstkami agarozy. Kolumnę równoważono 20 mm buforem fosforanowym (ph 7,0). Na kolumnę wprowadzano roztwór zawierający cząsteczki HCV uprzednio zatężony na filtrze do ultrafiltracji o wartości odcięcia masy cząsteczkowej od do i następnie rozcieńczony 20 mm buforem fosforanowym (ph 7,0).
66 67 Następnie przez kolumnę przepuszczano bufor fosforanowy (ph 7,0) w objętości stanowiącej około 10 objętości kolumny. Potem przez kolumnę przepuszczano kolejno bufory fosforanowe (ph 7,0), do których dodano 1 M albo 2M NaCl w objętościach stanowiących około 3-krotną objętość kolumny. Następnie przez kolumnę przepuszczano 20 mm bufor fosforanowy (ph 7,0), do którego dodano 2 M NaCl w objętości stanowiącej około 5-krotną objętość kolumny. W rezultacie, cząsteczki HCV eluowały z kolumny we frakcji nie związanej. Stosunek białka rdzeniowego do masy białka całkowitego wyniósł 3,33 w porównaniu do cząsteczek HCV przed oczyszczaniem kolumnowym, a zatem, stosunek cząsteczek HCV do całkowitego białka wzrósł. (4) Wirowanie w gradiencie gęstości sacharozy [0146] Cząsteczki HCV oczyszczano połączonymi metodami chromatografii kolumnowej i wirowania w gradiencie gęstości sacharozy, w odniesieniu do poprzednich przykładów. [0147] Na początku cząsteczki HCV oczyszczano na żelu Q Sepharose HP. Kolumnę zrównoważono buforem z 50 mm Tris.HCl (ph 8,0). Roztwór zawierający cząsteczki HCV uprzednio zatężony na filtrze do ultrafiltracji o wartości odcięcia masy cząsteczkowej od do i następnie rozcieńczony buforem 50 mm Tris.HCl (ph 8,0) wprowadzono na kolumnę. Następnie przez kolumnę przepuszczano bufor z 50 mm Tris.HCl (ph 8,0) w objętości stanowiącej około 10 objętości kolumny. Następnie przez kolumnę przepuszczano kolejno bufory z 50 mm Tris.HCl (ph 8,0), do których dodano 0,1 M NaCl, 0,3 M NaCl albo 1 M NaCl, w objętościach stanowiących około 3-krotne objętości kolumny. Następnie przez
67 68 kolumnę przepuszczono bufor z 50 mm Tris.HCl (ph 8,0), do którego dodano 1 M NaCl, w objętości stanowiącej około 5-krotną objętość kolumny (frakcja 1 M NaClW). W rezultacie, jak przedstawiono na fig. 16A, cząsteczki HCV były eluowane z kolumny we frakcji z 50 mm Tris.HCl (ph 8,0) z dodatkiem 0,3 M NaCl; we frakcji 50 mm Tris.HCl (ph 8,0) z dodatkiem 1,0 M NaCl; i we frakcji 1 M NaClW. Frakcje zawierające cząsteczki HCV zebrano. Stosunek białka rdzeniowego HCV do masy całkowitego białka wyniósł 2,29 w porównaniu do cząsteczek HCV przed oczyszczaniem kolumnowym, a więc stosunek cząsteczek HCV do całkowitego białka wzrósł. [0148] Następnie cząsteczki HCV oczyszczano metodą chromatografii powinowactwa z użyciem siarczanu cellulofiny. W każdej chromatografii kolumnę równoważono 20 mm buforem fosforanowym (ph 7,0). Na kolumnę wprowadzano roztwór zawierający cząsteczki HCV, uprzednio zatężony na filtrze do ultrafiltracji o wartości odcięcia masy cząsteczkowej od do , frakcje zawierające cząsteczki HCV oczyszczone na żelu Q Sepharose HP i następnie rozcieńczone 20 mm buforem fosforanowym (ph 7,0). Następnie przez kolumnę przepuszczano bufor fosforanowy (ph 7,0) w objętości stanowiącej około 10 objętości kolumny. Następnie przez kolumnę przepuszczano kolejno bufory fosforanowe (ph 7,0), do których dodano 0,25 M NaCl albo 1 M NaCl, w objętościach stanowiących około 3-krotną objętość kolumny. Następnie przez kolumnę przepuszczano 20 mm bufor fosforanowy (ph 7,0), do którego dodano 1 M NaCl w objętości stanowiącej około 5-krotną objętość kolumny. W rezultacie, jak to przedstawiono na fig. 16B, cząsteczki HCV eluowały głównie w 20 mm buforze fosforanowym (ph 7,0), do którego dodano 1,0 M NaCl.
68 69 Stosunek białka rdzeniowego HCV do masy białka całkowitego w 20 mm buforze fosforanowym (ph 7,0), do którego dodano 1 M NaCl wyniósł 31,4 w porównaniu do cząsteczek HCV przed oczyszczaniem kolumnowym. Tak więc, stosunek cząsteczek HCV do całkowitego białka wzrósł. [0149] Następnie, cząsteczki HCV oczyszczano metodą wirowania w gradiencie gęstości sacharozy. Frakcję eluowaną 20 mm buforem fosforanowym (ph 7,0), z dodatkiem 1 M NaCl z chromatografii na siarczanie cellulofiny zatężono metodą ultrafiltracji, stosując filtr odcinający masy cząsteczkowe od do i następnie rozcieńczano buforem TEN (bufor 10 mm Tris-HCl (ph 8,0), 0,1 M chlorek sodu i 1 mm kwas etylenodiaminotetraoctowy (ph 8,0)). Roztwór zawierający cząsteczki HCV nawarstwiono na roztwór otrzymany przez nałożenie warstw roztworów 60%, 50%, 40%, 30%, 20% i 10% sacharozy i otrzymany roztwór wirowano przy 390 k x g przez 18 godzin w 4 o C. Ponieważ cząsteczki HCV skupiły się we frakcji o gęstości właściwej około 1,2, frakcję tę zebrano. Stosunek białka rdzeniowego HCV do masy białka całkowitego w zebranej frakcji wyniósł 1,69 w porównaniu do cząsteczek HCV przed oczyszczaniem kolumnowym. Tak więc stosunek cząsteczek HCV do całkowitego białka wzrósł. [0150] We frakcji zawierającej cząsteczki HCV oczyszczone przez wirowanie w gradiencie sacharozy stosunek białka rdzeniowego HCV do masy białka całkowitego wskazał na około 120-krotne oczyszczenie w porównaniu z czystością przed rozpoczęciem chromatografii kolumnowej. Końcowa frakcja zawierała 10 9 kopii/ml cząsteczek wirusa.
69 70 [0151] Wszystkie cytowane w niniejszym opisie publikacje, patenty i zgłoszenia patentowe są włączone do niniejszego opisu w całości. Zastosowanie przemysłowe [0152] Wynalazek umożliwia wytwarzanie cząsteczek wirusa HCV o różnych genotypach w układzie hodowli komórkowej. Oznacza to, że nawet w przypadku szczepu HCV, który nie ma zdolności autonomicznej replikacji in vitro, takiego jak szczepy wirusa HCV wyizolowane od pacjentów, można część ich sekwencji RNA kodującą białka NS3, NS4, NS5A i NS5B zamienić na część sekwencji RNA kodującą białka NS3, NS4, NS5A i NS5B z JFH1, dzięki czemu powyższy szczep HCV nabędzie cechę autonomicznej replikacji w układzie hodowli komórkowej, wytwarzając cząsteczki HCV. Cząsteczki HCV według wynalazku można bezpośrednio użyć jako szczepionki do celów leczniczych. Genomowy RNA HCV albo cząsteczki wirusa uzyskane zgodnie z wynalazkiem można również wykorzystać jako wektor wirusowy dla obcego genu. Ponadto sposób według wynalazku może być również wykorzystany do badań nad procesem zakażenia wirusem HCV albo do produkcji systemu do prób przesiewowych dla różnych substancji, które wywierają wpływ na proces zakażenia wirusem HCV. [0153] Wykaz sekwencji wolny tekst SEQ ID Nr 1: sekwencja kodująca białka od NS3 do NS5 z JFH1 (sekwencja cdna) SEQ ID Nr 2: sekwencja kodująca białka od NS3 do NS5 z JFH1 (sekwencja cdna) SEQ ID Nr 3: białko NS5B z JFH1
70 71 SEQ ID Nr 4: syntetyczny peptyd zaprojektowany na podstawie fragmentu JFH1 E2 SEQ ID Nr 5: syntetyczny peptyd zaprojektowany na podstawie JFH1 E2 SEQ ID Nr 6: Starter (R6-130-S17) SEQ ID Nr 7: Starter (R6-290-R19) SEQ ID Nr 8: Sonda TaqMan (R6-148-S21FT) SEQ ID Nr 9: genomowy RNA wirusa zapalenia wątroby typu C o pełnej długości, wyprowadzony ze szczepu JFH1 (klon JFH-1) SEQ ID Nr 10: sekwencja genomowego RNA zawierająca 5 - UTR do regionu NS3 ze szczepu TH1 SEQ ID Nr 11: Chimerowy genomowy RNA wirusa zapalenia wątroby typu C, wyprowadzony ze szczepu JFH1 HCV (klon JFH-1) i szczepu TH HCV. [0154] WYKAZ SEKWENCJI <110> Tokyo Metropolitan Organization for Medical Research Toray Industries Inc. <120> Genomowy RNA wirusa zapalenia wątroby typu C mający zdolność autonomicznej replikacji
71 72 <210> 1 <211> 6013 <212> DNA <213> Wirus zapalenia wątroby typu C <220> <223> Twórca: Wakita, Takaji Twórca: Kato, Takanobu Twórca: Date, Tomoko Twórca: Bartenchlager, Ralf Twórca: Tanabe, Junichi Twórca: Sone, Saburou <220> <223> Sekwencja kodująca białka NS3 do NS5 z JFH1 (sekwencja cdna) <400> 1

72 73
73 74
74 75 <210> 2 <211> 1773 <212> DNA <213> Wirus zapalenia wątroby typu C <220> <221> CDS <222> (1) (1773) < 220> <223> Sekwencja kodująca białka NS3 do NS5 z JFH1 (sekwencja cdna)
75 76
76 77
77 78
78 79 <210> 3 <211> 591 <212> PRT <213> Wirus zapalenia wątroby typu C <220> <223> białko NS5B z JFH1
79 80
80 81
81 82 <210> 4 <211> 14 <212> PRT <213> Sekwencja sztuczna <220> <223> Opis sekwencji sztucznej: Syntetyczny peptyd wyprowadzony na podstawie fragmentu E2 z JFH1 <210> 5 <211> 14 <212> PRT <213> Sekwencja sztuczna <220> <223> Opis sekwencji sztucznej: Syntetyczny peptyd wyprowadzony na podstawie E2 z JFH1
82 83 <210> 6 <211> 17 <212> DNA <213> Sekwencja sztuczna <220> <223> Opis sekwencji sztucznej: Starter (R6-130-S17) <400> 6 cgggagagcc atagtgg <210> 7 <211> 19 <212> DNA <213> Sekwencja sztuczna <220> <223> Opis sekwencji sztucznej: Starter (R6-290-R19) <400> 7 agtaccacaa ggcctttcg <210> 8 <211> 21 <212> DNA <213> Sekwencja sztuczna <220> <223> Opis sekwencji sztucznej: Sonda TaqMan (R6-148-S21FT) <400> 8 ctgcggaacc ggtgagtaca c <210> 9 <211> 9707 <212> RNA <213> Człowiek (Homo sapiens) <220>
83 <223> genomowy RNA o pełnej długości wirusa zapalenia wątroby typu C, pochodzący ze szczepu JFH1 (klon JFH-1 ) 84
84 85

85 86

86 87

87 88 <210> 10 <211> 3748 <212> DNA <213> Wirus zapalenia wątroby typu C <220> <223> genomowa sekwencja RNA zawierająca region od 5 UTR do NS3 ze szczepu TH1

88 89
89 90 <210> 11 <211> <212> RNA <213> Człowiek (Homo sapiens) <220> <223> chimerowy genomowy RNA wirusa zapalenia wątroby typu C pochodzący ze szczepu JFH1 HCV (klon JFH-1)

90 91

91 92

92 93

93 94

94 95 Zastrzeżenia patentowe 1. Zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C, zawierający części genomowego RNA z dwóch lub z większej ilości szczepów wirusów zapalenia wątroby typu C, który to zmodyfikowany genomowy RNA zawiera nie podlegający translacji region 5, sekwencję kodującą białko rdzeniowe, sekwencję kodującą białko E1, sekwencję kodującą białko E2, sekwencję kodującą białko p7, sekwencję kodującą białko NS2, częściową sekwencję RNA
95 96 kodującą białka NS3, NS4, NS5A i NS5B ze szczepu JFH1 przedstawioną jako SEQ ID Nr: 1, oraz nie podlegający translacji region 3, i który to RNA ma zdolność autonomicznej replikacji, przy czym jednym ze szczepów wirusa zapalenia wątroby typu C jest szczep wirusa o genotypie 1b. 2. Zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C, zawierający części genomowego RNA z dwóch lub z większej ilości szczepów wirusów zapalenia wątroby typu C, który to zmodyfikowany genomowy RNA zawiera nie podlegający translacji region 5, sekwencję kodującą białko rdzeniowe, sekwencję kodującą białko E1, sekwencję kodującą białko E2, sekwencję kodującą białko p7, sekwencję kodującą białko NS2, sekwencję kodującą białko NS3, sekwencję kodującą białko NS4A, sekwencję kodującą białko NS4B, sekwencję kodującą białko NS5A, sekwencję kodującą białko NS5B ze szczepu JFH1, przedstawioną jako SEQ ID Nr: 2, oraz nie podlegający translacji region-3, i który to RNA ma zdolność autonomicznej replikacji, przy czym jednym ze szczepów wirusa zapalenia wątroby typu C jest szczep wirusa o genotypie 1b. 3. Zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według zastrz. 1 albo zastrz. 2, w którym szczep wirusowy o genotypie 1b wybiera się spośród szczepu HCV-con1, szczepu HCV-TH, szczepu HCV-J, szczepu HCV-JT i szczepu HCV-BK. 4. Zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według zastrz. 1 albo zastrz. 2, w
96 97 którym jeden ze szczepów wirusa zapalenia wątroby typu C jest szczepem HCV-con1 o genotypie 1b. 5. Zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według zastrz. 4, w którym sekwencja kodująca białko E1, białko E2, białko p7 i białko NS2 ze szczepu HCV con1 jest zligowana w dół od nie podlegającego translacji regionu 5 ze szczepu JFH1 i w dół od niego, znajduje się sekwencja kodująca białka NS3, NS4A, NS4B, NS5A i NS5B ze szczepu JFH1 i w dół od niej, znajduje się nie podlegający translacji region 3 ze szczepu JFH1. 6. Zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według zastrz. 1 albo zastrz. 2, w którym jednym ze szczepów wirusa zapalenia wątroby typu C jest szczep HCV-TH o genotypie 1b. 7. Zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według zastrz. 6, zawierający sekwencję nukleotydową przedstawioną jako SEQ ID Nr: Zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C, zawierający części genomowego RNA z dwóch szczepów wirusów zapalenia wątroby typu C, w którym to RNA, tymi szczepami wirusów zapalenia wątroby typu C są szczep JFH1 i szczep JCH1 i zmodyfikowany genomowy RNA zapalenia wątroby typu C zawiera sekwencję odpowiadającą pozycjom od 1 do 3866 sekwencji zdeponowanej w bibliotece GenBank pod numerem dostępu AB zligowanej z
97 98 sekwencją odpowiadającą pozycjom od 3867 do 9678 sekwencji zdeponowanej w bibliotece GenBank pod numerem dostępu AB047639, i ma zdolność autonomicznej replikacji. 9. Ukierunkowany na komórki wątroby wektor wirusowy, który to wektor zawiera zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według zastrz. 1 do zastrz Komórka, do której został wprowadzony zmodyfikowany genomowy RNA wirusa zapalenia wątroby typu C według zastrz. 1 do zastrz. 8, w której przebiega replikacja genomowego RNA wirusa zapalenia wątroby typu C i która ma zdolność generowania cząsteczek wirusa. 11. Cząsteczki wirusa zapalenia wątroby typu C, które otrzymano z hodowli uzyskanej przez hodowlę komórki według zastrz Sposób oczyszczania cząsteczek HCV przez poddanie cieczy lub produktu otrzymanego z homogenatu komórek, zawierających cząsteczki HCV według zastrz. 11, chromatografii kolumnowej i/lub użytemu w połączeniu z nią wirowaniu w gradiencie gęstości. 13. Sposób według zastrz. 12, w którym stosuje się chromatografię kolumnową będącą jednym lub większą ilością typów chromatografii wybranych spośród chromatografii jonowymiennej, chromatografii żelowej i chromatografii powinowactwa.
98 Sposób według zastrz. 13, w którym jako chromatografię jonowymienną stosuje się jeden lub większą ilość typów chromatografii wybranych spośród chromatografii anionowymiennej i kationowymiennej a jako chromatografię żelową stosuje się jeden lub większą ilość typów chromatografii z użyciem żywicy wybranej spośród żywic Sephacryl-S300, Sephacryl S400 i Sephacryl S500 a jako chromatografię powinowactwa stosuje się jeden lub większą ilość typów chromatografii z użyciem żywicy wybranej spośród siarczanu celulofiny, heparyny i lektyny. 15. Sposób według zastrz. 13, w którym jako chromatografię stosuje się chromatografię z użyciem siarczanu celulofiny. 16. Sposób według zastrz. 12, w którym wirowanie w gradiencie gęstości prowadzi się stosując jedną lub większą ilość substancji rozpuszczonych wybranych spośród chlorku cezu, sacharozy i polimerów cukrowych. 17. Sposób według zastrz. 12, w którym w sposobie oczyszczania prowadzi się odpowiednio chromatografię anionowymienną, chromatografię na siarczanie celulofiny i wirowanie w gradiencie gęstości przynajmniej raz, w dowolnym połączeniu. 18. Cząsteczki HCV, które otrzymuje się sposobem oczyszczania cząsteczek HCV, w którym ciecz lub roztwór uzyskany z homogenatu komórek, zawierający cząsteczki HCV według zastrz. 11, poddaje się
99 100 chromatografii kolumnowej i wirowaniu w gradiencie gęstości, stosowanych w połączeniu. 19. Cząsteczki HCV według zastrz. 18, oczyszczone sposobem, w którym jako chromatografię kolumnową stosuje się jeden lub większą ilość typów chromatografii wybranych spośród chromatografii jonowymiennej, chromatografii żelowej i chromatografii powinowactwa. 20. Cząsteczki HCV według zastrz. 18, oczyszczone sposobem, w którym jako chromatografię jonowymienną stosuje się jeden lub większą ilość typów chromatografii wybranych spośród chromatografii anionowymiennej i chromatografii kationowymiennej a jako chromatografię żelową stosuje się jeden lub większą ilość typów chromatografii z użyciem żywicy wybranej spośród żywic Sephacryl-S300, Sephacryl-S400 i Sephacryl-S500 i jako chromatografię powinowactwa stosuje się jeden lub większą ilość typów chromatografii z użyciem żywicy wybranej spośród siarczanu cellulofiny, heparyny i lektyny. 21. Cząsteczki HCV według zastrz. 18, oczyszczone sposobem, w którym jako chromatografię stosuje się chromatografię na siarczanie celulofiny. 22. Cząsteczki HCV według zastrz. 18, oczyszczone sposobem, w którym wirowanie w gradiencie gęstości prowadzi się z użyciem jednej lub większej ilości substancji rozpuszczonych, wybranych spośród chlorku cezu, sacharozy i polimerów cukrowych.
100 Cząsteczki HCV według zastrz. 18, które oczyszcza się sposobem oczyszczania, w którym prowadzi się w połączeniu, chromatografię anionowymienną, chromatografię na siarczanie celulofiny i wirowanie w gradiencie gęstości sacharozy. 24. Zastosowanie cząsteczek wirusa zapalenia wątroby typu C według zastrz. 11 albo zastrz. 18 jako antygenu do produkcji szczepionki i/lub neutralizującego przeciwciała przeciw zapaleniu wątroby typu C. 25. Komórka zakażona wirusem zapalenia wątroby typu C, która została zakażona cząsteczkami wirusa zapalenia wątroby typu C według zastrz. 11 albo zastrz Sposób wytwarzania cząsteczek wirusa zapalenia wątroby typu C, znamienny tym, że sposób ten obejmuje hodowanie komórki według zastrz. 10 i odzyskanie cząsteczek wirusa z hodowli. 27. Sposób wytwarzania komórki zakażonej wirusem zapalenia wątroby typu C, znamienny tym, że sposób ten obejmuje hodowanie komórki według zastrz. 10, odzyskanie supernatantu hodowli komórek i użycia tego supernatantu wobec innej komórki w celu zakażenia tej innej komórki cząsteczkami wirusa zawartymi w tym supernatancie hodowli. 28. Sposób prowadzenia prób przesiewowych na substancję przeciw wirusowi zapalenia wątroby typu C, znamienny tym, że sposób ten obejmuje hodowanie
101 102 komórek według zastrz. 10 albo zastrz. 25 w obecności badanej substancji i wykrywanie RNA wirusa zapalenia wątroby typu C albo cząsteczek wirusa w tej hodowli, oceniając działanie tej badanej substancji przeciw wirusowi zapalenia wątroby typu C. 29. Sposób wytwarzania szczepionki i/lub neutralizującego przeciwciała przeciw zapaleniu wątroby typu C przez użycie cząsteczek wirusa zapalenia wątroby typu C według zastrz. 11 albo zastrz. 18 jako antygenu. 30. Sposób replikacji i/lub wywoływania ekspresji obcego genu w komórce, znamienny tym, że sposób ten obejmuje insercję RNA kodującego ten obcy gen do zmodyfikowanego genomowego RNA wirusa zapalenia wątroby typu C według dowolnego z zastrz. od 1 do 8 i wprowadzenie genomowego RNA do komórki będącej przedmiotem zainteresowania, tak by wywołać w niej replikację lub ekspresję tego obcego genu. Tokyo Metropolita Organization for Medical Research TORAY INDUSTRIES, INC.
102 103
103 104
104 105
105 106
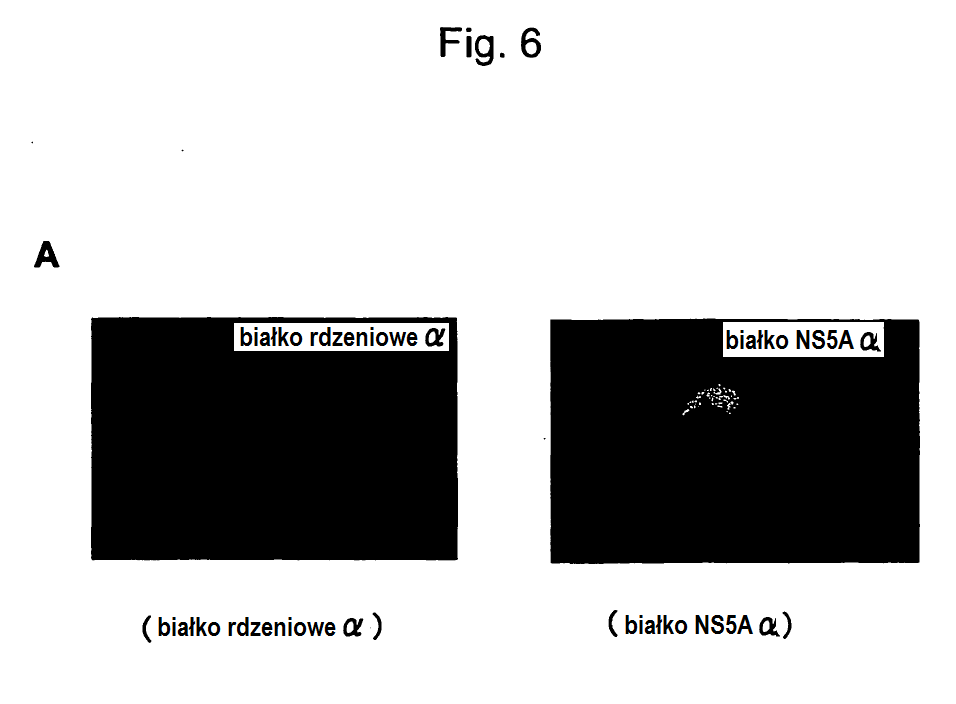
106 107
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE. Ewa Waszkowska ekspert UPRP
 WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1968711 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 05.01.2007 07712641.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1968711 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 05.01.2007 07712641.5
TECHNIKI ANALIZY RNA TECHNIKI ANALIZY RNA TECHNIKI ANALIZY RNA
 DNA 28SRNA 18/16S RNA 5SRNA mrna Ilościowa analiza mrna aktywność genów w zależności od wybranych czynników: o rodzaju tkanki o rodzaju czynnika zewnętrznego o rodzaju upośledzenia szlaku metabolicznego
DNA 28SRNA 18/16S RNA 5SRNA mrna Ilościowa analiza mrna aktywność genów w zależności od wybranych czynników: o rodzaju tkanki o rodzaju czynnika zewnętrznego o rodzaju upośledzenia szlaku metabolicznego
Klonowanie molekularne Kurs doskonalący. Zakład Geriatrii i Gerontologii CMKP
 Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680075 (13) T3 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2004
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680075 (13) T3 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2004
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1799953 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.08.2005 05770398.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1799953 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.08.2005 05770398.5
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609. (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609. (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609 (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.2 (13) (51) T3 Int.Cl. A61K 38/17 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609 (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.2 (13) (51) T3 Int.Cl. A61K 38/17 (2006.01)
Ćwiczenia 1 Wirtualne Klonowanie Prowadzący: mgr inż. Joanna Tymeck-Mulik i mgr Lidia Gaffke. Część teoretyczna:
 Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
Nowoczesne systemy ekspresji genów
 Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Hybrydyzacja kwasów nukleinowych
 Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 240040 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 09.07. 007077.0 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 240040 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 09.07. 007077.0 (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1747298 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.7 (51) Int. Cl. C22C14/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1747298 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.7 (51) Int. Cl. C22C14/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1730054 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 22.03.2005 05731932.9 (51) Int. Cl. B65G17/06 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1730054 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 22.03.2005 05731932.9 (51) Int. Cl. B65G17/06 (2006.01)
KLONOWANIE DNA REKOMBINACJA DNA WEKTORY
 KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2555663 (96) Data i numer zgłoszenia patentu europejskiego: 06.04.2011 11730434.5 (13) (51) T3 Int.Cl. A47L 15/42 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2555663 (96) Data i numer zgłoszenia patentu europejskiego: 06.04.2011 11730434.5 (13) (51) T3 Int.Cl. A47L 15/42 (2006.01)
Zawartość. Wstęp 1. Historia wirusologii. 2. Klasyfikacja wirusów
 Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2127498 (96) Data i numer zgłoszenia patentu europejskiego: 14.02.2008 08716843.1 (13) (51) T3 Int.Cl. H05B 41/288 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2127498 (96) Data i numer zgłoszenia patentu europejskiego: 14.02.2008 08716843.1 (13) (51) T3 Int.Cl. H05B 41/288 (2006.01)
Biologia Molekularna Podstawy
 Biologia Molekularna Podstawy Budowa DNA Budowa DNA Zasady: Purynowe: adenina i guanina Pirymidynowe: cytozyna i tymina 2 -deoksyryboza Grupy fosforanowe Budowa RNA Budowa RNA Zasady: purynowe: adenina
Biologia Molekularna Podstawy Budowa DNA Budowa DNA Zasady: Purynowe: adenina i guanina Pirymidynowe: cytozyna i tymina 2 -deoksyryboza Grupy fosforanowe Budowa RNA Budowa RNA Zasady: purynowe: adenina
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1740398 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 1.03.200 071703.9 (1) Int. Cl. B60C1/06 (2006.01) (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1740398 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 1.03.200 071703.9 (1) Int. Cl. B60C1/06 (2006.01) (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886669 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 02.08.2007 07113670.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886669 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 02.08.2007 07113670.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2480370 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 08.09.2010 10773557.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2480370 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 08.09.2010 10773557.3
Transformacja pośrednia składa się z trzech etapów:
 Transformacja pośrednia składa się z trzech etapów: 1. Otrzymanie pożądanego odcinka DNA z materiału genetycznego dawcy 2. Wprowadzenie obcego DNA do wektora 3. Wprowadzenie wektora, niosącego w sobie
Transformacja pośrednia składa się z trzech etapów: 1. Otrzymanie pożądanego odcinka DNA z materiału genetycznego dawcy 2. Wprowadzenie obcego DNA do wektora 3. Wprowadzenie wektora, niosącego w sobie
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 213136 (96) Data i numer zgłoszenia patentu europejskiego: 14.03.2008 08723469.6 (13) (1) T3 Int.Cl. F24D 19/ (2006.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 213136 (96) Data i numer zgłoszenia patentu europejskiego: 14.03.2008 08723469.6 (13) (1) T3 Int.Cl. F24D 19/ (2006.01) Urząd
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1890471 (96) Data i numer zgłoszenia patentu europejskiego: 19.10.2006 06791271.7 (13) (51) T3 Int.Cl. H04M 3/42 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1890471 (96) Data i numer zgłoszenia patentu europejskiego: 19.10.2006 06791271.7 (13) (51) T3 Int.Cl. H04M 3/42 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 223771 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.08 0886773.1 (13) (1) T3 Int.Cl. A47L 1/42 (06.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 223771 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.08 0886773.1 (13) (1) T3 Int.Cl. A47L 1/42 (06.01) Urząd
Dr. habil. Anna Salek International Bio-Consulting 1 Germany
 1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1787644 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 07.11.2006 06123574.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1787644 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 07.11.2006 06123574.3
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 28647 (96) Data i numer zgłoszenia patentu europejskiego: 30.03.09 091662.2 (13) (1) T3 Int.Cl. C07D 333/28 (06.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 28647 (96) Data i numer zgłoszenia patentu europejskiego: 30.03.09 091662.2 (13) (1) T3 Int.Cl. C07D 333/28 (06.01) Urząd
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych
 Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii
 Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
Hybrydyzacja kwasów nukleinowych
 Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja tego genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja tego genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 197092 (96) Data i numer zgłoszenia patentu europejskiego: 22.11.06 06824279.1 (13) (1) T3 Int.Cl. A61K 3/36 (06.01) A61P
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 197092 (96) Data i numer zgłoszenia patentu europejskiego: 22.11.06 06824279.1 (13) (1) T3 Int.Cl. A61K 3/36 (06.01) A61P
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii
 Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2003466 (96) Data i numer zgłoszenia patentu europejskiego: 12.06.2008 08460024.6 (13) (51) T3 Int.Cl. G01S 5/02 (2010.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2003466 (96) Data i numer zgłoszenia patentu europejskiego: 12.06.2008 08460024.6 (13) (51) T3 Int.Cl. G01S 5/02 (2010.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1859720. (96) Data i numer zgłoszenia patentu europejskiego: 15.02.2007 07003173.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1859720. (96) Data i numer zgłoszenia patentu europejskiego: 15.02.2007 07003173.") PL/EP 1859720 T3 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1859720 (96) Data i numer zgłoszenia patentu europejskiego: 15.02.2007 07003173.7 (13) (51) T3 Int.Cl. A47L
PL/EP 1859720 T3 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1859720 (96) Data i numer zgłoszenia patentu europejskiego: 15.02.2007 07003173.7 (13) (51) T3 Int.Cl. A47L
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1732433 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.01.2005 05702820.1
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1732433 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.01.2005 05702820.1
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 182634 (96) Data i numer zgłoszenia patentu europejskiego: 19.04.07 070963.1 (13) T3 (1) Int. Cl. F16H/17 F16H7/04 (06.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 182634 (96) Data i numer zgłoszenia patentu europejskiego: 19.04.07 070963.1 (13) T3 (1) Int. Cl. F16H/17 F16H7/04 (06.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2190940 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.09.2008 08802024.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2190940 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.09.2008 08802024.3
Replikacja wirusa HCV w linii komórkowej Huh 7.5. HCV replication in Huh 7.5 cell line
 MED. DOŚW. MIKROBIOL., 2012, 64: 239-244 Replikacja wirusa HCV w linii komórkowej Huh 7.5 HCV replication in Huh 7.5 cell line Marcin Komorowski, Agnieszka Kołakowska, Paulina Godzik, Kazimierz Madaliński
MED. DOŚW. MIKROBIOL., 2012, 64: 239-244 Replikacja wirusa HCV w linii komórkowej Huh 7.5 HCV replication in Huh 7.5 cell line Marcin Komorowski, Agnieszka Kołakowska, Paulina Godzik, Kazimierz Madaliński
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680966 (96) Data i numer zgłoszenia patentu europejskiego: 27.10.2004 04791390.0 (13) T3 (51) Int. Cl. A23L1/172 A23P1/08
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680966 (96) Data i numer zgłoszenia patentu europejskiego: 27.10.2004 04791390.0 (13) T3 (51) Int. Cl. A23L1/172 A23P1/08
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 71811 (96) Data i numer zgłoszenia patentu europejskiego: 29.09.06 06791167.7 (13) (1) T3 Int.Cl. H04Q 11/00 (06.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 71811 (96) Data i numer zgłoszenia patentu europejskiego: 29.09.06 06791167.7 (13) (1) T3 Int.Cl. H04Q 11/00 (06.01) Urząd
(96) Data i numer zgłoszenia patentu europejskiego:
 Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1690978 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2005 05101042.9 (13) T3 (51) Int. Cl. D06F81/08 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1690978 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2005 05101042.9 (13) T3 (51) Int. Cl. D06F81/08 (2006.01)
(Akty o charakterze nieustawodawczym) DECYZJE
 DECYZJE") 19.7.2019 PL L 193/1 II (Akty o charakterze nieustawodawczym) DECYZJE DECYZJA WYKONAWCZA KOMISJI (UE) 2019/1244 z dnia 1 lipca 2019 r. zmieniająca decyzję 2002/364/WE w odniesieniu do wymogów dotyczących
19.7.2019 PL L 193/1 II (Akty o charakterze nieustawodawczym) DECYZJE DECYZJA WYKONAWCZA KOMISJI (UE) 2019/1244 z dnia 1 lipca 2019 r. zmieniająca decyzję 2002/364/WE w odniesieniu do wymogów dotyczących
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1624265 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2005 05106119.0 (13) T3 (51) Int. Cl. F25D23/06 F25D25/02
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1624265 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2005 05106119.0 (13) T3 (51) Int. Cl. F25D23/06 F25D25/02
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2259949 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2009 09727379.1 (13) (51) T3 Int.Cl. B60L 11/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2259949 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2009 09727379.1 (13) (51) T3 Int.Cl. B60L 11/00 (2006.01)
Pytania Egzamin magisterski
 Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2383703 (96) Data i numer zgłoszenia patentu europejskiego: 29.04. 40068.1 (13) (1) T3 Int.Cl. G07B 1/06 (11.01) G08G 1/017
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2383703 (96) Data i numer zgłoszenia patentu europejskiego: 29.04. 40068.1 (13) (1) T3 Int.Cl. G07B 1/06 (11.01) G08G 1/017
Biologia Molekularna z Biotechnologią ===============================================================================================
 Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1810954 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.2006 06025226.9 (13) (51) T3 Int.Cl. C03B 9/41 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1810954 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.2006 06025226.9 (13) (51) T3 Int.Cl. C03B 9/41 (2006.01)
października 2013: Elementarz biologii molekularnej. Wykład nr 2 BIOINFORMATYKA rok II
 10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2052830. (96) Data i numer zgłoszenia patentu europejskiego: 21.10.2008 08018365.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2052830. (96) Data i numer zgłoszenia patentu europejskiego: 21.10.2008 08018365.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 202830 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21..2008 0801836.0 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 202830 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21..2008 0801836.0 (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1802536 (96) Data i numer zgłoszenia patentu europejskiego: 20.09.2004 04774954.4 (13) T3 (51) Int. Cl. B65D77/20 B65D85/72
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1802536 (96) Data i numer zgłoszenia patentu europejskiego: 20.09.2004 04774954.4 (13) T3 (51) Int. Cl. B65D77/20 B65D85/72
Agenda: Wynalazek jako własnośd intelektualna. Co może byd wynalazkiem. Wynalazek biotechnologiczny. Ochrona patentowa
 dr Bartosz Walter Agenda: Wynalazek jako własnośd intelektualna Co może byd wynalazkiem Wynalazek biotechnologiczny Ochrona patentowa Procedura zgłoszenia patentowego Gdzie, kiedy i jak warto patentowad
dr Bartosz Walter Agenda: Wynalazek jako własnośd intelektualna Co może byd wynalazkiem Wynalazek biotechnologiczny Ochrona patentowa Procedura zgłoszenia patentowego Gdzie, kiedy i jak warto patentowad
Ćwiczenia 1 Wirtualne Klonowanie. Prowadzący: mgr Anna Pawlik i mgr Maciej Dylewski. Część teoretyczna:
 Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią ===============================================================================================
Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią ===============================================================================================
Patentowanie wynalazków z dziedziny biotechnologii
 chroń swoje pomysły 3 Kongres Świata Przemysłu Farmaceutycznego Poznań, 16-17 czerwca 2011 Patentowanie wynalazków z dziedziny biotechnologii dr Izabela Milczarek Czym jest biotechnologia? Biotechnologia,
chroń swoje pomysły 3 Kongres Świata Przemysłu Farmaceutycznego Poznań, 16-17 czerwca 2011 Patentowanie wynalazków z dziedziny biotechnologii dr Izabela Milczarek Czym jest biotechnologia? Biotechnologia,
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 161679 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 24.06.0 064.7 (1) Int. Cl. B60R21/01 (06.01) (97) O udzieleniu
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 161679 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 24.06.0 064.7 (1) Int. Cl. B60R21/01 (06.01) (97) O udzieleniu
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii
 Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych aspekty praktyczne dr Małgorzata Kozłowska Ekspert
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych aspekty praktyczne dr Małgorzata Kozłowska Ekspert
Zestawy do izolacji DNA i RNA
 Syngen kolumienki.pl Gotowe zestawy do izolacji i oczyszczania kwasów nukleinowych Zestawy do izolacji DNA i RNA Katalog produktów 2012-13 kolumienki.pl Syngen Biotech Sp. z o.o., 54-116 Wrocław, ul. Ostródzka
Syngen kolumienki.pl Gotowe zestawy do izolacji i oczyszczania kwasów nukleinowych Zestawy do izolacji DNA i RNA Katalog produktów 2012-13 kolumienki.pl Syngen Biotech Sp. z o.o., 54-116 Wrocław, ul. Ostródzka
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1505553. (96) Data i numer zgłoszenia patentu europejskiego: 05.08.2004 04018511.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1505553. (96) Data i numer zgłoszenia patentu europejskiego: 05.08.2004 04018511.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 3 (96) Data i numer zgłoszenia patentu europejskiego: 0.08.04 0401811.8 (13) (1) T3 Int.Cl. G08C 17/00 (06.01) Urząd Patentowy
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 3 (96) Data i numer zgłoszenia patentu europejskiego: 0.08.04 0401811.8 (13) (1) T3 Int.Cl. G08C 17/00 (06.01) Urząd Patentowy
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 172874 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 04.0.2006 0611312. (1) Int. Cl. B23B31/28 (2006.01) (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 172874 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 04.0.2006 0611312. (1) Int. Cl. B23B31/28 (2006.01) (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2215272. (96) Data i numer zgłoszenia patentu europejskiego: 31.10.2008 08844624.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2215272. (96) Data i numer zgłoszenia patentu europejskiego: 31.10.2008 08844624.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 221272 (96) Data i numer zgłoszenia patentu europejskiego: 31.10.2008 08844624.0 (13) (1) T3 Int.Cl. C12Q 1/70 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 221272 (96) Data i numer zgłoszenia patentu europejskiego: 31.10.2008 08844624.0 (13) (1) T3 Int.Cl. C12Q 1/70 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2321656 (96) Data i numer zgłoszenia patentu europejskiego:.08.09 09807498.2 (13) (51) T3 Int.Cl. G01R /18 (06.01) G01R 19/
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2321656 (96) Data i numer zgłoszenia patentu europejskiego:.08.09 09807498.2 (13) (51) T3 Int.Cl. G01R /18 (06.01) G01R 19/
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1614553 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 01.07.2005 05014326.2 (51) Int. Cl. B60C27/06 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1614553 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 01.07.2005 05014326.2 (51) Int. Cl. B60C27/06 (2006.01)
Inżynieria genetyczna- 6 ECTS. Inżynieria genetyczna. Podstawowe pojęcia Część II Klonowanie ekspresyjne Od genu do białka
 Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Techniki molekularne w mikrobiologii SYLABUS A. Informacje ogólne
 Techniki molekularne w mikrobiologii A. Informacje ogólne Elementy sylabusu Nazwa jednostki prowadzącej kierunek Nazwa kierunku studiów Poziom kształcenia Profil studiów Forma studiów Rodzaj Rok studiów
Techniki molekularne w mikrobiologii A. Informacje ogólne Elementy sylabusu Nazwa jednostki prowadzącej kierunek Nazwa kierunku studiów Poziom kształcenia Profil studiów Forma studiów Rodzaj Rok studiów
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2057877 (96) Data i numer zgłoszenia patentu europejskiego: 03.11.2008 08019246.1 (13) (51) T3 Int.Cl. A01C 23/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2057877 (96) Data i numer zgłoszenia patentu europejskiego: 03.11.2008 08019246.1 (13) (51) T3 Int.Cl. A01C 23/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2074843. (96) Data i numer zgłoszenia patentu europejskiego: 27.09.2007 07818485.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2074843. (96) Data i numer zgłoszenia patentu europejskiego: 27.09.2007 07818485.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 74843 (96) Data i numer zgłoszenia patentu europejskiego: 27.09.07 0781848.0 (13) (1) T3 Int.Cl. H04W 4/12 (09.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 74843 (96) Data i numer zgłoszenia patentu europejskiego: 27.09.07 0781848.0 (13) (1) T3 Int.Cl. H04W 4/12 (09.01) Urząd
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 09.12.2005 05077837.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 09.12.2005 05077837.") RZECZPSPLITA PLSKA (12) TŁUMACZENIE PATENTU EURPEJSKIEG (19) PL (11) PL/EP 1671547 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 09.12.2005 05077837.2 (51) Int. Cl. A23B7/154 (2006.01) (97)
RZECZPSPLITA PLSKA (12) TŁUMACZENIE PATENTU EURPEJSKIEG (19) PL (11) PL/EP 1671547 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 09.12.2005 05077837.2 (51) Int. Cl. A23B7/154 (2006.01) (97)
mikrosatelitarne, minisatelitarne i polimorfizm liczby kopii
 Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2047071 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.07.2007 07786251.4
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2047071 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.07.2007 07786251.4
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1671552 (96) Data i numer zgłoszenia patentu europejskiego: 02.12.2005 05026319.3 (13) T3 (51) Int. Cl. A23L1/305 A23J3/16
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1671552 (96) Data i numer zgłoszenia patentu europejskiego: 02.12.2005 05026319.3 (13) T3 (51) Int. Cl. A23L1/305 A23J3/16
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2179743 (96) Data i numer zgłoszenia patentu europejskiego: 13.07.2009 09460028.5 (13) (51) T3 Int.Cl. A61K 38/18 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2179743 (96) Data i numer zgłoszenia patentu europejskiego: 13.07.2009 09460028.5 (13) (51) T3 Int.Cl. A61K 38/18 (2006.01)
Making the impossible possible: the metamorphosis of Polish Biology Olympiad
 Making the impossible possible: the metamorphosis of Polish Biology Olympiad Takao Ishikawa Faculty of Biology, University of Warsaw, Poland Performance of Polish students at IBO Gold Silver Bronze Merit
Making the impossible possible: the metamorphosis of Polish Biology Olympiad Takao Ishikawa Faculty of Biology, University of Warsaw, Poland Performance of Polish students at IBO Gold Silver Bronze Merit
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 187318 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 06.04.06 06731279.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 187318 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 06.04.06 06731279.3
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2210706 (96) Data i numer zgłoszenia patentu europejskiego: 21.01.2010 10000580.0 (13) (51) T3 Int.Cl. B24B 21/20 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2210706 (96) Data i numer zgłoszenia patentu europejskiego: 21.01.2010 10000580.0 (13) (51) T3 Int.Cl. B24B 21/20 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1854925 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2005 05826699.0 (13) (51) T3 Int.Cl. E03D 1/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1854925 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2005 05826699.0 (13) (51) T3 Int.Cl. E03D 1/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1510645 (96) Data i numer zgłoszenia patentu europejskiego: 20.08.2004 04019758.4 (13) (51) T3 Int.Cl. E06B 3/58 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1510645 (96) Data i numer zgłoszenia patentu europejskiego: 20.08.2004 04019758.4 (13) (51) T3 Int.Cl. E06B 3/58 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1658064 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.6 (51) Int. Cl. A61K31/37 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1658064 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.6 (51) Int. Cl. A61K31/37 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2131847. (96) Data i numer zgłoszenia patentu europejskiego: 27.02.2008 08716068.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2131847. (96) Data i numer zgłoszenia patentu europejskiego: 27.02.2008 08716068.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2131847 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.02.2008 08716068.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2131847 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.02.2008 08716068.5
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 221611 (96) Data i numer zgłoszenia patentu europejskiego: 19.01. 000481.1 (13) (1) T3 Int.Cl. B28C /42 (06.01) B60P 3/16
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 221611 (96) Data i numer zgłoszenia patentu europejskiego: 19.01. 000481.1 (13) (1) T3 Int.Cl. B28C /42 (06.01) B60P 3/16
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2086467 (96) Data i numer zgłoszenia patentu europejskiego: 26.11.2007 07824706.1 (13) (51) T3 Int.Cl. A61F 2/16 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2086467 (96) Data i numer zgłoszenia patentu europejskiego: 26.11.2007 07824706.1 (13) (51) T3 Int.Cl. A61F 2/16 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1701111 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 11.03.2005 05090064.6 (51) Int. Cl. F24H9/20 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1701111 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 11.03.2005 05090064.6 (51) Int. Cl. F24H9/20 (2006.01)
ROZPORZĄDZENIE KOMISJI (UE) / z dnia r.
 / z dnia r.") KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2328822 (96) Data i numer zgłoszenia patentu europejskiego: 02.09.2009 09782487.4 (13) (51) T3 Int.Cl. B65G 15/38 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2328822 (96) Data i numer zgłoszenia patentu europejskiego: 02.09.2009 09782487.4 (13) (51) T3 Int.Cl. B65G 15/38 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1477128 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 14.05.2004 04076445.8 (51) Int. Cl. A61D1/02 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1477128 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 14.05.2004 04076445.8 (51) Int. Cl. A61D1/02 (2006.01)
Oznaczenie polimorfizmu genetycznego cytochromu CYP2D6: wykrywanie liczby kopii genu
 Ćwiczenie 4 Oznaczenie polimorfizmu genetycznego cytochromu CYP2D6: wykrywanie liczby kopii genu Wstęp CYP2D6 kodowany przez gen występujący w co najmniej w 78 allelicznych formach związanych ze zmniejszoną
Ćwiczenie 4 Oznaczenie polimorfizmu genetycznego cytochromu CYP2D6: wykrywanie liczby kopii genu Wstęp CYP2D6 kodowany przez gen występujący w co najmniej w 78 allelicznych formach związanych ze zmniejszoną
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445326 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.10.2011 11186353.6
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445326 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.10.2011 11186353.6
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2326237 (96) Data i numer zgłoszenia patentu europejskiego: 07.07.2009 09780285.4 (13) (51) T3 Int.Cl. A47L 15/50 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2326237 (96) Data i numer zgłoszenia patentu europejskiego: 07.07.2009 09780285.4 (13) (51) T3 Int.Cl. A47L 15/50 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego: 06.09.2005 05788867.9
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego: 06.09.2005 05788867.9") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1786660 (96) Data i numer zgłoszenia patentu europejskiego: 06.09.2005 05788867.9 (13) T3 (51) Int. Cl. B62D25/08 B60G15/06
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1786660 (96) Data i numer zgłoszenia patentu europejskiego: 06.09.2005 05788867.9 (13) T3 (51) Int. Cl. B62D25/08 B60G15/06
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 4 Jak działają geny?
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 4 Jak działają geny?
Rok akademicki: 2014/2015 Kod: EIB-2-206-BN-s Punkty ECTS: 3. Kierunek: Inżynieria Biomedyczna Specjalność: Bionanotechnologie
 Nazwa modułu: Genetyka molekularna Rok akademicki: 2014/2015 Kod: EIB-2-206-BN-s Punkty ECTS: 3 Wydział: Elektrotechniki, Automatyki, Informatyki i Inżynierii Biomedycznej Kierunek: Inżynieria Biomedyczna
Nazwa modułu: Genetyka molekularna Rok akademicki: 2014/2015 Kod: EIB-2-206-BN-s Punkty ECTS: 3 Wydział: Elektrotechniki, Automatyki, Informatyki i Inżynierii Biomedycznej Kierunek: Inżynieria Biomedyczna
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2300459. (96) Data i numer zgłoszenia patentu europejskiego: 22.06.2009 09772333.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2300459. (96) Data i numer zgłoszenia patentu europejskiego: 22.06.2009 09772333.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2049 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.06.09 09772333.2 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2049 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.06.09 09772333.2 (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886585 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.07.2006 06291197.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886585 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.07.2006 06291197.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2764103 (96) Data i numer zgłoszenia patentu europejskiego: 12.12.2013 13824232.6 (13) (51) T3 Int.Cl. C12N 15/63 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2764103 (96) Data i numer zgłoszenia patentu europejskiego: 12.12.2013 13824232.6 (13) (51) T3 Int.Cl. C12N 15/63 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2084461 (96) Data i numer zgłoszenia patentu europejskiego: 27.11.2007 07847411.1 (13) (51) T3 Int.Cl. F24C 3/10 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2084461 (96) Data i numer zgłoszenia patentu europejskiego: 27.11.2007 07847411.1 (13) (51) T3 Int.Cl. F24C 3/10 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") PL/EP 1699990 T3 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1699990 (96) Data i numer zgłoszenia patentu europejskiego: 09.11.2004 04800186.1 (13) (51) T3 Int.Cl. E04G
PL/EP 1699990 T3 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1699990 (96) Data i numer zgłoszenia patentu europejskiego: 09.11.2004 04800186.1 (13) (51) T3 Int.Cl. E04G
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2584058. (96) Data i numer zgłoszenia patentu europejskiego: 21.10.2011 11186244.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2584058. (96) Data i numer zgłoszenia patentu europejskiego: 21.10.2011 11186244.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2584058 (96) Data i numer zgłoszenia patentu europejskiego: 21.10.2011 11186244.7 (13) (51) T3 Int.Cl. C22C 38/40 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2584058 (96) Data i numer zgłoszenia patentu europejskiego: 21.10.2011 11186244.7 (13) (51) T3 Int.Cl. C22C 38/40 (2006.01)