(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
|
|
|
- Sabina Karczewska
- 6 lat temu
- Przeglądów:
Transkrypt
 C07K 14/60 (06.01) Urząd Patentowy Rzeczypospolitej Polskiej (97) O udzieleniu patentu europejskiego ogłoszono: 12.08.")
1 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego: (13) (1) T3 Int.Cl. A61K 38/00 (06.01) C07K 14/60 (06.01) Urząd Patentowy Rzeczypospolitej Polskiej (97) O udzieleniu patentu europejskiego ogłoszono: Europejski Biuletyn Patentowy 1/33 EP B1 (4) Tytuł wynalazku: Mieszani agoniści na bazie GIP do leczenia zaburzeń metabolicznych i otyłości (30) Pierwszeństwo: US P US P US P US P (43) Zgłoszenie ogłoszono: w Europejskim Biuletynie Patentowym nr 11/13 (4) O złożeniu tłumaczenia patentu ogłoszono: Wiadomości Urzędu Patentowego 16/04 (73) Uprawniony z patentu: Indiana University Research and Technology Corporation, Indianapolis, US (72) Twórca(y) wynalazku: PL/EP T3 RICHARD D. DIMARCHI, Carmel, US TAO MA, Beijing, CN (74) Pełnomocnik: rzecz. pat. Marta Kawczyńska POLSERVICE KANCELARIA RZECZNIKÓW PATENTOWYCH SP. Z O.O. ul. Bluszczańska Warszawa Uwaga: W ciągu dziewięciu miesięcy od publikacji informacji o udzieleniu patentu europejskiego, każda osoba może wnieść do Europejskiego Urzędu Patentowego sprzeciw dotyczący udzielonego patentu europejskiego. Sprzeciw wnosi się w formie uzasadnionego na piśmie oświadczenia. Uważa się go za wniesiony dopiero z chwilą wniesienia opłaty za sprzeciw (Art. 99 (1) Konwencji o udzielaniu patentów europejskich).
2 P37239PL00 EP B1 Opis ODNIESIENIE DO ZGŁOSZEŃ POWIĄZANYCH [0001] Niniejsze zgłoszenie zastrzega pierwszeństwo z następujących zgłoszeń: tymczasowe zgłoszenie patentowe USA nr 61/073,274 dokonane 17 czerwca 08 r., tymczasowe zgłoszenie patentowe USA nr 61/078,171 dokonane 3 lipca 08 r., tymczasowe zgłoszenie patentowe USA nr 61/090,448 dokonane sierpnia 08 r. i tymczasowe zgłoszenie patentowe USA nr 61/11,349 dokonane lutego 09 r. INFORMACJE WSTĘPNE [0002] Preproglukagon jest polipeptydem prekursorowym o długości 18 aminokwasów ulegającym obróbce w różnych tkankach z utworzeniem szeregu różnych peptydów stanowiących pochodne proglukagonu, w tym glukagonu, peptydu glukagonopodobnego 1 (GLP-1), peptydu glukagonopodobnego 2 (GLP-2) i oksyntomoduliny (OXM), biorących udział w różnorodnych funkcjach fizjologicznych, w tym w homeostazie glukozy, wydzielaniu insuliny, opróżnianiu żołądka i wzroście tkanek w jelicie, a także w regulacji przyjmowania pokarmu. Glukagon jest peptydem o długości 29 aminokwasów odpowiadającym aminokwasom 33 do 61 w preproglukagonie, a GLP-1 jest wytwarzany jako peptyd o długości 37 aminokwasów odpowiadający aminokwasom 72 do 8 w preproglukagonie. [0003] Kiedy stężenie glukozy we krwi zaczyna spadać, glukagon hormon wytwarzany w trzustce przekazuje do wątroby sygnał prowadzący do rozkładu glikogenu i uwalniania glukozy, w wyniku czego poziomy glukozy we
3 krwi wzrastają do poziomu prawidłowego. GLP-1 ma odmienne działania biologiczne w porównaniu do glukagonu. Jego działania obejmują stymulację syntezy i wydzielania insuliny, hamowanie wydzielania glukagonu i hamowanie przyjmowania pokarmu. Stwierdzono, że GLP-1 zmniejsza hiperglikemię (podwyższone poziomy glukozy) u diabetyków. Eksendyna 4, peptyd z jadu jaszczurki wykazujący około 0% identyczności aminokwasów względem GLP-1, powoduje aktywację receptora GLP-1 i również wykazano, że zmniejsza hiperglikemię u diabetyków. [0004] Glukozozależny peptyd insulinotropowy (GIP) jest peptydem regulacyjnym w przewodzie pokarmowym o długości 42 aminokwasów stymulującym wydzielanie insuliny w komórkach beta trzustki w obecności glukozy. Jest on wytwarzany w wyniku obróbki proteolitycznej preprogip, prekursora o długości 133 aminokwasów. PODSUMOWANIE [000] Zgodnie z ujawnieniem w niniejszym dokumencie dostarcza się peptydy glukagonu, będące analogami natywnego glukagonu (NR. ID. SEKW.: 1) i wykazujące aktywność GIP. Wynalazek ujawnia ponadto sposoby zastosowania tych peptydów. [0006] Natywny glukagon nie powoduje aktywacji receptora GIP i normalnie ma około 1% aktywności natywnego GLP-1 wobec receptora GLP-1. W wyniku modyfikacji sekwencji natywnego glukagonu według niniejszego dokumentu otrzymuje się peptydy glukagonu, które mogą wykazywać silne działanie glukagonu równoważne lub większe od aktywności natywnego glukagonu (NR. ID. SEKW.: 1), silne działanie GIP równoważne lub większe od aktywności natywnego GIP (NR.
4 ID. SEKW.: 4) i(lub) silne działanie GLP-1 równoważne lub większe od aktywności natywnego GLP-1. GLP-1(7-36) amid (NR. ID. SEKW.: 3) lub GLP-1(7-37) (kwas) (NR. ID. SEKW.: 2) są formami GLP-1 o silnym działaniu biologicznym mającymi zasadniczo równoważne działanie wobec receptora GLP-1. [0007] Dane podane w niniejszym dokumencie pokazują, że peptydy mające zarówno działanie GIP, jak i działanie GLP-1 są szczególnie korzystne do wywołania utraty masy lub zapobiegania wzrostowi masy, a także w leczeniu hiperglikemii, w tym cukrzycy. Dane in vivo ujawniane w niniejszym dokumencie pokazują, że połączenie działania agonistycznego wobec GIP z działaniem agonistycznym wobec GLP-1 powoduje większe oddziaływanie na zmniejszenie masy niż sam GLP-1. Aktywność ta jest szczególnie nieoczekiwana w świetle ujawnień ze stanu techniki, według których działanie antagonistyczne wobec GIP jest pożądane, aby zmniejszyć dobowe przyjmowanie pokarmu i masę ciała, a także zwiększyć insulinowrażliwość i wydatek energetyczny. (Irwin i wsp., Diabetologia 0: (07); oraz Althage i wsp., J Biol Chem, publikacja elektroniczna z 17 kwietnia 08 r.). [0008] W związku z tym w jednym aspekcie wynalazek dostarcza się sposoby indukowania utraty masy lub zapobiegania wzrostowi masy, które obejmują podawanie pacjentowi tego wymagającemu skutecznej ilości związku, np. peptydu glukagonu, który wykazuje działanie zarówno wobec receptora GIP, jak i receptora GLP-1, a także ewentualnie wykazuje działanie wobec receptora glukagonu. Do takich związków należą związki działające
5 koagonistycznie wobec GIP/GLP-1 (koagoniści GIP/GLP-1) i triagonistycznie (triagoniści) wobec glukagonu/gip/glp-1 opisywane w niniejszym dokumencie. [0009] Zwiększoną aktywność wobec receptora GIP uzyskuje się dzięki modyfikacji aminokwasu w położeniu 1. Na przykład His w położeniu 1 jest podstawiona dużym aminokwasem aromatycznym, ewentualnie Tyr, Phe, Trp, amino-phe, nitro-phe, chloro-phe, sulfo-phe, 4- pirydylo-ala, metylo-tyr lub 3-amino-Tyr. [00] Zwiększoną aktywność wobec receptora GIP uzyskuje się dzięki modyfikacjom stabilizującym strukturę helisy alfa części C-końcowej (aminokwasy 12-29) peptydu glukagonu lub jego analogu. Na przykład mostek wewnątrzcząsteczkowy może utworzyć się poprzez wiązanie kowalencyjne między łańcuchami bocznymi dwóch aminokwasów w położeniach i oraz i+4 lub między położeniami j i j+3 lub między położeniami k i k+7. W pewnych przykładach wykonania mostek znajduje się między położeniami 12 i 16, 16 i, i 24, 24 i 28 lub 17 i. W innych przykładach wykonania oddziaływania niekowalencyjne, na przykład mostki solne, mogą utworzyć się między aminokwasami naładowanymi dodatnio i ujemnie w tych położeniach. Ewentualnie na przykład stabilizację struktury helisy alfa części C-końcowej peptydu glukagonu (w obrębie aminokwasów 12-29) uzyskuje się dzięki celowemu wprowadzeniu jednego lub większej liczby α,αdipodstawionych aminokwasów w położeniach pozwalających zachować pożądaną aktywność. W niektórych przykładach wykonania jedno, dwa, trzy, cztery lub więcej spośród położeń 16, 17, 18, 19,, 21, 24 lub 29 w peptydzie
6 glukagonu lub jego analogu jest podstawionych aminokwasem α,α-dipodstawionym. Na przykład substytucja położenia 16 w peptydzie glukagonu lub jego analogu przez kwas aminoizomasłowy (AIB) zapewnia ustabilizowaną helisę alfa w nieobecności mostka solnego lub laktamu. Takie peptydy uznaje się w niniejszym dokumencie za peptyd niezawierający mostka wewnątrzcząsteczkowego. W konkretnych aspektach stabilizację helisy alfa uzyskuje się dzięki wprowadzeniu jednego lub większej liczby α,αdipodstawionych aminokwasów bez wprowadzania kowalencyjnego mostka wewnątrzcząsteczkowego, np. mostka laktamowego, mostka disiarczkowego. Takie peptydy uznaje się w niniejszym dokumencie za peptyd niezawierający kowalencyjnego mostka wewnątrzcząsteczkowego. W niektórych przykładach wykonania jedno, dwa, trzy lub więcej spośród położeń 16,, 21 lub 24 jest podstawionych przez AIB. [0011] Zwiększoną aktywność wobec receptora GIP uzyskuje się dzięki modyfikacji aminokwasów w położeniach 27 i(lub) 28 oraz ewentualnie w położeniu 29. Na przykład Met w położeniu 27 jest podstawiona przez duży aminokwas alifatyczny, ewentualnie Leu, Asn w położeniu 28 jest podstawiona przez mały aminokwas alifatyczny, ewentualnie Ala, a Thr w położeniu 29 jest podstawiona przez mały aminokwas alifatyczny, ewentualnie Gly. [0012] Zwiększoną aktywność wobec receptora GIP uzyskuje się także dzięki modyfikacji aminokwasu w położeniu 12. Na przykład położenie 12 jest podstawione przez duży alifatyczny aminokwas niepolarny,
7 ewentualnie Ile. [0013] Zwiększoną aktywność wobec receptora GIP uzyskuje się także dzięki modyfikacji aminokwasu w położeniach 17 i(lub) 18. Na przykład położenie 17 jest podstawione przez resztę polarną, ewentualnie Gln, a położenie 18 jest podstawione przez mały aminokwas alifatyczny, ewentualnie Ala. [0014] Zwiększoną aktywność wobec receptora glukagonu uzyskuje się dzięki modyfikacji aminokwasu w położeniu 16 natywnego glukagonu (NR. ID. SEKW.: 1) opisanego w niniejszym dokumencie. [001] Zmniejszoną, utrzymaną lub zwiększoną aktywność wobec receptora glukagonu uzyskuje się np. dzięki modyfikacji aminokwasu w położeniu 3 zgodnie z opisem w niniejszym dokumencie. [0016] Przywrócenie aktywności glukagonu, która została zmniejszona wskutek modyfikacji aminokwasów w położeniach 1 i(lub) 2, uzyskuje się dzięki modyfikacjom stabilizującym strukturę helisy alfa części C-końcowej (aminokwasy 12-29) peptydu glukagonu lub jego analogu. Na przykład mostek wewnątrzcząsteczkowy może utworzyć się poprzez wiązanie kowalencyjne między łańcuchami bocznymi dwóch aminokwasów w położeniach i oraz i+4 lub między położeniami j i j+3, lub między położeniami k i k+7. W innych przykładach wykonania oddziaływania niekowalencyjne, takie jak mostki solne, mogą utworzyć się między aminokwasami naładowanymi dodatnio i ujemnie w tych położeniach. W jeszcze innych przykładach wykonania do tej części C-końcowej (aminokwasy 12-29) w położeniach pozwalających zachować pożądaną aktywność
8 wprowadza się lub podstawia jednym lub większą liczbą α,α-dipodstawionych aminokwasów. Na przykład jedno, dwa, trzy lub wszystkie spośród położeń 16,, 21 lub 24 jest podstawionych przez aminokwas α,αdipodstawiony, np. AIB. [0017] Zwiększoną aktywność wobec receptora GLP-1 uzyskuje się dzięki zastąpieniu kwasu karboksylowego w aminokwasie C-końcowym grupą o ładunku obojętnym, taką jak amid lub ester. [0018] Zwiększoną aktywność wobec receptora GLP-1 uzyskuje się dzięki modyfikacjom stabilizującym strukturę helisy alfa części C-końcowej (w obrębie aminokwasów 12-29) peptydu glukagonu lub jego analogu. W niektórych przykładach wykonania mostek wewnątrzcząsteczkowy może utworzyć się poprzez wiązanie kowalencyjne między łańcuchami bocznymi dwóch aminokwasów w położeniach i oraz i+4 lub między położeniami j i j+3, lub między położeniami k i k+7. W innych przykładach wykonania oddziaływania niekowalencyjne, takie jak mostki solne, mogą utworzyć się między aminokwasami naładowanymi dodatnio i ujemnie w tych położeniach. W jeszcze innych przykładach wykonania do tej części C-końcowej (aminokwasy 12-29) w położeniach pozwalających zachować pożądaną aktywność wprowadza się lub podstawia jednym lub większą liczbą α,α-dipodstawionych aminokwasów. Na przykład jedno, dwa, trzy lub wszystkie spośród położeń 16,, 21 lub 24 jest podstawionych przez aminokwas α,αdipodstawiony, np. AIB. [0019] Zwiększoną aktywność wobec receptora GLP-1 uzyskuje się dzięki modyfikacji aminokwasu w położeniu
9 zgodnie z opisem w niniejszym dokumencie. [00] Zwiększoną aktywność wobec receptora GLP-1 uzyskuje się dzięki dodaniu peptydu stanowiącego przedłużenie C-końcowe, na przykład GPSSGAPPPS (NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.: 96) na końcu C. Aktywność GLP-1 w takich analogach można dodatkowo zwiększyć dzięki modyfikacji aminokwasu w położeniu 18, 28 lub 29 bądź w położeniu 18 i 29, zgodnie z opisem w niniejszym dokumencie. [0021] Kolejne nieznaczne zwiększenie siły działania GLP-1 uzyskuje się dzięki modyfikacji aminokwasu w położeniu na dużą, aromatyczną resztę aminokwasową, ewentualnie Trp. [0022] Zmniejszoną aktywność wobec receptora GLP-1 uzyskuje się np. dzięki modyfikacji aminokwasu w położeniu 7, delecji aminokwasu lub aminokwasów po stronie C-końcowej aminokwasu w położeniu 27 lub 28, dzięki czemu otrzymuje się peptyd o długości 27 lub 28 aminokwasów, bądź przez ich kombinacjęzgodnie z opisem w niniejszym dokumencie. [0023] Utrzymanie aktywności po pegylowaniu uzyskuje się dzięki addycji GPSSGAPPPS (NR. ID. SEKW.: 9) na końcu C. [0024] Jak wykazano w niniejszym dokumencie, utrzymana lub zwiększona aktywność wobec każdego spośród receptora glukagonu, receptora GLP-1 i receptora GIP (w porównaniu do analogu na bazie glukagonu, zawierającego laktam, o aktywności GIP) uzyskuje się dzięki (i) substytucji aminokwasu Ser w położeniu 16 aminokwasem o wzorze IV:
10 9 [wzór IV], w którym n wynosi 1 do 16, lub 1 do, lub 1 do 7, lub 1 do 6, lub 2 do 6, lub 2, lub 3, lub 4, lub, przy czym każdy spośród R1 i R2 jest niezależnie wybrany z grupy składającej się z: H, C1-C18 alkil, (C1-C18 alkilo)oh, (C1-C18 alkilo)nh2, (C1-C18 alkilo)sh, (C0-C4 alkilo)(c3-c6)cykloalkil, (C0-C4 alkilo)(c2-c 1 2 heterocykl), (C0-C4 alkilo)(c6-c arylo)r7 i (C1-C4 alkilo)(c3-c9 heteroaryl), przy czym R7 oznacza H lub OH, a łańcuch boczny aminokwasu o wzorze IV zawiera wolną grupę aminową i (ii) substytucji aminokwasu Gln w położeniu aminokwasem alfa,alfa-dipodstawionym, np. AIB. W niektórych przykładach wykonania aminokwas w położeniu 16 stanowi Lys, a aminokwas w położeniu stanowi AIB. [002] Aktywność wobec każdego spośród receptora glukagonu, receptora GLP-1 i receptora GIP analogu zawierającego aminokwas o wzorze IV w położeniu 16 i aminokwas alfa,alfa-dipodstawiony w położeniu można dodatkowo zwiększyć w wyniku zwiększenia długości peptydu, np. w wyniku fuzji z peptydem stanowiącym przedłużenie C-końcowe, np. o długości około 1-21, około 9 do 21, około 6-18, około 9-12, lub około, lub 11 aminokwasów. W niektórych przykładach wykonania koniec C ulega przedłużeniu w wyniku fuzji z GPSSGAPPPS (NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.:
11 ), przy czym X stanowi Gly lub mały aminokwas alifatyczny lub niepolarny lub nieco polarny. W alternatywnych przykładach wykonania koniec C ulega przedłużeniu w wyniku fuzji z GPSSGAPPPS (NR. ID. SEKW.: 9) i 1-11 aminokwasów (np. 1- lub 1, 2, 3, 4,, 6, 7, 8, 9,, 11 aminokwasów) ulega fuzji z końcem C GPSSGAPPPS (NR. ID. SEKW.: 9) aminokwasów na końcu C GPSSGAPPPS (NR. ID. SEKW.: 9) mogą zawierać na przykład jeden lub większą liczbę małych aminokwasów alifatycznych, takich jak Ala lub Gly. W odniesieniu do tego przedłużenie na końcu C może na przykład stanowić GPSSGAPPPSXm, przy czym m wynosi 1 do 11 (np. 1 do ), a X stanowi Ala lub Gly. Ewentualnie 1 do 11 (np. 1 do ) aminokwasów w fuzji z końcem C NR. ID. SEKW.: 9 może stanowić kombinację różnych małych aminokwasów alifatycznych. Na przykład 1 do 11 (np. 1 do ) aminokwasów może stanowić kombinację reszt Ala i Gly. [0026] Zwiększenie aktywności wobec każdego spośród receptora glukagonu, GLP-1 i GIP analogu o aktywności GIP, na bazie glukagonu, w tym analogu zawierającego aminokwas o wzorze IV w położeniu 16 i aminokwas alfa,alfa-dipodstawiony w położeniu, można ponadto uzyskać w wyniku acylowania lub alkilowania aminokwasu znajdującego się w przedłużeniu na końcu C lub aminokwasu C-końcowego (np. aminokwasu dodawanego na końcu C przedłużenia C-końcowego). Acylowanie lub alkilowanie może dotyczyć aminokwasu znajdującego się na przykład w dowolnym spośród położeń 30, 31, 32, 33, 34, 3, 36, 37, 38, 39, 40, 41, 42, 43, 44, 4, 46, 47, 48, 49 i 0. W niektórych przykładach wykonania aminokwas ulegający acylowaniu lub alkilowaniu znajduje
12 się w położeniu 37, 38, 39, 40, 41, 42 lub 43. W niektórych przykładach wykonania aminokwas ulegający acylowaniu lub alkilowaniu stanowi Lys dołączoną do grupy acylowej lub alkilowej, np. C-C22. W niektórych przykładach wykonania Lys znajduje się po stronie C- końcowej przedłużenia C-końcowego składającego się z sekwencji aminokwasowej według NR. ID. SEKW.: 9, dzięki czemu Lys znajduje się w położeniu 40 analogu. Ewentualnie acylowane peptydy z przedłużeniem C- końcowym są także pegylowane, np. w położeniu 24. [0027] Zwiększenie aktywności analogu o aktywności GIP, na bazie glukagonu, wobec każdego spośród receptorów glukagonu, GLP-1 i GIP można ponadto uzyskać w wyniku acylowania lub alkilowania aminokwasu przez wstawkę (np. aminokwas, dipeptyd, tripeptyd, hydrofilowa wstawka dwufunkcyjna, hydrofobowa wstawka dwufunkcyjna). W niektórych przykładach wykonania analog o aktywności GIP, na bazie glukagonu, zawiera grupę acylową lub alkilową poprzez wstawkę, która jest dołączona do łańcucha bocznego aminokwasu w położeniu lub w położeniu 40 analogu. W innych przykładach wykonania analog zawiera przedłużenie C-końcowe o długości 1 do 21 aminokwasów po stronie C-końcowej aminokwasu w położeniu 29, a wstawka, która jest połączona kowalencyjnie z grupą acylową lub alkilową, jest dołączona do aminokwasu w przedłużeniu w położeniu odpowiadającym jednemu z położeń względem NR. ID. SEKW.: 1. W niektórych przykładach wykonania wstawka ma długość 3 do atomów. W specyficznych aspektach całkowita długość wstawki oraz grupy acylowej lub alkilowej wynosi około 14 do około 28 atomów. W
13 niniejszym dokumencie opisano ponadto odpowiednie wstawki w celach zwiększenia aktywności wobec jednego lub większej liczby spośród receptorów glukagonu, GLP-1 i GIP. [0028] Dowolne spośród modyfikacji zwiększających lub zmniejszających aktywność GIP, zwiększających lub zmniejszających aktywność wobec receptora glukagonu oraz zwiększających lub zmniejszających aktywność wobec receptora GLP-1 można zastosować pojedynczo lub w kombinacji. Dowolne spośród modyfikacji opisywanych powyżej można ponadto połączyć z innymi modyfikacjami zapewniającymi inne pożądane właściwości, takie jak zwiększona rozpuszczalność i(lub) stabilność i(lub) czas działania. Ewentualnie dowolne spośród modyfikacji opisywanych powyżej można połączyć z innymi modyfikacjami, które nie wpływają znacząco na rozpuszczalność lub stabilność lub aktywność. Przykładowe modyfikacje obejmują, ale nie ograniczająco: (A) Zwiększenie rozpuszczalności, na przykład w wyniku wprowadzenia jednego, dwóch, trzech lub większej liczby aminokwasów naładowanych w części C-końcowej natywnego glukagonu, korzystnie w położeniu po stronie C-końcowej względem położenia 27. Taki aminokwas naładowany można wprowadzić przez substytucję aminokwasu natywnego aminokwasem naładowanym, np. w położeniach 28 lub 29, lub ewentualnie przez dodanie aminokwasu naładowanego, np. po położeniu 27, 28 lub 29. W pewnych przykładach wykonania jeden, dwa, trzy lub wszystkie aminokwasy naładowane są naładowane ujemnie. W innych przykładach wykonania jeden, dwa, trzy lub wszystkie aminokwasy
14 naładowane są naładowane dodatnio. Takie modyfikacje zwiększają rozpuszczalność, np. zapewniają co najmniej 2-krotny, -krotny, -krotny, 1-krotny, 2-krotny, 30-krotny lub większy wzrost rozpuszczalności względem glukagonu natywnego w danym ph między około, a 8, np. ph 7, przy czym pomiaru dokonuje się po 24 godzinach w 2 C. (B) Zwiększenie rozpuszczalności i czasu działania lub okresu półtrwania w krwiobiegu przez dodanie ugrupowania hydrofilowego, takiego jak łańcuch poli(glikolu etylenowego), zgodnie z opisem w niniejszym dokumencie, np. w położeniu 16, 17,, 21, 24 lub 29, w obrębie przedłużenia C-końcowego lub na aminokwasie C-końcowym peptydu. (C) Zwiększenie rozpuszczalności i(lub) czasu działania lub okresu półtrwania w krwiobiegu i(lub) opóźnienie początku działania przez acylowanie lub alkilowanie peptydu glukagonu, zgodnie z opisem w niniejszym dokumencie. (D) Zwiększenie czasu działania lub okresu półtrwania w krwiobiegu w wyniku wprowadzenia odporności na trawienie przez dipeptydylopeptydazę IV (DPP IV) przez modyfikację aminokwasu w położeniu 1 lub 2 zgodnie z opisem w niniejszym dokumencie. (E) Zwiększenie stabilności dzięki modyfikacji Asp w położeniu 1, na przykład w wyniku delecji lub substytucji przez kwas glutaminowy, kwas homoglutaminowy, kwas cysteinowy lub kwas homocysteinowy. Takie modyfikacje mogą zmniejszyć degradację lub trawienie w ph w zakresie, do 8, na przykład utrzymanie co najmniej 7%, 80%, 90%, 9%,
15 %, 97%, 98% lub 99%, do 0% aktywności peptydu pierwotnego po 24 godzinach w 2 C. Takie modyfikacje zmniejszają trawienie wiązania peptydowego między Asp1-Ser16. (F) Zwiększenie stabilności dzięki modyfikacji Ser w położeniu 16, na przykład w wyniku substytucji przez Thr lub AIB. Takie modyfikacje także zmniejszają trawienie wiązania peptydowego między Asp1 a Ser16. (G) Zwiększenie stabilności dzięki modyfikacji metioniny w położeniu 27, na przykład w wyniku substytucji przez leucynę lub norleucynę. Takie modyfikacje mogą zmniejszyć degradację oksydacyjną. Stabilność można ponadto zwiększyć w wyniku modyfikacji Gln w położeniu lub 24, np. w wyniku substytucji przez Ala, Ser, Thr lub AIB. Takie modyfikacje mogą zmniejszyć degradację następującą w wyniku deamidacji Gln. Stabilność można zwiększyć w wyniku modyfikacji Asp w położeniu 21, np. w wyniku substytucji przez Glu. Takie modyfikacje mogą zmniejszyć degradację następującą w wyniku dehydratacji Asp w celu utworzenia cyklicznego sukcynoimidowego produktu pośredniego, po czym następuje izomeryzacja do izoasparaginianu. (H) Niekonserwatywne lub konserwatywne substytucje, addycje lub delecje, które nie zmieniają znacząco aktywności, na przykład konserwatywne substytucje w jednym lub większej liczbie spośród położeń 2,, 9,, 11, 12, 13, 14, 1, 16, 17, 18, 19,, 21, 24, 27, 28 lub 29; substytucja w jednym lub większej liczbie spośród tych położeń przez Ala; delecja aminokwasów w jednym lub większej liczbie spośród położeń 27, 28 lub 29; lub delecja aminokwasu 29, ewentualnie w połączeniu
16 z amidem lub estrem C-końcowym zamiast C-końcowej grupy kwasu karboksylowego; substytucja Lys w położeniu 12 przez Arg; substytucja Tyr w położeniu przez Val lub Phe. [0029] W niektórych przykładach wykonania peptydy glukagonu opisywane w niniejszym dokumencie mają wartość EC0 dla aktywności wobec aktywacji receptora GIP wynoszącą około 0 nm lub mniej, lub około 7, 0, 2,, 8, 6,, 4, 3, 2 lub 1 nm lub mniej. W niektórych przykładach wykonania peptydy glukagonu opisywane w niniejszym dokumencie mają wartość EC0 wobec receptora GIP wynoszącą około 0,001 nm, 0,01 nm lub 0,1 nm. W niektórych przykładach wykonania peptydy glukagonu opisywane w niniejszym dokumencie mają wartość EC0 wobec receptora GIP nie większą niż około 1 nm, 2 nm, 3 nm, 4 nm, nm, 6 nm, 8 nm, nm, 1 nm, nm, 2 nm, 30 nm, 40 nm, 0 nm, 7 nm lub 0 nm. W niektórych przykładach wykonania peptydy glukagonu mają wartość EC0 wobec aktywacji receptora glukagonu wynoszącą około 0 nm lub mniej, lub około 7, 0, 2,, 8, 6,, 4, 3, 2 lub 1 nm lub mniej. W niektórych przykładach wykonania peptydy glukagonu opisywane w niniejszym dokumencie mają wartość EC0 wobec receptora glukagonu wynoszącą około 0,001 nm, 0,01 nm lub 0,1 nm. W niektórych przykładach wykonania wartość EC0 wobec receptora glukagonu jest nie większa niż około 1 nm, 2 nm, 3 nm, 4 nm, nm, 6 nm, 8 nm, nm, 1 nm, nm, 2 nm, 30 nm, 40 nm, 0 nm, 7 nm lub 0 nm. W niektórych przykładach wykonania peptydy glukagonu mają wartość EC0 wobec aktywacji receptora GLP-1 wynoszącą około 0 nm lub mniej, lub około 7, 0, 2,, 8, 6,
17 , 4, 3, 2 lub 1 nm lub mniej. W niektórych przykładach wykonania peptydy glukagonu opisywane w niniejszym dokumencie mają wartość EC0 wobec receptora GLP-1 wynoszącą około 0,001 nm, 0,01 nm lub 0,1 nm. W niektórych przykładach wykonania wartość EC0 wobec receptora GLP-1 jest nie większa niż około 1 nm, 2 nm, 3 nm, 4 nm, nm, 6 nm, 8 nm, nm, 1 nm, nm, 2 nm, 30 nm, 40 nm, 0 nm, 7 nm lub 0 nm. Aktywację receptora można mierzyć za pomocą testów in vitro, w których dokonuje się pomiaru indukcji camp w komórkach HEK293 z nadekspresją receptora, np. dzięki testom w komórkach HEK293 poddanych kotransfekcji DNA kodującym receptor i gen lucyferazy połączony z elementem reagującym na camp zgodnie z opisem w przykładzie 16. [0030] W niektórych przykładach wykonania peptydy glukagonu wykazują co najmniej około 0,1%, 0,2%, 0,3%, 0,4%, 0,%, 0,6%, 0,7%, 0,8%, 0,9%,1%, %, %, %, 30%, 40%, 0%, 60%, 7%, 0%, 12%, 10%, 17% lub 0% lub większa aktywność wobec receptora GIP względem natywnego GIP (siła działania GIP). W niektórych przykładach wykonania peptydy glukagonu opisywane w niniejszym dokumencie wykazują nie więcej niż 00%, 000%, 0 000% lub % aktywności wobec receptora GIP względem natywnego GIP. W niektórych przykładach wykonania peptydy glukagonu wykazują co najmniej około 1%, %, %, %, 30%, 40%, 0%, 60%, 7%, 0%, 12%, 10%, 17%, 0%, %, 300%, 30%, 400%, 40% lub 00% lub większa aktywność wobec receptora glukagonu względem natywnego glukagonu (siła działania glukagonu). W niektórych przykładach wykonania peptydy glukagonu opisywane w niniejszym
18 dokumencie wykazują nie więcej niż 00%, 000%, 0 000% lub % aktywności wobec receptora glukagonu względem natywnego glukagonu. W niektórych przykładach wykonania peptydy glukagonu wykazują co najmniej około 0,1%, 0,2%, 0,3%, 0,4%, 0,%, 0,6%, 0,7%, 0,8%, 0,9%,1%, %, %, %, 30%, 40%, 0%, 60%, 7%, 0%, 12%, 10%, 17% lub 0% lub większa aktywność wobec receptora GLP-1 względem natywnego GLP- 1 (siła działania GLP-1). W niektórych przykładach wykonania peptydy glukagonu opisywane w niniejszym dokumencie wykazują nie więcej niż 00%, 000%, 0 000% lub % aktywności wobec receptora GLP- 1 względem natywnego GLP-1. Aktywność peptydu glukagonu wobec receptora względem natywnego liganda receptora oblicza się jako odwrotny stosunek wartości EC0 dla peptydu glukagonu względem natywnego liganda. [0031] W związku z tym jeden aspekt wynalazku ujawnia peptydy glukagonu, które wykazują działanie zarówno wobec receptora glukagonu, jak i receptora GIP ( działające koagonistycznie wobec glukagonu/gip ). Te peptydy glukagonu utraciły selektywność natywnego glukagonu wobec receptora glukagonu w porównaniu do receptora GIP. W niektórych przykładach wykonania wartość EC0 peptydu glukagonu wobec receptora GIP różni się o mniej niż około 0-krotnie, 40-krotnie, 30- krotnie lub -krotnie (jest większa lub mniejsza) od jego wartości EC0 wobec receptora glukagonu. W niektórych przykładach wykonania siła działania GIP peptydu glukagonu różni się o mniej niż około 00-, 40-, 400-, 30-, 300-, -, 0-, 10-, 0-, 7-, 0-, 2-, -, 1-, - lub -krotnie (jest większa lub
19 mniejsza) od jego siły działania glukagonu. W niektórych przykładach wykonania stosunek wartości EC0 peptydu glukagonu wobec receptora GIP podzielonej przez wartość EC0 peptydu glukagonu wobec receptora glukagonu jest mniejszy niż około 0, 7, 60, 0, 40, 30,, 1, lub. W niektórych przykładach wykonania stosunek wartości EC0 wobec receptora GIP podzielonej przez wartość EC0 wobec receptora glukagonu wynosi około 1 lub mniej niż około 1 (np. około 0,01, 0,013, 0,0167, 0,02, 0,02, 0,03, 0,0, 0,067, 0,1, 0,2). W niektórych przykładach wykonania stosunek siły działania GIP peptydu glukagonu w porównaniu do siły działania jako glukagonu peptydu glukagonu jest mniejszy niż około 00, 40, 400, 30, 300,, 0, 10, 0, 7, 60, 0, 40, 30,, 1, lub. W niektórych przykładach wykonania stosunek siły działania wobec receptora GIP podzielonej przez siłę działania wobec receptora glukagonu wynosi około 1 lub mniej niż około 1 (np. około 0,01, 0,013, 0,0167, 0,02, 0,02, 0,03, 0,0, 0,067, 0,1, 0,2). W niektórych przykładach wykonania aktywność GLP-1 zostaje znacząco zmniejszona lub zniesiona, np. w wyniku modyfikacji aminokwasu w położeniu 7, delecji aminokwasu lub aminokwasów po stronie C-końcowej aminokwasu w położeniu 27 lub 28, dzięki czemu otrzymuje się peptyd o długości 27 lub 28 aminokwasów, bądź przez ich kombinację. [0032] Inny aspekt wynalazku dostarcza peptydy glukagonu, które wykazują działanie wobec receptorów glukagonu, GIP i GLP-1 ( działające triagonistycznie wobec glukagonu/gip/glp-1 ). Te peptydy glukagonu
20 utraciły selektywność natywnego glukagonu wobec receptora glukagonu w porównaniu do zarówno receptorów GLP-1, jak i GIP. W niektórych przykładach wykonania wartość EC0 peptydu glukagonu wobec receptora GIP różni się o mniej niż około 0-krotnie, 40-krotnie, 30- krotnie lub -krotnie (jest większa lub mniejsza) od jego odpowiednich wartości EC0 wobec receptorów glukagonu i GLP-1. W niektórych przykładach wykonania siła działania GIP peptydu glukagonu różni się o mniej niż około 00-, 40-, 400-, 30-, 300-, -, 0-, 10-, 0-, 7-, 0-, 2-, -, 1-, - lub -krotnie (jest większa lub mniejsza) od jego sił działania jako glukagonu i GLP-1. W niektórych przykładach wykonania stosunek wartości EC0 peptydu o działaniu triagonistycznym wobec receptora GIP podzielonej przez wartość EC0 peptydu o działaniu triagonistycznym wobec receptora GLP-1 jest mniejszy niż około 0, 7, 60, 0, 40, 30,, 1, lub. W niektórych przykładach wykonania stosunek wartości EC0 wobec receptora GIP podzielonej przez wartość EC0 wobec receptora GLP-1 wynosi około 1 lub mniej niż około 1 (np. około 0,01, 0,013, 0,0167, 0,02, 0,02, 0,03, 0,0, 0,067, 0,1, 0,2). W niektórych przykładach wykonania stosunek siły działania GIP peptydu o działaniu triagonistycznym w porównaniu do siły działania GLP-1 peptydu o działaniu triagonistycznym jest mniejszy niż około 0, 7, 60, 0, 40, 30,, 1, lub. W niektórych przykładach wykonania stosunek siły działania wobec receptora GIP podzielonej przez siłę działania wobec receptora GLP-1 wynosi około 1 lub mniej niż około 1 (np. około 0,01, 0,013, 0,0167, 0,02, 0,02, 0,03, 0,0, 0,067, 0,1,
21 ,2). W powiązanych przykładach wykonania stosunek wartości EC0 peptydu o działaniu triagonistycznym wobec receptora GIP podzielonej przez wartość EC0 peptydu o działaniu triagonistycznym wobec receptora glukagonu jest mniejszy niż około 0, 7, 60, 0, 40, 30,, 1, lub. W niektórych przykładach wykonania stosunek wartości EC0 wobec receptora GIP podzielonej przez wartość EC0 wobec receptora glukagonu wynosi około 1 lub mniej niż około 1 (np. około 0,01, 0,013, 0,0167, 0,02, 0,02, 0,03, 0,0, 0,067, 0,1, 0,2). W niektórych przykładach wykonania stosunek siły działania GIP peptydu o działaniu triagonistycznym w porównaniu do siły działania glukagonu peptydu o działaniu triagonistycznym jest mniejszy niż około 00, 40, 400, 30, 300,, 0, 10, 0, 7, 60, 0, 40, 30,, 1, lub. W niektórych przykładach wykonania stosunek siły działania wobec receptora GIP podzielonej przez siłę działania wobec receptora glukagonu wynosi około 1 lub mniej niż około 1 (np. około 0,01, 0,013, 0,0167, 0,02, 0,02, 0,03, 0,0, 0,067, 0,1, 0,2). W niektórych przykładach wykonania stosunek wartości EC0 peptydu o działaniu triagonistycznym wobec receptora GLP-1 podzielonej przez wartość EC0 peptydu o działaniu triagonistycznym wobec receptora glukagonu jest mniejszy niż około 0, 7, 60, 0, 40, 30,, 1, lub. W niektórych przykładach wykonania stosunek wartości EC0 wobec receptora GLP-1 podzielonej przez wartość EC0 wobec receptora glukagonu wynosi około 1 lub mniej niż około 1 (np. około 0,01, 0,013, 0,0167, 0,02, 0,02, 0,03, 0,0, 0,067, 0,1, 0,2). W niektórych
22 przykładach wykonania stosunek siły działania GLP-1 peptydu o działaniu triagonistycznym w porównaniu do siły działania glukagonu o działaniu triagonistycznym jest mniejszy niż około 0, 7, 60, 0, 40, 30,, 1, lub. W niektórych przykładach wykonania stosunek siły działania wobec receptora GLP-1 podzielonej przez siłę działania wobec receptora glukagonu wynosi około 1 lub mniej niż około 1 (np. około 0,01, 0,013, 0,0167, 0,02, 0,02, 0,03, 0,0, 0,067, 0,1, 0,2). [0033] Jeszcze inny aspekt wynalazku dostarcza peptydy glukagonu, które wykazują działanie wobec receptorów GLP-1 i GIP, jednak w których aktywność glukagonu została znacznie zmniejszona lub zniesiona ( działające koagonistycznie wobec GIP/GLP-1 ), np. w wyniku modyfikacji aminokwasu w położeniu 3. Na przykład substytucja w tym położeniu przez aminokwas kwaśny, zasadowy lub hydrofobowy (kwas glutaminowy, ornitynę, norleucynę) zmniejsza aktywność glukagonu. W niektórych przykładach wykonania wartość EC0 peptydu glukagonu wobec receptora GIP różni się o mniej niż około 0- krotnie, 40-krotnie, 30-krotnie lub -krotnie (jest większa lub mniejsza) od jego wartości EC0 wobec receptora GLP-1. W niektórych przykładach wykonania siła działania GIP peptydu glukagonu różni się o mniej niż około 2-, -, 1-, - lub -krotnie (jest większa lub mniejsza) od jego siły działania GLP-1. W niektórych przykładach wykonania te peptydy glukagonu mają około % lub mniej aktywności natywnego glukagonu wobec receptora glukagonu, np. około 1-% lub około 0,1-% lub więcej niż około 0,1%, ale mniej niż około
23 %. W niektórych przykładach wykonania stosunek wartości EC0 peptydu glukagonu wobec receptora GIP podzielonej przez wartość EC0 peptydu glukagonu wobec receptora GLP-1 jest mniejszy niż około 0, 7, 60, 0, 40, 30,, 1, lub i nie mniejszy niż 1. W niektórych przykładach wykonania stosunek siły działania GIP peptydu glukagonu w porównaniu do siły działania GLP-1 peptydu glukagonu jest mniejszy niż około 0, 7, 60, 0, 40, 30,, 1, lub i nie mniejszy niż 1. [0034] Kolejny aspekt wynalazku dostarcza peptydy glukagonu, które wykazują działanie wobec receptora GIP, w przypadku których aktywność glukagonu i GLP-1 została znacznie zmniejszona lub zniesiona ( peptydy glukagonu działające agonistycznie wobec GIP ), np. w wyniku modyfikacji aminokwasów w położeniach 3 i 7. W niektórych przykładach wykonania te peptydy glukagonu mają około % lub mniej aktywności natywnego glukagonu wobec receptora glukagonu, np. około 1-% lub około 0,1-% lub więcej niż około 0,1%, 0,% lub 1%, ale mniej niż około 1%, % lub %. W niektórych przykładach wykonania te peptydy glukagonu mają ponadto około % lub mniej aktywności natywnego GLP-1 wobec receptora GLP-1, np. około 1-% lub około 0,1-% lub więcej niż około 0,1%, 0,% lub 1%, ale mniej niż około 1%, % lub %. [003] Zgodnie z niektórymi przykładami wykonania analog glukagonu (NR. ID. SEKW.: 1) mający aktywność agonistyczną wobec GIP zawiera NR. ID. SEKW.: 1 z (a) modyfikacją aminokwasu w położeniu 1 nadającą aktywność agonistyczną wobec GIP, (b) modyfikacją stabilizującą
24 strukturę helisy alfa części C-końcowej (aminokwasy 12-29) analogu i (c) ewentualnie 1 do (np. 1, 2, 3, 4,, 6, 7, 8, 9, ) dodatkowych modyfikacji aminokwasów. W niektórych przykładach wykonania analog wykazuje co najmniej około 1% aktywności natywnego GIP wobec receptora GIP lub dowolny inny poziom aktywności wobec receptora GIP zgodnie z opisem w niniejszym dokumencie. Modyfikacją stabilizującą strukturę helisy alfa może być dowolna spośród modyfikacji znanych w stanie techniki, na przykład dowolna z opisanych w niniejszym dokumencie. Patrz treść części Stabilizacja struktury helisy alfa. W niektórych przykładach wykonania modyfikacją stabilizującą strukturę helisy alfa jest modyfikacja wybrana z grupy składającej się z: (i) mostek laktamowy między łańcuchami bocznymi aminokwasów w położeniach i oraz i+4 lub między łańcuchami bocznymi aminokwasów w położeniach j i j+3, przy czym i wynosi 12, 13, 16, 17, lub 24 i przy czym j wynosi 17 oraz (ii) jeden, dwa, trzy lub wszystkie spośród aminokwasów w położeniach 16,, 21 i 24 analoga jest podstawionych przez aminokwas α,α-dipodstawiony. Takie analogi glukagonu mające aktywność agonistyczną wobec GIP opisano dodatkowo w niniejszym dokumencie. [0036] W niektórych przykładach wykonania dostarcza się tu analog glukagonu (NR. ID. SEKW.: 1) mający działanie agonistyczne wobec GIP z następującymi modyfikacjami: (a) modyfikacja aminokwasu w położeniu 1, (b) (i) mostek laktamowy między łańcuchami bocznymi aminokwasów w położeniach i oraz i+4 lub między łańcuchami bocznymi aminokwasów w położeniach j i j+3, przy czym i wynosi 12, 13, 16, 17, lub 24 i przy
25 czym j wynosi 17, lub (ii) substytucja aminokwasu przez aminokwas α,α-dipodstawiony w jednym, dwóch, trzech lub wszystkich spośród położeń 16,, 21 lub 24, (c) modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29 oraz (d) 1, 2, 3, 4,, 6 lub 8 dodatkowych modyfikacji aminokwasów, przy czym wartość EC0 analogu wobec aktywacji receptora GIP wynosi około 0 nm lub mniej. [0037] W niektórych przykładach wykonania (a) modyfikacja aminokwasu w położeniu 1 stanowi substytucję His w położeniu 1 przez duży aminokwas aromatyczny, ewentualnie Tyr, Phe, Trp, amino-phe, nitro-phe, chloro-phe, sulfo-phe, 4-pirydylo-Ala, metylo-tyr lub 3-amino-Tyr, (b) (i) mostek laktamowy znajduje się między aminokwasami w położeniach 16 i, przy czym jeden spośród aminokwasów w położeniach 16 i jest podstawiony przez Glu, a drugi spośród aminokwasów w położeniach 16 i jest podstawiony przez Lys, lub (ii) aminokwas α,α-dipodstawiony stanowi AIB, (c) Met w położeniu 27 jest podstawiona przez duży aminokwas alifatyczny, ewentualnie Leu, (d) Asn w położeniu 28 jest podstawiona przez mały aminokwas alifatyczny, ewentualnie Ala, oraz (e) Thr w położeniu 29 jest podstawiona przez mały aminokwas alifatyczny, ewentualnie Gly. [0038] Analog może zawierać dodatkowe modyfikacje, w tym bez ograniczania: (a) modyfikacja aminokwasu w położeniu 12, ewentualnie substytucja przez Ile,
26 (b) modyfikacje aminokwasów w położeniach 17 i 18, ewentualnie substytucja przez Q w położeniu 17 i A w położeniu 18, (c) addycja GPSSGAPPPS (NR. ID. SEKW.: 9) do końca C, lub dowolna ich kombinacja. [0039] Analog może alternatywnie lub dodatkowo zawierać dalsze modyfikacje, w tym bez ograniczania: (a) Ser w położeniu 2 podstawiona przez D-Ser, Ala, D- Ala, Gly, N-metylo-Ser, AIB, Val lub kwas α-amino-nmasłowy; (b) Tyr w położeniu podstawiona przez Trp, Lys, Orn, Glu, Phe lub Val; (c) połączenie grupy acylowej z Lys w położeniu ; (d) Lys w położeniu 12 podstawiona przez Arg; (e) Ser w położeniu 16 podstawiona przez Glu, Gln, kwas homoglutaminowy, kwas homocysteinowy, Thr, Gly lub AIB; (f) Arg w położeniu 17 podstawiona przez Gln, Lys lub Glu; (g) Arg w położeniu 18 podstawiona przez Ala, Ser, Thr, lub Gly; (h) Gln w położeniu podstawiona przez Ala, Ser, Thr, Lys, cytrulinę, Arg, Orn lub AIB; (i) Asp w położeniu 21 podstawiona przez Glu, kwas homoglutaminowy, kwas homocysteinowy; (j) Val w położeniu 23 podstawiona przez Ile; (k) Gln w położeniu 24 podstawiona przez Asn, Ala, Ser, Thr, Glu, Lys lub AIB; oraz (l) substytucja konserwatywna w dowolnym z położeń 2,, 9,, 11, 12, 13, 14, 1, 16, 17, 18, 19,, 21, 24, 27, 28 i 29, bądź dowolna ich kombinacja. [0040] W niektórych przykładach wykonania, jeśli peptyd
27 glukagonu nie jest pegylowany, wartość EC0 analogu wobec aktywacji receptora GIP wynosi około 4, 2, 1 nm lub mniej, lub analog ma co najmniej około 1%, 2%, 3%, 4% lub % aktywności natywnego GIP wobec receptora GIP. W powiązanych przykładach wykonania wartość EC0 niepegylowanego analogu wobec aktywacji receptora GLP-1 wynosi około 4, 2, 1 nm lub mniej lub ma on co najmniej około 1%, 2%, 3%, 4% lub % aktywności natywnego GLP-1 wobec receptora GLP-1. W jeszcze innych przykładach wykonania wartość EC0 niepegylowanego analogu wobec aktywacji receptora glukagonu wynosi około 4, 2, 1 nm lub mniej lub ma on co najmniej około %, %, 1% lub % aktywności natywnego glukagonu wobec receptora glukagonu. W niektórych przykładach wykonania niepegylowany analog ma mniej niż około 1% aktywności natywnego glukagonu wobec receptora glukagonu. W innych przykładach wykonania niepegylowany analog ma mniej niż około %, % lub 1% aktywności natywnego GLP-1 wobec receptora GLP-1. [0041] W niektórych przykładach wykonania peptyd glukagonu jest połączony kowalencyjnie z ugrupowaniem hydrofilowym w dowolnym spośród położeń aminokwasów 16, 17,, 21, 24 lub 29, po położeniu 29 na dodanym aminokwasie (np. położenie 30) w obrębie przedłużenia C-końcowego lub na aminokwasie C-końcowym. W niektórych przykładach wykonania to ugrupowanie hydrofilowe jest połączone kowalencyjnie z resztą Lys, Cys, Orn, homocysteiny lub acetylofenyloalaniny w dowolnym spośród tych położeń. Do przykładowych ugrupowań hydrofilowych należy poli(glikol etylenowy) (PEG), na przykład o masie cząsteczkowej około 00 daltonów do
28 około daltonów lub około 000 daltonów do około daltonów. [0042] W takich przykładach wykonania, w których analogi są połączone z ugrupowaniami hydrofilowymi, takimi jak PEG, względne wartości EC0 wobec jednego lub większej liczby receptorów mogą być wyższe, np. około -krotnie wyższe, w porównaniu do analogu niezawierającego ugrupowania hydrofilowego. Na przykład wartość EC0 pegylowanego analogu wobec aktywacji receptora GIP wynosi około nm lub mniej lub analog ma co najmniej około 0,1%, 0,2%, 0,3%, 0,4% lub 0,% aktywności natywnego GIP wobec receptora GIP. W powiązanych przykładach wykonania wartość EC0 pegylowanego analogu wobec aktywacji receptora GLP-1 wynosi około nm lub mniej lub ma on co najmniej około 0,1%, 0,2%, 0,3%, 0,4% lub 0,% aktywności natywnego GLP-1 wobec receptora GLP-1. W jeszcze innych powiązanych przykładach wykonania wartość EC0 pegylowanego analogu wobec aktywacji receptora glukagonu wynosi około nm lub mniej lub ma on co najmniej około 0,%, 1%, 1,% lub 2% aktywności natywnego glukagonu wobec receptora glukagonu. W niektórych przykładach wykonania analog ma mniej niż około 1% aktywności natywnego glukagonu wobec receptora glukagonu. W innych przykładach wykonania analog ma mniej niż około %, % lub 1% aktywności natywnego GLP-1 wobec receptora GLP-1. [0043] Peptyd glukagonu może być częścią dimeru, trimeru lub multimeru wyższego rzędu zawierającego co najmniej dwa, trzy lub więcej peptydów połączonych łącznikiem, przy czym co najmniej jeden lub obydwa
29 peptydy stanowią peptyd glukagonu. Dimer może stanowić homodimer lub heterodimer. W niektórych przykładach wykonania łącznik jest wybrany z grupy składającej się z dwufunkcyjnego tiolowego łącznika sieciującego i dwufunkcyjnego aminowego łącznika sieciującego. W niektórych przykładach wykonania łącznik stanowi PEG, np. PEG o masie kda, PEG o masie kda. W niektórych przykładach wykonania łącznik stanowi wiązanie disiarczkowe. Na przykład każdy monomer w dimerze może zawierać resztę Cys (np. Cys końcowa lub w położeniu wewnętrznym), a atom siarki w każdej reszcie Cys uczestniczy w tworzeniu wiązania disiarczkowego. W niektórych aspektach wynalazku monomery są połączone poprzez aminokwasy końcowe (np. N-końcowe lub C- końcowe), przez aminokwasy wewnętrzne lub poprzez aminokwas końcowy co najmniej jednego monomeru i aminokwas wewnętrzny co najmniej jednego innego monomeru. W konkretnych aspektach monomery nie są połączone poprzez aminokwas N-końcowy. W niektórych aspektach monomery w multimerze są połączone ze sobą w orientacji ogon do ogona, w której aminokwasy C- końcowe każdego monomeru są połączone ze sobą. Ugrupowanie sprzężone może być połączone kowalencyjnie z dowolnym spośród peptydów glukagonu opisanych w niniejszym dokumencie, w tym z dimerem, trimerem lub multimerem wyższego rzędu. [0044] Dowolne spośród modyfikacji opisanych w niniejszym dokumencie, zwiększających aktywność wobec receptora glukagonu, zachowujących częściową aktywność wobec receptora glukagonu, zwiększających rozpuszczalność, zwiększających stabilność lub
30 zmniejszających degradację można zastosować do peptydów glukagonu pojedynczo lub w kombinacji. W niektórych przykładach wykonania peptydy glukagonu są rozpuszczalne w stężeniu co najmniej 1 mg/ml w ph między 6 a 8, lub między 6 a 9, lub między 7 a 9 (np. ph 7) i ewentualnie zachowują co najmniej 9% peptydu pierwotnego (np. % lub mniej peptydu pierwotnego ulega degradacji lub trawieniu) po 24 godzinach w 2 C. [004] Dostarcza się jałowe kompozycje farmaceutyczne zawierające dopuszczalny farmaceutycznie nośnik lub rozcieńczalnik i zestawy zawierające urządzenia. Dostarcza się sposoby zmniejszania wzrostu masy lub indukowania utraty masy obejmujące podawanie pacjentowi tego wymagającemu takich kompozycji farmaceutycznych w ilości skutecznej do zmniejszenia wzrostu masy lub indukowania utraty masy. Dostarcza się sposoby leczenia cukrzycy obejmujące podawanie pacjentowi tego wymagającemu takich kompozycji farmaceutycznych w ilości skutecznej do zmniejszenia poziomów glukozy we krwi. [0046] Wszystkie sposoby terapeutyczne, kompozycje farmaceutyczne, zestawy i inne podobne przykłady wykonania opisywane w niniejszym dokumencie przewidują, że zastosowanie określeń peptydy, pochodne o działaniu agonistycznym, koagonistycznym, triagonistycznym lub analogi obejmują wszystkie ich dopuszczalne farmaceutycznie sole lub estry. [0047] Celem powyższego streszczenia nie jest zdefiniowanie każdego aspektu wynalazku, a w innych częściach, na przykład w Opisie szczegółowym, podano dodatkowe przykłady wykonania. Przewiduje się, że cały
31 dokument ma stanowić jednolite ujawnienie oraz należy rozumieć, że uwzględnia się wszystkie możliwe kombinacje cech opisywanych w niniejszym dokumencie, nawet jeśli danej kombinacji cech nie ujęto łącznie w tym samym zdaniu lub akapicie lub części niniejszego dokumentu. [0048] Ponadto wynalazek obejmuje dowolny jeden lub wszystkie przykłady wykonania wynalazku mające w dowolnym sensie węższy zakres od zmian zdefiniowanych w określonych akapitach niniejszego dokumentu. Na przykład, jeśli pewne aspekty wynalazku opisuje się w postaci kategorii, należy rozumieć, że każdy element kategorii stanowi pojedynczo przykład wykonania wynalazku, a kombinacje dwóch lub większej liczby elementów kategorii stanowią przykłady wykonania wynalazku. SKRÓCONY OPIS RYSUNKÓW [0049] Figura 1 przedstawia wykres zmiany procentowej masy ciała myszy w zależności od czasu (dni) po podaniu samego nośnika (niewypełnione odwrócone trójkąty), chimery 2 AIB2 Cys24 (40K PEG) (wypełnione kwadraty), antagonisty GIP Pro3 Cys24 GIP-NH2 (1-42) 40K PEG (niewypełnione romby), agonisty GIP AIB2 Cys24 GIP (1-42) 40K PEG (niewypełnione proste trójkąty z linią kropkowaną) lub niepowiązanego hormonu peptydowego (zacienione odwrócone trójkąty). Figura 2 przedstawia wykres przyjmowania pokarmu (w gramach) przez myszy w zależności od czasu po podaniu samego nośnika (niewypełnione odwrócone trójkąty), chimery 2 AIB2 Cys24 (40K PEG) (wypełnione kwadraty),
32 antagonisty GIP Pro3 Cys24 GIP-NH2 (1-42) 40K PEG (niewypełnione romby), agonisty GIP AIB2 Cys24 GIP (1-42) 40K PEG (niewypełnione proste trójkąty z linią kropkowaną) lub niepowiązanego hormonu peptydowego (zacienione odwrócone trójkąty). Figura 3 przedstawia wykres zmiany poziomów glukozy we krwi (mg/dl) u myszy w dniu 7 po podaniu samego nośnika (czarny słupek), chimery 2 AIB2 Cys24 (40K PEG) (biały słupek), antagonisty GIP Pro3 Cys24 GIP-NH2 (1-42) 40K PEG (zacieniony słupek), agonisty GIP AIB2 Cys24 GIP (1-42) 40K PEG (słupek kreskowany poziomo) lub niepowiązanego hormonu peptydowego (słupek kreskowany pionowo). Figura 4 przedstawia wykres zmiany procentowej masy ciała myszy w zależności od czasu (dni) po podaniu samego nośnika (wypełnione odwrócone trójkąty), chimery 2 AIB2 (niewypełnione romby), chimery 2 AIB2 laktamu (niewypełnione trójkąty), peptydu o działaniu triagonistycznym MT-170 (niewypełnione kwadraty), peptydu o działaniu koagonistycznym wobec GIP/glukagonu MT-182 (wypełnione romby), peptydu o działaniu koagonistycznym wobec GLP-1/GIP MT-178 (zacienione trójkąty z linią kropkowaną) lub peptydu o działaniu koagonistycznym wobec GIP/glukagonu MT-179 (wypełnione kwadraty). Należy zauważyć, że PEGylowany MT-179 działa triagonistycznie. Figura przedstawia wykres zmiany procentowej masy ciała myszy w zależności od czasu (dni) po podaniu samego nośnika (niewypełnione proste trójkąty), agonisty GLP-1 E 16 w stężeniu nmol/kg (wypełnione odwrócone trójkąty) lub 3 nmol/kg (niewypełnione
33 kwadraty), peptydu o działaniu triagonistycznym MT-170 w stężeniu nmol/kg (niewypełnione odwrócone trójkąty) lub 3 nmol/kg (wypełnione romby) lub peptydu o działaniu koagonistycznym wobec GLP-1/GIP MT-178 w stężeniu nmol/kg (szare odwrócone trójkąty) lub 3 nmol/kg (szare kwadraty). Figura 6 przedstawia wykres zmiany poziomów glukozy we krwi (mg/dl) u myszy w dniu 7 po podaniu samego nośnika (czarny słupek), agonisty GLP-1 E 16 w stężeniu nmol/kg (biały słupek) lub 3 nmol/kg (szary słupek), peptydu o działaniu triagonistycznym MT-170 w stężeniu nmol/kg (słupek kreskowany poziomo) lub 3 nmol/kg (słupek kreskowany pionowo) lub peptydu o działaniu koagonistycznym wobec GLP-1/GIP MT-178 w stężeniu nmol/kg (słupek kreskowany ukośnie w górę) lub 3 nmol/kg (słupek kreskowany ukośnie w dół). Figura 7 przedstawia wykres poziomów glukozy we krwi (mg/dl) w zależności od czasu przed i po wstrzyknięciu glukozy (podanej w punkcie czasowym 0) u myszy, którym wstrzyknięto (w punkcie czasowym -60) kontrolę (nośnik), kontrolę (peptyd o działaniu agonistycznym wobec GLP-1), analog glukagonu zawierający laktam (cykliczny), pegylowany o aktywności GIP ( mt-178 ) lub analog glukagonu niezawierający laktamu (liniowy), pegylowany o aktywności GIP ( mt-274 ) w dawce 1, 3 lub nmol/kg/tydzień. Dane na tym rysunku nie obejmują danych czterech myszy, ponieważ stwierdzono u nich zachowanie agresywne i znaczne zmniejszenie masy ciała. Figura 8 przedstawia wykres poziomu glukozy we krwi (mg/dl) w zależności od czasu przed i po wstrzyknięciu glukozy (podanej w punkcie czasowym 0) u myszy, którym
34 wstrzyknięto (24 godziny przed wstrzyknięciem glukozy) kontrolę (nośnik), kontrolę (peptyd o działaniu agonistycznym wobec GLP-1), mt-178 lub mt-274 w dawce 1, 3, lub nmol/kg/tydzień. Dane na tym rysunku nie obejmują danych czterech myszy, ponieważ stwierdzono u nich zachowanie agresywne i znaczne zmniejszenie masy ciała. Figura 9 przedstawia wykres poziomów glukozy we krwi (mg/dl) u myszy 0 lub 7 dni po wstrzyknięciu kontroli (nośnik), kontroli (peptyd o działaniu agonistycznym wobec GLP-1), mt-178 lub mt-274 w dawce 1, 3 lub nmol/kg/tydzień. Dane na tym rysunku nie obejmują danych czterech myszy, ponieważ stwierdzono u nich zachowanie agresywne i znaczne zmniejszenie masy ciała. Figura przedstawia wykres zmiany procentowej masy ciała u myszy 0, 1, 3, i 7 dni po wstrzyknięciu kontroli (podłoże), kontroli (peptyd o działaniu agonistycznym wobec GLP-1), mt-178 lub mt-274 w dawce 1, 3 lub nmol/kg/tydzień. Dane na tym rysunku nie obejmują danych czterech myszy, ponieważ stwierdzono u nich zachowanie agresywne i znaczne zmniejszenie masy ciała. Figura 11 przedstawia wykres poziomów glukozy we krwi (mg/dl) u myszy 0 lub 7 dni po wstrzyknięciu kontroli (nośnik), kontroli (peptyd o działaniu agonistycznym wobec GLP-1), mt-178, mt-178(te), mt-274 lub mt-274(te) w dawce lub 3 nmol/kg/tydzień. TE oznacza grupę PEG dołączoną do Cys w położeniu 40. Figura 12 przedstawia wykres zmiany poziomu glukozy we krwi (mg/dl) u myszy 7 dni po wstrzyknięciu kontroli (nośnik), kontroli (peptyd o działaniu agonistycznym
35 wobec GLP-1), mt-178, mt-178(te), mt-274 lub mt-274(te) w dawce lub 3 nmol/kg/tydzień. TE oznacza grupę PEG dołączoną do Cys w położeniu 40. Figura 13 przedstawia wykres zmiany procentowej masy ciała u myszy 0, 1, 3,, 7 i dni po wstrzyknięciu kontroli (nośnik), kontroli (peptyd o działaniu agonistycznym wobec GLP-1), mt-178, mt-178(te), mt-274 lub mt-274(te) w dawce lub 3 nmol/kg/tydzień. TE oznacza grupę PEG dołączoną do Cys w położeniu 40. Figura 14 przedstawia wykres zmiany procentowej masy ciała u myszy 7 dni po wstrzyknięciu kontroli (nośnik), kontroli (peptyd o działaniu agonistycznym wobec GLP- 1), mt-178, mt-178(te), mt-274 lub mt-274(te) w dawce lub 3 nmol/kg/tydzień. TE oznacza grupę PEG dołączoną do Cys w położeniu 40. Figura 1 przedstawia wykres poziomów glukozy we krwi (mg/dl) u myszy 0 lub 7 dni po wstrzyknięciu kontroli (nośnik), kontroli (peptyd o działaniu agonistycznym wobec GLP-1), mt-178, mt-274, liniowego niepegylowanego nieacylowanego peptydu ( mt-311 ), peptydu liniowego acylowanego kwasem tłuszczowym C14 ( mt-309 ), peptydu liniowego acylowanego kwasem tłuszczowym C16 ( mt-298 ) lub peptydu liniowego acylowanego kwasem tłuszczowym C18 ( mt-3 ) w dawce nmol/kg. Figura 16 przedstawia wykres zmiany procentowej masy ciała u myszy 0, 1, 3, i 7 dni po wstrzyknięciu kontroli (nośnik), kontroli (peptyd o działaniu agonistycznym wobec GLP-1), mt-178, mt-274, mt-311, mt- 309, mt-298 lub mt-3 w dawce nmol/kg. Figura 17 przedstawia wykres zmiany procentowej masy ciała u myszy 7 dni po wstrzyknięciu kontroli (nośnik),
36 kontroli (peptyd o działaniu agonistycznym wobec GLP- 1), mt-178, mt-274, mt-311, mt-309, mt-298, lub mt-3 w dawce nmol/kg. Figura 18 przedstawia wykres zmiany poziomów glukozy we krwi (mg/dl) u myszy 0 lub 7 dni po wstrzykiwaniu raz na dobę przez 7 dni kontroli (nośnik), liraglutydu (acylowany analog GLP-1), niepegylowanego peptydu liniowego acylowanego kwasem tłuszczowym C14 ( mt- 260 ), niepegylowanego peptydu liniowego acylowanego kwasem tłuszczowym C16 ( mt-261 ) lub niepegylowanego peptydu liniowego acylowanego kwasem tłuszczowym C18 ( mt-262 ) w dawce 2 lub 12 nmol/kg. Figura 19 przedstawia wykres zmiany procentowej masy ciała u myszy 0, 1, 3, i 7 dni po wstrzyknięciu kontroli (nośnik), liraglutydu, mt-260, mt-261 lub mt- 262 w dawce 2 lub 12 nmol/kg. Figura przedstawia wykres zmiany procentowej masy ciała u myszy 7 dni po wstrzyknięciu kontroli (nośnik), liraglutydu, mt-260, mt-261 lub mt-262 w dawce 2 lub 12 nmol/kg. Figura 21 przedstawia wykres zmiany masy ciała (g) u myszy 0, 1, 3, i 7 dni po pierwszym wstrzyknięciu kontroli (nośnik), liraglutydu (30 nmol/kg/dobę) lub mt-261 (0,3, 1, 3, lub 30 nmol/kg/dobę). Figura 22 przedstawia wykres masy tłuszczu u myszy 7 dni po pierwszym wstrzyknięciu kontroli (nośnik), liraglutydu (30 nmol/kg/dobę) lub mt-261 (0,3, 1, 3, lub 30 nmol/kg/dobę). Figura 23 przedstawia wykres poziomów glukozy we krwi (mg/dl) u myszy 0 i 7 dni po pierwszym wstrzyknięciu kontroli (nośnik), liraglutydu (30 nmol/kg/dobę) lub
37 mt-261 (0,3, 1, 3, lub 30 nmol/kg/dobę). Figura 24 przedstawia wykres liniowy zmiany masy ciała (zmiana procentowa) w zależności od czasu u myszy, którym wstrzyknięto mt-263, eksendynę 4 lub kontrolę (nośnik) w dawkach (nmol/kg/dobę) podanych w (). Figura 2 przedstawia wykres słupkowy całkowitej zmiany masy ciała (%) (mierzonej w dniu 7 w porównaniu z dniem 0) u myszy, którym wstrzyknięto mt-263, eksendynę 4 lub kontrolę (nośnik) w dawkach (nmol/kg/dobę) podanych w (). Figura 26 przedstawia wykres słupkowy zmiany poziomów glukozy we krwi (mg/dl) (mierzonych w dniu 7 w porównaniu z dniem 0) u myszy, którym wstrzyknięto mt- 263, eksendynę 4 lub kontrolę (nośnik) w dawkach (nmol/kg/dobę) podanych w (). Figura 27 przedstawia wykres zmiany procentowej masy ciała u myszy 0, 1, 3, i 7 dni po pierwszym wstrzyknięciu kontroli (nośnik), liraglutydu, mt-277, mt-278 lub mt-279. Figura 28 przedstawia wykres poziomów glukozy we krwi (mg/dl) u myszy 0 lub 7 dni po pierwszym wstrzyknięciu kontroli (nośnik), liraglutydu, mt-277, mt-278 lub mt Figura 29 przedstawia wykres zmiany całkowitej masy ciała (%) u myszy mierzonej 7 dni po podaniu mt-331, mt-311 lub kontroli (nośnik). Dawki (nmol/kg) podano w (). Figura 30 przedstawia wykres całkowitego przyjmowania pokarmu (g) przez myszy mierzonego 7 dni po podaniu mt- 331, mt-311 lub kontroli (nośnik). Dawki (nmol/kg) podano w ().
38 Figura 31 przedstawia wykres zmiany całkowitej poziomów glukozy we krwi u myszy mierzonych 7 dni po podaniu mt- 331, mt-311 lub kontroli (nośnik). Dawki (nmol/kg) podano w (). Figura 32 przedstawia wykres zmiany całkowitej masy ciała u myszy mierzonej 7 dni po podaniu mt-331, mt-33 lub kontroli (nośnik) w określonej dawce (nmol/kg) podanej w (). Figura 33 przedstawia wykres całkowitego przyjmowania pokarmu (g) przez myszy mierzonego 7 dni po podaniu mt- 331, mt-33 lub kontroli (nośnik) w określonej dawce (nmol/kg) podanej w (). Figura 34 przedstawia wykres zmiany poziomów glukozy we krwi (mg/dl) u myszy mierzonych 7 dni po podaniu mt- 331, mt-33 lub kontroli (nośnik) w określonej dawce (nmol/kg) podanej w (). Figura 3 przedstawia wykres zmiany całkowitej masy ciała (%) u myszy mierzonej 7 dni po pierwszym podaniu mt-277, mt-278, mt-279 lub kontroli (nośnik). Figura 36 przedstawia wykres zmiany całkowitej masy ciała (%) u myszy mierzonej 6 dni po pierwszym podaniu mt-261, mt-309 lub kontroli (nośnik). Figura 37 przedstawia wykres poziomów glukozy we krwi (mg/dl) u myszy mierzonych 6 dni po pierwszym podaniu mt-261, mt-309 lub kontroli (nośnik). Pierwszy słupek w każdej parze słupków o tym samym cieniowaniu przedstawia poziomy glukozy we krwi mierzone w dniu 0, a drugi słupek w każdej parze stanowi poziomy w dniu 6. Figura 38 przedstawia wykres słupkowy zmiany całkowitej masy ciała (%) mierzonej 6 dni po pierwszym podaniu mt- 261 (w porównaniu z masą ciała mierzoną w pierwszym
39 dniu podawania) u myszy, którym wstrzyknięto kontrolę (nośnik) lub mt-261 jak dodatkowo opisano w niniejszym dokumencie. OPIS SZCZEGÓŁOWY [000] Niniejszy wynalazek dotyczy analogu glukagonu (NR. ID. SEKW.: 1) mającego działanie agonistyczne wobec GIP z modyfikacjami zdefiniowanymi w zastrzeżeniach. Te modyfikacje zdefiniowane w zastrzeżeniach opisano dodatkowo szczegółowo poniżej. [001] Niniejszy wynalazek dotyczy ponadto peptydu zdefiniowanego w zastrzeżeniach 12 do 17. [002] Niniejszy wynalazek dotyczy ponadto koniugatu, dimeru lub peptydu fuzyjnego zawierającego analog lub peptyd zdefiniowany w zastrzeżeniach. [003] Niniejszy wynalazek dotyczy ponadto kompozycji farmaceutycznej zawierającej analog zdefiniowany w zastrzeżeniach, peptyd zdefiniowany w zastrzeżeniach, koniugat, dimer lub peptyd fuzyjny zdefiniowany w zastrzeżeniach, lub ich kombinację, oraz dopuszczalny farmaceutycznie nośnik. [004] Niniejszy wynalazek dotyczy ponadto zestawu zawierającego kompozycję farmaceutyczną zdefiniowaną w zastrzeżeniach i urządzenie do podawania wspomnianej kompozycji farmaceutycznej pacjentowi, przy czym ewentualnie urządzenie obejmuje strzykawkę zawierającą kompozycję farmaceutyczną. [00] Niniejszy wynalazek dotyczy ponadto analogu zdefiniowanego w zastrzeżeniach lub peptydu zdefiniowanego w zastrzeżeniach do zastosowania do zmniejszania wzrostu masy lub indukowania utraty masy lub leczenia cukrzycy.
40 [006] Niniejszy wynalazek dotyczy ponadto zastosowania analogu zdefiniowanego w zastrzeżeniach lub peptydu zdefiniowanego w zastrzeżeniach do wytwarzania leku do zmniejszania wzrostu masy lub indukowania utraty masy lub leczenia cukrzycy. DEFINICJE [007] W opisie i zastrzeżeniach wynalazku stosuje się następującą terminologię zgodnie z definicjami podanymi poniżej. [008] Określenie około w znaczeniu stosowanym w niniejszym dokumencie oznacza więcej lub mniej od podanej wartości lub zakresu wartości o procent, jednak nie oznacza to, że przypisuje się jakąkolwiek wartość lub zakres wartości wyłącznie do takiej szerszej definicji. Każda wartość lub zakres wartości poprzedzone określeniem około ma także obejmować przykład wykonania podanej wartości bezwzględnej lub zakresu wartości. [009] W znaczeniu stosowanym w niniejszym dokumencie określenie dopuszczalny farmaceutycznie nośnik obejmuje dowolne spośród standardowych nośników farmaceutycznych, na przykład roztwór sodu soli fizjologicznej buforowanej fosforanem, wodę, emulsje, takie jak emulsję typu olej w wodzie lub woda w oleju, oraz różne rodzaje środków zwilżających. Określenie to obejmuje ponadto dowolne środki zarejestrowane przez urząd rejestracyjny rządu federalnego USA lub wymienione w Farmakopei USA do stosowania u zwierząt, w tym ludzi. [0060] W znaczeniu stosowanym w niniejszym dokumencie określenie dopuszczalna farmaceutycznie sól oznacza
41 sole związków zachowujących aktywność biologiczną związku macierzystego, które nie są niepożądane biologicznie lub z innych względów. Wiele spośród związków ujawnianych w niniejszym dokumencie może tworzyć sole z kwasami i(lub) zasadami dzięki obecności grup aminowych i(lub) karboksylowych bądź grup do nich podobnych. [0061] Dopuszczalne farmaceutycznie sole addycyjne z zasadami można otrzymać z zasad nieorganicznych i organicznych. Do soli pochodzących od zasad nieorganicznych należą, jedynie w charakterze przykładu, sole sodu, potasu, litu, amonu, wapnia i magnezu. Do soli pochodzących od zasad organicznych należą, ale bez ograniczania, sole amin pierwszorzędowych, drugorzędowych i trzeciorzędowych. [0062] Dopuszczalne farmaceutycznie sole addycyjne z kwasami można otrzymać z kwasów nieorganicznych i organicznych. Sole pochodzące od kwasów nieorganicznych obejmują sole z kwasem chlorowodorowym, kwasem bromowodorowym, kwasem siarkowym, kwasem azotowym, kwasem fosforowym i podobne. Do soli pochodzących od kwasów organicznych należą sole z kwasem octowym, kwasem propionowym, kwasem glikolowym, kwasem pirogronowym, kwasem szczawiowym, kwasem jabłkowym, kwasem malonowym, kwasem bursztynowym, kwasem maleinowym, kwasem fumarowym, kwasem winowym, kwasem cytrynowym, kwasem benzoesowym, kwasem cynamonowym, kwasem migdałowym, kwasem metanosulfonowym, kwasem etanosulfonowym, kwasem p-toluenosulfonowym, kwasem salicylowym i podobne. [0063] W znaczeniu stosowanym w niniejszym dokumencie
42 określenie leczenie obejmuje profilaktykę określonego stanu lub zaburzenia bądź złagodzenie objawów towarzyszących określonemu stanowi lub zaburzeniu i(lub) zapobieganie lub zniesienie wspomnianych objawów. Na przykład w znaczeniu stosowanym w niniejszym dokumencie określenie leczenie cukrzycy będzie oznaczać ogólnie zmianę poziomów glukozy we krwi w stronę poziomów prawidłowych i może obejmować zwiększenie lub zmniejszenie poziomów glukozy we krwi w zależności od konkretnej sytuacji. [0064] W znaczeniu stosowanym w niniejszym dokumencie ilość skuteczna lub ilość skuteczna terapeutycznie peptydu glukagonu oznacza nietoksyczną, ale wystarczającą ilość peptydu, aby uzyskać pożądane działanie. Na przykład jednym pożądanym działaniem jest zapobieganie lub leczenie hipoglikemii, mierzonej na przykład na podstawie wzrostu poziomu glukozy we krwi. Alternatywne pożądane działanie peptydów glukagonu według niniejszego ujawnienia będzie obejmować leczenie hiperglikemii, np. mierzonej na podstawie zmian poziomu glukozy we krwi w stronę wartości normalnych, lub indukowanie utraty masy /zapobieganie wzrostowi masy, np. mierzonych na podstawie zmniejszenia masy ciała, lub zapobieganie lub zmniejszanie wzrostu masy ciała bądź normalizacja dystrybucji tłuszczu w organizmie. Ilość która jest skuteczna będzie różnić się w zależności od osobnika, w zależności od wieku i stanu ogólnego osoby, sposobu podawania i podobnych. W związku z tym nie zawsze jest możliwe dokładne określenie ilości skutecznej. Jednak specjalista w dziedzinie może określić odpowiednią ilość skuteczną
43 w każdym pojedynczym przypadku na podstawie rutynowych doświadczeń. [006] Określenie pozajelitowy oznacza nie poprzez przewód pokarmowy, ale pewną inną drogą, na przykład podskórnie, domięśniowo, dordzeniowo lub dożylnie. [0066] W znaczeniu stosowanym w niniejszym dokumencie określenia oczyszczony i podobne określenia oznaczają wyodrębnienie cząsteczki lub związku w postaci zasadniczo niezawierającej zanieczyszczeń towarzyszących normalnie tej cząsteczce lub związkowi w środowisku natywnym lub naturalnym. W znaczeniu stosowanym w niniejszym dokumencie określenie oczyszczony nie oznacza czystości absolutnej; raczej ma ono oznaczać definicję względną. Określenie polipeptyd oczyszczony stosuje się w niniejszym dokumencie do opisu polipeptydu, który został oddzielony od innych związków, w tym nieograniczająco od cząsteczek kwasów nukleinowych, lipidów i węglowodanów. [0067] Określenie wyodrębniony wymaga, aby dany materiał został usunięty ze swojego pierwotnego środowiska (np. środowiska naturalnego, jeśli występuje naturalnie). Na przykład występujący naturalnie polinukleotyd obecny u zwierzęcia żywego nie jest wyodrębniony, ale ten sam polinukleotyd oddzielony od niektórych lub wszystkich materiałów towarzyszących w układzie naturalnym jest wyodrębniony. [0068] W znaczeniu stosowanym w niniejszym dokumencie określenie peptyd obejmuje sekwencję 3 lub więcej aminokwasów i zazwyczaj mniej niż 0 aminokwasów, przy czym aminokwasy stanowią aminokwasy występujące
44 naturalnie lub niewystępujące naturalnie. Aminokwasy niewystępujące naturalnie oznaczają aminokwasy, które nie występują naturalnie in vivo, ale które mimo tego można wprowadzić do struktur peptydowych opisywanych w niniejszym dokumencie. [0069] W znaczeniu stosowanym w niniejszym dokumencie określenia polipeptyd i białko są określeniami wykorzystywanymi zamiennie w znaczeniu polimeru aminokwasów, bez odniesienia do długości polimeru. Zazwyczaj polipeptydy i białka mają długość polimeru większą niż w przypadku peptydów. [0070] Peptyd glukagonu w znaczeniu stosowanym w niniejszym dokumencie obejmuje dowolny peptyd zawierający sekwencję aminokwasową według NR. ID. SEKW.: 1 lub dowolny analog sekwencji aminokwasowej według NR. ID. SEKW.: 1, w tym substytucje, addycje, delecje lub modyfikacje potranslacyjne aminokwasów (np. metylacja, acylowanie, alkilowanie, ubikwitynacja, wewnątrzcząsteczkowe wiązanie kowalencyjne, takie jak utworzenie mostka laktamowego, PEGylowanie i podobne) peptydu, przy czym analog stymuluje aktywność receptora glukagonu lub GLP-1 lub GIP, np. na podstawie pomiaru wytwarzania camp przy użyciu testu opisanego w przykładzie 16. [0071] Określenie agonista glukagonu dotyczy kompleksu zawierającego peptyd glukagonu, który stymuluje aktywność receptora glukagonu, np. na podstawie pomiaru wytwarzania camp przy użyciu testu opisanego w przykładzie 16. [0072] W znaczeniu stosowanym w niniejszym dokumencie modyfikacja aminokwasu oznacza substytucję, addycję
45 lub delecję aminokwasu i obejmuje substytucję lub addycję dowolnego spośród aminokwasów występujących powszechnie w białkach człowieka, a także aminokwasów nietypowych lub niewystępujących naturalnie. W całej treści zgłoszenia wszystkie odniesienia do określonego położenia aminokwasu za pomocą liczby (np. położenie 28) dotyczą tego aminokwasu w tym położeniu w glukagonie natywnym (NR. ID. SEKW.: 1) lub odpowiadającego położenia aminokwasu w dowolnych jego analogach. Na przykład odniesienie w niniejszym dokumencie do położenia 28 będzie oznaczać odpowiadające położenie 27 w przypadku analogu glukagonu, w którym pierwszy aminokwas według NR. ID. SEKW.: 1 został usunięty. Podobnie odniesienie w niniejszym dokumencie do położenia 28 będzie oznaczać odpowiadające położenie 29 w przypadku analogu glukagonu, w którym jeden aminokwas został dodany przed końcem N według NR. ID. SEKW.: 1. Aminokwasy nietypowe są dostępne w sprzedaży na przykład od Sigma-Aldrich (Milwaukee, WI), ChemPep Inc. (Miami, FL) i Genzyme Pharmaceuticals (Cambridge, MA). Aminokwasy nietypowe można zakupić u dostawców komercyjnych, zsyntetyzować de novo lub poddać modyfikacjom chemicznym bądź otrzymać pochodne z innych aminokwasów. [0073] W znaczeniu stosowanym w niniejszym dokumencie określenie glukagon natywny oznacza peptyd składający się z sekwencji według NR. ID. SEKW.: 1, określenie GIP natywny oznacza peptyd składający się z sekwencji według NR. ID. SEKW.: 4, a określenie GLP-1 natywny stanowi określenie ogólne oznaczające GLP-1(7-36) amid (składający się z sekwencji według NR. ID. SEKW.: 3),
46 GLP-1(7-37) kwas (składający się z sekwencji według NR. ID. SEKW.: 2) lub mieszaninę tych dwóch związków. W znaczeniu stosowanym w niniejszym dokumencie odniesienie ogólne do glukagonu lub GIP lub GLP-1 przy braku żadnego dodatkowego określenia ma oznaczać odpowiednio natywny glukagon lub natywny GIP lub natywny GLP-1. [0074] W znaczeniu stosowanym w niniejszym dokumencie substytucja aminokwasu oznacza podstawienie jednej reszty aminokwasowej inną resztą aminokwasową. [007] W znaczeniu stosowanym w niniejszym dokumencie określenie konserwatywna substytucja aminokwasu definiuje się w niniejszym dokumencie jako wymianę w obrębie jednej z następujących pięciu grup: I. Małe reszty alifatyczne, niepolarne lub nieco polarne: Ala, Ser, Thr, Pro, Gly; II. Reszty polarne naładowane ujemnie oraz ich amidy i estry: Asp, Asn, Glu, Gln, kwas cysteinowy i kwas homocysteinowy; III. Reszty polarne naładowane dodatnio: His, Arg, Lys; ornityna (Orn) IV. Duże niepolarne reszty alifatyczne: Met, Leu, Ile, Val, Cys, norleucyna (Nle), homocysteina V. Duże reszty aromatyczne: Phe, Tyr, Trp, acetylofenyloalanina [0076] W znaczeniu stosowanym w niniejszym dokumencie określenie ogólne łańcuch poli(glikolu etylenowego) lub łańcuch PEG oznacza mieszaniny polimerów kondensacyjnych tlenku etylenu i wody, w łańcuchu
47 rozgałęzionym lub prostym, przedstawione wzorem ogólnym H(OCH2CH2)nOH, w którym n wynosi co najmniej 9. Jeśli nie podano dodatkowej charakterystyki, określenie to ma obejmować polimery glikolu etylenowego mające średnią całkowitą masę cząsteczkową wybraną z zakresu 00 do daltonów. Terminy łańcuch poli(glikolu etylenowego) lub łańcuch PEG stosuje się w połączeniu z przedrostkiem liczbowym, aby określić ich przybliżoną średnią masę cząsteczkową. Na przykład PEG- 000 oznacza łańcuch poli(glikolu etylenowego) mający całkowitą średnią masę cząsteczkową około 000. [0077] W znaczeniu stosowanym w niniejszym dokumencie określenie pegylowany i podobne terminy oznaczają związek, który zmodyfikowano względem jego stanu natywnego przez połączenie z tym związkiem łańcucha poli(glikolu etylenowego). Pegylowany peptyd glukagonu stanowi peptyd glukagonu zawierający łańcuch PEG połączony kowalencyjnie z peptydem glukagonu. [0078] W znaczeniu stosowanym w niniejszym dokumencie odniesienie ogólne do peptydu ma obejmować peptydy zawierające zmodyfikowane końce aminowe i karboksylowe. Na przykład łańcuch aminokwasów zawierający grupę amidową zamiast końcowego kwasu karboksylowego ma obejmować sekwencję aminokwasową przedstawioną aminokwasami standardowymi. [0079] W znaczeniu stosowanym w niniejszym dokumencie łącznik stanowi wiązanie, cząsteczkę lub grupę cząsteczek wiążących ze sobą dwie odrębne jednostki. Łączniki mogą zapewniać optymalne oddzielenie dwóch jednostek lub mogą ponadto stanowić nietrwałe połączenie umożliwiające oddzielenie od siebie tych
48 dwóch jednostek. Do wiązań nietrwałych należą grupy rozszczepialne światłem, ugrupowania nietrwałe pod wpływem kwasów, ugrupowania nietrwałe pod wpływem zasad i grupy rozszczepialne enzymami. [0080] W znaczeniu stosowanym w niniejszym dokumencie dimer stanowi kompleks zawierający dwie podjednostki połączone ze sobą kowalencyjnie przez łącznik. Określenie dimer, stosowane bez żadnych kwalifikatorów, obejmuje zarówno homodimery, jak i heterodimery. Homodimer zawiera dwie identyczne podjednostki, a heterodimer zawiera dwie podjednostki różniące się od siebie, jednak te dwie podjednostki są zasadniczo podobne do siebie. [0081] W znaczeniu stosowanym w niniejszym dokumencie określenie aminokwas naładowany oznacza aminokwas zawierający łańcuch boczny naładowany ujemnie (tzn. poddany deprotonacji) lub naładowany dodatnio (tzn. protonowany) w roztworze wodnym w ph fizjologicznym. Na przykład do aminokwasów naładowanych ujemnie należą kwas asparaginowy, kwas glutaminowy, kwas cysteinowy, kwas homocysteinowy i kwas homoglutaminowy, a do aminokwasów naładowanych dodatnio należą arginina, lizyna i histydyna. Aminokwasy naładowane obejmują aminokwasy naładowane spośród aminokwasów występujących powszechnie w białkach człowieka, a także aminokwasy nietypowe lub niewystępujące naturalnie. [0082] W znaczeniu stosowanym w niniejszym dokumencie określenie aminokwas kwaśny oznacza aminokwas zawierający drugie ugrupowanie kwasowe, w tym na przykład grupę kwasu karboksylowego lub kwasu sulfonowego.
49 [0083] W znaczeniu stosowanym w niniejszym dokumencie określenie selektywność cząsteczki wobec pierwszego receptora względem receptora drugiego oznacza następujący stosunek: wartość EC0 cząsteczki wobec drugiego receptora podzielona przez wartość EC0 cząsteczki wobec receptora pierwszego. Na przykład cząsteczka mająca wartość EC0 wynoszącą 1 nm wobec pierwszego receptora i wartość EC0 wynoszącą 0 nm wobec drugiego receptora ma 0-krotnie większą selektywność wobec pierwszego receptora niż wobec drugiego receptora. [0084] W znaczeniu stosowanym w niniejszym dokumencie siła działania glukagonu cząsteczki oznacza stosunek wartości EC0 cząsteczki wobec receptora glukagonu podzielonej przez wartość EC0 natywnego glukagonu wobec receptora glukagonu. [008] W znaczeniu stosowanym w niniejszym dokumencie siła działania GIP cząsteczki oznacza stosunek wartości EC0 cząsteczki wobec receptora GIP podzielonej przez wartość EC0 natywnego GIP wobec receptora GIP. [0086] W znaczeniu stosowanym w niniejszym dokumencie siła działania GLP-1 cząsteczki oznacza stosunek wartości EC0 cząsteczki wobec receptora GLP-1 podzielonej przez wartość EC0 natywnego GLP-1 wobec receptora GLP-1. [0087] W znaczeniu stosowanym w niniejszym dokumencie określenie alkil oznacza liniowy lub rozgałęziony węglowodór zawierający podaną liczbę atomów węgla. Do przykładów alkilów należą grupy metylowa, etylowa i liniowa propylowa.
50 [0088] W znaczeniu stosowanym w niniejszym dokumencie określenie heteroalkil oznacza liniowy lub rozgałęziony węglowodór zawierający podaną liczbę atomów węgla i co najmniej jeden heteroatom w szkielecie struktury. Do odpowiednich heteroatomów na potrzeby niniejszego dokumentu należą między innymi N, S i O. [0089] W znaczeniu stosowanym w niniejszym dokumencie określenie cykloalkil oznacza cykliczną grupę węglowodorową zawierającą podaną liczbę atomów węgla, np. cyklopropyl, cyklobutyl, cykloheksyl i cyklopentyl. [0090] W znaczeniu stosowanym w niniejszym dokumencie określenie heterocykl oznacza cykliczną grupę węglowodorową zawierającą podaną liczbę atomów węgla oraz jeden do trzech heteroatomów wybranych niezależnie z grupy składającej się z tlenu, azotu i siarki. Do nieograniczających przykładów grup heterocykloalkilowych należą piperydyna, tetrahydrofuran, tetrahydropiran, dihydrofuran, morfolina, tiofen i podobne. [0091] W znaczeniu stosowanym w niniejszym dokumencie określenie aryl oznacza monocykliczną lub policykliczną grupę aromatyczną, korzystnie monocykliczną lub bicykliczną grupę aromatyczną, np. fenylową lub naftylową, zawierającą podaną liczbę atomów węgla. Jeśli nie podano inaczej, grupa arylowa może być niepodstawiona lub podstawiona. [0092] W znaczeniu stosowanym w niniejszym dokumencie określenie heteroaryl oznacza monocykliczną lub policykliczną grupę aromatyczną zawierającą podaną liczbę atomów węgla oraz co najmniej jeden heteroatom
51 wybrany z grupy składającej się z tlenu, azotu i siarki. Jeśli nie podano inaczej, grupa arylowa może być niepodstawiona lub podstawiona. PRZYKŁADY WYKONANIA [0093] Modyfikacje ujawniane w niniejszym dokumencie umożliwiają manipulację glukagonem (NR. ID. SEKW.: 1) w celu utworzenia peptydów glukagonu, które wykazują zwiększoną aktywność GIP, aktywność glukagonu i(lub) aktywność GLP-1. Inne modyfikacje ujawniane w niniejszym dokumencie wydłużają okres półtrwania, zwiększają rozpuszczalność lub zwiększają stabilność otrzymywanego peptydu. Jeszcze inne modyfikacje ujawniane w niniejszym dokumencie nie mają wpływu na aktywność lub można je przeprowadzić nie niszcząc aktywności pożądanej lub pożądanych. Dowolną spośród kombinacji służących temu samemu celowi (np. zwiększenie aktywności GIP) można zastosować pojedynczo lub w kombinacji. Dowolną spośród pojedynczych kombinacji lub ich zestawów nadających zwiększone właściwości można zastosować pojedynczo lub w kombinacji, np. zwiększoną aktywność GIP i(lub) GLP-1 można połączyć ze zwiększonym okresem półtrwania. [0094] W pewnych przykładach wykonania peptyd glukagonu może zawierać ogółem 1, do 2, do 3, do 4, do, do 6, do 7, do 8, do 9 lub do modyfikacji aminokwasów względem sekwencji natywnego glukagonu. W niektórych przykładach wykonania takie peptydy glukagonu zachowują co najmniej 22, 23, 24, 2, 26, 27 lub 28 spośród aminokwasów występujących naturalnie w odpowiadających położeniach natywnego glukagonu (np. zawierają 1-7, 1- lub 1-3 modyfikacji względem glukagonu występującego
52 naturalnie). W powiązanych przykładach wykonania 1, 2, 3, 4,, 6 lub więcej spośród modyfikacji aminokwasów mogą stanowić substytucje, addycje lub delecje niekonserwatywne. W powiązanych przykładach wykonania 1, 2, 3, 4,, 6 lub więcej spośród modyfikacji aminokwasów mogą stanowić substytucje konserwatywne. W niektórych przykładach wykonania 1, 2, 3, 4 lub substytucji niekonserwatywnych wprowadza się w dowolnym spośród położeń 2,, 7,, 11, 12, 13, 14, 17, 18, 19,, 21, 24, 27, 28 lub 29 i do dodatkowych substytucji konserwatywnych wprowadza się w dowolnym spośród tych położeń. W niektórych przykładach wykonania 1, 2, lub 3 modyfikacje aminokwasów wprowadza się w obrębie aminokwasów w położeniach 1-16 oraz 1, 2 lub 3 modyfikacje aminokwasów wprowadza się w obrębie aminokwasów w położeniach Modyfikacje mające wpływ na aktywność GIP [009] Zwiększoną aktywność wobec receptora GIP uzyskuje się dzięki modyfikacji aminokwasu w położeniu 1. Na przykład His w położeniu 1 jest podstawiona dużym aminokwasem aromatycznym, ewentualnie Tyr, Phe, Trp, amino-phe, nitro-phe, chloro-phe, sulfo-phe, 4- pirydylo-ala, metylo-tyr lub 3-amino-Tyr. Nieoczekiwanie kombinacja Tyr w położeniu 1 ze stabilizacją helisy alfa w obrębie regionu odpowiadającego aminokwasom zapewnia peptyd glukagonu powodujący aktywację receptora GIP, a także receptora GLP-1 i receptora glukagonu. Stabilizację struktury helisy alfa można uzyskać np. dzięki utworzeniu kowalencyjnego lub niekowalencyjnego mostka wewnątrzcząsteczkowego lub substytucji i(lub) insercji
53 aminokwasów w obrębie położeń z użyciem aminokwasu stabilizującego helisę alfa (np. aminokwasu α,α-dipodstawionego). [0096] Zwiększoną aktywność wobec receptora GIP uzyskuje się ponadto dzięki modyfikacji aminokwasów w położeniach 27 i(lub) 28 oraz ewentualnie w położeniu 29. Na przykład Met w położeniu 27 jest podstawiona przez duży aminokwas alifatyczny, ewentualnie Leu, Asn w położeniu 28 jest podstawiona przez mały aminokwas alifatyczny, ewentualnie Ala, a Thr w położeniu 29 jest podstawiona przez mały aminokwas alifatyczny, ewentualnie Gly. Substytucja przez LAG w położeniach zapewnia zwiększoną aktywność GIP względem natywnej sekwencji MNT w tych położeniach. [0097] Zwiększoną aktywność wobec receptora GIP uzyskuje się także dzięki modyfikacji aminokwasu w położeniu 12. Na przykład położenie 12 jest podstawione przez duży alifatyczny aminokwas niepolarny, ewentualnie Ile. [0098] Zwiększoną aktywność wobec receptora GIP uzyskuje się także dzięki modyfikacji aminokwasu w położeniach 17 i(lub) 18. Na przykład położenie 17 jest podstawione przez resztę polarną, ewentualnie Gln, a położenie 18 jest podstawione przez mały aminokwas alifatyczny, ewentualnie Ala. Substytucja przez QA w położeniach 17 i 18 zapewnia zwiększoną aktywność GIP względem sekwencji natywnego RR w tych położeniach. [0099] Dowolną spośród modyfikacji opisanych powyżej, które zwiększają aktywność wobec receptora GIP, można zastosować pojedynczo lub w kombinacji. Kombinacje modyfikacji zwiększających aktywność wobec receptora
54 GIP zapewniają zasadniczo większą aktywność GIP niż dowolne spośród takich modyfikacji rozpatrywane oddzielnie. Modyfikacje mające wpływ na aktywność glukagonu [00] W niektórych przykładach wykonania dostarcza się analogi glukagonu mające zwiększoną siłę działania i ewentualnie zwiększoną rozpuszczalność i stabilność. W jednym przykładzie wykonania zwiększoną siłę działania glukagonu uzyskuje się dzięki modyfikacji aminokwasu w położeniu 16 natywnego glukagonu (NR. ID. SEKW.: 1). W ramach nieograniczającego przykładu taką zwiększoną siłę działania można uzyskać dzięki substytucji naturalnie występującej seryny w położeniu 16 przez kwas glutaminowy lub przez inny aminokwas zawierający naładowany ujemnie, który zawiera łańcuch boczny o długości 4 atomów, lub ewentualnie przez dowolny spośród glutaminy, kwasu homoglutaminowego lub kwasu homocysteinowego lub aminokwasu naładowanego zawierającego łańcuch boczny zawierający co najmniej jeden heteroatom (np. N, O, S, P) i o długości łańcucha bocznego około 4 (lub 3-) atomów. W niektórych przykładach wykonania peptyd glukagonu zachowuje swoją pierwotną selektywność wobec receptora glukagonu względem receptorów GLP-1. [01] Aktywność wobec receptora glukagonu można zmniejszyć dzięki modyfikacji aminokwasu w położeniu 3, np. substytucji naturalnie występującej glutaminy w położeniu 3 przez aminokwas kwaśny, zasadowy lub hydrofobowy. Na przykład substytucja w położeniu 3 przez kwas glutaminowy, ornitynę lub norleucynę znacznie zmniejsza lub znosi aktywność wobec receptora
55 glukagonu. [02] Utrzymaną lub zwiększoną aktywność wobec receptora glukagonu można uzyskać dzięki modyfikacji Gln w położeniu 3 przez analog glutaminy. Na przykład peptyd glukagonu zawierający analog glutaminy w położeniu 3 może wykazywać około %, około %, około %, około 0% lub około 8% lub większą aktywność natywnego glukagonu (NR. ID. SEKW.: 1) wobec receptora glukagonu. W niektórych przykładach wykonania peptyd glukagonu zawierający analog glutaminy w położeniu 3 może wykazywać około %, około 0%, około 7%, około 0%, około 0% lub około 00% lub większą aktywność odpowiadającego peptydu glukagonu mającego tę samą sekwencję aminokwasową co peptyd zawierający analog glutaminy, oprócz zmodyfikowanego aminokwasu w położeniu 3 (np. NR. ID. SEKW.: lub NR. ID. SEKW.: 21) wobec receptora glukagonu. W niektórych przykładach wykonania peptyd glukagonu zawierający analog glutaminy w położeniu 3 wykazuje zwiększoną aktywność wobec receptora glukagonu, ale ta zwiększona aktywność stanowi nie więcej niż 00%, 000%, 0 000% lub % aktywności natywnego glukagonu lub odpowiadającego peptydu glukagonu zawierającego tę samą sekwencję aminokwasową co peptyd zawierający analog glutaminy, oprócz zmodyfikowanego aminokwasu w położeniu 3. [03] W niektórych przykładach wykonania analog glutaminy stanowi występujący naturalnie lub niewystępujący naturalnie aminokwas zawierający łańcuch boczny według struktury I, II lub III:
56 Struktura I Struktura II Struktura III w których R 1 oznacza C0-3 alkil lub C0-3 heteroalkil; R 2 oznacza NHR 4 lub C1-3 alkil; R 3 oznacza C1-3 alkil; R 4 oznacza H lub C1-3 alkil; X oznacza NH, O lub S, a Y oznacza NHR 4, SR 3 lub OR 3. W niektórych przykładach wykonania X oznacza NH lub Y oznacza NHR 4. W niektórych przykładach wykonania R 1 oznacza C0-2 alkil lub C1 1 2 heteroalkil. W niektórych przykładach wykonania R 2 oznacza NHR 4 lub C1 alkil. W niektórych przykładach wykonania R 4 oznacza H lub C 1 alkil. W pewnych przykładach wykonania dostarcza się aminokwas zawierający łańcuch boczny według struktury I, w której R 1 oznacza CH2-S, X oznacza NH, a R 2 oznacza CH3 (acetamidometylocysteina, C(Acm)); R 1 oznacza CH2, X oznacza NH, a R 2 oznacza CH3 (kwas acetylodiaminobutanowy, Dab(Ac)); R 1 oznacza C0 alkil, X oznacza NH, R 2 oznacza NHR 4, a R 4 oznacza H (kwas karbamoilodiaminopropanowy, Dap(mocznik)); lub R 1 oznacza CH2-CH2, X oznacza NH, a R 2 oznacza CH3 (acetyloornityna, Orn(Ac)). W pewnych przykładach
57 wykonania dostarcza się aminokwas zawierający łańcuch boczny według struktury II, w której R 1 oznacza CH2, Y oznacza NHR 4, a R 4 oznacza CH3 (metyloglutamina, Q(Me)); W pewnych przykładach wykonania dostarcza się aminokwas zawierający łańcuch boczny według struktury III, w której R 1 oznacza CH2, a R 4 oznacza H (sulfotlenek metioniny, M(O)). W konkretnych przykładach wykonania aminokwas w położeniu 3 jest podstawiony przez Dab(Ac). Na przykład agoniści glukagonu mogą zawierać sekwencję aminokwasową według którejkolwiek z NR. ID. SEKW.: ,, 21 i Modyfikacje mające wpływ na aktywność GLP-1 [04] Zwiększoną aktywność wobec receptora GLP-1 uzyskuje się dzięki zastąpieniu kwasu karboksylowego w aminokwasie C-końcowym grupą o ładunku obojętnym, taką jak amid lub ester. [0] Zwiększoną aktywność wobec receptora GLP-1 uzyskuje się także dzięki stabilizacji struktury helisy alfa części C-końcowej glukagonu (w obrębie aminokwasów 12-29), np. przez utworzenie mostka wewnątrzcząsteczkowego między łańcuchami bocznymi dwóch aminokwasów lub substytucję i(lub) insercję aminokwasów w obrębie położeń z użyciem aminokwasu stabilizującego helisę alfa (np. aminokwasu α,αdipodstawionego). Łańcuchy boczne tych aminokwasów można połączyć ze sobą poprzez wiązanie wodorowe lub oddziaływania jonowe, na przykład przez utworzenie mostków solnych lub wiązań kowalencyjnych. W niektórych przykładach wykonania mostek tworzy się między aminokwasami rozdzielonymi trzema rozdzielającymi aminokwasami, tzn. aminokwasem w położeniu i i
58 aminokwasem w położeniu i+4, przy czym i jest dowolną liczbą całkowitą od 12 do 2 (np. 12, 13, 14, 1, 16, 17, 18, 19,, 21, 22, 23, 24 lub 2). W pewnych przykładach wykonania łańcuchy boczne par aminokwasów 12 i 16, 13 i 17, 16 i, 17 i 21, i 24 lub 24 i 28 (par aminokwasów, w których i = 12, 16, lub 24) są połączone ze sobą i w ten sposób stabilizują helisę alfa glukagonu. [06] W innych przykładach wykonania mostek tworzy się między aminokwasami rozdzielonymi dwoma rozdzielającymi aminokwasami, tzn. aminokwasem w położeniu j i aminokwasem w położeniu j+3, przy czym j jest dowolną liczbą całkowitą od 12 do 26 (np. 12, 13, 14, 1, 16, 17, 18, 19,, 21, 22, 23, 24, 2 lub 26). W pewnych przykładach wykonania j wynosi 17. W dodatkowych przykładach wykonania mostek tworzy się między aminokwasami rozdzielonymi sześcioma rozdzielającymi aminokwasami, tzn. aminokwasem w położeniu k i aminokwasem w położeniu k+7, przy czym k jest dowolną liczbą całkowitą od 12 do 22 (np. 12, 13, 14, 1, 16, 17, 18, 19,, 21 lub 22). W jednym przykładzie wykonania k wynosi 17. [07] W niektórych przykładach wykonania mostek lub łącznik ma długość około 8 (lub około 7-9) atomów, w szczególności, jeśli mostek występuje między położeniami i oraz i+4. W niektórych przykładach wykonania mostek lub łącznik ma długość około 6 (lub około -7) atomów, w szczególności, jeśli mostek występuje między położeniami j i j+3. [08] W niektórych przykładach wykonania mostki wewnątrzcząsteczkowe tworzą się przez (a) substytucję
59 naturalnie występującej seryny w położeniu 16 przez kwas glutaminowy lub przez inny aminokwas naładowany ujemnie, który zawiera łańcuch boczny o długości 4 atomów lub ewentualnie przez dowolny spośród glutaminy, kwasu homoglutaminowego lub kwasu homocysteinowego lub aminokwasu naładowanego zawierającego łańcuch boczny zawierający co najmniej jeden heteroatom (np. N, O, S, P) i o długości łańcucha bocznego około 4 (lub 3-) atomów oraz (b) substytucję naturalnie występującej glutaminy w położeniu przez inny aminokwas hydrofilowy zawierający łańcuch boczny naładowany lub mający zdolność do tworzenia wiązania wodorowego i o długości co najmniej około (lub około 4-6) atomów, na przykład lizynę, cytrulinę, argininę lub ornitynę. Łańcuchy boczne takich aminokwasów w położeniach 16 i mogą tworzyć mostek solny lub być połączone kowalencyjnie. [09] W jednym przykładzie wykonania te dwa aminokwasy są połączone ze sobą tworząc pierścień laktamowy. Wielkość pierścienia laktamowego może się zmieniać w zależności od długości łańcuchów bocznych aminokwasów, i w jednym przykładzie wykonania laktam tworzy się przez połączenie łańcuchów bocznych aminokwasu lizyny z łańcuchem bocznym kwasu glutaminowego. Kierunek wiązania amidowego w pierścieniu laktamowym można odwrócić (np. można utworzyć pierścień laktamowy między łańcuchami bocznymi Lys12 i Glu16 lub ewentualnie między Glu 12 i Lys16). [01] W niektórych przykładach wykonania stabilizację struktury helisy alfa w części C-końcowej peptydu glukagonu uzyskuje się dzięki utworzeniu mostka
60 wewnątrzcząsteczkowego innego niż mostek laktamowy. Na przykład do odpowiednich metod łączenia wiązaniami kowalencyjnymi należy dowolna jedna lub większa liczba spośród metatezy olefin, cyklizacji z użyciem lantioniny, utworzenia mostka disiarczkowego lub mostka zawierającego zmodyfikowaną siarkę, zastosowania α,ωdiaminoalkanowych struktur wiążących, utworzenia mostków metal-atom, i inne środki cyklizacji peptydów są stosowane do stabilizacji helisy alfa. [0111] Siłę działania wobec receptora GLP-1 można dodatkowo zwiększyć dzięki substytucji alaniny zamiast natywnej argininy w położeniu 18. [0112] Dowolną spośród modyfikacji opisanych powyżej, które zwiększają aktywność wobec receptora GLP-1, można zastosować pojedynczo lub w kombinacji. Kombinacje modyfikacji zwiększających aktywność wobec receptora GLP-1 zapewniają zasadniczo większą aktywność GLP-1 niż dowolne spośród takich modyfikacji rozpatrywane oddzielnie. Na przykład wynalazek dostarcza peptydy glukagonu zawierające modyfikacje w położeniu 16, w położeniu i na C-końcowej grupie kwasu karboksylowego, ewentualnie z wiązaniem kowalencyjnym między aminokwasami w położeniach 16 i ; peptydy glukagonu zawierające modyfikacje w położeniu 16 i na C-końcowej grupie kwasu karboksylowego; peptydy glukagonu zawierające modyfikacje w położeniach 16 i, ewentualnie z wiązaniem kowalencyjnym między aminokwasami w położeniach 16 i oraz peptydy glukagonu zawierające modyfikacje w położeniu i na C-końcowej grupie kwasu karboksylowego. [0113] Aktywność GLP-1 można zmniejszyć dzięki
61 60 1 zastosowaniu (i) C-końcowej grupy alfa-karboksylanowej, (ii) substytucji Thr w położeniu 7 przez aminokwas niezawierający grupy hydroksylowej, np. Abu lub Ile, (iii) delecji aminokwasu lub aminokwasów C-końcowych wobec aminokwasu w położeniu 27 lub 28 (np. delecję aminokwasu w położeniu 28, delecję aminokwasu w położeniach 28 i 29), dzięki czemu otrzymuje się peptyd o długości 27 lub 28 aminokwasów, lub (iv) ich kombinacji. Modyfikacje mające wpływ na aktywność wobec każdego spośród receptorów glukagonu, GLP-1 i GIP [0114] Zwiększoną aktywność wobec każdego spośród receptora glukagonu, receptora GLP-1 i receptora GIP zapewnia się dzięki (i) substytucji aminokwasu Ser w położeniu 16 przez aminokwas o wzorze IV: [wzór IV], w którym n wynosi 1 do 16, lub 1 do, lub 1 do 7, lub 1 do 6, lub 2 do 6, lub 2, lub 3, lub 4, lub, przy czym każdy spośród R1 i R2 jest niezależnie wybrany z grupy składającej się z: H, C1-C18 alkil, (C1-C18 alkilo)oh, (C1-C18 alkilo)nh2, (C1-C18 alkilo)sh, (C0-C4 alkilo)(c3-c6)cykloalkil, (C0-C4 alkilo)(c2-c 2 heterocykl), (C0-C4 alkilo)(c6-c arylo)r7 i (C1-C4 alkilo)(c3-c9 heteroaryl), przy czym R7 oznacza H lub OH, a łańcuch boczny aminokwasu o wzorze IV zawiera wolną grupę aminową i (ii) substytucji aminokwasu Gln w
62 położeniu aminokwasem alfa,alfa-dipodstawionym, np. AIB. W niektórych przykładach wykonania aminokwas w położeniu 16 stanowi Orn, Dab, Lys lub homolys, a aminokwas w położeniu stanowi AIB. W konkretnych przykładach wykonania aminokwas w położeniu 16 stanowi Lys, a aminokwas w położeniu stanowi AIB. [011] Aktywność wobec każdego spośród receptora glukagonu, receptora GLP-1 i receptora glukagonu analogu zawierającego aminokwas o wzorze IV w położeniu 16 i aminokwas alfa,alfa-dipodstawiony w położeniu można dodatkowo zwiększyć w wyniku zwiększenia długości peptydu, np. przez fuzję z peptydem stanowiącym przedłużenie C-końcowe, np. o długości około 1-21, około 9 do 21, około 6-18, około 9-12, lub około, lub 11 aminokwasów. W niektórych przykładach wykonania koniec C jest przedłużony w wyniku fuzji z GPSSGAPPPS (NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.: 96), przy czym X stanowi Gly lub mały aminokwas alifatyczny lub niepolarny lub nieco polarny. W alternatywnych przykładach wykonania koniec C jest przedłużony w wyniku fuzji z GPSSGAPPPS (NR. ID. SEKW.: 9) i 1-11 aminokwasów jest poddanych fuzji z końcem C GPSSGAPPPS (NR. ID. SEKW.: 9). Na przykład przedłużenie C-końcowe analogu może zawierać GPSSGAPPPS (NR. ID. SEKW.: 9), a następnie 1, 2, 3, 4,, 6, 7, 8, 9, lub 11 dodatkowych aminokwasów na końcu C według NR. ID. SEKW.: 9. Tych 1-11 dodatkowych aminokwasów może na przykład stanowić mały aminokwas alifatyczny, taki jak Ala. W odniesieniu do tego przedłużenie C-końcowe może na przykład zawierać sekwencję aminokwasową GPSSGAPPPSAm, w której m wynosi
63 do 11. [0116] Zwiększoną aktywność wobec każdego spośród receptorów glukagonu, GLP-1 i GIP analogu o aktywności GIP na bazie glukagonu, w tym analogu zawierającego aminokwas o wzorze IV w położeniu 16 i aminokwas alfa,alfa-dipodstawiony w położeniu, można ponadto uzyskać w wyniku acylowania lub alkilowania aminokwasu znajdującego się w przedłużeniu na końcu C lub aminokwasu C-końcowego (np. aminokwasu dodawanego na końcu C przedłużenia C-końcowego). Acylowanie lub alkilowanie może dotyczyć aminokwasu znajdującego się na przykład w dowolnym spośród położeń 30, 31, 32, 33, 34, 3, 36, 37, 38, 39, 40, 41, 42, 43, 44, 4, 46, 47, 48, 49 i 0 analogu z przedłużeniem C-końcowym. W niektórych przykładach wykonania aminokwas ulegający acylowaniu lub alkilowaniu znajduje się w położeniu 37, 38, 39, 40, 41, 42 lub 43 analogu z przedłużeniem C- końcowym. W niektórych przykładach wykonania aminokwas ulegający acylowaniu lub alkilowaniu stanowi aminokwas o wzorze I, II lub III, np. Lys dołączoną do grupy acylowej lub alkilowej, np. C-C22. W niektórych przykładach wykonania Lys znajduje się po stronie C- końcowej wobec przedłużenia C-końcowego składającego się z NR. ID. SEKW.: 9, w taki sposób, że Lys, Dab, Orn lub homolys znajduje się w położeniu 40 analogu. Ewentualnie peptydy z przedłużeniem C-końcowym są także pegylowane, np. w dowolnym z położeń opisywanych w niniejszym dokumencie (np. położenie 24). [0117] Zwiększenie aktywności wobec każdego spośród receptorów glukagonu, GLP-1 i GIP analogu o aktywności GIP na bazie glukagonu można ponadto uzyskać w wyniku
64 acylowania lub alkilowania aminokwasu przez wstawkę (np. aminokwas, dipeptyd, tripeptyd, hydrofilową wstawkę dwufunkcyjną, hydrofobową wstawkę dwufunkcyjną). W niektórych przykładach wykonania analog o aktywności GIP będący pochodną glukagonu zawiera grupę acylową lub alkilową poprzez wstawkę, która jest dołączona do łańcucha bocznego aminokwasu w położeniu analogu. W innych przykładach wykonania analog zawiera przedłużenie C-końcowe o długości 1 do 21 aminokwasów (np. przedłużenie zawierające sekwencję aminokwasową według NR. ID. SEKW.: 9 lub 96) po stronie C-końcowej aminokwasu w położeniu 29, a wstawka, która jest połączona kowalencyjnie z grupą acylową lub alkilową, jest dołączona do aminokwasu w przedłużeniu w położeniu odpowiadającym jednemu z położeń względem NR. ID. SEKW.: 1. W konkretnych przykładach wykonania wstawka jest dołączona do aminokwasu w położeniu 40 względem NR. ID. SEKW.: 1. W niektórych przykładach wykonania wstawka ma długość 3 do atomów. W konkretnych aspektach całkowita długość wstawki oraz grupy acylowej lub alkilowej wynosi około 14 do około 28 atomów. Na przykład wstawka może stanowić aminokwas, w tym między innym dowolny spośród opisywanych w niniejszym dokumencie. Ponadto na przykład wstawka może stanowić dipeptyd lub tripeptyd zawierający aminokwasy opisywane w niniejszym dokumencie. Wstawka w konkretnych aspektach stanowi jeden z następujących dipeptydów: Ala-Ala, βala-βala lub γglu-γglu. W niniejszym dokumencie opisano ponadto dodatkowe odpowiednie wstawki w celu zwiększenia aktywności wobec jednego lub większej liczby spośród
65 receptorów glukagonu, GLP-1 i GIP. Modyfikacje zwiększające odporność na DPP-IV [0118] Modyfikacje w położeniu 1 i(lub) 2 mogą zwiększać odporność peptydu na trawienie przez dipeptydylopeptydazę IV (DPP IV). Na przykład aminokwas w położeniu 2 może zostać podstawiony przez D-serynę, D-alaninę, walinę, glicynę, N-metyloserynę, N- metyloalaninę lub kwas aminoizomasłowy. W niektórych przykładach wykonania aminokwas w położeniu 1 może zostać podstawiony przez D-histydynę, desaminohistydynę, hydroksylohistydynę, acetylohistydynę, homohistydynę, N-metylohistydynę, alfa-metylohistydynę, kwas imidazolooctowy lub kwas alfa,alfa-dimetyloimidazolooctowy (DMIA). [0119] Stwierdzono, że modyfikacje w położeniu 2 (np. AIB w położeniu 2) i w niektórych przypadkach modyfikacje w położeniu 1 (np. DMIA w położeniu 1) mogą zmniejszać aktywność glukagonu, niekiedy znacząco; nieoczekiwanie to zmniejszenie aktywności glukagonu można przywrócić dzięki stabilizacji struktury helisy alfa części C-końcowej glukagonu (wokół aminokwasów 12-29), np. przez utworzenie wiązania kowalencyjnego między łańcuchami bocznymi dwóch aminokwasów zgodnie z opisem w niniejszym dokumencie. W niektórych przykładach wykonania wiązanie kowalencyjne znajduje się między aminokwasami w położeniach i i i+4 lub w położeniach j i j+3, np. między położeniami 12 i 16, 16 i, i 24, 24 i 28 lub 17 i. W pewnych przykładach wykonania to wiązanie kowalencyjne stanowi mostek laktamowy między kwasem glutaminowym w położeniu 16 a lizyną w położeniu. W niektórych przykładach
66 wykonania to wiązanie kowalencyjne stanowi mostek wewnątrzcząsteczkowy inny niż mostek laktamowy. Na przykład do odpowiednich metod łączenia wiązaniami kowalencyjnymi (tzn. sposobów tworzenia kowalencyjnego mostka wewnątrzcząsteczkowego) należy dowolna jedna lub większa liczba spośród metatezy olefin, cyklizacji z użyciem lantioniny, tworzenia mostka disiarczkowego lub zawierającego zmodyfikowaną siarkę, zastosowania α,ωdiaminoalkanowych struktur wiążących, tworzenia mostków metal-atom oraz innych sposobów cyklizacji peptydów. Modyfikacje zmniejszające degradację [01] W jeszcze innych pewnych przykładach wykonania dowolny spośród peptydów glukagonu można dodatkowo zmodyfikować, aby zwiększyć stabilność przez modyfikację aminokwasu w położeniu 1 i(lub) 16 według NR. ID. SEKW.: 1 w celu zmniejszenia degradacji peptydu z upływem czasu, w szczególności w buforach kwaśnych lub alkalicznych. Takie modyfikacje zmniejszają trawienie wiązania peptydowego Asp1-Ser16. W pewnych przykładach wykonania modyfikacja aminokwasu w położeniu 1 stanowi delecję lub substytucję Asp przez kwas glutaminowy, kwas homoglutaminowy, kwas cysteinowy lub kwas homocysteinowy. W innych niektórych przykładach wykonania modyfikacja aminokwasu w położeniu 16 stanowi delecję lub substytucję Ser przez Thr lub AIB. W innych niektórych przykładach wykonania Ser w położeniu 16 jest podstawiona przez kwas glutaminowy lub przez inny aminokwas naładowany ujemnie, który zawiera łańcuch boczny o długości 4 atomów lub ewentualnie przez dowolny spośród glutaminy, kwasu homoglutaminowego lub kwasu homocysteinowego.
67 [0121] W niektórych przykładach wykonania reszta metioniny występująca w położeniu 27 peptydu natywnego jest zmodyfikowana, np. przez delecję lub substytucję. Takie modyfikacje mogą zapobiegać degradacji oksydacyjnej peptydu. W niektórych przykładach wykonania Met w położeniu 27 jest podstawiona przez leucynę, izoleucynę lub norleucynę. W niektórych konkretnych przykładach wykonania Met w położeniu 27 jest podstawiona przez leucynę lub norleucynę. [0122] W niektórych przykładach wykonania Gln w położeniu i(lub) 24 jest zmodyfikowana, np. przez delecję lub substytucję. Takie modyfikacje mogą zmniejszyć degradację następującą w wyniku deamidacji Gln. W niektórych przykładach wykonania Gln w położeniu i(lub) 24 jest podstawiona przez Ala lub AIB. W niektórych przykładach wykonania Gln w położeniu i(lub) 24 jest podstawiona przez Lys, Arg, Orn lub cytrulinę. [0123] W niektórych przykładach wykonania Asp w położeniu 21 zostaje zmodyfikowana, np. przez delecję lub substytucję. Takie modyfikacje mogą zmniejszyć degradację następującą w wyniku dehydratacji Asp i utworzenia cyklicznego sukcynoimidowego produktu pośredniego, po czym następuje izomeryzacja do izoasparaginianu. W niektórych przykładach wykonania położenie 21 jest podstawione przez Glu, kwas homoglutaminowy lub kwas homocysteinowy. W pewnych konkretnych przykładach wykonania położenie 21 jest podstawione przez Glu. Inne modyfikacje [0124] Niektóre położenia w natywnym peptydzie
68 glukagonu można modyfikować utrzymując co najmniej niektóre spośród aktywności peptydu macierzystego. W związku z tym zgłaszający przewidują, że jeden lub większa liczba spośród aminokwasów znajdujących się w położeniach 2,,, 11, 12, 13, 14, 17, 18, 19,, 21, 24, 27, 28 lub 29 może być podstawionych przez aminokwas inny niż występujący w naturalnym peptydzie glukagonu i nadal można utrzymać aktywność wobec receptora glukagonu. [012] W niektórych przykładach wykonania położenie 18 jest podstawione przez aminokwas wybrany z grupy składającej się z Ala, Ser lub Thr. W niektórych przykładach wykonania aminokwas w położeniu jest podstawiony przez Ser, Thr, Lys, Arg, Orn, cytrulinę lub AIB. W niektórych przykładach wykonania położenie 21 jest podstawione przez Glu, kwas homoglutaminowy lub kwas homocysteinowy. W niektórych przykładach wykonania peptyd glukagonu zawiera 1 do modyfikacji aminokwasów wybranych spośród położeń 16, 17, 18,, 21, 23, 24, 27, 28 i 29. W pewnych przykładach wykonania modyfikacje stanowią substytucje jednego lub większej liczby aminokwasów wybranych z grupy składającej się z Gln17, Ala18, Glu21, Ile23, Ala24, Val27 i Gly29. W niektórych przykładach wykonania 1 do 2 aminokwasów wybranych spośród położeń różni się od peptydu macierzystego. W innych przykładach wykonania 1 do 2 aminokwasów wybranych spośród położeń różni się od peptydu macierzystego. W jeszcze innych przykładach wykonania modyfikacje stanowią Gln17, Ala18, Glu21, Ile23 i Ala24. [0126] W niektórych przykładach wykonania jeden lub
69 większą liczbę aminokwasów dodaje się do końca karboksylowego peptydu glukagonu. Aminokwas ten wybiera się zazwyczaj jako jeden z aminokwasów podstawowych i w niektórych przykładach wykonania aminokwas ten zawiera grupę amidową zamiast kwasu karboksylowego w natywnym aminokwasie. W pewnych przykładach wykonania dodawany aminokwas jest wybrany z grupy składającej się z kwasu glutaminowego i kwasu asparaginowego oraz glicyny. [0127] Inne modyfikacje, które nie niszczą aktywności, obejmują W lub R. [0128] W niektórych przykładach wykonania peptydy glukagonu ujawniane w niniejszym dokumencie ulegają modyfikacji w wyniku skrócenia końca C o jedną lub dwie reszty aminokwasowe przy utrzymaniu podobnej aktywności i siły działania wobec receptorów glukagonu, GLP-1 i(lub) GIP. W odniesieniu do tego aminokwas w położeniu 29 i(lub) 28 może ulec delecji. Stabilizacja struktury helisy alfa [0129] Stabilizacja struktury helisy alfa w części C- końcowej peptydu glukagonu (wokół aminokwasów 12-29) zapewnia zwiększoną aktywność GLP-1 i(lub) GIP oraz przywróca aktywność glukagonu, która została zmniejszona wskutek modyfikacji aminokwasów w położeniach 1 i(lub) 2. Stabilizację struktury helisy alfa można uzyskać np. dzięki utworzeniu kowalencyjnego lub niekowalencyjnego mostka wewnątrzcząsteczkowego lub substytucji i(lub) insercji aminokwasów wokół położeń z użyciem aminokwasu stabilizującego helisę alfa (np. aminokwasu α,α-dipodstawionego). [0130] W niektórych przykładach wykonania mostek
70 wewnątrzcząsteczkowy tworzy się między dwoma łańcuchami bocznymi aminokwasów, dzięki czemu następuje stabilizacja struktury trójwymiarowej części na końcu karboksylowym (np. aminokwasy 12-29) peptydu glukagonu. Łańcuchy boczne dwóch aminokwasów można połączyć ze sobą wiązaniami niekowalencyjnymi, np. przez wiązania wodorowe, oddziaływania jonowe, takie jak utworzenie mostków solnych, lub przez wiązania kowalencyjne. Kiedy łańcuchy boczne dwóch aminokwasów są połączone ze sobą poprzez jedno lub większą liczbę wiązań kowalencyjnych, można uznać, że peptyd w niniejszym dokumencie zawiera kowalencyjny mostek wewnątrzcząsteczkowy. Kiedy łańcuchy boczne dwóch aminokwasów są połączone ze sobą wiązaniami niekowalencyjnymi, np. wiązaniami wodorowymi, oddziaływaniami jonowymi, można uznać, że peptyd w niniejszym dokumencie zawiera niekowalencyjny mostek wewnątrzcząsteczkowy. [0131] W niektórych przykładach wykonania mostek wewnątrzcząsteczkowy tworzy się między dwoma aminokwasami rozdzielonymi 3 aminokwasami, np. aminokwasami w położeniach i oraz i+4, przy czym i jest dowolną liczbą całkowitą między 12 a 2 (np. 12, 13, 14, 1, 16, 17, 18, 19,, 21, 22, 23, 24 i 2). W szczególności łańcuchy boczne par aminokwasów 12 i 16, 16 i, i 24 lub 24 i 28 (par aminokwasów, w których i = 12, 16, lub 24) są połączone ze sobą i w ten sposób stabilizują helisę alfa glukagonu. Ewentualnie i może wynosić 17. [0132] W niektórych konkretnych przykładach wykonania, w których aminokwasy w położeniach i oraz i+4 są połączone mostkiem wewnątrzcząsteczkowym, wielkość
71 łącznika wynosi około 8 atomów lub około 7-9 atomów. [0133] W innych przykładach wykonania mostek wewnątrzcząsteczkowy tworzy się między dwoma aminokwasami rozdzielonymi dwoma aminokwasami, np. aminokwasami w położeniach j i j+3, przy czym j jest dowolną liczbą całkowitą między 12 a 26 (np. 12, 13, 14, 1, 16, 17, 18, 19,, 21, 22, 23, 24, 2 i 26). W pewnych konkretnych przykładach wykonania j wynosi 17. [0134] W niektórych konkretnych przykładach wykonania, w których aminokwasy w położeniach j i j+3 są połączone mostkiem wewnątrzcząsteczkowym, wielkość łącznika wynosi około 6 atomów lub około do 7 atomów. [013] W jeszcze innych przykładach wykonania mostek wewnątrzcząsteczkowy tworzy się między dwoma aminokwasami rozdzielonymi 6 aminokwasami, np. aminokwasami w położeniach k i k+7, przy czym k jest dowolną liczbą całkowitą od 12 do 22 (np. 12, 13, 14, 1, 16, 17, 18, 19,, 21 i 22). W pewnych konkretnych przykładach wykonania k wynosi 12, 13 lub 17. W pewnym przykładzie wykonania k wynosi 17. [0136] Do przykładów parowania aminokwasów zdolnych do utworzenia wiązania kowalencyjnego, w wyniku czego powstaje mostek łączący złożony z sześciu atomów, należą Orn i Asp, Glu i aminokwas o wzorze I, w którym n wynosi 2, oraz kwas homoglutaminowy i aminokwas o wzorze I, w którym n wynosi 1, przy czym wzór I stanowi:
![71 1 2 w którym n = 1 do 4 [wzór I] [0137] Do przykładów par aminokwasów zdolnych do utworzenia wiązania kowalencyjnego, w wyniku czego powstaje mostek łączący złożony z siedmiu atomów, należą](/docs-images/65/53262204/images/72-0.jpg "Orn-Glu (pierścień laktamowy); Lys-Asp (laktam); lub Homoser-Homoglu (lakton).")
72 w którym n = 1 do 4 [wzór I] [0137] Do przykładów par aminokwasów zdolnych do utworzenia wiązania kowalencyjnego, w wyniku czego powstaje mostek łączący złożony z siedmiu atomów, należą Orn-Glu (pierścień laktamowy); Lys-Asp (laktam); lub Homoser-Homoglu (lakton). Do przykładów parowania aminokwasów, które mogą utworzyć łącznik złożony z ośmiu atomów, należą Lys-Glu (laktam); Homolys-Asp (laktam); Orn-Homoglu (laktam); 4-amino-Phe-Asp (laktam); lub Tyr-Asp (lakton). Do przykładów parowania aminokwasów, które mogą utworzyć łącznik złożony z dziewięciu atomów, należą Homolys-Glu (laktam); Lys- Homoglu (laktam); 4-amino-Phe-Glu (laktam); lub Tyr-Glu (lakton). Dowolny z łańcuchów bocznych tych aminokwasów może być dodatkowo podstawiony przez dodatkowe grupy chemiczne, o ile nie jest zakłócona struktura trójwymiarowa helisy alfa. Specjalista w dziedzinie może przewidzieć alternatywne parowania lub alternatywne analogi aminokwasów, w tym pochodne zmodyfikowane chemicznie, które utworzą strukturę stabilizującą o podobnej wielkości i pożądanym działaniu. Na przykład mostek disiarczkowy homocysteina-homocysteina ma długość 6 atomów i można go dodatkowo zmodyfikować, aby uzyskać pożądane działanie. Nawet bez wiązania kowalencyjnego parowania
73 aminokwasów opisane powyżej lub podobne parowania, które specjalista w dziedzinie może przewidzieć, mogą także zapewniać dodatkową stabilność helisy alfa dzięki wiązaniom niekowalencyjnym, na przykład przez utworzenie mostków solnych lub oddziaływań przez wiązania wodorowe. [0138] Wielkość pierścienia laktamowego może się zmieniać w zależności od długości łańcuchów bocznych aminokwasów, i w jednym przykładzie wykonania laktam tworzy się przez połączenie łańcuchów bocznych aminokwasu lizyny z łańcuchem bocznym kwasu glutaminowego. Dodatkowe pewne przykłady wykonania obejmują następujące parowania, ewentualnie z mostkiem laktamowym: Glu w położeniu 12 z Lys w położeniu 16; naturalna Lys w położeniu 12 z Glu w położeniu 16; Glu w położeniu 16 z Lys w położeniu ; Lys w położeniu 16 z Glu w położeniu ; Glu w położeniu z Lys w położeniu 24; Lys w położeniu z Glu w położeniu 24; Glu w położeniu 24 z Lys w położeniu 28; Lys w położeniu 24 z Glu w położeniu 28. Kierunek wiązania amidowego w pierścieniu laktamowym można ewentualnie odwrócić (np. pierścień laktamowy można utworzyć między łańcuchami bocznymi Lys12 i Glu16 lub ewentualnie między Glu 12 i Lys16). [0139] Do stabilizacji helisy alfa w peptydach stanowiących analogi glukagonu można wykorzystać mostki wewnątrzcząsteczkowe inne niż mostek laktamowy. W jednym przykładzie wykonania mostek wewnątrzcząsteczkowy stanowi mostek hydrofobowy. W tym przypadku mostek wewnątrzcząsteczkowy tworzy się ewentualnie między łańcuchami bocznymi dwóch
74 aminokwasów znajdującymi się po stronie hydrofobowej helisy alfa w peptydzie stanowiącym analog glukagonu. Na przykład jednym z aminokwasów połączonym mostkiem hydrofobowym może być aminokwas w położeniu, 14 i 18. [0140] W jednym konkretnym aspekcie do sieciowania jednego lub dwóch zwrotów helisy alfa w peptydzie glukagonu przy pomocy całkowicie węglowodorowego układu sieciującego można zastosować metatezę olefin. Peptyd glukagonu w tym przypadku może zawierać aminokwasy α- metylowane zawierające olefinowe łańcuchy boczne o różnej długości i mające konfigurację stereochemiczną R lub S w położeniach i oraz i+4 lub i+7. Na przykład strona olefinowa może zawierać (CH2)n, w którym n jest dowolną liczbą całkowitą między 1 a 6. W jednym przykładzie wykonania n wynosi 3 dla długości sieciowania 8 atomów. Odpowiednie metody tworzenia takich mostków wewnątrzcząsteczkowych opisano w stanie techniki. Patrz na przykład Schafmeister i wsp., J. Am. Chem. Soc. 122: (00) oraz Walensky i wsp., Science 30: (04). Ewentualnie peptyd glukagonu może zawierać reszty O-allilo-Ser znajdujące się na sąsiednich zwrotach helisy, połączone ze sobą mostkiem w reakcji metatezy z zamknięciem pierścienia katalizowanym rutenem. Takie procedury sieciowania opisano na przykład w Blackwell i wsp., Angew, Chem., Int. Ed. 37: (1998). [0141] W innym konkretnym aspekcie do sieciowania jednego zwrotu helisy alfa wykorzystuje się nienaturalny aminokwas tiodialaninowy, lantioninę, który powszechnie stosuje się jako związek
75 peptydomimetyczny wobec cysteiny. Odpowiednie metody cyklizacji z użyciem lantioniny są znane w stanie techniki. Patrz na przykład Matteucci i wsp., Tetrahedron Letters 4: (04); Mayer i wsp., J. Peptide Res. 1: (1998); Polinsky i wsp., J. Med. Chem. 3: (1992); Osapay i wsp., J. Med. Chem. 40: (1997); Fukase i wsp., Bull. Chem. Soc. Jpn. 6: (1992); Harpp i wsp., J. Org. Chem. 36: (1971); Goodman i Shao, Pure Appl. Chem. 68: (1996); oraz Osapay i Goodman, J. Chem. Soc. Chem. Commun (1993). [0142] W niektórych przykładach wykonania do stabilizacji helisy alfa peptydu glukagonu stosuje się α,ω-diaminoalkanowe struktury wiążące, np. 1,4- diaminopropan i 1,-diaminopentan, między dwoma resztami Glu w położeniach i oraz i+7. Takie struktury wiążące prowadzą do utworzenia mostka o długości 9 lub więcej atomów, w zależności od długości diaminoalkanowej struktury wiążącej. Odpowiednie metody wytwarzania peptydów usieciowanych za pomocą takich struktur wiążących opisano w stanie techniki. Patrz na przykład Phelan i wsp., J. Am. Chem. Soc. 119: (1997). [0143] W jeszcze innym przykładzie wykonania wynalazku do sieciowania jednego lub dwóch zwrotów helisy alfa w peptydzie glukagonu stosuje się mostek disiarczkowy. Ewentualnie do stabilizacji helisy alfa w peptydzie glukagonu stosuje się zmodyfikowany mostek disiarczkowy, w którym jeden lub oba atomy siarki są zastąpione grupą metylenową, co powoduje makrocyklizację izosteryczną. Odpowiednie metody
76 modyfikacji peptydów mostkami disiarczkowymi lub cyklizacji z użyciem siarki opisano na przykład w Jackson i wsp., J. Am. Chem. Soc. 113: (1991) oraz Rudinger i Jost, Experientia : (1964). [0144] W jeszcze innym przykładzie wykonania helisę alfa w peptydzie glukagonu stabilizuje się przez połączenie atomu metalu z dwiema resztami His lub parą His i Cys w położeniach i oraz i+4. Atomem metalu może być na przykład Ru(III), Cu(II), Zn(II) lub Cd(II). Takie metody stabilizacji helisy alfa z użyciem wiązania metalu są znane w stanie techniki. Patrz na przykład Andrews i Tabor, Tetrahedron : (1999); Ghadiri i wsp., J. Am. Chem. Soc. 112: (1990); oraz Ghadiri i wsp., J. Am. Chem. Soc. 119: (1997). [014] Helisę alfa w peptydzie glukagonu można ewentualnie ustabilizować innymi środkami cyklizacji peptydów, których przegląd podano w Davies, J. Peptide. Sci. 9: (03). Helisę alfa można stabilizować przez utworzenie mostka amidowego, mostka tioeterowego, mostka tioestrowego, mostka mocznikowego, mostka karbaminianowego, mostka sulfonamidowego i podobne. Na przykład mostek tioestrowy można utworzyć między końcem C a łańcuchem bocznym reszty Cys. Ewentualnie tioester można utworzyć poprzez łańcuchy boczne aminokwasów zawierających tiol (Cys) i kwas karboksylowy (np. Asp, Glu). W innej metodzie środek sieciujący, na przykład kwas dikarboksylowy, np. kwas suberynowy (kwas oktanodiowy), itd. może wprowadzić połączenie między dwiema grupami funkcyjnymi łańcucha bocznego aminokwasów, na przykład wolną grupą aminową,
77 hydroksylową, tiolową i ich kombinacjami. [0146] Według jednego przykładu wykonania helisę alfa w peptydzie glukagonu stabilizuje się dzięki wprowadzeniu aminokwasów hydrofobowych w położeniach i oraz i+4. Na przykład i może stanowić Tyr, a i+4 może stanowić Val lub Leu; i może stanowić Phe, a i+4 może stanowić Cys lub Met; i może stanowić Cys, a i+4 może stanowić Met lub i może stanowić Phe, a i+4 może stanowić Ile. Należy rozumieć, że na potrzeby niniejszego dokumentu powyższe parowania aminokwasów można odwrócić, tak że podany aminokwas w położeniu i może ewentualnie znajdować się w i+4, a aminokwas i+4 może znajdować się w położeniu i. [0147] Według innych przykładów wykonania wynalazku helisę alfa stabilizuje się dzięki wprowadzeniu (w wyniku substytucji lub insercji aminokwasów) jednego lub większej liczby aminokwasów stabilizujących helisę alfa w części C-końcowej peptydu glukagonu (w obrębie aminokwasów 12-29). W konkretnym przykładzie wykonania aminokwasem stabilizującym helisę alfa jest aminokwas α,α-dipodstawiony, w tym, ale nieograniczająco kwas aminoizomasłowy (AIB), aminokwas dipodstawiony tą samą lub inną grupą wybraną spośród metylu, etylu, propylu i n-butylu, lub cyklooktanem lub cykloheptanem (np. kwas 1-aminocyklooktano-1-karboksylowy). W niektórych przykładach wykonania jedno, dwa, trzy, cztery lub więcej spośród położeń 16, 17, 18, 19,, 21, 24 lub 29 w peptydzie glukagonu jest podstawionych aminokwasem α,α-dipodstawionym. W konkretnym przykładzie wykonania jedno, dwa, trzy lub wszystkie spośród położeń 16,, 21 lub 24 jest podstawionych przez aminokwas α,α-
78 dipodstawiony, np. AIB. Na przykład peptyd glukagonu może zawierać substytucję położenia 16 przez AIB w nieobecności mostka wewnątrzcząsteczkowego, np. niekowalencyjnego mostka wewnątrzcząsteczkowego (np. mostka solnego) lub kowalencyjnego mostka wewnątrzcząsteczkowego (np. laktamu). Takie peptydy niezawierające mostka wewnątrzcząsteczkowego łatwo jest otrzymać, co jest korzystne. [0148] Według niektórych przykładów wykonania peptyd glukagonu niezawierający mostka wewnątrzcząsteczkowego zawiera jedną lub większą liczbę substytucji w obrębie położeń aminokwasów przez aminokwas α,αdipodstawiony oraz grupę acylową lub alkilową połączoną kowalencyjnie z łańcuchem bocznym aminokwasu w peptydzie glukagonu, np. aminokwasu w położeniach lub 40 peptydu glukagonu. W konkretnych przykładach wykonania grupa acylowa lub alkilowa jest nienatywna względem aminokwasu występującego naturalnie. W pewnych aspektach grupa acylowa lub alkilowa jest nienatywna względem aminokwasu w położeniu. Takie acylowane lub alkilowane peptydy glukagonu niezawierające mostka wewnątrzcząsteczkowego wykazują zwiększoną aktywność wobec receptorów GLP-1 i glukagonu w porównaniu z odpowiadającymi im peptydami nieacylowanymi. Dodatkowe zwiększenie aktywności wobec receptorów GLP-1 i glukagonu można uzyskać dzięki acylowanym peptydom glukagonu niezawierającym mostka wewnątrzcząsteczkowego przez wprowadzenie wstawki między grupą acylową lub alkilową a łańcuchem bocznym aminokwasu w położeniach lub 40 peptydu. Acylowanie i alkilowanie, z wprowadzaniem lub bez wprowadzania wstawek, opisano
79 78 1 poniżej w niniejszym dokumencie. [0149] W konkretnych przykładach wykonania acylowany lub alkilowany peptyd glukagonu bądź ich analog zawiera ponadto modyfikację selektywnie zmniejszającą aktywność wobec receptora GLP-1. Na przykład acylowany lub alkilowany peptyd glukagonu bądź ich analog zawiera jedno lub kombinację spośród: C-końcowego alfakarboksylanu, delecji aminokwasów C-końcowych względem aminokwasu w położeniu 27 lub 28 (np. delecja aminokwasu w położeniu 29, delecji aminokwasów w położeniach 28 i 29), substytucji Thr w położeniu 7 przez duży, alifatyczny aminokwas niepolarny, np. Ile. W niektórych przykładach wykonania położenie 16 lub położenie jest podstawione przez aminokwas α,αdipodstawiony, np. AIB. W niektórych przykładach wykonania położenie jest podstawione przez aminokwas α,α-dipodstawiony, np. AIB. W niektórych przykładach wykonania położenie jest podstawione przez aminokwas α,α-dipodstawiony, np. AIB, a położenie 16 jest podstawione przez aminokwas o wzorze IV [wzór IV], 2 w którym n wynosi 1 do 16, lub 1 do, lub 1 do 7, lub 1 do 6, lub 2 do 6, lub 2, lub 3, lub 4, lub, przy
80 79 czym każdy spośród R1 i R2 jest niezależnie wybrany z grupy składającej się z: H, C1-C18 alkil, (C1-C18 alkilo)oh, (C1-C18 alkilo)nh2, (C1-C18 alkilo)sh, (C0-C4 alkilo)(c3-c6)cykloalkil, (C0-C4 alkilo)(c2-c heterocykl), (C0-C4 alkilo)(c6-c arylo)r7 i (C1-C4 alkilo)(c3-c9 heteroaryl), przy czym R7 oznacza H lub OH, a łańcuch boczny aminokwasu o wzorze IV zawiera wolną grupę aminową. W konkretnych przykładach wykonania aminokwas o wzorze IV stanowi kwas 2,3- diaminopropionowy (DAP), kwas 2,4-diaminomasłowy (DAB), Orn, Lys lub homolys. Kombinacja aminokwasu o wzorze IV w położeniu 16 i aminokwasu alfa,alfa-dipodstawionego korzystnie zapewnia zwiększoną aktywność wobec każdego spośród receptorów glukagonu, GLP-1 i GIP. Łączenie ugrupowań hydrofilowych [010] W innym przykładzie wykonania rozpuszczalność peptydów glukagonu ujawnianych w niniejszym dokumencie zwiększa się dzięki połączeniu kowalencyjnemu ugrupowania hydrofilowego z peptydem. Ugrupowania hydrofilowe można dołączyć do peptydów glukagonu w dowolnych odpowiednich warunkach wykorzystywanych w reakcji białka z cząsteczką aktywowanego polimeru. Można zastosować dowolne metody znane w stanie techniki, w tym acylowanie, alkilowanie redukcyjne, addycję Michaela, alkilowanie tiolu lub inne chemoselektywne metody sprzęgania i ligacji poprzez grupę reaktywną w ugrupowaniu PEG (np. grupę aldehydową, aminową, estrową, tiolową, α-haloacetylową, maleimidową lub hydrazynową) z grupą reaktywną w związku docelowym (np. grupę aldehydową, aminową, estrową, tiolową, α-haloacetylową, maleimidową lub

81 80 1 hydrazynową). Do grup aktywujących, które można wykorzystać do połączenia polimeru rozpuszczalnego w wodzie z jednym lub większą liczbą białek, należą bez ograniczania, grupa sulfonowa, maleimidowa, sulfhydrylowa, tiolowa, triflatowa, tresylatowa, azyrydynowa, oksyranowa, -pirydylowa i alfahalogenowana grupa acylowa (np. kwas alfa-jodooctowy, kwas alfa-bromooctowy, kwas alfa-chlorooctowy). Jeśli wybrany polimer zostaje dołączony do peptydu w wyniku alkilowania redukcyjnego, powinien on zawierać pojedynczy aldehyd reaktywny, dzięki czemu kontroluje się stopień polimeryzacji. Patrz na przykład Kinstler i wsp., Adv. Drug. Delivery Rev. 4: (02); Roberts i wsp., Adv. Drug Delivery Rev. 4: (02); oraz Zalipsky i wsp., Adv. Drug Delivery Rev. 16: (199). [011] W konkretnym aspekcie wynalazku resztę aminokwasową w peptydzie glukagonu zawierającą tiol modyfikuje się ugrupowaniem hydrofilowym, takim jak PEG. W niektórych przykładach wykonania tiol modyfikuje się PEG aktywowanym maleimidem w reakcji addycji Michaela, dzięki czemu otrzymuje się peptyd PEGylowany zawierający wiązanie eterowe pokazane poniżej. 2 [012] W niektórych przykładach wykonania tiol modyfikuje się PEG aktywowanym haloacetylem w reakcji substytucji nukleofilowej, dzięki czemu otrzymuje się peptyd PEGylowany zawierający wiązanie tioeterowe
![81 pokazane poniżej. 1 2 [013] Do odpowiednich ugrupowań hydrofilowych należą poli(glikol etylenowy) (PEG), poli(glikol propylenowy), polioksyetylowane poliole (np.](/docs-images/65/53262204/images/82-0.jpg "POG), polioksyetylowany sorbitol, polioksyetylowana glukoza, polioksyetylowany glicerol (POG), polioksyalkileny, aldehyd propionowy poli(glikolu etylenowego), kopolimery glikolu etylenowego/glikolu")
82 81 pokazane poniżej. 1 2 [013] Do odpowiednich ugrupowań hydrofilowych należą poli(glikol etylenowy) (PEG), poli(glikol propylenowy), polioksyetylowane poliole (np. POG), polioksyetylowany sorbitol, polioksyetylowana glukoza, polioksyetylowany glicerol (POG), polioksyalkileny, aldehyd propionowy poli(glikolu etylenowego), kopolimery glikolu etylenowego/glikolu propylenowego, glikol monometoksypolietylenowy, mono-(c1-c) alkoksy- lub aryloksypoli(glikol etylenowy), karboksymetyloceluloza, poliacetale, poli(alkohol winylowy) (PVA), poliwinylopirolidon, poli-1,3-dioksolan, poli-1,3,6- trioksan, kopolimer etylenu i bezwodnika maleinowego, poli(beta-aminokwasy) (homopolimery lub kopolimery przypadkowe), poli(n-winylopirolidon)poli(glikol etylenowy), homopolimery glikolu propropylenowego (PPG) i inne tlenki polialkilenu, kopolimery poli(tlenku propylenu) i tlenku etylenu, kwasy kolonowe (ang. colonic acids) lub inne polimery polisacharydów, fikol lub dekstran i ich mieszaniny. [014] Ugrupowanie hydrofilowe, np. łańcuch poli(glikolu etylenowego) według niektórych przykładów wykonania, ma masę cząsteczkową wybraną z zakresu około 00 do około daltonów. W jednym przykładzie wykonania ugrupowanie hydrofilowe, np. PEG, ma masę cząsteczkową wybraną z zakresu około 00 do około 000 daltonów lub około 00 do około 000 daltonów. W innym
83 przykładzie wykonania ugrupowanie hydrofilowe, np. PEG, ma masę cząsteczkową około 000 do około 000 daltonów. W innych pewnych przykładach wykonania ugrupowanie hydrofilowe, np. PEG, ma masę cząsteczkową około 000 do około daltonów. [01] W jednym przykładzie wykonania jako ugrupowanie hydrofilowe wykorzystuje się dekstrany. Dekstrany są polimerami polisacharydów złożonymi z podjednostek glukozy, połączonych głównie wiązaniami α1-6. Dekstran jest dostępny w wielu zakresach masy cząsteczkowej, np. około 1 kd do około 0 kd lub od około,, 1 lub kd do około, 30, 40, 0, 60, 70, 80 lub 90 kd. [016] Przewiduje się zastosowanie polimerów liniowych lub rozgałęzionych. Otrzymane preparaty koniugatów mogą być zasadniczo monodyspersyjne lub polidyspersyjne i mogą zawierać około 0,, 0,7, 1, 1,2, 1, lub 2 ugrupowań polimerowych na peptyd. [017] W jednym przykładzie wykonania ugrupowanie hydrofilowe stanowi łańcuch poli(glikolu etylenowego) (PEG), ewentualnie połączony z peptydem w jednym lub więcej spośród położeń 16, 17, 21, 24, 29, położeniu w obrębie przedłużenia C-końcowego, np. 30, 31, 32, 33, 34, 3, 36, 37, 38, 39 lub na aminokwasie C-końcowym (np. 40). W niektórych przykładach wykonania aminokwas natywny w tym położeniu jest podstawiony przez aminokwas zawierający łańcuch boczny odpowiedni do sieciowania z ugrupowaniami hydrofilowymi, aby ułatwić połączenie ugrupowania hydrofilowego z peptydem. W pewnych przykładach wykonania aminokwas natywny w tym położeniu jest podstawiony przez resztę Lys, Cys, Orn, homocysteiny lub acetylofenyloalaniny. W innych
84 przykładach wykonania do peptydu na końcu C dodaje się aminokwas zmodyfikowany, tak że zawiera grupę hydrofilową. Inne modyfikacje zwiększające rozpuszczalność [018] W innym przykładzie wykonania rozpuszczalność dowolnego spośród peptydów glukagonu można zwiększyć dzięki substytucjom i(lub) addycjom aminokwasów wprowadzającym aminokwas naładowany do części C- końcowej peptydu, korzystnie w położeniu C-końcowym względem położenia 27 według NR. ID. SEKW.: 1. Ewentualnie jeden, dwa lub trzy aminokwasy naładowane można wprowadzić w obrębie części C-końcowej, korzystnie w położeniu C-końcowym względem położenia 27. W niektórych przykładach wykonania aminokwas lub aminokwasy natywne w położeniach 28 i(lub) 29 są podstawione przez jeden lub dwa aminokwasy naładowane, i(lub) w dodatkowym przykładzie wykonania do końca C peptydu dodaje się ponadto jeden do trzech aminokwasów naładowanych. W pewnych przykładach wykonania jeden, dwa lub wszystkie aminokwasy naładowane mają ładunek ujemny. W niektórych przykładach wykonania aminokwas naładowany ujemnie (kwaśny) stanowi kwas asparaginowy lub kwas glutaminowy. Do peptydu glukagonu można wprowadzić dodatkowe modyfikacje, np. substytucje konserwatywne, które nadal umożliwiają zachowanie aktywności GIP (i ewentualnie aktywności GLP-1 i(lub) aktywności glukagonu). Koniugaty i fuzje [019] Niniejsze ujawnienie obejmuje ponadto inne koniugaty, w których peptydy glukagonu według wynalazku są połączone, ewentualnie poprzez wiązanie kowalencyjne
85 i ewentualnie przez łącznik, z ugrupowaniem sprzęgającym. Połączenie można uzyskać dzięki kowalencyjnym wiązaniom chemicznym, siłom fizycznym, na przykład elektrostatycznym, wodorowym, jonowym, van der Waalsa, bądź oddziaływaniom hydrofobowym lub hydrofilowym. Można zastosować różnorodne niekowalencyjne układy sprzęgające, w tym biotynaawidyna, ligand/receptor, enzym/substrat, kwas nukleinowy/białko wiążące kwas nukleinowy, lipid/białko wiążące lipid, partnerów cząsteczek adhezji komórkowej; lub dowolnych partnerów wiązania lub ich fragmenty wykazujące wobec siebie powinowactwo. [0160] Peptyd można połączyć z ugrupowaniami sprzęgającymi poprzez bezpośrednie wiązanie kowalencyjne w wyniku reakcji docelowych reszt aminokwasu w peptydzie z organicznym środkiem derywatyzującym, który może reagować z wybranymi łańcuchami bocznymi lub N- lub C-końcowymi resztami w tych aminokwasach docelowych. Do grup reaktywnych w peptydzie lub koniugacie należą np. grupa aldehydowa, aminowa, estrowa, tiolowa, α-haloacetylowa, maleimidowa lub hydrazynowa. Do środków derywatyzujących należą na przykład ester maleimidobenzoilowy sulfosukcynoimidu (sprzęganie poprzez reszty cysteiny), N- hydroksysukcynoimid (poprzez reszty lizyny), aldehyd glutarowy, bezwodnik bursztynowy lub inne środki znane w stanie techniki. Ewentualnie ugrupowania sprzęgające można połączyć z peptydem pośrednio poprzez nośniki pośrednie, takie jak nośniki polisacharydowe lub polipeptydowe. Do przykładów nośników polisacharydowych należy aminodekstran. Do przykładów odpowiednich
86 nośników polipeptydowych należą polilizyna, kwas poliglutaminowy, kwas poliasparaginowy, ich kopolimery oraz mieszane polimery tych aminokwasów i inne, np. seryny, które nadają pożądane właściwości rozpuszczalności otrzymywanemu zmodyfikowanemu nośnikowi. [0161] Reszty cysteinylowe poddaje się najczęściej reakcji z α-halooctanami (i odpowiednimi aminami), takimi jak kwas chlorooctowy lub chloroacetamid, uzyskując pochodne karboksymetylowe lub karboksyamidometylowe. Reszty cysteinylowe derywatyzuje się ponadto w reakcji z bromotrifluoroacetonem, kwasem alfa-bromo-β-(-imidazoilo)propionowym, fosforanem chloroacetylu, N-alkilomaleimidami, disiarczkiem 3- nitro-2-pirydylu, disiarczkiem metylo-2-pirydylu, p- chlorortęciobenzoesanem, 2-chlorortęcio-4-nitrofenolem lub chloro-7-nitrobenzo-2-oksa-1,3-diazolem. [0162] Reszty histydylowe derywatyzuje się w reakcji z pirowęglanem dietylu w ph,-7,0, ponieważ środek ten jest stosunkowo specyficzny wobec histydylowego łańcucha bocznego. Użyteczny jest także bromek parabromofenacylu; reakcję korzystnie prowadzi się w 0,1 M kakodylanie sodu w ph 6,0. [0163] Reszty lizynylowe i końcowe aminowe reszty poddaje się reakcji z bezwodnikami bursztynowymi lub innych kwasów karboksylowych. Derywatyzacja z użyciem tych środków prowadzi do odwrócenia ładunku reszt lizynylowych. Do innych odpowiednich odczynników do uzyskiwania pochodnych reszt zawierających grupy alfaaminowe należą imidoestry, takie jak pikolinoimidan metylu, fosforan pirydoksalu, pirydoksal,
87 chloroborowodorek, kwas trinitrobenzenosulfonowy, O- metyloizomocznik, 2,4-pentanodion i reakcja katalizowana transaminazą z glioksylanem. [0164] Reszty arginylowe modyfikuje się za pomocą reakcji z jednym lub większą liczbą typowych odczynników, między innymi fenyloglioksalem, 2,3- butanodionem, 1,2-cykloheksanodionem i ninhydryną. Derywatyzacja reszt argininy wymaga, aby reakcję prowadzić w warunkach zasadowych, ze względu na wysokie pka grupy funkcyjnej guanidyny. Te odczynniki mogą ponadto reagować z grupami lizyny, a także z grupą epsilon-aminową argininy. [016] Określoną modyfikację reszt tyrozylowych można przeprowadzić w szczególności ze względu na wprowadzanie znaczników widmowych do reszt tyrozylowych w wyniku reakcji z aromatycznymi związkami diazoniowymi lub tetranitrometanem. Najczęściej do otrzymywania odpowiednio związków O-acetylotyrozylowych i pochodnych 3-nitrowych można wykorzystywać N-acetyloimidazol i tetranitrometan. [0166] Karboksylowe grupy boczne (aspartylowe lub glutamylowe) modyfikuje się w sposób selektywny w reakcji z karbodiimidami (R-N.dbd.C.dbd.N-R ), w których R i R są różnymi grupami alkilowymi, takimi jak 1-cykloheksylo-3-[2-morfolinylo-(4- etylo)]karbodiimid lub 1-etylo-3-(4-azonia-4,4- dimetylopentylo)karbodiimid. Ponadto reszty aspartylowe i glutamylowe przekształca się w reszty asparaginylowe i glutaminylowe w reakcji z jonami amonowymi. [0167] Do innych modyfikacji należy hydroksylowanie proliny i lizyny, fosforylacja grup hydroksylowych
88 reszt serylowych lub treonylowych, metylacja grup alfaaminowych łańcuchów bocznych lizyny, argininy i histydyny (T. E. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, str (1983)), deamidacja asparagin lub glutaminy, acetylowanie aminy N-końcowej i(lub) amidowanie lub estryfikacja C-końcowej grupy kwasu karboksylowego. [0168] Inny rodzaj modyfikacji kowalencyjnej obejmuje sprzęganie chemiczne lub enzymatyczne glikozydów z peptydem. Cukier lub cukry można połączyć z (a) argininą i histydyną, (b) wolnymi grupami karboksylowymi, (c) wolnymi grupami sulfhydrylowymi, takimi jak te w cysteinie, (d) wolnymi grupami hydroksylowymi, takimi jak te w serynie, treoninie lub hydroksyprolinie, (e) resztami aromatycznymi, takimi jak te w tyrozynie lub tryptofanie, lub (f) grupą amidową glutaminy. Te metody opisano w WO87/0330 opublikowanym 11 września 1987 r. oraz w Aplin i Wriston, CRC Crit. Rev. Biochem., str (1981). [0169] Do przykładowych ugrupowań sprzęgających, które można połączyć z dowolnym spośród peptydów glukagonu opisanych w niniejszym dokumencie, należą, ale nieograniczająco, heterologiczny peptyd lub polipeptyd (w tym na przykład białko osocza), środek kierujący, immunoglobulina lub jej część (np. region zmienny, CDR lub region Fc), znacznik diagnostyczny, taki jak radioizotop, fluorofor lub znacznik enzymatyczny, polimer, w tym polimery rozpuszczalne w wodzie lub inne środki terapeutyczne lub diagnostyczne. W jednym przykładzie wykonania dostarcza się koniugat
89 zawierający peptyd glukagonu według niniejszego wynalazku i białko osocza, przy czym białko osocza wybrane jest z grupy składającej się z albuminy, transferyny, fibrynogenu i globulin. [0170] W niektórych przykładach wykonania łącznik zawiera łańcuch atomów o długości od 1 do około 60 lub 1 do 30 atomów lub dłuższy, 2 do atomów, 2 do atomów, do atomów lub o długości do atomów. W niektórych przykładach wykonania wszystkie atomy w łańcuchu stanowią atomy węgla. W niektórych przykładach wykonania atomy w łańcuchu w szkielecie łącznika wybrane są z grupy składającej się z C, O, N i S. Atomy w łańcuchu i łączniki można wybierać w zależności od ich oczekiwanej rozpuszczalności (hydrofilowości), tak by uzyskać koniugat o większej rozpuszczalności. W niektórych przykładach wykonania łącznik dostarcza grupy funkcyjnej poddawanej trawieniu przez enzym lub inny katalizator lub w wyniku warunków hydrolitycznych występujących w tkance lub narządzie lub komórce docelowej. W niektórych przykładach wykonania łącznik ma wystarczającą długość, aby zmniejszyć możliwość zawady sterycznej. Jeśli łącznik stanowi wiązanie kowalencyjne lub wiązanie peptydylowe, a koniugat stanowi polipeptyd, cały koniugat może stanowić białko fuzyjne. Takie łączniki peptydylowe mogą mieć dowolną długość. Przykładowe łączniki mają długość od około 1 do 0 aminokwasów, do 0, 3 do, do, do 1, lub do 30 aminokwasów długości. Takie białka fuzyjne można ewentualnie wytworzyć metodami inżynierii genetycznej z rekombinacją znanymi specjaliście w dziedzinie.
90 [0171] Jak stwierdzono powyżej, w niektórych przykładach wykonania peptydy glukagonu są sprzężone, np. w fuzji z immunoglobuliną lub jej częścią (np. regionem zmiennym, CDR lub regionem Fc). Do znanych rodzajów immunoglobulin (Ig) należą IgG, IgA, IgE, IgD lub IgM. Region Fc stanowi region C-końcowy łańcucha ciężkiego Ig, odpowiedzialny za wiązanie z receptorami Fc zapewniającymi aktywności, takie jak możliwość ponownego wykorzystania (która zwiększa okres półtrwania), cytotoksyczność komórkową zależną od przeciwciał (ang. AADC) i cytotoksyczność zależną od dopełniacza (ang. CDC). [0172] Na przykład według niektórych definicji region Fc łańcucha ciężkiego ludzkiej IgG obejmuje odcinek od Cys226 do końca C łańcucha ciężkiego. Region zawiasowy obejmuje zasadniczo odcinek od Glu216 do Pro230 ludzkiej IgG1 (regiony zawiasowe innych izotypów IgG można dopasować do sekwencji IgG1 na podstawie dopasowania cystein uczestniczących w wiązaniach cysteinowych). Region Fc IgG zawiera dwie domeny stałe, CH2 i CH3. Domena CH2 regionu Fc ludzkiej IgG obejmuje zazwyczaj odcinek od aminokwasu 231 do aminokwasu 341. Domena CH3 regionu Fc ludzkiej IgG obejmuje zazwyczaj odcinek od aminokwasu 342 do 447. Dokonuje się odniesienia do numeracji aminokwasów w immunoglobulinach lub fragmentach immunoglobulin lub ich regionach, przy czym wszystkie opierają się na publikacji Kabat i wsp. 1991, Sequences of Proteins of Immunological Interest, U.S. Department of Public Health, Bethesda, Md. W powiązanych przykładach wykonania region Fc może zawierać jeden lub większą
91 liczbę natywnych lub zmodyfikowanych regionów stałych łańcucha ciężkiego immunoglobuliny innego niż CH1, na przykład regiony CH2 i CH3 IgG i IgA lub regiony CH3 i CH4 IgE. [0173] Do odpowiednich ugrupowań sprzęgających należą części sekwencji immunoglobuliny obejmujące miejsce wiązania FcRn. FcRn, receptor ratujący, odpowiada za ponowne wykorzystanie immunoglobulin oraz ich powrót do krwiobiegu. Region części Fc IgG wiążący się z receptorem FcRn opisano na podstawie krystalografii rentgenowskiej (Burmeister i wsp. 1994, Nature 372:379). Główny obszar kontaktu Fc z FcRn znajduje się w pobliżu połączenia domen CH2 i CH3. Wszystkie kontakty Fc-FcRn znajdują się w obrębie pojedynczego łańcucha ciężkiego Ig. Główne miejsca kontaktu obejmują reszty aminokwasowe 248, -27, 272, 28, 288, , i 314 w domenie CH2 oraz reszty aminokwasowe , 428 i w domenie CH3. [0174] Niektóre ugrupowania sprzęgające mogą, ale nie muszą, obejmować miejsce lub miejsca wiązania FcγR. FcγR są odpowiedzialne za ADCC i CDC. Przykładami położeń w obrębie regionu Fc tworzącymi kontakt bezpośredni z FcγR są aminokwasy (region zawiasu dolnego), aminokwasy (pętla B/C), aminokwasy (pętla C /E) i aminokwasy (pętla F/G) (Sondermann i wsp., Nature 406: , 00). Region zawiasu dolnego IgE bierze ponadto udział w wiązaniu FcRI (Henry i wsp., Biochemistry 36, , 1997). Reszty biorące udział w wiązaniu receptora IgA opisano w Lewis i wsp. (J Immunol. 17: , 0). Reszty aminokwasowe biorące udział
92 w wiązaniu receptora IgE opisano w Sayers i wsp. (J Biol Chem. 279(34):33-, 04). [017] Modyfikacje aminokwasów można wprowadzić w regionie Fc immunoglobuliny. Takie warianty regionów Fc zawierają co najmniej jedną modyfikację aminokwasu w domenie CH3 regionu Fc (reszty ) i(lub) co najmniej jedną modyfikację aminokwasu w domenie CH2 regionu Fc (reszty ). Do mutacji, co do których sądzi się, że zapewniają zwiększone powinowactwo do FcRn, należą T26A, T307A, E380A i N434A (Shields i wsp. 01, J. Biol. Chem. 276:691). Inne mutacje mogą zmniejszać wiązanie regionu Fc z FcγRI, FcγRIIA, FcγRIIB i(lub) FcγRIIIA bez znacznego zmniejszenia powinowactwa do FcRn. Na przykład substytucja Asn w położeniu 297 regionu Fc przez Ala lub inny aminokwas usuwa wysoce konserwatywne miejsce N-glikozylacji i może doprowadzić do zmniejszonej immunogenności jednocześnie z wydłużeniem okresu półtrwania regionu Fc, a także zmniejszonego wiązania z FcγRs (Routledge i wsp. 199, Transplantation 60:847; Friend i wsp. 1999, Transplantation 68:1632; Shields i wsp. 199, J. Biol. Chem. 276:691). Wprowadzono modyfikacje aminokwasów w położeniach IgG1, które zmniejszają wiązanie z FcγRs (Ward i Ghetie 199, Therapeutic Immunology 2:77 oraz Armour i wsp. 1999, Eur. J. Immunol. 29:2613). Niektóre przykładowe substytucje aminokwasów opisano w patentach USA 7,3,008 i 7,381,408. [0176] Niniejsze ujawnienie obejmuje ponadto peptydy lub białka fuzyjne glukagonu, w których drugi peptyd lub polipeptyd został poddany fuzji z końcem, np. końcem karboksylowym peptydu glukagonu. W niektórych
93 przykładach wykonania drugim peptydem dodawanym do końca karboksylowego peptydu glukagonu jest NR. ID. SEKW.: 9 (GPSSGAPPPS), NR. ID. SEKW.: 97 (KRNRNNIA) lub NR. ID. SEKW.: 98 (KRNR) połączone z aminokwasem 29 peptydu glukagonu. W innych przykładach wykonania drugi peptyd stanowi XGPSSGAPPPS (NR. ID. SEKW.: 96), przy czym X wybiera się spośród jednego z podstawowych aminokwasów, np. kwasu glutaminowego, kwasu asparaginowego lub glicyny. W jednym przykładzie wykonania X oznacza aminokwas, na przykład Cys, który zawiera ponadto ugrupowanie hydrofilowe połączone kowalencyjnie z łańcuchem bocznym tego aminokwasu. Takie przedłużenia C-końcowe zwiększają rozpuszczalność i mogą ponadto zwiększyć aktywność GIP lub GLP-1. W niektórych przykładach wykonania, w których peptyd glukagonu zawiera ponadto przedłużenie na końcu karboksylowym, przy czym aminokwas na końcu karboksylowym przedłużenia kończy się raczej grupą amidową lub grupą estrową, a nie kwasem karboksylowym. [0177] W niektórych przykładach wykonania, np. w peptydach glukagonu zawierających przedłużenie C- końcowe, treonina w położeniu 29 natywnego peptydu glukagonu jest zastąpiona glicyną. Na przykład peptyd glukagonu zawierający substytucję glicyną za treoninę w położeniu 29 i zawierający przedłużenie C-końcowe GPSSGAPPPS (NR. ID. SEKW.: 9) ma czterokrotnie większą siłę działania wobec receptora GLP-1 względem natywnego glukagonu zmodyfikowanego tak, że zawiera to samo przedłużenie C-końcowe. Tę substytucję T29G można wykorzystać w połączeniu z innymi modyfikacjami ujawnianymi w niniejszym dokumencie, aby zwiększyć
94 powinowactwo peptydów glukagonu do receptora GLP-1. Na przykład substytucję T29G można połączyć z substytucjami aminokwasów S16E i NK, ewentualnie z mostkiem laktamowym między aminokwasami 16 i oraz ewentualnie z dodaniem łańcucha PEG zgodnie z opisem w niniejszym dokumencie. [0178] W niektórych przykładach wykonania aminokwas dodaje się na końcu C i dodatkowy aminokwas jest wybieraby z grupy składającej się z kwasu glutaminowego, kwasu asparaginowego i glicyny. [0179] Niniejsze ujawnienie obejmuje ponadto multimery zmodyfikowanych peptydów glukagonu ujawnianych w niniejszym dokumencie. Dwa lub więcej spośród zmodyfikowanych peptydów glukagonu można połączyć ze sobą stosując standardowe środki łączące i procedury znane specjaliście w dziedzinie. Na przykład dimery można utworzyć między dwoma zmodyfikowanymi peptydami glukagonu przez zastosowanie dwufunkcyjnych tiolowych środków sieciujących i dwufunkcyjnych aminowych środków sieciujących, w szczególności w przypadku peptydów glukagonu z substytucją resztami cysteiny, lizyny, ornityny, homocysteiny lub acetylofenyloalaniny. Acylowanie i alkilowanie [0180] Według niektórych przykładów wykonania peptydy glukagonu ujawniane w niniejszym dokumencie modyfikuje się tak, że zawierają grupę acylową lub grupę alkilową, np. grupę acylową lub alkilową, która nie jest natywna w aminokwasach występujących naturalnie. Acylowanie lub alkilowanie może zwiększyć okres półtrwania peptydów glukagonu w krążeniu. Acylowanie lub alkilowanie może korzystnie opóźnić początek działania i(lub) wydłużyć
95 okres działania wobec receptorów glukagonu i(lub) GLP-1 i(lub) zwiększyć odporność na proteazy, takie jak DPP- IV i(lub) zwiększyć rozpuszczalność. Aktywność wobec receptorów glukagonu i(lub) GLP-1 i(lub) GIP peptydu glukagonu po acylowaniu może być utrzymana. W niektórych przykładach wykonania siła działania acylowanych peptydów glukagonu jest porównywalna do nieacylowanych wariantów peptydów glukagonu. W alternatywnych przykładach wykonania siła działania acylowanych peptydów glukagonu zwiększa się w porównaniu do nieacylowanego wariantu peptydów glukagonu. [0181] W niektórych przykładach wykonania wynalazek dostarcza peptyd glukagonu zmodyfikowany tak, że zawiera grupę acylową lub grupę alkilową połączoną kowalencyjnie z aminokwasem w położeniu peptydu glukagonu. Peptyd glukagonu może ponadto zawierać wstawkę między aminokwasem w położeniu peptydu glukagonu a grupą acylową lub grupą alkilową. W niektórych przykładach wykonania grupa acylowa stanowi kwas tłuszczowy lub kwas żółciowy bądź ich sole, np. kwas tłuszczowy C4 do C30, kwas tłuszczowy C8 do C24, kwas cholowy, C4 do C30 alkil, C8 do C24 alkil, lub alkil zawierający ugrupowanie steroidowe kwasu żółciowego. Wstawka stanowi dowolne ugrupowanie z odpowiednimi grupami reaktywnymi do dołączenia grup acylowych lub alkilowych. W pewnych przykładach wykonania wstawka zawiera aminokwas, dipeptyd, tripeptyd lub stanowi wstawkę dwufunkcyjną hydrofilową lub dwufunkcyjną hydrofobową. W niektórych przykładach wykonania wstawkę wybiera się z grupy składającej się
96 z: Trp, Glu, Asp, Cys i wstawki zawierającej NH2(CH2CH2O)n(CH2)mCOOH, przy czym m jest dowolną liczbą całkowitą od 1 do 6, a n jest dowolną liczbą całkowitą od 2 do 12. Takie acylowane lub alkilowane peptydy glukagonu mogą ponadto zawierać ugrupowanie hydrofilowe, ewentualnie poli(glikolu etylenowego). Dowolny spośród powyższych peptydów glukagonu może zawierać dwie grupy acylowe lub dwie grupy alkilowe bądź ich kombinację. [0182] Acylowanie można przeprowadzić w dowolnym położeniu w obrębie peptydu glukagonu, w tym dowolnym z położeń 1-29, w położeniu w obrębie przedłużenia C- końcowego lub na aminokwasie N- lub C-końcowym, pod warunkiem, że aktywność GIP (i ewentualnie aktywność GLP-1 i(lub) glukagonu) zostaje zachowana, a nawet zwiększona. Acylowanie może nastąpić na przykład na dowolnym aminokwasie dodawanym do sekwencji glukagonu (NR. ID. SEKW.: 1), np. na końcu N lub C. Do nieograniczających przykładów należą położenia 1,,, 11, 12, 13, 14, 16, 17, 18, 19,, 21, 24, 27, 28, 29, 30, 31, 32, 33, 34, 3, 36, 37, 38, 39, 40, 41, 42, 43, 44, 4, 46, 47, 48, 49 lub 0 w peptydzie glukagonu. Grupa acylowa może być połączona kowalencyjnie bezpośrednio z aminokwasem w peptydzie glukagonu lub pośrednio z aminokwasem w peptydzie glukagonu przez wstawkę, przy czym wstawka znajduje się między aminokwasem w peptydzie glukagonu a grupą acylową. Peptydy glukagonu mogą być acylowane w tym samym położeniu aminokwasu, w którym dołączone jest ugrupowanie hydrofilowe, lub w innym położeniu aminokwasu. Do nieograniczających przykładów należy
97 acylowanie w położeniu lub położeniu 40 i pegylowanie w jednym lub większej liczbie położeń w części C-końcowej peptydu glukagonu, np. w położeniu 24, 28 lub 29 w obrębie przedłużenia C-końcowego lub na końcu C (np. przez dodanie C-końcowej Cys). [0183] W niektórych przykładach wykonania peptyd glukagonu modyfikuje się tak, że zawiera przedłużenie o długości około 1 do około 21 aminokwasów po stronie C- końcowej peptydu glukagonu według NR. ID. SEKW.: 1 lub jego analogu i co najmniej jeden z aminokwasów w przedłużeniu jest acylowany lub alkilowany. Na przykład zmodyfikowany peptyd glukagonu może zawierać przedłużenie o długości około 1 do około 21 aminokwasów po stronie C-końcowej aminokwasu w położeniu 29 peptydu glukagonu według NR. ID. SEKW.: 1 lub jego analogu. Ewentualnie, jeśli peptyd glukagonu lub jego analog jest skrócony o jeden lub dwa aminokwasy, przedłużenie o długości około 1 do około 21 aminokwasów może znajdować się po stronie C-końcowej aminokwasu w położeniu 27 lub 28 peptydu glukagonu lub jego analogu. W związku z tym aminokwas acylowany lub alkilowany w obrębie przedłużenia C-końcowego może na przykład stanowić dowolny spośród aminokwasów w położeniu 28, 29, 30, 31, 32, 33, 34, 3, 36, 37, 38, 39, 40, 41, 42, 43, 44, 4, 46, 47, 48, 49 lub 0 peptydu glukagonu z przedłużeniem C-końcowym. W niektórych przykładach wykonania przedłużenie C-końcowe zawiera sekwencję aminokwasową według NR. ID. SEKW.: 9 lub 96. W niektórych przykładach wykonania peptyd glukagonu zawiera przedłużenie C-końcowe zawierające sekwencję aminokwasową według NR. ID. SEKW.: 9 oraz 1 do 11
98 dodatkowych aminokwasów na końcu C według NR. ID. SEKW.: 9, który to dodatkowy aminokwas lub aminokwasy są acylowane lub alkilowane, zgodnie z opisem w niniejszym dokumencie. W konkretnych przykładach wykonania aminokwas acylowany lub alkilowany stanowi resztę Dab, Orn, Lys lub homolys i znajduje się w położeniu 40 w obrębie przedłużenia C-końcowego w peptydzie glukagonu lub jego analogu. [0184] Zgodnie z jednym przykładem wykonania peptyd glukagonu jest zmodyfikowany tak, że zawiera grupę acylową dołączoną do peptydu glukagonu poprzez wiązanie estrowe, tioestrowe lub amidowe na potrzeby wydłużenia okresu półtrwania w krążeniu i(lub) opóźnienia początku działania i(lub) wydłużenia okresu działania i(lub) zwiększenia odporności na proteazy, takie jak DPP-IV. [018] W konkretnym aspekcie wynalazku peptyd glukagonu jest zmodyfikowany tak, że zawiera grupę acylową w wyniku acylowania bezpośredniego grupy aminowej, hydroksylowej lub tiolowej w łańcuchu bocznym aminokwasu w peptydzie glukagonu. W niektórych przykładach wykonania peptyd glukagonu ulega acylowaniu bezpośredniemu poprzez grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym aminokwasu. W niektórych przykładach wykonania acylowanie odbywa się w położeniu,, 24, 29 lub 40. W odniesieniu do tego acylowany peptyd glukagonu może zawierać sekwencję aminokwasową według NR. ID. SEKW.: 1 lub sekwencję aminokwasową zmodyfikowaną względem niej, zawierającą jedną lub większą liczbę spośród modyfikacji aminokwasów opisanych w niniejszym dokumencie, przy czym co najmniej jeden spośród aminokwasów w położeniach,
99 98, 24, 29 i 40 jest zmodyfikowany na dowolny aminokwas zawierający grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym. W niektórych konkretnych przykładach wykonania wynalazku acylowanie bezpośrednie peptydu glukagonu odbywa się poprzez grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym aminokwasu w położeniu lub 40. [0186] W niektórych przykładach wykonania aminokwas zawierający grupę aminową w łańcuchu bocznym stanowi aminokwas o wzorze I: 1 w którym n = 1 do 4 [wzór I] W pewnych przykładach wykonania aminokwas o wzorze I stanowi aminokwas, w którym n wynosi 4 (Lys) lub n wynosi 3 (Orn). [0187] W innych przykładach wykonania aminokwas zawierający grupę hydroksylową w łańcuchu bocznym stanowi aminokwas o wzorze II: w którym n = 1 do 4 [wzór II]
100 99 W pewnych przykładach wykonania aminokwas o wzorze II stanowi aminokwas, w którym n wynosi 1 (Ser). [0188] W jeszcze innych przykładach wykonania aminokwas zawierający grupę tiolową w łańcuchu bocznym stanowi aminokwas o wzorze III: 1 2 w którym n = 1 do 4 [wzór III] W pewnych przykładach wykonania aminokwas o wzorze II stanowi aminokwas, w którym n wynosi 1 (Cys). [0189] W jeszcze innych przykładach wykonania aminokwas zawierający grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym stanowi aminokwas dipodstawiony zawierający tę samą strukturę o wzorze I, wzorze II lub wzorze III, oprócz tego, że atom wodoru połączony z atomem węgla alfa aminokwasu o wzorze I, wzorze II lub wzorze III jest zastąpiony drugim łańcuchem bocznym. [0190] W jednym przykładzie wykonania wynalazku acylowany peptyd glukagonu zawiera wstawkę między peptydem a grupą acylową. W niektórych przykładach wykonania peptyd glukagonu jest połączony kowalencyjnie ze wstawką, która jest połączona kowalencyjnie z grupą acylową. [0191] Aminokwas, do którego dołączona jest wstawka, może stanowić dowolny aminokwas (np. aminokwas pojedynczo lub podwójnie α-podstawiony) zawierający
101 ugrupowanie umożliwiające połączenie ze wstawką. Na przykład odpowiedni jest aminokwas zawierający łańcuch boczny NH2, -OH lub -COOH (np. Lys, Orn, Ser, Asp lub Glu). W odniesieniu do tego acylowany peptyd glukagonu może zawierać sekwencję aminokwasową według NR. ID. SEKW.: 1 lub sekwencję aminokwasową zmodyfikowaną względem niej, zawierającą jeden lub większą liczbę spośród modyfikacji aminokwasów opisanych w niniejszym dokumencie, przy czym co najmniej jeden spośród aminokwasów w położeniach,, 24, 29 i 40 jest zmodyfikowany na dowolny aminokwas zawierający grupę aminową, hydroksylową lub karboksylanową w łańcuchu bocznym. [0192] W niektórych przykładach wykonania wstawką jest aminokwas zawierający grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym bądź dipeptyd lub tripeptyd zawierający aminokwas zawierający grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym. [0193] Jeśli acylowanie następuje poprzez grupę aminową wstawki, acylowanie takie może odbywać się poprzez grupę alfa-aminową aminokwasu lub grupę aminową w łańcuchu bocznym. W przypadku, w którym grupa alfaaminowa jest acylowana, aminokwas we wstawce może stanowić dowolny aminokwas. Na przykład aminokwas we wstawce może stanowić aminokwas hydrofobowy, np. Gly, Ala, Val, Leu, Ile, Trp, Met, Phe, Tyr, kwas 6- aminoheksanowy, kwas -aminowalerianowy, kwas 7- aminoheptanowy i kwas 8-aminooktanowy. Ewentualnie aminokwas we wstawce może stanowić resztę kwaśną, np. Asp i Glu. [0194] W przypadku, w którym grupa aminowa łańcucha
102 bocznego aminokwasu we wstawce jest acylowana, aminokwas we wstawce stanowi aminokwas zawierający grupę aminową w łańcuchu bocznym, np. aminokwas o wzorze I (np. Lys lub Orn). W tym przypadku jest możliwe, że zarówno grupa alfa-aminowa, jak i grupa aminowa łańcucha bocznego aminokwasu we wstawce jest acylowana, tak że peptyd glukagonu jest diacylowany. Przykłady wykonania wynalazku obejmują takie cząsteczki diacylowane. [019] Jeśli acylowanie następuje poprzez grupę hydroksylową wstawki, aminokwas lub jeden z aminokwasów w dipeptydzie lub tripeptydzie może stanowić aminokwas o wzorze II. W konkretnym pewnym przykładzie wykonania ten aminokwas stanowi Ser. [0196] Jeśli acylowanie następuje poprzez grupę tiolową wstawki, aminokwas lub jeden z aminokwasów w dipeptydzie lub tripeptydzie może stanowić aminokwas o wzorze III. W konkretnym pewnym przykładzie wykonania ten aminokwas stanowi Cys. [0197] W niektórych przykładach wykonania wstawka stanowi hydrofilową wstawkę dwufunkcyjną. W niektórych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera dwie lub więcej grup reaktywnych, np. grupę aminową, hydroksylową, tiolową i karboksylową, bądź dowolne ich kombinacje. W niektórych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera grupę hydroksylową i karboksylan. W innych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera grupę aminową i karboksylan. W innych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera grupę tiolową i karboksylan. W konkretnych przykładach
103 wykonania wstawka zawiera aminopoli(alkiloksy)karboksylan. W odniesieniu do tego wstawka może zawierać na przykład NH2(CH2CH2O)n(CH2)mCOOH, w którym m jest dowolną liczbą całkowitą od 1 do 6, a n jest dowolną liczbą całkowitą od 2 do 12, taki jak np. kwas 8-amino-3,6- dioksaoktanowy, dostępny w sprzedaży od Peptides International, Inc. (Louisville, KY). [0198] W niektórych przykładach wykonania wstawka stanowi hydrofobową wstawkę dwufunkcyjną. Hydrofobowe wstawki dwufunkcyjne są znane w stanie techniki. Patrz np. Bioconjugate Techniques, G. T. Hermanson (Academic Press, San Diego, CA, 1996). W niektórych przykładach wykonania hydrofobowa wstawka dwufunkcyjna zawiera dwie lub więcej grup reaktywnych, np. grupę aminową, hydroksylową, tiolową i karboksylową, bądź dowolne ich kombinacje. W niektórych przykładach wykonania hydrofobowa wstawka dwufunkcyjna zawiera grupę hydroksylową i karboksylan. W innych przykładach wykonania hydrofobowa wstawka dwufunkcyjna zawiera grupę aminową i karboksylan. W innych przykładach wykonania hydrofobowa wstawka dwufunkcyjna zawiera grupę tiolową i karboksylan. Odpowiednie hydrofobowe wstawki dwufunkcyjne zawierające karboksylan i grupę hydroksylową lub grupę tiolową są znane w stanie techniki i należą do nich na przykład kwas 8- hydroksyoktanowy i kwas 8-merkaptooktanowy. [0199] W niektórych przykładach wykonania wstawka dwufunkcyjna nie stanowi kwasu dikarboksylowego zawierającego nierozgałęzione grupy metylenowe o długości 1-7 atomów węgla między grupami
104 karboksylanowymi. W niektórych przykładach wykonania wstawka dwufunkcyjna stanowi kwas dikarboksylowy zawierający nierozgałęzione grupy metylenowe o długości 1-7 atomów węgla między grupami karboksylanowymi. [00] Wstawka (np. aminokwas, dipeptyd, tripeptyd, hydrofilowa wstawka dwufunkcyjna lub hydrofobowa wstawka dwufunkcyjna) w konkretnych przykładach wykonania ma długość 3 do atomów (np. 6 do atomów (np. 6, 7, 8, 9 lub atomów). W bardziej konkretnych przykładach wykonania wstawka ma długość około 3 do atomów (np. 6 do atomów), a grupa acylowa stanowi tłuszczową grupę acylową C12 do C18, np. tłuszczową grupę acylową C14, tłuszczową grupę acylową C16, tak że całkowita długość wstawki oraz grupy acylowej wynosi 14 do 28 atomów, np. około 14, 1, 16, 17, 18, 19,, 21, 22, 23, 24, 2, 26, 27 lub 28 atomów. W niektórych przykładach wykonania długość wstawki i grupy acylowej wynosi 17 do 28 (np. 19 do 26, 19 do 21) atomów. [01] Według niektórych powyższych przykładów wykonania wstawka dwufunkcyjna może stanowić aminokwas syntetyczny lub występujący naturalnie (w tym ale nieograniczającodowolny spośród opisywanych w niniejszym dokumencie) zawierający szkielet aminokwasowy o długości 3 do atomów (np. kwas 6- aminoheksanowy, kwas -aminowalerianowy, kwas 7- aminoheptanowy i kwas 8-aminooktanowy). Ewentualnie wstawka może stanowić wstawkę dipeptydową lub tripeptydową zawierającą szkielet peptydowy o długości 3 do atomów (np. 6 do atomów). Każdy aminokwas we wstawce dipeptydowej lub tripeptydowej może być taki sam lub inny od pozostałych aminokwasów w dipeptydzie
105 lub tripeptydzie i może być niezależnie wybrany z grupy składającej się z: aminokwasów występujących naturalnie i(lub) niewystępujących naturalnie, w tym na przykład dowolnego spośród izomerów D lub L aminokwasów występujących naturalnie (Ala, Cys, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp, Tyr) lub dowolnego spośród izomerów D lub L aminokwasów niewystępujących naturalnie wybranych z grupy składającej się z: β-alaniny (β-ala), N-αmetyloalaniny (Me-Ala), kwasu aminomasłowego (Abu), kwasu γ-aminomasłowego (γ-abu), kwasu aminoheksanowego (ε-ahx), kwasu aminoizomasłowego (Aib), kwasu aminometylopirolokarboksylowego, kwasu aminopiperydynokarboksylowego, aminoseryny (Ams), kwasu aminotetrahydropirano-4-karboksylowego, arginino-nmetoksy-n-metyloamidu, kwasu β-asparaginowego (β-asp), kwasu azetydynokarboksylowego, 3-(2- benzotiazolilo)alaniny, α-tert-butyloglicyny, kwasu 2- amino--ureido-n-walerianowego (cytrulina, Cit), p- cykloheksyloalaniny (Cha), acetamidometylocysteiny, kwasu diaminobutanowego (Dab), kwasu diaminopropionowego (Dpr), dihydroksyfenyloalaniny (DOPA), dimetylotiazolidyny (DMTA), kwasu γ- glutaminowego (γ-glu), homoseryny (Hse), hydroksyproliny (Hyp), izoleucyno-n-metoksy-nmetyloamidu, metyloizoleucyny (MeIle), kwasu izonipekotynowego (Isn), metyloleucyny (MeLeu), metylolizyny, dimetylolizyny, trimetylolizyny, metanoproliny, sulfotlenku metioniny (Met(O)), metioninosulfonu (Met(O2)), norleucyny (Nle), metylonorleucyny (Me-Nle), norwaliny (Nva), ornityny
106 (Orn), kwasu para-aminobenzoesowego (PABA), penicyloaminy (Pen), metylofenyloalaniny (MePhe), 4- chlorofenyloalaniny (Phe(4-CI)), 4-fluorofenyloalaniny (Phe(4-F)), 4-nitrofenyloalaniny (Phe(4-NO2)), 4- cyjanofenyloalaniny ((Phe(4-CN)), fenyloglicyny (Phg), piperydynyloalaniny, piperydynyloglicyny, 3,4- dehydroproliny, pirolidynyloalaniny, sarkozyny (Sar), selenocysteiny (Sec), O-benzylofosfoseryny, kwasu 4- amino-3-hydroksy-6-metyloheptanowego (Sta), kwasu 4- amino--cykloheksylo-3-hydroksypentanowego (ACHPA), kwasu 4-amino-3-hydroksy--fenylopentanowego (AHPPA), kwasu 1,2,3,4,-tetrahydroizochinolino-3-karboksylowego (Tic), tetrahydropiranoglicyny, tienyloalaniny (Thi), O-benzylofosfotyrozyny, O-fosfotyrozyny, metoksytyrozyny, etoksytyrozyny, O-(bisdimetyloaminofosfono)-tyrozyny, siarczanu tyrozynotetrabutyloaminy, metylowaliny (MeVal) i alkilowanego kwasu 3-merkaptopropionowego. [02] W niektórych przykładach wykonania wstawka zawiera wypadkowy ładunek ujemny, np. zawiera jeden lub dwa aminokwasy naładowane ujemnie. W niektórych przykładach wykonania dipeptyd nie stanowi żadnego z dipeptydów o strukturze ogólnej A-B, w której A jest wybrany z grupy składającej się z Gly, Gln, Ala, Arg, Asp, Asn, Ile, Leu, Val, Phe i Pro, przy czym B jest wybrany z grupy składającej się z Lys, His, Trp. W niektórych przykładach wykonania wstawka dipeptydowa jest wybrana z grupy składającej się z: Ala-Ala, β-alaβ-ala, Leu-Leu, Pro-Pro, kwasu γ-aminomasłowego-kwasu γ-aminomasłowego i γ-glu-γ-glu. [03] W pewnych przykładach wykonania peptyd glukagonu
107 jest zmodyfikowany tak, że zawiera grupę acylową w wyniku acylowania grupy aminowej, hydroksylowej lub tiolowej we wstawce, która to wstawka jest dołączona do łańcucha bocznego aminokwasu w położeniu,, 24, 29 lub 40 lub na aminokwasie C-końcowym w peptydzie glukagonu. [04] W jeszcze bardziej konkretnych przykładach wykonania grupa acylowa jest dołączona do aminokwasu w położeniu lub 40 peptydu glukagonu oraz, ewentualnie, długość wstawki i grupy acylowej wynosi 14 do 28 atomów. Aminokwas w położeniu lub 40 stanowi w niektórych aspektach aminokwas o wzorze I, np. Lys, lub aminokwas dipodstawiony podobny do wzoru I. W bardziej konkretnych przykładach wykonania peptyd glukagonu nie zawiera mostka wewnątrzcząsteczkowego, np. kowalencyjnego mostka wewnątrzcząsteczkowego. Peptyd glukagonu może na przykład stanowić peptyd zawierający jeden lub więcej aminokwasów alfa,alfa-dipodstawionych, np. AIB, do stabilizacji struktury helisy alfa peptydu. Jak pokazano w niniejszym dokumencie, takie peptydy zawierające acylowaną wstawkę dołączoną kowalencyjnie do łańcucha bocznego aminokwasu w położeniu 40 wykazują zwiększoną siłę działania wobec receptorów GIP, GLP-1 i glukagonu. [0] Odpowiednie metody acylowania peptydów poprzez grupy aminowe, hydroksylowe i tiolowe są znane w stanie techniki. Na przykład patrz przykład 19 (dotyczący metod acylowania poprzez aminę), Miller, Biochem Biophys Res Commun 218: (1996); Shimohigashi i Stammer, Int J Pept Protein Res 19: 4-62 (1982); oraz Previero i wsp., Biochim Biophys Acta 263: 7-13 (1972)
108 (dotyczący metod acylowania poprzez hydroksyl); oraz San i Silvius, J Pept Res 66: (0) (dotyczący metod acylowania poprzez tiol); Bioconjugate Chem. Chemical Modifications of Proteins: History and Applications strony 1, 2-12 (1990); Hashimoto i wsp., Pharmacuetical Res. Synthesis of Palmitoyl Derivatives of Insulin and their Biological Activity tom 6, nr 2 str (1989). [06] Grupa acylowa w acylowanym peptydzie glukagonu może mieć dowolną wielkość, np. łańcuch atomów węgla o dowolnej długości, oraz może być liniowa lub rozgałęziona. W pewnych konkretnych przykładach wykonania wynalazku grupa acylowa stanowi kwas tłuszczowy C4 do C30. Na przykład grupa acylowa może stanowić dowolny spośród kwasu tłuszczowego C4, kwasu tłuszczowego C6, kwasu tłuszczowego C8, kwasu tłuszczowego C, kwasu tłuszczowego C12, kwasu tłuszczowego C14, kwasu tłuszczowego C16, kwasu tłuszczowego C18, kwasu tłuszczowego C, kwasu tłuszczowego C22, kwasu tłuszczowego C24, kwasu tłuszczowego C26, kwasu tłuszczowego C28 lub kwasu tłuszczowego C30. W niektórych przykładach wykonania grupa acylowa stanowi kwas tłuszczowy C8 do C, np. kwas tłuszczowy C14 lub kwas tłuszczowy C16. [07] W alternatywnym przykładzie wykonania grupa acylowa stanowi kwas żółciowy. Kwas żółciowy może stanowić dowolny odpowiedni kwas żółciowy, w tym bez ograniczania kwas cholowy, kwas chenodeoksycholowy, kwas deoksycholowy, kwas litocholowy, kwas taurocholowy, kwas glikocholowy i kwas cholesterolowy. [08] W niektórych przykładach wykonania wynalazku
109 peptyd glukagonu jest zmodyfikowany tak, że zawiera grupę acylową w wyniku acylowania alkanu długołańcuchowego przez peptyd glukagonu. W konkretnych aspektach alkan długołańcuchowy zawiera grupę aminową, hydroksylową lub tiolową (np. oktadecyloaminę, tetradekanol i heksadekanotiol), która reaguje z grupą karboksylową lub jej formą aktywowaną w peptydzie glukagonu. Grupa karboksylowa lub jej forma aktywowana w peptydzie glukagonu może stanowić część łańcucha bocznego aminokwasu (np. kwasu glutaminowego, kwasu asparaginowego) w peptydzie glukagonu lub może stanowić część szkieletu peptydowego. [09] W niektórych przykładach wykonania peptyd glukagonu jest zmodyfikowany tak, że zawiera grupę acylową w wyniku acylowania alkanu długołańcuchowego przez wstawkę dołączoną do peptydu glukagonu. W konkretnych aspektach alkan długołańcuchowy zawiera grupę aminową, hydroksylową lub tiolową, która reaguje z grupą karboksylową wstawki, lub jej formą aktywowaną. Odpowiednie wstawki zawierające grupę karboksylową, lub jej formę aktywowaną są opisane w niniejszym dokumencie i obejmują na przykład wstawki dwufunkcyjne, np. aminokwasy, dipeptydy, tripeptydy, hydrofilowe wstawki dwufunkcyjne i hydrofobowe wstawki dwufunkcyjne. [02] W znaczeniu stosowanym w niniejszym dokumencie określenie aktywowana forma grupy karboksylowej oznacza grupę karboksylową mającą wzór ogólny R(C=O)X, w którym X oznacza grupę opuszczającą, a R oznacza peptyd glukagonu lub wstawkę. Na przykład do aktywowanych form grup karboksylowych mogą należeć, ale nieograniczająco, chlorki, bezwodniki i estry acylowe.
110 W niektórych przykładach wykonania aktywowana grupa karboksylowa stanowi ester z grupą opuszczającą estru N-hydroksysukcynimidowego (NHS). [0211] W odniesieniu do tych aspektów wynalazku, w których alkan długołańcuchowy jest acylowany przez peptyd glukagonu lub wstawkę, alkan długołańcuchowy może mieć dowolną wielkość i może zawierać łańcuch atomów węgla o dowolnej długości. Alkan długołańcuchowy może być liniowy lub rozgałęziony. W niektórych aspektach alkan długołańcuchowy stanowi C4 do C30 alkan. Na przykład alkan długołańcuchowy może stanowić dowolny spośród C4 alkanu, C6 alkanu, C8 alkanu, C alkanu, C12 alkanu, C14 alkanu, C16 alkanu, C18 alkanu, C alkanu, C22 alkanu, C24 alkanu, C26 alkanu, C28 alkanu lub C30 alkanu. W niektórych przykładach wykonania alkan długołańcuchowy obejmuje C8 do C alkan, np. C14 alkan, C16 alkan lub C18 alkan. [0212] Ponadto w niektórych przykładach wykonania grupa aminowa, hydroksylowa lub tiolowa peptydu glukagonu jest acylowana przez kwas cholesterolowy. W konkretnych przykładach wykonania peptyd glukagonu jest połączony z kwasem cholesterolowym poprzez zmodyfikowaną wstawkę Cys. [0213] Acylowane peptydy glukagonu opisywane w niniejszym dokumencie mogą być dodatkowo zmodyfikowane tak, żeby zawierały ugrupowanie hydrofilowe. W niektórych konkretnych przykładach wykonania ugrupowanie hydrofilowe może zawierać łańcuch poli(glikolu etylenowego) (PEG). Wprowadzenie ugrupowania hydrofilowego może nastąpić za pomocą dowolnych odpowiednich środków, takich jak dowolny ze
111 sposobów opisywanych w niniejszym dokumencie. W odniesieniu do tego acylowany peptyd glukagonu może zawierać NR. ID. SEKW.: 1, w tym dowolne spośród modyfikacji opisanych w niniejszym dokumencie, przy czym co najmniej jeden spośród aminokwasów w położeniu,, 24, 29 i 40 zawiera grupę acylową i co najmniej jeden spośród aminokwasów w położeniu 16, 17, 21, 24, 29 lub 40, położeniu w obrębie przedłużenia C-końcowego lub aminokwas C-końcowy są zmodyfikowane na Cys, Lys, Orn, homo-cys lub Ac-Phe, a łańcuch boczny tego aminokwasu jest połączony kowalencyjnie z ugrupowaniem hydrofilowym (np. PEG). W niektórych przykładach wykonania grupa acylowa jest dołączona w położeniu lub 40, ewentualnie poprzez wstawkę zawierającą Cys, Lys, Orn, homo-cys lub Ac-Phe, a ugrupowanie hydrofilowe jest wprowadzone na reszcie Cys w położeniu 24. [0214] Ewentualnie acylowany peptyd glukagonu może zawierać wstawkę, przy czym wstawka ta jest zarówno acylowana, jak i zmodyfikowana, tak że zawiera ugrupowanie hydrofilowe. Do nieograniczających przykładów odpowiednich wstawek należą wstawka zawierająca jeden lub więcej aminokwasów wybranych z grupy składającej się z Cys, Ac-Cys, Lys, Orn, homo-cys i Ac-Phe. [021] W konkretnym aspekcie wynalazku acylowany peptyd glukagonu zawiera sekwencję aminokwasową według którejkolwiek z NR. ID. SEKW.: 1-6, , , , , , , , 148, 11, 12, 14, 16, 18, 160, 162, 163, 16, 166, 231, , 27 i 28.
112 [0216] Według niektórych przykładów wykonania peptyd glukagonu jest zmodyfikowany tak, że zawiera grupę alkilową, np. grupę alkilową niewystępującą naturalnie w aminokwasie (np. grupę alkilową, która jest nienatywna w aminokwasie występującym naturalnie). Bez ograniczania się do żadnej konkretnej teorii uważa się, że alkilowanie peptydów glukagonu pozwala uzyskać podobne, a nawet identyczne efekty co acylowanie peptydów glukagonu, np. wydłużony okres półtrwania w krążeniu, opóźniony początek działania, wydłużony okres działania, zwiększoną odporność na proteazy, takie jak DPP-IV, i zwiększoną siłę działania wobec receptorów GLP-1, GIP i glukagonu. [0217] Alkilowanie można przeprowadzić w dowolnych położeniach w obrębie peptydu glukagonu, w tym dowolnym z położeń 1-29, w położeniu w obrębie przedłużenia C- końcowego lub na aminokwasie N- lub C-końcowym, pod warunkiem, że aktywność GIP (i ewentualnie aktywność GIP i(lub) glukagonu) jest zachowana, a nawet zwiększona. Alkilowanie może nastąpić na przykład na dowolnym aminokwasie dodawanym do sekwencji glukagonu (NR. ID. SEKW.: 1), np. na końcu N lub C. Do nieograniczających przykładów należą położenia 1,,, 11, 12, 13, 14, 16, 17, 18, 19,, 21, 24, 27, 28, 29, 30, 31, 32, 33, 34, 3, 36, 37, 38, 39, 40, 41, 42, 43, 44, 4, 46, 47, 48, 49 lub 0. Grupa alkilowa może być połączona kowalencyjnie bezpośrednio z aminokwasem w peptydzie glukagonu lub pośrednio z aminokwasem w peptydzie glukagonu przez wstawkę, przy czym wstawka znajduje się między aminokwasem w peptydzie glukagonu a grupą alkilową. Peptydy glukagonu mogą być alkilowane w
113 tym samym położeniu aminokwasu, w którym dołączone jest ugrupowanie hydrofilowe, lub w innym położeniu aminokwasu. Do nieograniczających przykładów należy alkilowanie w położeniu lub 40 i pegylowanie w jednym lub większej liczbie położeń w części C-końcowej peptydu glukagonu, np. położeniu 24, 28, 29 lub 40 w obrębie przedłużenia C-końcowego lub na końcu C (np. przez dodanie C-końcowej Cys). [0218] W konkretnym aspekcie wynalazku peptyd glukagonu jest zmodyfikowany tak, że zawiera grupę alkilową w wyniku alkilowania bezpośredniego grupy aminowej, hydroksylowej lub tiolowej w łańcuchu bocznym aminokwasu w peptydzie glukagonu. W niektórych przykładach wykonania peptyd glukagonu ulega alkilowaniu bezpośredniemu poprzez grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym aminokwasu. W niektórych przykładach wykonania alkilowanie odbywa się w położeniu,, 24, 29 lub 40. W odniesieniu do tego alkilowany peptyd glukagonu może zawierać sekwencję aminokwasową według NR. ID. SEKW.: 1 lub sekwencję aminokwasową zmodyfikowaną względem niej, zawierającą jeden lub większą liczbę spośród modyfikacji aminokwasów opisanych w niniejszym dokumencie, przy czym co najmniej jeden spośród aminokwasów w położeniach,, 24, 29 i 40 jest zmodyfikowany na dowolny aminokwas zawierający grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym. W niektórych konkretnych przykładach wykonania wynalazku alkilowanie bezpośrednie peptydu glukagonu odbywa się poprzez grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym aminokwasu w położeniu.
114 [0219] W niektórych przykładach wykonania aminokwas zawierający grupę aminową w łańcuchu bocznym stanowi aminokwas o wzorze I. W pewnych przykładach wykonania aminokwas o wzorze I stanowi aminokwas, w którym n wynosi 4 (Lys) lub n wynosi 3 (Orn). [02] W innych przykładach wykonania aminokwas zawierający grupę hydroksylową w łańcuchu bocznym stanowi aminokwas o wzorze II. W pewnych przykładach wykonania aminokwas o wzorze II stanowi aminokwas, w którym n wynosi 1 (Ser). [0221] W jeszcze innych przykładach wykonania aminokwas zawierający grupę tiolową w łańcuchu bocznym stanowi aminokwas o wzorze III. W pewnych przykładach wykonania aminokwas o wzorze III stanowi aminokwas, w którym n wynosi 1 (Cys). [0222] W jeszcze innych przykładach wykonania aminokwas zawierający grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym stanowi aminokwas dipodstawiony zawierający tę samą strukturę o wzorze I, wzorze II lub wzorze III, oprócz tego, że atom wodoru połączony z atomem węgla alfa aminokwasu o wzorze I, wzorze II lub wzorze III jest zastąpiony drugim łańcuchem bocznym. [0223] W jednym przykładzie wykonania wynalazku alkilowany peptyd glukagonu zawiera wstawkę między peptydem a grupą alkilową. W niektórych przykładach wykonania peptyd glukagonu jest połączony kowalencyjnie ze wstawką, która jest połączona kowalencyjnie z grupą alkilową. W pewnych przykładach wykonania peptyd glukagonu jest zmodyfikowany tak, że zawiera grupę alkilową w wyniku alkilowania grupy aminowej, hydroksylowej lub tiolowej we wstawce, która to wstawka
115 jest dołączona do łańcucha bocznego aminokwasu w położeniu,, 24, 29 lub 40 w peptydzie glukagonu. Aminokwas, do którego dołączona jest wstawka, może stanowić dowolny aminokwas (np. aminokwas α-podstawiony pojedynczo lub aminokwas α,α-dipodstawiony) zawierający ugrupowanie umożliwiające połączenie ze wstawką. Na przykład odpowiedni jest aminokwas zawierający łańcuch boczny NH2, -OH lub -COOH (np. Lys, Orn, Ser, Asp lub Glu). W odniesieniu do tego alkilowany peptyd glukagonu może zawierać sekwencję aminokwasową według NR. ID. SEKW.: 1 lub sekwencję aminokwasową zmodyfikowaną względem niej, zawierającą jeden lub większą liczbę spośród modyfikacji aminokwasów opisanych w niniejszym dokumencie, przy czym co najmniej jeden spośród aminokwasów w położeniach,, 24, 29 i 40 jest zmodyfikowany na dowolny aminokwas zawierający grupę aminową, hydroksylową lub karboksylanową w łańcuchu bocznym. [0224] W niektórych przykładach wykonania wstawką jest aminokwas zawierający grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym bądź dipeptyd lub tripeptyd zawierający aminokwas zawierający grupę aminową, hydroksylową lub tiolową w łańcuchu bocznym. [022] Jeśli alkilowanie następuje poprzez grupę aminową wstawki, alkilowanie takie może odbywać się poprzez grupę alfa-aminową aminokwasu lub grupę aminową w łańcuchu bocznym. W przypadku, w którym grupa alfaaminowa jest alkilowana, aminokwas we wstawce może stanowić dowolny aminokwas. Na przykład aminokwas we wstawce może stanowić aminokwas hydrofobowy, np. Gly, Ala, Val, Leu, Ile, Trp, Met, Phe, Tyr, kwas 6-
116 aminoheksanowy, kwas -aminowalerianowy, kwas 7- aminoheptanowy i kwas 8-aminooktanowy. Ewentualnie aminokwas we wstawce może stanowić resztę kwaśną, np. Asp i Glu, pod warunkiem, że alkilowanie następuje na grupie alfa-aminowej reszty kwaśnej. W przypadku, w którym grupa aminowa łańcucha bocznego aminokwasu we wstawce jest alkilowana, aminokwas we wstawce stanowi aminokwas zawierający grupę aminową w łańcuchu bocznym, np. aminokwas o wzorze I (np. Lys lub Orn). W tym przypadku jest możliwe, że zarówno grupa alfa-aminowa, jak i grupa aminowa łańcucha bocznego aminokwasu we wstawce jest alkilowana, tak że peptyd glukagonu jest dialkilowany. Przykłady wykonania wynalazku obejmują takie cząsteczki dialkilowane. [0226] Jeśli alkilowanie następuje poprzez grupę hydroksylową wstawki, aminokwas lub jeden z aminokwasów w dipeptydzie lub tripeptydzie może stanowić aminokwas o wzorze II. W konkretnym pewnym przykładzie wykonania ten aminokwas stanowi Ser. [0227] Jeśli acylowanie następuje poprzez grupę tiolową wstawki, aminokwas lub jeden z aminokwasów w dipeptydzie lub tripeptydzie może stanowić aminokwas o wzorze III. W konkretnym pewnym przykładzie wykonania ten aminokwas stanowi Cys. [0228] W niektórych przykładach wykonania wstawka stanowi hydrofilową wstawkę dwufunkcyjną. W niektórych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera dwie lub więcej grup reaktywnych, np. grupę aminową, hydroksylową, tiolową i karboksylową bądź dowolne ich kombinacje. W niektórych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera
117 grupę hydroksylową i karboksylan. W innych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera grupę aminową i karboksylan. W innych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera grupę tiolową i karboksylan. W konkretnym przykładzie wykonania wstawka zawiera aminopoli(alkiloksy)karboksylan. W odniesieniu do tego wstawka może zawierać na przykład NH2(CH2CH2O)n(CH2)mCOOH, w którym m jest dowolną liczbą całkowitą od 1 do 6, a n jest dowolną liczbą całkowitą od 2 do 12, taki jak np. kwas 8-amino-3,6- dioksaoktanowy, dostępny w sprzedaży od Peptides International, Inc. (Louisville, KY). [0229] W niektórych przykładach wykonania wstawka stanowi hydrofobową wstawkę dwufunkcyjną. W niektórych przykładach wykonania hydrofobowa wstawka dwufunkcyjna zawiera dwie lub więcej grup reaktywnych, np. grupę aminową, hydroksylową, tiolową i karboksylową bądź dowolne ich kombinacje. W niektórych przykładach wykonania hydrofobowa wstawka dwufunkcyjna zawiera grupę hydroksylową i karboksylan. W innych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera grupę aminową i karboksylan. W innych przykładach wykonania hydrofilowa wstawka dwufunkcyjna zawiera grupę tiolową i karboksylan. Odpowiednie hydrofobowe wstawki dwufunkcyjne zawierające karboksylan i grupę hydroksylową lub grupę tiolową są znane w stanie techniki i należą do nich na przykład kwas 8- hydroksyoktanowy i kwas 8-merkaptooktanowy. [0230] Wstawka (np. aminokwas, dipeptyd, tripeptyd, hydrofilowa wstawka dwufunkcyjna lub hydrofobowa
118 wstawka dwufunkcyjna) w konkretnych przykładach wykonania ma długość 3 do atomów (np. 6 do atomów (np. 6, 7, 8, 9 lub atomów)). W bardziej konkretnych przykładach wykonania wstawka ma długość około 3 do atomów (np. 6 do atomów), a alkil stanowi grupę C12 do C18 alkilową, np. grupę alkilową C14, grupę alkilową C16, tak że całkowita długość wstawki oraz grupy alkilowej wynosi 14 do 28 atomów, np. około 14, 1, 16, 17, 18, 19,, 21, 22, 23, 24, 2, 26, 27 lub 28 atomów. W niektórych przykładach wykonania długość wstawki i grupy alkilowej wynosi 17 do 28 (np. 19 do 26, 19 do 21) atomów. [0231] Według niektórych powyższych przykładów wykonania wstawka dwufunkcyjna może stanowić aminokwas syntetyczny lub niewystępujący naturalnie, zawierający szkielet aminokwasowy o długości 3 do atomów (np. kwas 6-aminoheksanowy, kwas -aminowalerianowy, kwas 7- aminoheptanowy i kwas 8-aminooktanowy). Ewentualnie wstawka może stanowić wstawkę w postaci dipeptydu lub tripeptydu zawierającego szkielet peptydowy o długości 3 do atomów (np. 6 do atomów). Wstawka dipeptydowa lub tripeptydowa może składać się z aminokwasów występujących naturalnie i(lub) niewystępujących naturalnie, w tym na przykład dowolnego spośród aminokwasów wymienionych w niniejszym dokumencie. W niektórych przykładach wykonania wstawka zawiera wypadkowy ładunek ujemny, np. zawiera jeden lub dwa aminokwasy naładowane ujemnie. W niektórych przykładach wykonania wstawkę dipeptydową wybiera się z grupy składającej się z: Ala-Ala, β-ala-β-ala, Leu-Leu, Pro-Pro, kwasu γ-aminomasłowego-kwasu γ-aminomasłowego
119 i γ-glu-γ-glu. [0232] Odpowiednie metody alkilowania peptydów poprzez grupy aminowe, hydroksylowe i tiolowe są znane w stanie techniki. Na przykład do wytworzenia wiązania eterowego między grupą hydroksylową w peptydzie glukagonu a grupą alkilową można zastosować syntezę eterów Williamsona. Ponadto reakcja substytucji nukleofilowej peptydu za pomocą halogenku alkilu może dać dowolne spośród wiązania eterowego, tioeterowego lub aminowego. [0233] Grupa alkilowa w alkilowanym peptydzie glukagonu może mieć dowolną wielkość, np. łańcuch atomów węgla o dowolnej długości, oraz może być liniowa lub rozgałęziona. W niektórych przykładach wykonania wynalazku grupa alkilowa stanowi C4 do C30 alkil. Na przykład grupa alkilowa może stanowić dowolny spośród C4 alkilu, C6 alkilu, C8 alkilu, C alkilu, C12 alkilu, C14 alkilu, C16 alkilu, C18 alkilu, C alkilu, C22 alkilu, C24 alkilu, C26 alkilu, C28 alkilu lub C30 alkilu. W niektórych przykładach wykonania wynalazku grupa alkilowa stanowi C8 do C alkil, np. C14 alkil lub C16 alkil. [0234] W niektórych konkretnych przykładach wykonania grupa alkilowa zawiera ugrupowanie steroidowe kwasu żółciowego, np. kwasu cholowego, kwasu chenodeoksycholowego, kwasu deoksycholowego, kwasu litocholowego, kwasu taurocholowego, kwasu glikocholowego i kwasu cholesterolowego. [023] W niektórych przykładach wykonania wynalazku peptyd glukagonu jest zmodyfikowany tak, żeby zawierał grupę alkilową w wyniku reakcji nukleofilowego alkanu długołańcuchowego z peptydem glukagonu, przy czym
120 peptyd glukagonu zawiera grupę opuszczającą odpowiednią do substytucji nukleofilowej. W konkretnych aspektach grupa nukleofilowa alkanu długołańcuchowego zawiera grupę aminową, hydroksylową lub tiolową (np. oktadecyloaminę, tetradekanol i heksadekanotiol). Grupa opuszczająca w peptydzie glukagonu może stanowić część łańcucha bocznego aminokwasu lub może stanowić część szkieletu peptydowego. Do odpowiednich grup opuszczających należą na przykład N- hydroksysukcynoimid, halogenowce i estry sulfonianowe. [0236] W niektórych przykładach wykonania peptyd glukagonu jest zmodyfikowany tak, że zawiera grupę alkilową w wyniku reakcji nukleofilowego alkanu długołańcuchowego ze wstawką dołączoną do peptydu glukagonu, przy czym wstawka zawiera grupę opuszczającą. W konkretnych aspektach alkan długołańcuchowy zawiera grupę aminową, hydroksylową lub tiolową. W niektórych przykładach wykonania wstawka zawierająca grupę opuszczającą może stanowić dowolną spośród wstawek omawianych w niniejszym dokumencie, np. aminokwasy, dipeptydy, tripeptydy, hydrofilowe wstawki dwufunkcyjne i hydrofobowe wstawki dwufunkcyjne zawierające ponadto odpowiednią grupę opuszczającą. [0237] W odniesieniu do tych aspektów wynalazku, w których alkan długołańcuchowy jest alkilowany przez peptyd glukagonu lub wstawkę, alkan długołańcuchowy może mieć dowolną wielkość i może zawierać łańcuch atomów węgla o dowolnej długości. Alkan długołańcuchowy może być liniowy lub rozgałęziony. W niektórych aspektach alkan długołańcuchowy stanowi C4 do C30 alkan. Na przykład alkan długołańcuchowy może stanowić
121 dowolny spośród C4 alkanu, C6 alkanu, C8 alkanu, C alkanu, C12 alkanu, C14 alkanu, C16 alkanu, C18 alkanu, C alkanu, C22 alkanu, C24 alkanu, C26 alkanu, C28 alkanu, lub C30 alkanu. W niektórych przykładach wykonania alkan długołańcuchowy obejmuje C8 do C alkan, np. C14 alkan, C16 alkan, lub C18 alkan. [0238] Ponadto w niektórych przykładach wykonania alkilowanie może nastąpić między peptydem glukagonu a ugrupowaniem cholesterolu. Na przykład grupa hydroksylowa cholesterolu może zastąpić grupę opuszczającą w alkanie długołańcuchowym tworząc produkt stanowiący cholesterolopeptyd glukagonu. [0239] Alkilowane peptydy glukagonu opisywane w niniejszym dokumencie mogą być dodatkowo modyfikowane tak, żeby zawierały ugrupowanie hydrofilowe. W niektórych konkretnych przykładach wykonania ugrupowanie hydrofilowe może zawierać łańcuch poli(glikolu etylenowego) (PEG). Wprowadzenie ugrupowania hydrofilowego może nastąpić za pomocą dowolnych odpowiednich metod, na przykład dowolnej z metod opisywanych w niniejszym dokumencie. W odniesieniu do tego alkilowany peptyd glukagonu może zawierać NR. ID. SEKW.: 1 lub sekwencję aminokwasową zmodyfikowaną względem niej, zawierającą jeden lub większą liczbę spośród modyfikacji aminokwasów opisanych w niniejszym dokumencie, w których co najmniej jeden z aminokwasów w położeniu,, 24, 29, i 40 zawiera grupę alkilową i co najmniej jeden z aminokwasów w położeniu 16, 17, 21, 24, 29 i 40, w położeniu w przedłużeniu C-końcowym lub aminokwas C- końcowy są zmodyfikowane na Cys, Lys, Orn, homo-cys lub
122 Ac-Phe, a łańcuch boczny tego aminokwasu jest połączony kowalencyjnie z ugrupowaniem hydrofilowym (np. PEG). W niektórych przykładach wykonania grupa alkilowa jest dołączona w położeniu lub 40, ewentualnie poprzez wstawkę zawierającą Cys, Lys, Orn, homo-cys lub Ac-Phe, a ugrupowanie hydrofilowe zostaje wprowadzone na reszcie Cys w położeniu 24. [0240] Ewentualnie alkilowany peptyd glukagonu może zawierać wstawkę, przy czym wstawka ta jest zarówno alkilowana, jak i zmodyfikowana, tak żeby zawierała ugrupowanie hydrofilowe. Do nieograniczających przykładów odpowiednich wstawek należą wstawka zawierająca jeden lub więcej aminokwasów wybranych z grupy składającej się z Cys, Lys, Orn, homo-cys i Ac- Phe. Przykłady wykonania [0241] Zgodnie z niektórymi przykładami wykonania analog glukagonu (NR. ID. SEKW.: 1) mający aktywność agonistyczną wobec GIP zawiera NR. ID. SEKW.: 1 z (a) modyfikacją aminokwasu w położeniu 1 nadającą aktywność agonistyczną wobec GIP, (b) modyfikacją stabilizującą strukturę helisy alfa części C-końcowej (aminokwasy 12-29) analogu, i (c) ewentualnie 1 do (np. 1, 2, 3, 4,, 6, 7, 8, 9, ) dodatkowych modyfikacji aminokwasów. W niektórych przykładach wykonania analog wykazuje co najmniej około 1% aktywności natywnego GIP wobec receptora GIP lub dowolny inny poziom aktywności wobec receptora GIP zgodnie z opisem w niniejszym dokumencie. [0242] W niektórych przykładach wykonania modyfikacją stabilizującą strukturę helisy alfa jest modyfikacja zapewniająca lub wprowadzająca mostek
123 wewnątrzcząsteczkowy, w tym na przykład kowalencyjny mostek wewnątrzcząsteczkowy, taki jak dowolny z opisanych w niniejszym dokumencie. Kowalencyjnym mostkiem wewnątrzcząsteczkowym w niektórych przykładach wykonania jest mostek laktamowy. Mostek laktamowy w analogu tych przykładów wykonania może stanowić mostek laktamowy zgodnie z opisem w niniejszym dokumencie. Patrz np. informacje dotyczące mostków laktamowych w części Stabilizacja struktury helisy alfa. Na przykład mostek laktamowy może stanowić mostek znajdujący się między łańcuchami bocznymi aminokwasów w położeniach i oraz i+4 lub między łańcuchami bocznymi aminokwasów w położeniach j i j+3, przy czym i wynosi 12, 13, 16, 17, lub 24, i przy czym j wynosi 17. W niektórych przykładach wykonania mostek laktamowy może znajdować się między aminokwasami w położeniach 16 i, przy czym jeden spośród aminokwasów w położeniach 16 i jest podstawiony przez Glu, a drugi spośród aminokwasów w położeniach 16 i jest podstawiony przez Lys. [0243] W alternatywnych przykładach wykonania modyfikacja stabilizująca strukturę helisy alfa stanowi wprowadzenie jednego, dwóch, trzech lub czterech aminokwasów α,α-dipodstawionych w położeniu lub położeniach 16,, 21 i 24 w analogu. W niektórych przykładach wykonania aminokwas α,α-dipodstawiony stanowi AIB. W niektórych aspektach aminokwas α,αdipodstawiony (np. AIB) występuje w położeniu, a aminokwas w położeniu 16 jest podstawiony przez aminokwas naładowany dodatnio, taki jak na przykład aminokwas o wzorze IV, opisany w niniejszym dokumencie.
124 Aminokwas o wzorze IV może stanowić homolys, Lys, Orn lub kwas 2,4-diaminomasłowy (Dab). [0244] W konkretnych aspektach wynalazku modyfikacja aminokwasu w położeniu 1 stanowi substytucję His przez aminokwas niezawierający imidazolowego łańcucha bocznego, np. duży aminokwas aromatyczny (np. Tyr). [024] W niektórych aspektach analog glukagonu zawiera modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29. Na przykład Met w położeniu 27 może być podstawiona przez duży aminokwas alifatyczny, ewentualnie Leu, Asn w położeniu 28 może zostać podstawiona przez mały aminokwas alifatyczny, ewentualnie Ala, a Thr w położeniu 29 może być podstawiona przez mały aminokwas alifatyczny, ewentualnie Gly, bądź kombinację dwóch lub trzech spośród powyższych. W konkretnych przykładach wykonania analog glukagonu zawiera Leu w położeniu 27, Ala w położeniu 28 i Gly lub Thr w położeniu 29. [0246] W niektórych przykładach wykonania wynalazku analog glukagonu zawiera przedłużenie o długości 1 do 21 aminokwasów po stronie C-końcowej aminokwasu w położeniu 29. Przedłużenie to może zawierać sekwencję aminokwasową według sekwencji na przykład NR. ID. SEKW.: 9 lub 96. Dodatkowo lub alternatywnie analog glukagonu może zawierać przedłużenie, przy czym 1-6 aminokwasów w przedłużeniu stanowią aminokwasy naładowane dodatnio. Aminokwasy naładowane dodatnio mogą stanowić aminokwasy o wzorze IV, w tym ale nieograniczająco, Lys, homolys, Orn i Dab. [0247] Analog glukagonu w niektórych przykładach wykonania jest acylowany lub alkilowany zgodnie z
125 opisem w niniejszym dokumencie. Na przykład grupa acylowa lub alkilowa może być dołączona do analogu glukagonu ze wstawką lub bez niej, w położeniu lub 40 analogu, zgodnie z poniższym opisem w niniejszym dokumencie. Analog może być dodatkowo lub alternatywnie zmodyfikowany tak, żeby zawierał ugrupowanie hydrofilowe, zgodnie z poniższym opisem w niniejszym dokumencie. Ponadto w niektórych przykładach wykonania analog zawiera dowolną jedną lub kombinację następujących modyfikacji: (a) Ser w położeniu 2 podstawiona przez D-Ser, Ala, D- Ala, Gly, N-metylo-Ser, AIB, Val lub kwas α-amino-nmasłowy; (b) Tyr w położeniu podstawiona przez Trp, Lys, Orn, Glu, Phe lub Val; (c) połączenie grupy acylowej z Lys w położeniu ; (d) Lys w położeniu 12 podstawiona przez Arg lub Ile; (e) Ser w położeniu 16 podstawiona przez Glu, Gln, kwas homoglutaminowy, kwas homocysteinowy, Thr, Gly lub AIB; (f) Arg w położeniu 17 podstawiona przez Gln; (g) Arg w położeniu 18 podstawiona przez Ala, Ser, Thr, lub Gly; (h) Gln w położeniu podstawiona przez Ser, Thr, Ala, Lys, cytrulinę, Arg, Orn lub AIB; (i) Asp w położeniu 21 podstawiona przez Glu, kwas homoglutaminowy, kwas homocysteinowy; (j) Val w położeniu 23 podstawiona przez Ile; (k) Gln w położeniu 24 podstawiona przez Asn, Ser, Thr, Ala lub AIB; (l) oraz substytucja konserwatywna w dowolnym z położeń 2,, 9,, 11, 12, 13, 14, 1, 16, 8, 19,, 21, 24,
126 , 28 i 29. [0248] W pewnych przykładach wykonania analog glukagonu (NR. ID. SEKW.: 1) mający działanie agonistyczne wobec GIP zawiera następujące modyfikacje: (a) modyfikacja aminokwasu w położeniu 1 nadająca aktywność agonistyczną wobec GIP, (b) mostek laktamowy między łańcuchami bocznymi aminokwasów w położeniach i oraz i+4 lub między łańcuchami bocznymi aminokwasów w położeniach j i j+3, przy czym i wynosi 12, 13, 16, 17, lub 24 i przy czym j wynosi 17, (c) modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29, np. modyfikacje aminokwasów w położeniu 27 i(lub) 28, oraz (d) 1-9 lub 1-6 dodatkowych modyfikacji aminokwasów, np. 1, 2, 3, 4,, 6, 7, 8 lub 9 dodatkowych modyfikacji aminokwasów, a wartość EC0 analogu wobec aktywacji receptora GIP wynosi około nm lub mniej. [0249] Mostek laktamowy w analogu z tych przykładów wykonania może stanowić mostek laktamowy zgodnie z opisem w niniejszym dokumencie. Patrz np. informacje dotyczące mostków laktamowych w części Stabilizacja struktury helisy alfa. Na przykład mostek laktamowy może znajdować się między aminokwasami w położeniach 16 i, przy czym jeden spośród aminokwasów w położeniach 16 i jest podstawiony przez Glu, a drugi spośród aminokwasów w położeniach 16 i jest podstawiony przez Lys. [0] Według tych przykładów wykonania analog może zawierać na przykład sekwencję aminokwasową według którejkolwiek z NR. ID. SEKW.: -94.
127 [021] W innych niektórych przykładach wykonania analog glukagonu (NR. ID. SEKW.: 1) mający działanie agonistyczne wobec GIP zawiera następujące modyfikacje: (a) modyfikacja aminokwasu w położeniu 1 nadająca aktywność agonistyczną wobec GIP, (b) jeden, dwa, trzy lub wszystkie spośród aminokwasów w położeniach 16,, 21 i 24 analogu jest podstawionych przez aminokwas α,α-dipodstawiony, (c) modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29, np. modyfikacje aminokwasów w położeniu 27 i(lub) 28, oraz (d) 1-9 lub 1-6 dodatkowych modyfikacji aminokwasów, np. 1, 2, 3, 4,, 6, 7, 8 lub 9 dodatkowych modyfikacji aminokwasów, a wartość EC0 analogu wobec aktywacji receptora GIP wynosi około nm lub mniej. [022] Aminokwas α,α-dipodstawiony w analogu według tych przykładów wykonania może stanowić dowolny aminokwas α,α-dipodstawiony, w tym ale nieograniczająco kwas aminoizomasłowy (AIB), aminokwas dipodstawiony tą samą lub inną grupą wybraną spośród metylu, etylu, propylu i n-butylu lub cyklooktanem lub cykloheptanem (np. kwas 1-aminocyklooktano-1-karboksylowy). W niektórych przykładach wykonania aminokwas α,αdipodstawiony stanowi AIB. W niektórych przykładach wykonania aminokwas w położeniu jest podstawiony przez aminokwas α,α-dipodstawiony, np. AIB. [023] Według tych przykładów wykonania analog może zawierać na przykład sekwencję aminokwasową według którejkolwiek z NR. ID. SEKW.: , , i
![127 [024] W jeszcze innych niektórych przykładach wykonania analog glukagonu (NR. ID. SEKW.](/docs-images/65/53262204/images/128-0.jpg ": 1) mający działanie agonistyczne wobec GIP zawiera następujące modyfikacje: (a) modyfikacja aminokwasu w położeniu 1 nadająca aktywność agonistyczną wobec GIP, (b) substytucja aminokwasu Ser w")
128 127 [024] W jeszcze innych niektórych przykładach wykonania analog glukagonu (NR. ID. SEKW.: 1) mający działanie agonistyczne wobec GIP zawiera następujące modyfikacje: (a) modyfikacja aminokwasu w położeniu 1 nadająca aktywność agonistyczną wobec GIP, (b) substytucja aminokwasu Ser w położeniu 16 przez aminokwas o wzorze IV: [wzór IV], w którym n wynosi 1 do 16, lub 1 do, lub 1 do 7, lub 1 do 6, lub 2 do 6, przy czym każdy spośród R1 i R2 jest niezależnie wybrany z grupy składającej się z: H, C1-C18 alkil, (C1-C18 alkilo)oh, (C1-C18 alkilo)nh2, (C1-1 2 C18 alkilo)sh, (C0-C4 alkilo)(c3-c6)cykloalkil, (C0-C4 alkilo)(c2-c heterocykl), (C0-C4 alkilo)(c6-c arylo)r7 i (C1-C4 alkilo)(c3-c9 heteroaryl), przy czym R7 oznacza H lub OH, a łańcuch boczny aminokwasu o wzorze IV zawiera wolną grupę aminową, (c) substytucja aminokwasu Gln w położeniu aminokwasem alfa,alfa-dipodstawionym, (d) modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29, np. modyfikacje aminokwasów w położeniu 27 i(lub) 28, oraz (e) 1-9 lub 1-6 dodatkowych modyfikacji aminokwasów, np. 1, 2, 3, 4,, 6, 7, 8 lub 9 dodatkowych
129 modyfikacji aminokwasów, a wartość EC0 analogu wobec aktywacji receptora GIP wynosi około nm lub mniej. [02] Aminokwas o wzorze IV w analogu według tych przykładów wykonania może stanowić dowolny aminokwas, taki jak na przykład aminokwas o wzorze IV, w którym n wynosi 1, 2, 3, 4,, 6, 7, 8, 9,, 11, 12, 13, 14, 1 lub 16. W niektórych przykładach wykonania n wynosi 2, 3, 4 lub, w którym to przypadku aminokwas stanowi odpowiednio Dab, Orn, Lys lub homolys. [026] Aminokwas alfa,alfa-dipodstawiony w analogu według tych przykładów wykonania może stanowić dowolny aminokwas alfa,alfa-dipodstawiony, w tym, ale nieograniczająco, kwas aminoizomasłowy (AIB), aminokwas dipodstawiony tą samą lub inną grupą wybraną spośród metylu, etylu, propylu i n-butylu lub cyklooktanem lub cykloheptanem (np. kwas 1-aminocyklooktano-1- karboksylowy). W niektórych przykładach wykonania aminokwas alfa,alfa-dipodstawiony stanowi AIB. [027] Według tych przykładów wykonania analog może zawierać na przykład sekwencję aminokwasową według którejkolwiek z NR. ID. SEKW.: [028] W jeszcze innych niektórych przykładach wykonania analog glukagonu (NR. ID. SEKW.: 1) mający aktywność agonistyczną wobec GIP zawiera: (a) modyfikację aminokwasu w położeniu 1 nadającą aktywność agonistyczną wobec GIP, oraz (b) przedłużenie o długości około 1 do około 21 aminokwasów po stronie C-końcowej aminokwasu w położeniu 29, przy czym co najmniej jeden z aminokwasów w przedłużeniu jest acylowany lub alkilowany,
130 przy czym wartość EC0 analogu wobec aktywacji receptora GIP wynosi około nm lub mniej. [029] W niektórych przykładach wykonania aminokwas acylowany lub alkilowany stanowi aminokwas o wzorze I, II lub III. W bardziej konkretnych przykładach wykonania aminokwas o wzorze I stanowi Dab, Orn, Lys lub homolys. Ponadto w niektórych przykładach wykonania przedłużenie o długości około 1 do około 21 aminokwasów zawiera sekwencję aminokwasową GPSSGAPPPS (NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.: 96), przy czym X stanowi dowolny aminokwas, lub GPSSGAPPPK (NR. ID. SEKW.: 170) lub XGPSSGAPPPK (NR. ID. SEKW.: 171) lub XGPSSGAPPPSK (NR. ID. SEKW.: 172), w których X stanowi Gly lub mały aminokwas alifatyczny lub niepolarny lub nieco polarny. W niektórych przykładach wykonania około 1 do około 21 aminokwasów może zawierać sekwencje zawierające jedną lub większą liczbę substytucji konserwatywnych względem NR. ID. SEKW.: 9, 96, 170, 171 lub 172. W niektórych przykładach wykonania aminokwas acylowany lub alkilowany znajduje się w położeniu 37, 38, 39, 40, 41, 42 lub 43 analogu z przedłużeniem C-końcowym. W niektórych przykładach wykonania aminokwas acylowany lub alkilowany znajduje się w położeniu 40 analogu z przedłużeniem C-końcowym. [0260] W niektórych przykładach wykonania analog mający aktywność agonistyczną wobec GIP zawiera ponadto modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29, np. modyfikacje aminokwasów w położeniu 27 i(lub) 28. [0261] W dowolnym spośród powyższych przykładów wykonania modyfikacja aminokwasu w położeniu 1 nadająca
131 aktywność agonistyczną wobec GIP może stanowić substytucję His przez aminokwas niezawierający imidazolowego łańcucha bocznego. Modyfikacją aminokwasu w położeniu 1 może na przykład być substytucja His przez duży aminokwas aromatyczny. W niektórych przykładach wykonania duży aminokwas aromatyczny stanowi dowolny spośród opisywanych w niniejszym dokumencie, w tym na przykład Tyr. [0262] Ponadto w odniesieniu do powyższych przykładów wykonania modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29 mogą stanowić dowolne spośród modyfikacji w tych położeniach opisanych w niniejszym dokumencie. Na przykład Met w położeniu 27 może być podstawiona przez duży aminokwas alifatyczny, ewentualnie Leu, Asn w położeniu 28 może być podstawiona przez mały aminokwas alifatyczny, ewentualnie Ala, i(lub) Thr w położeniu 29 może być podstawiona przez mały aminokwas alifatyczny, ewentualnie Gly. Ewentualnie analog może zawierać takie modyfikacje aminokwasów w położeniu 27 i(lub) 28. [0263] Analog według powyższych niektórych przykładów wykonania może ponadto zawierać 1-9 lub 1-6 kolejnych dodatkowych modyfikacji aminokwasów, np. 1, 2, 3, 4,, 6, 7, 8 lub 9 dodatkowych modyfikacji aminokwasów, takich jak na przykład dowolna spośród modyfikacji opisanych w niniejszym dokumencie, które zwiększają lub zmniejszają aktywność wobec dowolnego spośród receptorów GIP, GLP-1 i glukagonu, zwiększają rozpuszczalność, zwiększają czas działania lub okres półtrwania w krążeniu, opóźniają początek działania lub zwiększają stabilność. Analog może ponadto zawierać na
132 przykład modyfikację aminokwasu w położeniu 12, ewentualnie substytucję przez Ile, i(lub) modyfikacje aminokwasów w położeniach 17 i 18, ewentualnie substytucję przez Q w położeniu 17 i A w położeniu 18 i(lub) addycję GPSSGAPPPS (NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.: 96) lub sekwencje zawierające jedną lub większą liczbę substytucji konserwatywnych względem NR. ID. SEKW.: 9 lub 96 na końcu C. Analog może zawierać jedną lub więcej z następujących modyfikacji: (i) Ser w położeniu 2 podstawiona przez D-Ser, Ala, D- Ala, Gly, N-metylo-Ser, AIB, Val lub kwas α-amino-nmasłowy; (ii) Tyr w położeniu podstawiona przez Trp, Lys, Orn, Glu, Phe lub Val; (iii) połączenie grupy acylowej z Lys w położeniu ; (iv) Lys w położeniu 12 podstawiona przez Arg; (v) Ser w położeniu 16 podstawiona przez Glu, Gln, kwas homoglutaminowy, kwas homocysteinowy, Thr, Gly lub AIB; (vi) Arg w położeniu 17 podstawiona przez Gln; (vii) Arg w położeniu 18 podstawiona przez Ala, Ser, Thr, lub Gly; (viii) Gln w położeniu podstawiona przez Ala, Ser, Thr, Lys, cytrulinę, Arg, Orn lub AIB; (ix) Asp w położeniu 21 podstawiona przez Glu, kwas homoglutaminowy, kwas homocysteinowy; (x) Val w położeniu 23 podstawiona przez Ile; (xi) Gln w położeniu 24 podstawiona przez Asn, Ala, Ser, Thr lub AIB; oraz (xii) substytucja konserwatywna w dowolnym spośród położeń 2,, 9,, 11, 12, 13, 14, 1, 16, 17, 18,
133 ,, 21, 24, 27, 28 i 29. Analog w niektórych przykładach wykonania zawiera kombinację modyfikacji (i) do (xii). Alternatywnie lub dodatkowo analog może zawierać modyfikację aminokwasu w położeniu 3 (np. substytucję aminokwasu Gln przez Glu), przy czym analog ma mniej niż 1% aktywności glukagonu wobec receptora glukagonu. Alternatywnie lub dodatkowo analog może zawierać modyfikację aminokwasu w położeniu 7 (np. substytucję aminokwasu Thr przez aminokwas niezawierający grupy hydroksylowej, np. Abu lub Ile), przy czym analog ma mniej niż około % aktywności GLP- 1 wobec receptora GLP-1. [0264] W odniesieniu do niektórych przykładów wykonania analog może być połączony kowalencyjnie z ugrupowaniem hydrofilowym. W niektórych przykładach wykonania analog jest połączony kowalencyjnie z ugrupowaniem hydrofilowym w dowolnym spośród położeń aminokwasów 16, 17,, 21, 24, 29, 40 lub na końcu C. W niektórych przykładach wykonania analog zawiera przedłużenie C- końcowe (np. sekwencję aminokwasową według NR. ID. SEKW.: 9) oraz addycję aminokwasu zawierającego ugrupowanie hydrofilowe, tak że ugrupowanie hydrofilowe jest połączone kowalencyjnie z analogiem w położeniu 40. [026] W niektórych przykładach wykonania ugrupowanie hydrofilowe jest połączone kowalencyjnie z Lys, Cys, Orn, homocysteiną lub acetylofenyloalaniną w analogu. Lys, Cys, Orn, homocysteina lub acetylofenyloalanina może stanowić aminokwas natywny w sekwencji glukagonu (NR. ID. SEKW.: 1) lub może to być aminokwas zastępujący aminokwas natywny w NR. ID. SEKW.: 1. W
![133 niektórych przykładach wykonania, w których ugrupowanie hydrofilowe jest dołączone do Cys, połączenie ugrupowania hydrofilowego może zawierać strukturę 1 [0266] W odniesieniu do analogów](/docs-images/65/53262204/images/134-0.jpg "zawierających ugrupowanie hydrofilowe, takie ugrupowanie hydrofilowe może stanowić dowolne spośród opisywanych w niniejszym dokumencie. Patrz np. treść części Łączenie ugrupowań hydrofilowych.")
134 133 niektórych przykładach wykonania, w których ugrupowanie hydrofilowe jest dołączone do Cys, połączenie ugrupowania hydrofilowego może zawierać strukturę 1 [0266] W odniesieniu do analogów zawierających ugrupowanie hydrofilowe, takie ugrupowanie hydrofilowe może stanowić dowolne spośród opisywanych w niniejszym dokumencie. Patrz np. treść części Łączenie ugrupowań hydrofilowych. W niektórych przykładach wykonania ugrupowanie hydrofilowe stanowi poli(glikol etylenowy)(peg). PEG w niektórych przykładach wykonania ma masę cząsteczkową wynoszącą około 00 daltonów do około daltonów, np. około 000 daltonów do około daltonów. [0267] W odniesieniu do niektórych przykładów wykonania analog może zawierać zmodyfikowany aminokwas, w przypadku którego łańcuch boczny jest połączony kowalencyjnie z grupą acylową lub alkilową (np. grupą acylową lub alkilową, która jest nienatywna w aminokwasie występującym naturalnie). Analog acylowany lub alkilowany może być zgodny z peptydami acylowanymi lub alkilowanymi opisanymi w części Acylowanie i alkilowanie. W niektórych przykładach wykonania grupa
135 acylowa stanowi tłuszczową grupę acylową C4 do C30, taką jak na przykład tłuszczowa grupa acylowa lub alkilowa C, tłuszczowa grupa acylowa lub alkilowa C12, tłuszczowa grupa acylowa lub alkilowa C14, tłuszczowa grupa acylowa lub alkilowa C16, tłuszczowa grupa acylowa lub alkilowa C18, grupa acylowa lub alkilowa C lub grupa acylowa lub alkilowa C22. Grupa acylowa lub alkilowa może być połączona kowalencyjnie z dowolnym aminokwasem w analogu, w tym, ale nieograniczająco z aminokwasem w położeniu lub 40, lub z aminokwasem C-końcowym. W niektórych przykładach wykonania analog zawiera przedłużenie C-końcowe (np. sekwencję aminokwasową według NR. ID. SEKW.: 9) oraz addycję aminokwasu zawierającego grupę acylową lub alkilową, tak że grupa acylowa lub alkilowa jest połączona kowalencyjnie z analogiem w położeniu 40. W niektórych przykładach wykonania grupa acylowa lub alkilowa jest połączona kowalencyjnie z łańcuchem bocznym aminokwasu o wzorze I, II lub III, np. resztą Lys. Grupa acylowa lub alkilowa może być połączona kowalencyjnie z aminokwasem natywnym w sekwencji glukagonu (NR. ID. SEKW.: 1) lub może być połączona z aminokwasem dodanym do sekwencji według NR. ID. SEKW.: 1 lub do sekwencji według NR. ID. SEKW.: 1, a następnie NR. ID. SEKW.: 9 (na końcu N lub C) lub może być połączona z aminokwasem zastępującym aminokwas natywny, np. Tyr w położeniu według NR. ID. SEKW.: 1. [0268] W powyższych niektórych przykładach wykonania, w których analog zawiera grupę acylową lub alkilową, analog taki może być dołączony do grupy acylowej lub alkilowej przez wstawkę, zgodnie z opisem w niniejszym
136 dokumencie. Wstawka może na przykład mieć długość 3 do atomów i może na przykład stanowić aminokwas (np. kwas 6-aminoheksanowy, dowolny aminokwas zgodnie z niniejszym dokumentem), dipeptyd (np. Ala-Ala, βalaβala, Leu-Leu, Pro-Pro, γglu-γglu), tripeptyd bądź hydrofilowy lub hydrofobowy łącznik dwufunkcyjny. W niektórych aspektach całkowita długość wstawki oraz grupy acylowej lub alkilowej wynosi około 14 do około 28 atomów. [0269] W jeszcze innych niektórych przykładach wykonania analog glukagonu mający działanie agonistyczne wobec GIP zawiera sekwencję aminokwasową według którejkolwiek spośród NR. ID. SEKW.: 227, 228, 229 lub 230, która zawiera ponadto następujące modyfikacje: (a) ewentualnie modyfikację aminokwasu w położeniu 1 nadającą aktywność agonistyczną wobec GIP, (b) przedłużenie o długości około 1 do około 21 aminokwasów po stronie C-końcowej aminokwasu w położeniu 29, przy czym co najmniej jeden z aminokwasów w przedłużeniu jest acylowany lub alkilowany, oraz (d) do 6 dalszych modyfikacji aminokwasów, przy czym wartość EC0 analogu wobec aktywacji receptora GIP wynosi około nm lub mniej. [0270] W niektórych aspektach aminokwas acylowany lub alkilowany stanowi aminokwas o wzorze I, II lub III. W bardziej konkretnych przykładach wykonania aminokwas o wzorze I stanowi Dab, Orn, Lys lub homolys. Ponadto w niektórych przykładach wykonania około 1 do około 21 aminokwasów zawiera sekwencję aminokwasową GPSSGAPPPS (NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.:
137 ), przy czym X stanowi dowolny aminokwas, lub GPSSGAPPPK (NR. ID. SEKW.: 170) lub XGPSSGAPPPK (NR. ID. SEKW.: 171) lub XGPSSGAPPPSK (NR. ID. SEKW.: 172), w których X stanowi Gly lub mały aminokwas alifatyczny lub niepolarny lub nieco polarny. W niektórych przykładach wykonania około 1 do około 21 aminokwasów może zawierać sekwencje zawierające jedną lub większą liczbę substytucji konserwatywnych względem NR. ID. SEKW.: 9, 96, 170, 171 lub 172. W niektórych przykładach wykonania aminokwas acylowany lub alkilowany znajduje się w położeniu 37, 38, 39, 40, 41, 42 lub 43 analogu z przedłużeniem C-końcowym. W niektórych przykładach wykonania aminokwas acylowany lub alkilowany znajduje się w położeniu 40 analogu z przedłużeniem C-końcowym. [0271] W dowolnym spośród powyższych przykładów wykonania aminokwas w położeniu 1 nadający aktywność agonistyczną wobec GIP może stanowić aminokwas niezawierający imidazolowego łańcucha bocznego. Aminokwas w położeniu 1 może na przykład stanowić duży aminokwas aromatyczny. W niektórych przykładach wykonania duży aminokwas aromatyczny stanowi dowolny spośród opisywanych w niniejszym dokumencie, w tym na przykład Tyr. [0272] Analog powyższych niektórych przykładów wykonania może ponadto zawierać 1-6 dodatkowych modyfikacji aminokwasów, takich jak na przykład dowolna spośród modyfikacji opisanych w niniejszym dokumencie, które zwiększają lub zmniejszają aktywność wobec dowolnego spośród receptorów GIP, GLP-1 i glukagonu, zwiększają rozpuszczalność, zwiększają czas działania
138 lub okres półtrwania w krążeniu, opóźniają początek działania lub zwiększają stabilność. [0273] W niektórych aspektach analogi glukagonu opisane w powyższym pewnym przykładzie wykonania zawierają dodatkowe modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29. Modyfikacje w tych położeniach mogą stanowić dowolne spośród modyfikacji opisywanych w niniejszym dokumencie w odniesieniu do tych położeń. Na przykład względem NR. ID. SEKW.: 227, 228, 229 lub 230 położenie 27 może być podstawione przez duży aminokwas alifatyczny (np. Leu, Ile lub norleucynę) lub Met, położenie 28 może być podstawione przez inny mały aminokwas alifatyczny (np. Gly lub Ala) lub Asn i(lub) położenie 29 może być podstawione przez inny mały aminokwas alifatyczny (np. Ala lub Gly) lub Thr. Ewentualnie analog może zawierać takie modyfikacje aminokwasów w położeniu 27 i(lub) 28. [0274] Analog może ponadto zawierać jedną lub większą z następujących modyfikacji dodatkowych: (i) aminokwas w położeniu 2 stanowi dowolny spośród D- Ser, Ala, D-Ala, Gly, N-metylo-Ser, AIB, Val lub kwasu α-amino-n-masłowego; (ii) aminokwas w położeniu stanowi Tyr, Trp, Lys, Orn, Glu, Phe lub Val; (iii) połączenie grupy acylowej z Lys w położeniu ; (iv) aminokwas w położeniu 12 stanowi Ile, Lys lub Arg; (v) aminokwas w położeniu 16 stanowi dowolny spośród Ser, Glu, Gln, kwasu homoglutaminowego, kwasu homocysteinowego, Thr, Gly lub AIB; (vi) aminokwas w położeniu 17 stanowi Gln lub Arg; (vii) aminokwas w położeniu 18 stanowi dowolny spośród
139 Ala, Arg, Ser, Thr lub Gly; (viii) aminokwas w położeniu stanowi dowolny spośród Ala, Ser, Thr, Lys, cytruliny, Arg, Orn lub AIB bądź innego aminokwasu alfa,alfa-dipodstawionego; (ix) aminokwas w położeniu 21 stanowi dowolny spośród Glu, Asp, kwasu homoglutaminowego, kwasu homocysteinowego; (x) aminokwas w położeniu 23 stanowi Val lub Ile; (xi) aminokwas w położeniu 24 stanowi dowolny spośród Gln, Asn, Ala, Ser, Thr lub AIB; oraz (xii) jedna lub większa liczba substytucji konserwatywnych w dowolnym spośród położeń 2,, 9,, 11, 12, 13, 14, 1, 16, 17, 18, 19,, 21, 24, 27, 28 i 29. Analog w niektórych przykładach wykonania zawiera kombinację modyfikacji (i) do (xii). Alternatywnie lub dodatkowo analog może zawierać modyfikację aminokwasu w położeniu 3 (np. substytucję aminokwasu Gln przez Glu), przy czym analog ma mniej niż 1% aktywności glukagonu wobec receptora glukagonu. Alternatywnie lub dodatkowo analog może zawierać modyfikację aminokwasu w położeniu 7 (np. substytucję aminokwasu Thr przez aminokwas niezawierający grupy hydroksylowej, np. Abu lub Ile), przy czym analog ma mniej niż około % aktywności GLP- 1 wobec receptora GLP-1. [027] W odniesieniu do niektórych przykładów wykonania analog może być połączony kowalencyjnie z ugrupowaniem hydrofilowym. W niektórych przykładach wykonania analog jest połączony kowalencyjnie z ugrupowaniem hydrofilowym w dowolnym spośród położeń aminokwasów 16, 17,, 21, 24, 29, 40 lub na końcu C. W niektórych

140 139 przykładach wykonania analog zawiera ugrupowanie hydrofilowe połączone kowalencyjnie z analogiem w położeniu 24. [0276] W niektórych przykładach wykonania ugrupowanie hydrofilowe jest połączone kowalencyjnie z Lys, Cys, Orn, homocysteiną lub acetylofenyloalaniną w analogu. Lys, Cys, Orn, homocysteina lub acetylofenyloalanina może stanowić aminokwas natywny w NR. ID. SEKW.: 1, 227, 228, 229 lub 230 lub może stanowić aminokwas podstawiony. W niektórych przykładach wykonania, w których ugrupowanie hydrofilowe jest połączone do Cys, połączenie może zawierać strukturę 1 2 [0277] W odniesieniu do analogów zawierających ugrupowanie hydrofilowe, takie ugrupowanie hydrofilowe może stanowić dowolne spośród opisywanych w niniejszym dokumencie. Patrz np. treść części Łączenie ugrupowań hydrofilowych. W niektórych przykładach wykonania ugrupowanie hydrofilowe stanowi poli(glikol etylenowy)(peg). PEG w niektórych przykładach wykonania ma masę cząsteczkową wynoszącą około 00 daltonów do około daltonów, np. około 000 daltonów do około daltonów. [0278] W odniesieniu do niektórych przykładów wykonania analog może zawierać zmodyfikowany aminokwas w obrębie
141 przedłużenia C-końcowego, w przypadku którego łańcuch boczny jest połączony kowalencyjnie z grupą acylową lub alkilową. Analog acylowany lub alkilowany może być zgodny z peptydami acylowanymi lub alkilowanymi opisanymi w części Acylowanie i alkilowanie. W niektórych przykładach wykonania grupa acylowa stanowi tłuszczową grupę acylową C4 do C30, taką jak na przykład tłuszczowa grupa acylowa lub alkilowa C, tłuszczowa grupa acylowa lub alkilowa C12, tłuszczowa grupa acylowa lub alkilowa C14, tłuszczowa grupa acylowa lub alkilowa C16, tłuszczowa grupa acylowa lub alkilowa C18, grupa acylowa lub alkilowa C lub grupa acylowa lub alkilowa C22. Grupa acylowa lub alkilowa może być połączona kowalencyjnie z dowolnym aminokwasem w analogu, w tym ale nieograniczająco, z aminokwasem w położeniu lub 40, lub z aminokwasem C-końcowym. W niektórych przykładach wykonania grupa acylowa lub alkilowa jest połączona kowalencyjnie z łańcuchem bocznym aminokwasu o wzorze I, II lub III, np. z resztą Lys. Grupa acylowa lub alkilowa jest połączona kowalencyjnie z aminokwasem natywnym w NR. ID. SEKW.: 1, 227, 228, 229 lub 230 lub może być połączona z aminokwasem podstawionym. Grupa acylowa lub alkilowa jest połączona kowalencyjnie z aminokwasem natywnym w NR. ID. SEKW.: 9, 96, 171 lub 172 lub może być połączona z aminokwasem podstawionym. [0279] W powyższych niektórych przykładach wykonania, w których analog zawiera grupę acylową lub alkilową, analog taki może być dołączony do grupy acylowej lub alkilowej przez wstawkę, zgodnie z opisem w niniejszym dokumencie. Wstawka może na przykład mieć długość 3 do
142 atomów i może na przykład stanowić aminokwas (np. kwas 6-aminoheksanowy, dowolny aminokwas zgodnie z opisem w niniejszym dokumencie), dipeptyd (np. Ala-Ala, βala-βala, Leu-Leu, Pro-Pro, γglu-γglu), tripeptyd lub hydrofilowy lub hydrofobowy łącznik dwufunkcyjny. W niektórych aspektach całkowita długość wstawki oraz grupy acylowej lub alkilowej wynosi około 14 do około 28 atomów. [0280] W pewnych bardzo konkretnych przykładach wykonania analog według wynalazku zawiera sekwencję aminokwasową wybraną z grupy składającej się z NR. ID. SEKW.: , , 166, 192-7, i 223 lub wybraną z grupy składającej się z NR. ID. SEKW.: , i 22. [0281] Ponadto do konkretnych przykładów analogów według wynalazku należą,ale bez ograniczania, dowolne spośród wymienionych w tabelach 1-3. [0282] W jeszcze innych niektórych przykładach wykonania analog glukagonu mający działanie agonistyczne wobec GIP zawiera grupę acylową lub alkilową (np. grupę acylową lub alkilową, która jest nienatywna w aminokwasie występującym naturalnie), przy czym grupa acylowa lub alkilowa jest dołączona do wstawki, przy czym (i) wstawka jest dołączona do łańcucha bocznego aminokwasu w położeniu w analogu; lub (ii) analog zawiera przedłużenie o długości 1 do 21 aminokwasów po stronie C-końcowej aminokwasu w położeniu 29, a wstawka jest połączona z łańcuchem bocznym aminokwasu odpowiadającego jednemu z położeń względem NR. ID. SEKW.: 1, przy czym wartość EC0 analogu wobec aktywacji receptora GIP wynosi około
143 142 1 nm lub mniej. [0283] W takich przykładach wykonania analog może zawierać sekwencję aminokwasową według NR. ID. SEKW.: 1 z (i) modyfikacją aminokwasu w położeniu 1 nadającą aktywność agonistyczną wobec GIP, (ii) modyfikacjami aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29, (iii) co najmniej jednym z następujących: (A) analog zawiera mostek laktamowy między łańcuchami bocznymi aminokwasów w położeniach i oraz i+4 lub między łańcuchami bocznymi aminokwasów w położeniach j i j+3, przy czym i wynosi 12, 13, 16, 17, lub 24 i przy czym j wynosi 17; (B) jeden, dwa, trzy lub wszystkie spośród aminokwasów w położeniach 16,, 21 i 24 analogu jest podstawionych przez aminokwas α,α-dipodstawiony; lub (C) analog zawiera (i) substytucję aminokwasu Ser w położeniu 16 przez aminokwas o wzorze IV: [wzór IV], przy czym n wynosi 1 do 7, przy czym każdy spośród R 1 i R2 jest niezależnie wybrany z grupy składającej się z: H, C1-C18 alkil, (C1-C18 alkilo)oh, (C1-C18 alkilo)nh2, (C1-C18 alkilo)sh, (C0-C4 alkilo)(c3-c6)cykloalkil, (C0-C4 2 alkilo)(c2-c heterocykl), (C0-C4 alkilo)(c6-c arylo)r7 i (C1-C4 alkilo)(c3-c9 heteroaryl), przy czym R7 oznacza
144 H lub OH, a łańcuch boczny aminokwasu o wzorze IV zawiera wolną grupę aminową; oraz (ii) substytucję aminokwasu Gln w położeniu aminokwasem alfa,alfadipodstawionym i (iv) do 6 dodatkowych modyfikacji aminokwasów. [0284] Aminokwas alfa,alfa-dipodstawiony w analogu według tych przykładów wykonania może stanowić dowolny aminokwas alfa,alfa-dipodstawiony, w tym bez ograniczania, kwas aminoizomasłowy (AIB), aminokwas dipodstawiony tą samą lub inną grupą wybraną spośród metylu, etylu, propylu i n-butylu lub cyklooktanem lub cykloheptanem (np. kwas 1-aminocyklooktano-1- karboksylowy). W niektórych przykładach wykonania aminokwas alfa,alfa-dipodstawiony stanowi AIB. [028] Aminokwas o wzorze IV w analogu według tych przykładów wykonania może stanowić dowolny aminokwas, taki jak na przykład aminokwas o wzorze IV, w którym n wynosi 1, 2, 3, 4,, 6, 7, 8, 9,, 11, 12, 13, 14, 1 lub 16. W niektórych przykładach wykonania n wynosi 2, 3, 4 lub, w którym to przypadku aminokwas stanowi odpowiednio Dab, Orn, Lys lub homolys. [0286] W dowolnym spośród powyższych przykładów wykonania modyfikacja aminokwasu w położeniu 1 nadająca aktywność agonistyczną wobec GIP może stanowić substytucję His przez aminokwas niezawierający imidazolowego łańcucha bocznego. Modyfikacją aminokwasu w położeniu 1 może na przykład być substytucja His przez duży aminokwas aromatyczny. W niektórych przykładach wykonania duży aminokwas aromatyczny stanowi dowolny spośród opisywanych w niniejszym dokumencie, w tym na przykład Tyr.
145 [0287] Ponadto w odniesieniu do powyższych przykładów wykonania modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29 mogą stanowić dowolne spośród modyfikacji w tych położeniach opisanych w niniejszym dokumencie. Na przykład Met w położeniu 27 może być podstawiona przez duży aminokwas alifatyczny, ewentualnie Leu, Asn w położeniu 28 może być podstawiona przez mały aminokwas alifatyczny, ewentualnie Ala, i(lub) Thr w położeniu 29 może być podstawiona przez mały aminokwas alifatyczny, ewentualnie Gly. Ewentualnie analog może zawierać takie modyfikacje aminokwasów w położeniu 27 i(lub) 28. [0288] Analog według powyższych niektórych przykładów wykonania może ponadto zawierać 1-9 lub 1-6 kolejnych dodatkowych modyfikacji aminokwasów, np. 1, 2, 3, 4,, 6, 7, 8 lub 9 dodatkowych modyfikacji aminokwasów, takich jak na przykład dowolna spośród modyfikacji opisanych w niniejszym dokumencie, które zwiększają lub zmniejszają aktywność wobec dowolnego spośród receptorów GIP, GLP-1 i glukagonu, zwiększają rozpuszczalność, zwiększają czas działania lub okres półtrwania w krążeniu, opóźniają początek działania lub zwiększają stabilność. Analog może ponadto zawierać na przykład modyfikację aminokwasów w położeniu 12, ewentualnie substytucję przez Ile, i(lub) modyfikacje aminokwasów w położeniach 17 i 18, ewentualnie substytucję przez Q w położeniu 17 i A w położeniu 18, i(lub) addycję GPSSGAPPPS (NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.: 96) lub sekwencje zawierające jedną lub większą liczbę substytucji konserwatywnych względem NR. ID. SEKW.: 9 lub 96 na
146 końcu C. Analog może zawierać jedną lub więcej z następujących modyfikacji: (i) Ser w położeniu 2 podstawiona przez D-Ser, Ala, D- Ala, Gly, N-metylo-Ser, AIB, Val lub kwas α-amino-nmasłowy; (ii) Tyr w położeniu podstawiona przez Trp, Lys, Orn, Glu, Phe lub Val; (iii) połączenie grupy acylowej z Lys w położeniu ; (iv) Lys w położeniu 12 podstawiona przez Arg; (v) Ser w położeniu 16 podstawiona przez Glu, Gln, kwas homoglutaminowy, kwas homocysteinowy, Thr, Gly, Lys lub AIB; (vi) Arg w położeniu 17 podstawiona przez Gln; (vii) Arg w położeniu 18 podstawiona przez Ala, Ser, Thr lub Gly; (viii) Gln w położeniu podstawiona przez Ala, Ser, Thr, Lys, cytrulinę, Arg, Orn lub AIB; (ix) Asp w położeniu 21 podstawiona przez Glu, kwas homoglutaminowy, kwas homocysteinowy; (x) Val w położeniu 23 podstawiona przez Ile; (xi) Gln w położeniu 24 podstawiona przez Asn, Ala, Ser, Thr lub AIB; oraz (xii) substytucja konserwatywna w dowolnym spośród położeń 2,, 9,, 11, 12, 13, 14, 1, 16, 17, 18, 19,, 21, 24, 27, 28 i 29. Analog w niektórych przykładach wykonania zawiera kombinację modyfikacji (i) do (xii). Alternatywnie lub dodatkowo analog może zawierać modyfikację aminokwasu w położeniu 3 (np. substytucję aminokwasu Gln przez Glu), przy czym analog ma mniej niż 1% aktywności glukagonu wobec receptora glukagonu. Ewentualnie lub dodatkowo
147 analog może zawierać modyfikację aminokwasu w położeniu 7 (np. substytucję aminokwasu Thr przez aminokwas niezawierający grupy hydroksylowej, np. Abu lub Ile), delecję aminokwasu lub aminokwasów C-końcowych wobec aminokwasu w położeniu 27 lub 28, dzięki czemu otrzymuje się peptyd o długości 27 lub 28 aminokwasów bądź ich kombinację, przy czym analog ma mniej niż około % aktywności GLP-1 wobec receptora GLP-1. [0289] W odniesieniu do niektórych przykładów wykonania analog może być połączony kowalencyjnie z ugrupowaniem hydrofilowym. W niektórych przykładach wykonania analog jest połączony kowalencyjnie z ugrupowaniem hydrofilowym w dowolnym spośród położeń aminokwasów 16, 17,, 21, 24, 29, 40 lub na końcu C. W niektórych przykładach wykonania analog zawiera przedłużenie C- końcowe (np. sekwencję aminokwasową według NR. ID. SEKW.: 9) oraz addycję aminokwasu zawierającego ugrupowanie hydrofilowe, tak że ugrupowanie hydrofilowe jest połączone kowalencyjnie z analogiem w położeniu 40. [0290] W niektórych przykładach wykonania ugrupowanie hydrofilowe jest połączone kowalencyjnie z Lys, Cys, Orn, homocysteiną lub acetylofenyloalaniną w analogu. Lys, Cys, Orn, homocysteina lub acetylofenyloalanina może stanowić aminokwas natywny w sekwencji glukagonu (NR. ID. SEKW.: 1) lub może to być aminokwas zastępujący aminokwas natywny według NR. ID. SEKW.: 1. W niektórych przykładach wykonania, w których ugrupowanie hydrofilowe jest dołączone do Cys, połączenie ugrupowania hydrofilowego może zawierać strukturę
148 [0291] W odniesieniu do analogów zawierających ugrupowanie hydrofilowe, takie ugrupowanie hydrofilowe może stanowić dowolne spośród opisywanych w niniejszym dokumencie. Patrz np. treść części Łączenie ugrupowań hydrofilowych. W niektórych przykładach wykonania ugrupowanie hydrofilowe stanowi poli(glikol etylenowy)(peg). PEG w niektórych przykładach wykonania ma masę cząsteczkową wynoszącą około 00 daltonów do około daltonów, np. około 000 daltonów do około daltonów. [0292] W niektórych przykładach wykonania, w których analog zawiera grupę acylową lub alkilową, dołączoną do analogu przez wstawkę, wstawka może stanowić dowolną wstawkę zgodnie z opisem w niniejszym dokumencie. Wstawka może na przykład mieć długość 3 do atomów i może na przykład stanowić aminokwas (np. kwas 6- aminoheksanowy, dowolny aminokwas opisany w niniejszym dokumencie), dipeptyd (np. Ala-Ala, βala-βala, Leu-Leu, Pro-Pro, γglu-γglu), tripeptyd lub hydrofilowy lub hydrofobowy łącznik dwufunkcyjny. W niektórych aspektach całkowita długość wstawki oraz grupy acylowej lub alkilowej wynosi około 14 do około 28 atomów. [0293] Grupa acylowa lub alkilowa stanowi dowolną grupę acylową lub alkilową zgodnie z opisem w niniejszym
149 dokumencie, na przykład grupę acylową lub alkilową, która jest nienatywna względem aminokwasu występującego naturalnie. W niektórych przykładach wykonania grupa acylowa lub alkilowa stanowi tłuszczową grupę acylową C4 do C30, taką jak na przykład tłuszczowa grupa acylowa lub alkilowa C, tłuszczowa grupa acylowa lub alkilowa C12, tłuszczowa grupa acylowa lub alkilowa C14, tłuszczowa grupa acylowa lub alkilowa C16, tłuszczowa grupa acylowa lub alkilowa C18, grupa acylowa lub alkilowa C lub grupa acylowa lub alkilowa C22 lub grupa alkilowa C4 do C30. W konkretnych przykładach wykonania grupa acylowa stanowi tłuszczową grupę acylową C12 do C18 (np. tłuszczową grupę acylową C14 lub C16). [0294] W niektórych przykładach wykonania przedłużenie o długości około 1 do około 21 aminokwasów po stronie C-końcowej aminokwasu w położeniu 29 analogu zawiera sekwencję aminokwasową GPSSGAPPPS (NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.: 96), w których X stanowi dowolny aminokwas, lub GPSSGAPPPK (NR. ID. SEKW.: 170) lub XGPSSGAPPPK (NR. ID. SEKW.: 171) lub XGPSSGAPPPSK (NR. ID. SEKW.: 172), w których X stanowi Gly lub mały aminokwas alifatyczny lub niepolarny lub nieco polarny. W niektórych przykładach wykonania około 1 do około 21 aminokwasów może zawierać sekwencje zawierające jedną lub większą liczbę substytucji konserwatywnych względem NR. ID. SEKW.: 9, 96, 170, 171 lub 172. W niektórych przykładach wykonania aminokwas acylowany lub alkilowany znajduje się w położeniu 37, 38, 39, 40, 41, 42 lub 43 analogu z przedłużeniem C-końcowym. W niektórych przykładach
150 wykonania aminokwas acylowany lub alkilowany znajduje się w położeniu 40 analogu z przedłużeniem C-końcowym. Kompozycje farmaceutyczne i metody leczenia [029] W niektórych aspektach wynalazek dostarcza kompozycję farmaceutyczną zawierającą dowolny z nowych peptydów glukagonu ujawnianych w niniejszym dokumencie, korzystnie jałową i korzystnie na poziomie czystości co najmniej 90%, 91%, 92%, 93%, 94%, 9%, 96%, 97%, 98% lub 99% oraz dopuszczalny farmaceutycznie rozcieńczalnik, nośnik lub zaróbke. Takie kompozycje mogą zawierać peptyd glukagonu w stężeniu co najmniej A, przy czym A wynosi 0,001 mg/ml, 0,01 mg/ml, 0,1 mg/ml, 0, mg/ml, 1 mg/ml, 2 mg/ml, 3 mg/ml, 4 mg/ml, mg/ml, 6 mg/ml, 7 mg/ml, 8 mg/ml, 9 mg/ml, mg/ml, 11 mg/ml, 12 mg/ml, 13 mg/ml, 14 mg/ml, 1 mg/ml, 16 mg/ml, 17 mg/ml, 18 mg/ml, 19 mg/ml, mg/ml, 21 mg/ml, 22 mg/ml, 23 mg/ml, 24 mg/ml, 2 mg/ml lub więcej. W innych przykładach wykonania takie kompozycje mogą zawierać peptyd glukagonu w stężeniu co najwyżej B, przy czym B wynosi 30 mg/ml, 2 mg/ml, 24 mg/ml, 23, mg/ml, 22 mg/ml, 21 mg/ml, mg/ml, 19 mg/ml, 18 mg/ml, 17 mg/ml, 16 mg/ml, 1 mg/ml, 14 mg/ml, 13 mg/ml, 12 mg/ml, 11 mg/ml mg/ml, 9 mg/ml, 8 mg/ml, 7 mg/ml, 6 mg/ml, mg/ml, 4 mg/ml, 3 mg/ml, 2 mg/ml, 1 mg/ml lub 0,1 mg/ml. W niektórych przykładach wykonania kompozycje mogą zawierać peptyd glukagonu w zakresie stężenia A do B mg/ml, na przykład 0,001 do 30,0 mg/ml. W jednym przykładzie wykonania kompozycje farmaceutyczne zawierają roztwory wodne sterylizowane i ewentualnie przechowywane w różnych pojemnikach. Związki według niniejszego wynalazku można
151 wykorzystywać w niektórych przykładach wykonania do otrzymywania roztworów w postaci gotowych preparatów do wstrzykiwania. W innych przykładach wykonania kompozycje farmaceutyczne zawierają liofilizowany proszek. Kompozycje farmaceutyczne mogą być ponadto pakowane jako część zestawu zawierającego urządzenie jednorazowego użytku do podawania kompozycji pacjentowi. Pojemniki lub zestawy mogą być oznakowane do przechowywania w temperaturze pokojowej otoczenia lub w temperaturze chłodziarki. [0296] Peptydy glukagonu można podawać pacjentowi stosując dowolną standardową drogę podawania, w tym pozajelitowo, na przykład dożylnie, dootrzewnowo, podskórnie lub domięśniowo, dokanałowo, przezskórnie, doodbytniczo, doustnie, donosowo lub wziewnie. W jednym przykładzie wykonania kompozycję podaje się podskórnie lub domięśniowo. [0297] W jednym przykładzie wykonania zestaw zawiera urządzenie do podawania kompozycji glukagonu pacjentowi, np. igłę ze strzykawką, urządzenie w postaci wstrzykiwacza (pena), wstrzykiwacz strumieniowy lub inny wstrzykiwacz bezigłowy. Zestaw może alternatywnie lub dodatkowo zawierać jeden lub więcej pojemników, np. fiolek, probówek, butelek, ampułkostrzykawek pojedynczych lub wielokomorowych, wkładów, pomp infuzyjnych (zewnętrznych lub do wszczepiania), wstrzykiwaczy strumieniowych, wyrobów w postaci napełnionych wstrzykiwaczy i podobnych, ewentualnie zawierających peptyd glukagonu w postaci liofilizowanej lub w roztworze wodnym. Zestawy zawierają ponadto korzystnie instrukcje stosowania. W
152 niektórych przykładach wykonania urządzenie w zestawie stanowi urządzenie do dawkowania aerozolu, przy czym kompozycja znajduje się w urządzeniu do podawania aerozolu. W innym przykładzie wykonania zestaw zawiera strzykawkę i igłę, a w jednym przykładzie wykonania jałowa kompozycja glukagonu znajduje się w strzykawce. [0298] Zgodnie z jednym przykładem wykonania dostarcza się kompozycję farmaceutyczną, przy czym kompozycja ta zawiera analog glukagonu o aktywności GIP według niniejszego ujawnienia lub jego dopuszczalną farmaceutycznie sól i dopuszczalny farmaceutycznie nośnik. Kompozycja farmaceutyczna może zawierać dowolny składnik dopuszczalny farmaceutycznie, w tym na przykład środki zakwaszające, środki dodatkowe, adsorbenty, gazy pędne do aerozolu, środki wypierające powietrze, środki alkalizujące, środki przeciwzbrylające, środki przeciwkrzepliwe, przeciwdrobnoustrojowe środki konserwujące, przeciwutleniacze, środki antyseptyczne, zasady, środki wiążące, środki buforujące, środki chelatujące, środki powlekające, środki barwiące, środki suszące, detergenty, rozcieńczalniki, środki dezynfekcyjne, środki ułatwiające rozpad, środki dyspergujące, środki ułatwiające rozpuszczanie, barwniki, środki zmiękczające, środki emulgujące, stabilizatory emulsji, wypełniacze, środki błonotwórcze, środki wzmacniające smak, środki aromatyzujące, środki zwiększające płynność, środki żelujące, środki granulujące, środki zatrzymujące wilgoć (humektanty), środki smarujące, środki mukoadhezyjne, podłoża do maści, maści, nośniki oleiste, zasady organiczne, podłoża do pastylek,
153 pigmenty, plastyfikatory, środki polerujące, środki konserwujące, środki maskujące, środki przenikające przez skórę, środki solubilizujące, rozpuszczalniki, środki stabilizujące, podłoża do czopków, środki powierzchniowo czynne, surfaktanty, środki ułatwiające tworzenie zawiesiny, środki słodzące, środki terapeutyczne, środki zagęszczające, środki zapewniające toniczność, środki zapobiegające toksyczności, środki zwiększające lepkość, środki pochłaniające wodę, współrozpuszczalniki mieszające się z wodą, środki do zmiękczania wody lub środki zwilżające. [0299] W niektórych przykładach wykonania kompozycja farmaceutyczna zawiera dowolny jeden lub kombinację następujących składników: akacja, acesulfam potasu, cytrynian acetylotributylu, cytrynian acetylotrietylu, agar, albumina, alkohol, alkohol bezwodny, alkohol zdenaturowany, alkohol rozcieńczony, kwas aleurytowy, kwas alginowy, poliestry alifatyczne, tlenek glinu, wodorotlenek glinu, stearynian glinu, amylopektyna, α- amyloza, kwas askorbinowy, palmitynian askorbylu, aspartam, woda bakteriostatyczna do wstrzykiwań, bentonit, magma bentonitowa, chlorek benzalkoniowy, chlorek benzetoniowy, kwas benzoesowy, alkohol benzylowy, benzoesan benzylu, bronopol, butylowany hydroksyanizol, butylowany hydroksytoluen, butyloparaben, butyloparaben sodu, alginian wapnia, askorbinian wapnia, węglan wapnia, cyklaminian wapnia, wodorofosforan wapnia bezwodny, wodorofosforan wapnia odwodniony, fosforan wapnia, propionian wapnia, krzemian wapnia, sorbinian wapnia, stearynian wapnia,
154 siarczan wapnia, siarczan wapnia półwodny, olej rzepakowy, karbomer, dwutlenek węgla, sól wapniowa karboksymetylocelulozy, sól sodowa karboksymetylocelulozy, β-karoten, karagen, olej rycynowy, uwodorniony olej rycynowy, kationowy wosk emulgujący, octan celulozy, ftalan octanu celulozy, etyloceluloza, celuloza mikrokrystaliczna, celuloza sproszkowana, krzemionkowana celuloza mikrokrystaliczna, sól sodowa karboksymetylocelulozy, alkohol cetostearylowy, cetrimid, alkohol cetylowy, chlorheksydyna, chlorobutanol, chlorokrezol, cholesterol, octan chlorheksydyny, glukonian chlorheksydyny, chlorowodorek chlorheksydyny, chlorodifluoroetan (HCFC), chlorodifluorometan, chlorofluorowęglowodory (CFC), chlorofenoksyetanol, chloroksylenol, syrop kukurydziany w postaci stałej, bezwodny kwas cytrynowy, kwas cytrynowy jednowodny, masło kakaowe, środki barwiące, olej kukurydziany, olej bawełniany, krezol, m-krezol, o-krezol, p-krezol, sól sodowa kroskarmelozy, krospowidon, kwas cyklaminowy, cyklodekstryny, dekstrany, dekstryna, dekstroza dekstroza bezwodna, diazolidynylomocznik, ftalan dibutylu, sebacynian dibutylu, dietanolamina, ftalan dietylu, difluoroetan (HFC), dimetylo-β-cyklodekstryna, związki typu cyklodekstryn, na przykład Captisol, eter dimetylowy, ftalan dimetylu, wersenian dipotasu, wersenian disodu, wodorofosforan disodowy, dokuzan wapnia, dokuzan potasu, dokuzan sodu, galusan dodecylu, bromek dodecylotrimetyloamoniowy, wersenian wapnia i disodu, kwas wersenowy, eglumina, alkohol etylowy, etyloceluloza, galusan etylu, laurynian etylu,
155 etylomaltol, oleinian etylu, etyloparaben, sól potasowa etyloparabenu, sól sodowa etyloparabenu, etylowanilina, fruktoza, fruktoza płynna, fruktoza mielona, fruktoza niepirogenna, fruktoza sproszkowana, kwas fumarowy, żelatyna, glukoza, glukoza płynna, mieszaniny glicerydów nasyconych roślinnych kwasów tłuszczowych, gliceryna, behenian glicerylu, monooleinian glicerylu, monostearynian glicerylu, samoemulgujący monostearynian glicerylu, palmitostearynian glicerylu, glicyna, glikole, glikofurol, guma guar, heptafluoropropan (HFC), bromek heksadecylotrimetyloamoniowy, stężony syrop fruktozowy, albumina surowicy ludzkiej, węglowodory (HC), rozcieńczony kwas solny, utwardzony olej roślinny, typu II, hydroksyetyloceluloza, 2- hydroksyetylo-β-cyklodekstryna, hydroksypropyloceluloza, hydroksypropyloceluloza o niskim stopniu podstawienia, 2-hydroksypropylo-βcyklodekstryna, hydroksypropylometyloceluloza, ftalan hydroksypropylometylocelulozy, imidomocznik, indygokarmin, wymieniacze jonowe, tlenki żelaza, alkohol izopropylowy, mirystynian izopropylu, palmitynian izopropylu, izotoniczna sól fizjologiczna, kaolin, kwas mlekowy, laktitol, laktoza, lanolina, alkohole lanolinowe, lanolina bezwodna, lecytyna, krzemian magnezu i glinu, węglan magnezu, zwykły węglan magnezu, węglan magnezu bezwodny, wodorotlenek węglan magnezu (zasadowy węglan magnezu), wodorotlenek magnezu, laurylosiarczan magnezu, tlenek magnezu, krzemian magnezu, stearynian magnezu, trikrzemian magnezu, trikrzemian magnezu bezwodny, kwas jabłkowy, słód, maltitol, roztwór maltitolu, maltodekstryna,
156 maltol, maltoza, mannitol, triglicerydy średniołańcuchowe, meglumina, mentol, metyloceluloza, metakrylan metylu, oleinian metylu, metyloparaben, sól potasowa metyloparabenu, sól sodowa metyloparabenu, celuloza mikrokrystaliczna i sól sodowa karboksymetylocelulozy, olej mineralny, lekki olej mineralny, olej mineralny i alkohole lanolinowe, olej, oliwa z oliwek, monoetanolamina, montmorylonit, galusan oktylu, kwas oleinowy, kwas palmitynowy, parafina, olej arachidowy, wazelina, wazelina i alkohole lanolinowe, glazura farmaceutyczna, fenol, fenol skroplony, fenoksyetanol, fenoksypropanol, alkohol fenyloetylowy, octan fenylortęciowy, boran fenylortęciowy, azotan fenylortęciowy, polakrylina, sól potasowa polakryliny, poloksamer, polidekstroza, poli(glikol etylenowy), poli(tlenek etylenu), poliakrylany, polimery blokowe polietylenu i polioksypropylenu, polimetakrylany, etery alkilowe polioksyetylenu, pochodne polioksyetylenowane oleju rycynowego, estry polioksyetylenowanego sorbitolu i kwasów tłuszczowych, stearyniany polioksyetylenu, poli(alkohol winylowy), poliwinylopirolidon, alginian potasu, benzoesan potasu, wodorowęglan potasu, wodorosiarczyn potasu, chlorek potasu, cytrynian potasu, cytrynian potasu bezwodny, wodorofosforan potasu, metasiarczyn potasu, diwodorofosforan potasu, propionian potasu, sorbinian potasu, powidon, propanol, kwas propionowy, węglan propylenu, glikol propylenowy, alginian glikolu propylenowego, galusan propylu, propyloparaben, sól potasowa propyloparabenu, sól sodowa propyloparabenu, siarczan protaminy, olej rzepakowy, roztwór Ringera, sacharyna, sacharynian
157 amonu, sacharynian wapnia, sacharynian sodu, olej z krokosza, saponit, białka surowicy, olej sezamowy, krzemionka koloidalna, koloidalny dwutlenek krzemu, alginian sodu, askorbinian sodu, benzoesan sodu, wodorowęglan sodu, wodorosiarczyn sodu, chlorek sodu, bezwodny cytrynian sodu, cytrynian sodu odwodniony, chlorek sodu, cyklaminian sodu, wersenian sodu, dodecylosiarczan sodu, laurylosiarczan sodu, metasiarczyn sodu, wodorofosforan sodu, diwodorofosforan sodu, fosforan sodu, bezwodny propionian sodu, propionian sodu, sorbinian sodu, glikolan sodowy skrobi, stearylofumaran sodu, siarczyn sodu, kwas sorbowy, estry sorbitanu (estry sorbitanu i kwasów tłuszczowych), sorbitol, roztwór sorbitolu 70%, olej sojowy, wosk olbrotowy, skrobia, skrobia kukurydziana, skrobia ziemniaczana, skrobia preżelowana, skrobia kukurydziana do sterylizowania, kwas stearynowy, oczyszczony kwas stearynowy, alkohol stearylowy, sacharoza, cukry, cukier do prasowania, cukier cukierniczy, sfery cukrowe, cukier inwertowany, Sugartab, żółcień pomarańczowa FCF, parafina syntetyczna, talk, kwas winowy, tartrazyna, tetrafluoroetan (HFC), olej kakaowy, tiomersal, dwutlenek tytanu, alfa-tokoferol, octan tokoferylu, kwaśny bursztynian alfa-tokoferylu, beta-tokoferol, delta-tokoferol, gamma-tokoferol, tragakant, triacetyna, cytrynian tributylu, trietanolamina, cytrynian trietylu, trimetylo-β-cyklodekstryna, bromek trimetylotetradecyloamoniowy, bufor tris, wersenian trisodu, wanilina, uwodorniony olej roślinny typu I, woda, woda miękka, woda twarda, woda niezawierająca
158 dwutlenku węgla, woda niepirogenna, woda do wstrzykiwań, woda jałowa do inhalacji, woda jałowa do wstrzykiwań, woda jałowa do irygacji, woski, anionowy wosk emulgujący, wosk karnauba, kationowy wosk emulgujący, wosk cetyloestrowy, wosk mikrokrystaliczny, niejonowy wosk emulgujący, wosk do czopków, wosk biały, wosk żółty, wazelina biała, tłuszcz z wełny, guma ksantanowa, ksylitol, zeina, propionian cynku, sole cynku, stearynian cynku lub dowolna zaróbkawedług Handbook of Pharmaceutical Excipients, wydanie trzecie, A. H. Kibbe (Pharmaceutical Press, Londyn, Wielka Brytania, 00), która to publikacja zostaje włączona w całości na mocy odniesienia. Remington s Pharmaceutical Sciences, wydanie szesnaste, E. W. Martin (Mack Publishing Co., Easton, Pa., 1980), ujawnia różne składniki wykorzystywane do otrzymywania preparatów kompozycji dopuszczalnych farmaceutycznie i znane metody ich otrzymywania. Z wyjątkiem okoliczności, w których typowy środek jest niezgodny z kompozycjami farmaceutycznymi, przewiduje się jego zastosowanie w kompozycjach farmaceutycznych. W kompozycjach można także stosować dodatkowe substancje czynne. [0300] Preparaty farmaceutyczne ujawniane w niniejszym dokumencie można określić jako krótko działające, o szybkim uwalnianiu, długo działające lub o uwalnianiu przzedłużonym, zgodnie z opisem poniżej. Preparaty farmaceutyczne można ponadto otrzymywać do uwalniania natychmiastowego, uwalniania kontrolowanego lub uwalniania wolnego. Niniejsze kompozycje mogą ponadto zawierać na przykład micele lub liposomy bądź pewną inną formę kapsułkowaną lub można je podawać w postaci
159 do uwalniania przedłużonego zapewniającej efekt wydłużonego przebywania i(lub) dostarczania. Ujawniane preparaty farmaceutyczne można podawać według dowolnego schematu, w tym na przykład codziennie (1 raz na dobę, 2 razy na dobę, 3 razy na dobę, 4 razy na dobę, razy na dobę, 6 razy na dobę), co dwa dni, co trzy dni, co cztery dni, co pięć dni, co sześć dni, raz na tydzień, raz na dwa tygodnie, raz na trzy tygodnie, raz na miesiąc lub raz na dwa miesiące. [0301] W niektórych przykładach wykonania powyższy składnik lub składniki mogą występować w kompozycji farmaceutycznej w dowolnym stężeniu, takim jak na przykład co najmniej A, przy czym A wynosi 0,0001% wag./obj., 0,001% wag./obj., 0,01% wag./obj., 0,1% wag./obj., 1% wag./obj., 2% wag./obj., % wag./obj., % wag./obj., % wag./obj., 30% wag./obj., 40% wag./obj., 0% wag./obj., 60% wag./obj., 70% wag./obj., 80% wag./obj. lub 90% wag./obj.. W niektórych przykładach wykonania powyższy składnik lub składniki mogą występować w kompozycji farmaceutycznej w dowolnym stężeniu, takim jak na przykład co najwyżej B, przy czym B wynosi 90% wag./obj., 80% wag./obj., 70% wag./obj., 60% wag./obj., 0% wag./obj., 40% wag./obj., 30% wag./obj., % wag./obj., % wag./obj., % wag./obj., 2% wag./obj., 1% wag./obj., 0,1% wag./obj., 0,001% wag./obj. lub 0,0001%. W innych przykładach wykonania powyższy składnik lub składniki mogą występować w kompozycji farmaceutycznej w dowolnym zakresie stężenia, takim jak na przykład od około A do około B. W niektórych przykładach wykonania A wynosi 0,0001%, a B wynosi 90%.
160 [0302] Preparaty kompozycji farmaceutycznych można otrzymywać tak, aby uzyskać ph zgodne fizjologicznie. W niektórych przykładach wykonania ph kompozycji farmaceutycznej może wynosić co najmniej, co najmniej,, co najmniej 6, co najmniej 6,, co najmniej 7, co najmniej 7,, co najmniej 8, co najmniej 8,, co najmniej 9, co najmniej 9,, co najmniej lub co najmniej,, aż do ph 11 włącznie w zależności od preparatu i drogi podania. W niektórych przykładach wykonania kompozycje farmaceutyczne mogą zawierać środki buforujące do uzyskania ph zgodnego fizjologicznie. Środki buforujące mogą zawierać dowolne związki zdolne do buforowania w pożądanym ph, takie jak na przykład bufory fosforanowe (np. PBS), trietanolamina, Tris, bicyna, TAPS, tricyna, HEPES, TES, MOPS, PIPES, kakodylan, MES i inne. W niektórych przykładach wykonania moc buforu wynosi co najmniej 0, mm, co najmniej 1 mm, co najmniej mm, co najmniej mm, co najmniej mm, co najmniej 30 mm, co najmniej 40 mm, co najmniej 0 mm, co najmniej 60 mm, co najmniej 70 mm, co najmniej 80 mm, co najmniej 90 mm, co najmniej 0 mm, co najmniej 1 mm, co najmniej 10 mm lub co najmniej 0 mm. W niektórych przykładach wykonania moc buforu wynosi nie więcej niż 300 mm (np. co najwyżej 0 mm, co najwyżej 0 mm, co najwyżej 90 mm, co najwyżej 80 mm, co najwyżej 70 mm, co najwyżej 60 mm, co najwyżej 0 mm, co najwyżej 40 mm, co najwyżej 30 mm, co najwyżej mm, co najwyżej mm, co najwyżej mm, co najwyżej 1 mm). [0303] Peptydy glukagonu działające koagonistycznie wobec GIP/GLP-1, koagonistycznie wobec glukagonu/gip i
161 triagonistycznie wobec glukagonu/gip/glp-1 można wykorzystywać w dowolnym wskazaniu w związku z każdą z ich aktywności opisanych wcześniej jako użyteczne. Na przykład aktywność glukagonu może zwiększać poziomy glukozy, służyć do buforowania insuliny lub zmniejszać motorykę jelit podczas badania radiologicznego. Aktywność GLP-1 może obniżyć poziomy glukozy, które to działanie jest użyteczne w leczeniu hiperglikemii, np. cukrzycy. Aktywność GLP-1 może ponadto indukować utratę masy lub zapobiegać wzrostowi masy, np. dzięki zmniejszeniu łaknienia. Aktywność GIP może ponadto obniżać poziomy glukozy, które to działanie jest użyteczne w leczeniu hiperglikemii, np. cukrzycy. [0304] Związki działające koagonistycznie wobec GIP/GLP-1 i triagonistycznie wobec glukagonu/gip/glp-1 są szczególnie korzystne do indukowania utraty masy lub zapobiegania wzrostowi masy, a także w leczeniu hiperglikemii, w tym cukrzycy. Dane in vivo ujawniane w niniejszym dokumencie pokazują, że połączenie działania agonistycznego wobec GIP z działaniem agonistycznym wobec GLP-1 powoduje większe oddziaływanie na zmniejszenie masy niż sam GLP-1. Aktywność ta jest szczególnie nieoczekiwana w świetle ujawnień ze stanu techniki, według których działanie antagonistyczne wobec GIP jest pożądane, aby zmniejszyć dobowe przyjmowanie pokarmu i masę ciała, a także zwiększyć insulinowrażliwość i wydatek energetyczny (Irwin i wsp., Diabetologia 0: (07); oraz Althage i wsp., J Biol Chem, publikacja elektroniczna z 17 kwietnia 08 r.). [030] Dane in vivo ujawniane w niniejszym dokumencie
162 pokazują ponadto, że połączenie działania agonistycznego wobec GIP z działaniem agonistycznym wobec GLP-1 zmniejsza poziomy glukozy. [0306] Oznacza to, że oczekuje się, iż peptydy glukagonu opisane w niniejszym dokumencie będą wykorzystywane do zmniejszenia, lub utrzymania masy ciała, lub do leczenia hiperglikemii, lub do obniżania poziomu glukozy we krwi, lub do normalizacji i(lub) stabilizacji poziomu glukozy we krwi. [0307] W niektórych przykładach wykonania dostarcza się sposób leczenia hiperglikemii lub sposób zmniejszania wzrostu masy lub indukowania utraty masy, który obejmuje podawanie ilości skutecznej roztworu wodnego zawierającego peptyd glukagonu według wynalazku. W kolejnych przykładach wykonania ujawnia się sposoby leczenia cukrzycy obejmujące podawanie skojarzone dawki konwencjonalnej lub dawki zmniejszonej insuliny i peptydu glukagonu według wynalazku. Dostarcza się ponadto sposoby leczenia cukrzycy za pomocą peptydu glukagonu według wynalazku, bez podawania skojarzonego insuliny. [0308] Oczekuje się, że takie sposoby leczenia hiperglikemii będą użyteczne w przypadku różnych postaci hiperglikemii, w tym cukrzycy, cukrzycy typu I, cukrzycy typu II lub cukrzycy ciążowej, insulinozależnej lub insulinoniezależnej, i ograniczenia powikłań cukrzycy, w tym nefropatii, retinopatii i chorób naczyniowych. [0309] Oczekuje się, że sposoby ograniczania łaknienia lub ułatwiania utraty masy ciała będą użyteczne w zmniejszaniu masy ciała, zapobieganiu wzrostowi masy
163 ciała lub leczeniu otyłości z różnych przyczyn, w tym otyłości polekowej, a także ograniczania powikłań związanych z otyłością, w tym chorób naczyniowych (choroby naczyń wieńcowych, udaru, choroby naczyń obwodowych, reperfuzji po niedokrwieniu itd.), nadciśnienia, początku cukrzycy typu II, hiperlipidemii i chorób mięśniowo-szkieletowych. [03] Zespół metaboliczny, znany także jako zespół metaboliczny X, zespół insulinooporności lub zespół Reavena, jest zaburzeniem, na które cierpi ponad 0 milionów Amerykanów. Zespół metaboliczny charakteryzuje się zazwyczaj połączeniem co najmniej trzech lub większej liczby z następujących czynników ryzyka: (1) otyłość brzuszna (nadmiar tkanki tłuszczowej w jamie brzusznej i w jej okolicy), (2) dyslipidemia aterogenna (zaburzenia dotyczące tłuszczu we krwi, w tym wysoki poziom triglicerydów, niski poziom cholesterolu HDL i wysoki poziom cholesterolu LDL, nasilające narastanie blaszek na ścianach tętnic), (3) podwyższone ciśnienie krwi, (4) insulinooporność lub nietolerancja glukozy, () stan prozakrzepowy (np. wysoki poziom fibrynogenu lub inhibitora aktywatora plazminogenu 1 we krwi), i (6) stan prozapalny (np. podwyższony poziom białka C- reaktywnego we krwi). Do innych czynników ryzyka mogą należeć starzenie, zaburzenia równowagi hormonalnej i predyspozycje genetyczne. [0311] Zespół metaboliczny jest związany ze zwiększonym ryzykiem choroby wieńcowej oraz innych zaburzeń związanych z narastaniem blaszki w naczyniach, takich jak udar i choroba naczyń obwodowych, określanych jako miażdżycopochodna choroba układu sercowo-naczyniowego
164 (ASCVD). U pacjentów z zespołem metabolicznym może dojść do progresji od stanu insulinooporności we wczesnych stadiach do pełnoobjawowej cukrzycy typu II z dodatkowym wzrostem ryzyka ASCVD. Bez ograniczania się do żadnej konkretnej teorii należy stwierdzić, że związek między insulinoopornością, zespołem metabolicznym i chorobą naczyń może obejmować jeden lub większą liczbę równoległych mechanizmów chorobotwórczych, w tym zaburzenie rozszerzania naczyń stymulowanego insuliną, zmniejszenie dostępności NO związane z insulinoopornością wskutek zwiększenia stresu oksydacyjnego oraz zaburzenia dotyczące hormonów pochodzących z adipocytów, takich jak adiponektyna (Lteif i Mather, Can. J. Cardiol. (suppl. B):66B-76B (04)). [0312] Według ustaleń National Cholesterol Education Program Adult Treatment Panel (ATP III) z 01 r. kryteria dla zespołu metabolicznego są spełnione, jeśli u tej samej osoby występują dowolne z trzech następujących cech: (a) otyłość brzuszna (obwód w pasie ponad 2 cm u mężczyzn i ponad 88 cm u kobiet); (b) triglicerydy w surowicy (10 mg/dl lub więcej); (c) cholesterol HDL (40 mg/dl lub mniej u mężczyzn i 0 mg/dl lub mniej u kobiet); (d) ciśnienie krwi (130/8 lub więcej); oraz (e) stężenie glukozy we krwi na czczo (1 mg/dl lub więcej). Według Światowej Organizacji Zdrowia (WHO) osoba mająca wysokie poziomy insuliny (podwyższony poziom glukozy we krwi na czczo lub wyłącznie podwyższony poziom glukozy po posiłku) i co najmniej dwie z następujących cech spełnia kryteria zespołu metabolicznego: (a) otyłość brzuszna (stosunek
165 obwodu w pasie do obwodu w biodrach przekraczający 0,9, współczynnik masy ciała co najmniej 30 kg/m 2 lub obwód w pasie ponad 37 cali); (b) profil lipidowy wykazujący poziom triglicerydów wynoszący co najmniej 10 mg/dl lub cholesterol HDL poniżej 3 mg/dl; (c) ciśnienie krwi 140/90 lub więcej lub aktualne leczenie podwyższonego ciśnienia krwi). (Mathur, Ruchi, Metabolic Syndrome, red. Shiel, Jr., William C., MedicineNet.com, 11 maja 09 r.). [0313] Na potrzeby niniejszego dokumentu, jeśli osoba spełnia warunki według jednego lub obydwu kryteriów określonych przez National Cholesterol Education Program Adult Treatment Panel z 01 r. lub WHO, uznaje się, że osoba taka cierpi na zespół metaboliczny. [0314] Bez ograniczania się do żadnej konkretnej teorii peptydy glukagonu opisywane w niniejszym dokumencie są użyteczne w leczeniu zespołu metabolicznego. W związku z tym w niniejszym dokumencie ujawnia się sposób zapobiegania lub leczenia zespołu metabolicznego bądź ograniczenia jednego, dwóch, trzech lub większej liczby jego czynników ryzyka u osobnika, obejmujący podawanie osobnikowi peptydu glukagonu opisanego w niniejszym dokumencie w ilości skutecznej do zapobiegania lub leczenia zespołu metabolicznego bądź jego czynnika ryzyka. [031] Niealkoholowa stłuszczeniowa choroba wątroby (ang. nonalcoholic fatty liver disease, NAFLD) dotyczy szerokiego spektrum chorób wątroby od prostego otłuszczenia wątroby (stłuszczenie) do niealkoholowego stłuszczeniowego zapalenia wątroby (ang. nonalcoholic steatohepatitis, NASH) i marskości (nieodwracalnego
166 zaawansowanego bliznowacenia wątroby). Cechą wspólną wszystkich stadiów NAFLD jest gromadzenie tłuszczu (naciek tłuszczowy) w komórkach wątroby (hepatocytach). Proste stłuszczenie wątroby stanowi nieprawidłowe gromadzenie pewnego rodzaju tłuszczu, triglicerydów, w komórkach wątroby bez stanu zapalnego lub bliznowacenia. W przypadku NASH gromadzeniu tłuszczu towarzyszy stan zapalny o różnym nasileniu (zapalenie wątroby) i bliznowacenie (zwłóknienie) wątroby. Komórki zapalne mogą niszczyć komórki wątroby (martwica wątrobowokomórkowa). W określeniach stłuszczeniowe zapalenie wątroby i stłuszczeniowa martwica stłuszczenie oznacza naciek tłuszczowy, zapalenie wątroby oznacza stan zapalny wątroby, a martwica oznacza zniszczone komórki wątroby. NASH może ostatecznie prowadzić do bliznowacenia wątroby (zwłóknienie), a następnie do nieodwracalnego, zaawansowanego bliznowacenia (marskości). Marskość powodowana przez NASH stanowi ostatnie i najcięższe stadium w obrębie spektrum NAFLD. (Mendler, Michel, Fatty Liver: Nonalcoholic Fatty Liver Disease (NAFLD) and Nonalcoholic Steatohepatitis (NASH), red. Schoenfield, Leslie J., MedicineNet.com, 29 sierpnia 0 r.). [0316] Alkoholowa choroba wątroby lub poalkoholowa choroba wątroby obejmuje trzy patologicznie odmienne choroby wątroby związane z nadmiernym spożywaniem alkoholu lub przez nie wywołane: otłuszczenie wątroby (stłuszczenie), przewlekłe lub ostre zapalenie wątroby i marskość. Alkoholowe zapalenie wątroby może obejmować zakres od łagodnego zapalenia wątroby z nieprawidłowymi
167 wynikami badań laboratoryjnych, które są jedynym przejawem choroby, do ciężkiej dysfunkcji choroby z powikłaniami, takimi jak żółtaczka (żółta barwa skóry wywołana przez zatrzymywanie bilirubiny), encefalopatia wątrobowa (dysfunkcja neurologiczna spowodowana niewydolnością wątroby), wodobrzusze (gromadzenie płynów w jamie brzusznej), krwawiące żylaki przełyku (żyły żylakowe w przełyku), nieprawidłowa krzepliwość krwi i śpiączka. W ujęciu histologicznym alkoholowe zapalenie wątroby ma charakterystyczny obraz ze zwyrodnieniem hepatocytów przebiegającym z rozdęciem, stanem zapalnym z neutrofilami i niekiedy ciałkami Mallory ego (nieprawidłowymi skupieniami komórkowych białek filamentów pośrednich). Marskość charakteryzuje się anatomicznie licznymi guzkami w wątrobie w połączeniu ze zwłóknieniem. (Worman, Howard J., Alcoholic Liver Disease, strona internetowa Columbia University Medical Center). [0317] Bez ograniczania się do żadnej konkretnej teorii peptydy glukagonu opisywane w niniejszym dokumencie są użyteczne w leczeniu alkoholowej choroby wątroby, NAFLD lub dowolnego ich stadium, w tym na przykład stłuszczenia, stłuszczeniowego zapalenia wątroby, zapalenia wątroby, stanu zapalnego wątroby, NASH, marskości lub ich powikłań. W związku z tym w niniejszym dokumencie ujawnia się sposób zapobiegania lub leczenia alkoholowej choroby wątroby, NAFLD lub dowolnego ich stadium u osobnika, obejmujący podawanie osobnikowi peptydu glukagonu opisanego w niniejszym dokumencie w ilości skutecznej do zapobiegania lub leczenia alkoholowej choroby wątroby, NAFLD lub ich
168 stadium. Takie sposoby leczenia obejmują zmniejszenie jednego, dwóch, trzech lub większej liczby z następujących: zawartość tłuszczu w wątrobie, wystąpienie lub progresja marskości, wystąpienie raka wątrobowokomórkowego, objawy stanu zapalnego, np. nieprawidłowe poziomy enzymów wątrobowych (np. aminotransferazy asparaginianowej AST i(lub) aminotransferazy alaninowej ALT lub LDH), podwyższony poziom ferrytyny w surowicy, podwyższony poziom bilirubiny w surowicy i(lub) objawy zwłóknienia, np. podwyższone poziomy TGF-beta. W korzystnych przykładach wykonania peptydy glukagonu stosuje się w leczeniu pacjentów, u których nastąpiła progresja wykraczająca poza proste otłuszczenie wątroby (stłuszczenie) i występują objawy stanu zapalnego lub zapalenia wątroby. Takie sposoby mogą na przykład doprowadzić do zmniejszenia poziomów AST i(lub) ALT. [0318] Peptydy glukagonu według wynalazku można podawać w monoterapii lub w skojarzeniu z innymi środkami przeciwcukrzycowymi lub przeciwko otyłości. Do środków przeciwcukrzycowych znanych w stanie techniki lub w fazie badań należą insulina, sulfonylomoczniki, takie jak tolbutamid (Orinase), acetoheksamid (Dymelor), tolazamid (Tolinase), chlorpropamid (Diabinese), glipizyd (Glucotrol), gliburyd (Diabeta, Micronase, Glynase), glimepiryd (Amaryl) lub gliklazyd (Diamicron); meglitynidy, takie jak repaglinid (Prandin) lub nateglinid (Starlix); biguanidy, takie jak metformina (Glucophage) lub fenformina; tiazolidynodiony, takie jak rozyglitazon (Avandia), pioglitazon (Actos) lub troglitazon (Rezulin) lub inne
169 inhibitory PPARγ; inhibitory alfa-glukozydazy powodujące hamowanie trawienia węglowodanów, takie jak miglitol (Glyset), akarboza (Precose/Glucobay); eksenatyd (Byetta) lub pramlintyd; inhibitory dipeptydylopeptydazy 4 (DPP-4), takie jak wildagliptyna lub sitagliptyna; inhibitory SGLT (transportera glukozy zależnego od sodu 1); aktywatory glukokinazy (GKA); antagoniści receptorów glukagonu (GRA); lub inhibitory FBPazy (1,6-bisfosfatazy fruktozy). [0319] Do środków przeciwko otyłości znanych w stanie techniki lub w fazie badań należą leptyna i czynnik wzrostu fibroblastów 21 (FGF-21), środki hamujące łaknienie, na przykład stymulanty typu fenetyloaminy, fentermina (ewentualnie z fenfluraminą lub deksfenfluraminą), dietylopropion (Tenuate ), fendimetrazyna (Prelu-2, Bontril ), benzfetamina (Didrex ), sybutramina (Meridia, Reductil ); rimonabant (Acomplia ), inne środki działające antagonistycznie na receptory kanabinoidowe; oksyntomodulina; chlorowodorek fluoksetyny (Prozac); Qnexa (topiramat i fentermina), Excalia (bupropion i zonisamid) lub Contrave (bupropion i naltrekson); lub inhibitory lipazy podobne do ksenikalu (Orlistat) lub Cetilistat (znany także jako ATL-962) lub GT [03] Peptydy glukagonu ujawniane w niniejszym dokumencie, które zachowują działania glukagonu polegające na zwiększeniu poziomu glukozy u pacjentów z hipoglikemią, można stosować w leczeniu hipoglikemii, np. do zapobiegania lub leczenia ostrej, okresowej lub nocnej hipoglikemii. Takie peptydy glukagonu można ponadto podawać w skojarzeniu z insuliną w celu
170 buforowania działań insuliny i aby ułatwić utrzymanie stabilnych poziomów glukozy we krwi u diabetyków. W takich przykładach wykonania dostarcza się udoskonalony sposób regulacji poziomów glukozy we krwi u pacjentów zależnych od insuliny. Peptydy glukagonu według niniejszego ujawnienia można podawać w skojarzeniu z insuliną w postaci pojedynczej kompozycji, podawać jednocześnie w postaci odrębnych roztworów lub ewentualnie insulinę i peptyd glukagonu można podawać w różnym czasie względem siebie. W niektórych przykładach wykonania sposób obejmuje etapy podawania insuliny w ilości skutecznej terapeutycznie do kontroli cukrzycy i podawania nowego zmodyfikowanego peptydu glukagonu według niniejszego ujawnienia w ilości skutecznej terapeutycznie do zapobiegania hipoglikemii, przy czym wspomniane etapy podawania przeprowadza się w odstępie nie więcej niż dwunastu godzin. Dokładny stosunek zmodyfikowanego peptydu glukagonu względem podawanej insuliny będzie zależał po części od określenia poziomów glukozy u pacjenta i innych parametrów klinicznych. Normalizacja poziomu we krwi oznacza, że poziom glukozy we krwi powraca do wartości prawidłowych (np. obniżenie poziomu glukozy we krwi, jeśli jest on wyższy od prawidłowego, lub podwyższenie poziomu glukozy we krwi, jeśli jest on niższy od prawidłowego). Stabilizacja poziomu glukozy we krwi oznacza zmniejszenie maksymalnej zmienności poziomu glukozy we krwi w pewnym okresie, np. 8 godzin, 16 godzin, 24 godzin, 2 dni, 3 dni, 4 dni, dni, 6 dni lub 1 tygodnia. Na przykład podawanie peptydu glukagonu powoduje utrzymanie poziomu glukozy we krwi w okresie
171 bliższym normalnemu zakresowi wartości glukozy niż gdyby nie podano peptydu glukagonu. [0321] Peptydy glukagonu ujawniane w niniejszym dokumencie, które zachowują pożądaną aktywność, można wykorzystać do indukowania przemijającego porażenia jelit do zastosowań radiologicznych bądź do leczenia innych chorób metabolicznych wynikających z niskich poziomów glukagonu we krwi. W takich przykładach wykonania dostarcza się sposób indukowania przemijającego porażenia przewodu pokarmowego. Sposób ten obejmuje etap podawania pacjentowi jednego lub większej liczby peptydów glukagonu ujawnianych w niniejszym dokumencie. [0322] W innym aspekcie wynalazek dostarcza sposób zmniejszania wzrostu masy lub indukowania utraty masy lub leczenia hiperglikemii obejmujący skojarzone podawanie pacjentowi tego wymagającemu cząsteczki agonisty receptora GIP i cząsteczki agonisty receptora GLP-1 w ilościach skutecznych do zmniejszenia wzrostu masy lub indukowania utraty masy lub zmniejszenia łaknienia. Te dwie cząsteczki mogą znajdować się razem w tej samej kompozycji. Ewentualnie w takich sposobach można podawać cząsteczkę powodującą aktywację receptorów zarówno GIP, jak i GLP-1. Kombinacja różnych właściwości agonistów receptorów, tzn. aktywacji, zapewnia nieoczekiwane działanie addytywne lub synergiczne bądź inną nieoczekiwaną korzyść lub korzyści kliniczne. Według takich sposobów przewiduje się podawanie z konwencjonalną dawką insuliny, zmniejszoną dawką insuliny lub bez insuliny. Do przykładowych cząsteczek agonistów receptorów GIP
172 należy GIP lub analogi GIP, np. które zachowują co najmniej 0%, 60%, 70% lub 80% identyczności sekwencji na podstawie kontroli wzrokowej po dopasowaniu w celu maksymalizacji reszt pasujących. Do przykładowych cząsteczek agonistów receptorów GLP-1 należy GLP-1, analogi GLP-1, np. które zachowują co najmniej 0%, 60%, 70% lub 80% identyczności sekwencji na podstawie kontroli wzrokowej po dopasowaniu w celu maksymalizacji reszt pasujących, analogi eksendyny 4, np. które zachowują co najmniej 0%, 60%, 70% lub 80% identyczności sekwencji na podstawie kontroli wzrokowej po dopasowaniu w celu maksymalizacji reszt pasujących bądź ich pochodne. Do przykładowych cząsteczek wykazujących obie aktywności należą peptydy glukagonu według wynalazku, analogi GLP-1 powodujące aktywację zarówno receptorów GLP-1, jak i GIP, fuzje GIP i GLP-1 lub fuzje analogów GIP i analogów GLP-1 bądź ich zmodyfikowane chemicznie pochodne. [0323] Sposoby leczenia ujawniane w niniejszym dokumencie, które obejmują, ale nieograniczająco, leczenie hipoglikemii, mogą obejmować etapy podawania ujawnianych niniejszym peptydów glukagonu pacjentowi stosując dowolną standardową drogę podawania, w tym pozajelitowo, na przykład dożylnie, dootrzewnowo, podskórnie lub domięśniowo, dokanałowo, przezskórnie, doodbytniczo, doustnie, donosowo lub wziewnie. [0324] Według niektórych przykładów wykonania z takich sposobów wyklucza się, w zależności od konkretnej choroby, wszelkie analogi glukagonu lub analogi GLP-1 znane w stanie techniki, wobec których ujawniono, że są użyteczne w leczeniu tej określonej choroby. W innym
173 przykładzie wykonania mogą także być wykluczone peptydy opisane w patencie USA nr 6,864,069 jako działające zarówno jako agoniści GLP-1, jak i antagoniści glukagonu do leczenia cukrzycy. W innym przykładzie wykonania ujawnienie może wykluczać zastosowanie antagonistów glukagonu w leczeniu cukrzycy, takich jak antagoniści opisani w Unson i wsp., J. Biol. Chem., 264: (1989), Ahn i wsp., J. Med. Chem., 44: (01), oraz Sapse i wsp., Mol. Med., 8(): (02). W niektórych przykładach wykonania może zostać wykluczona oksyntomodulina lub peptyd glukagonu zawierający 8 C-końcowych aminokwasów oksyntomoduliny (NR. ID. SEKW.: 97). [032] Oksyntomodulina, występujący naturalnie hormon trawienny stwierdzany w jelicie cienkim, powoduje utratę masy (patrz Diabetes 0; 4: ). Oksyntomodulina jest peptydem o długości 37 aminokwasów zawierającym sekwencję glukagonu o długości 29 aminokwasów (tzn. NR. ID. SEKW.: 1), a następnie przedłużenie na końcu karboksylowym o długości 8 aminokwasów według NR. ID. SEKW.: 97 (KRNRNNIA). Mimo że niniejszy wynalazek przewiduje, że peptydy glukagonu opisywane w niniejszym dokumencie mogą ewentualnie być połączone z tym przedłużeniem na końcu karboksylowym o długości 8 aminokwasów (NR. ID. SEKW.: 97), wynalazek w niektórych przykładach wykonania ponadto konkretnie przewiduje peptydy glukagonu niezawierające przedłużenia na końcu karboksylowym o długości 8 kolejnych aminokwasów według NR. ID. SEKW.: 97. [0326] Dowolny z poniższych peptydów zostaje wykluczony ze związków według wynalazku, jednak dalsze jego
174 modyfikacje wykazujące pożądane działanie koagonistyczne lub triagonistyczne, kompozycje farmaceutyczne, zestawy i sposoby leczenia z użyciem takich związków mogą zostać włączone do wynalazku. Peptyd według NR. ID. SEKW.: 1 z substytucją [Arg12] i z amidem C-końcowym. Peptyd według NR. ID. SEKW.: 1 z substytucjami [Arg12,Lys] i z amidem C-końcowym. Peptyd według NR. ID. SEKW.: 1 z substytucjami [Arg12,Lys24] i z amidem C-końcowym. Peptyd według NR. ID. SEKW.: 1 z substytucjami [Arg12,Lys29] i z amidem C-końcowym. Peptyd według NR. ID. SEKW.: 1 z substytucją [Glu9]. Peptyd według NR. ID. SEKW.: 1 niezawierający His1, z substytucjami [Glu9, Glu16, Lys29] i amidem C-końcowym. Peptyd według NR. ID. SEKW.: 1 z substytucjami [Glu9, Glu16, Lys29] i z amidem C-końcowym. Peptyd według NR. ID. SEKW.: 1 z substytucjami [Lys13, Glu17] połączonymi poprzez mostek laktamowy i z amidem C-końcowym. Peptyd według NR. ID. SEKW.: 1 z substytucjami [Lys17, Glu21] połączonymi poprzez mostek laktamowy i z amidem C-końcowym; peptyd według NR. ID. SEKW.: 1 niezawierający His1, z substytucjami [Glu, Lys24] połączonymi poprzez mostek laktamowy; peptydy ujawniane w PCT/US08/0387, dokonanym 13 lutego 08 r., PCT/US06/017494, dokonanym maja 06 r.; PCT/US07/01841, dokonanym 17 sierpnia 07 r.; PCT/GB0/0007, dokonanym 2 lutego 0 r.; PCT/GB00/089, dokonanym 29 marca 00 r.; PCT/US06/000, dokonanym lutego 06 r. 30
175 PRZYKŁADY [0327] Związki według wynalazku można otrzymywać standardowymi metodami syntezy, metodami rekombinacji DNA lub dowolnymi innymi metodami otrzymywania peptydów i białek fuzyjnych. Mimo że niektóre aminokwasy nienaturalne nie mogą ulegać ekspresji standardowymi metodami rekombinacji DNA, metody ich otrzymywania są znane w stanie techniki. Związki według wynalazku zawierające części niepeptydowe można syntetyzować za pomocą standardowych reakcji chemii organicznej, a także, w stosownych przypadkach, standardowych reakcji chemii peptydów. Dodatkowe dane dotyczące glukagonu i aktywności GLP-1 peptydów glukagonu ujawniono ponadto w PCT/US08/0387, dokonanym 13 lutego 08 r. PRZYKŁAD 1 Ogólny protokół syntezy: [0328] Analogi glukagonu zsyntetyzowano wykorzystując pojedyncze sprzęganie Fast Boc z aktywacją HBTU rozpoczynając od 0,2 mmol żywicy Boc Thr(OBzl)Pam w zmodyfikowanym syntezatorze peptydów Applied Biosystem 430 A. Aminokwasy Boc i HBTU otrzymano od Midwest Biotech (Fishers, IN). Zastosowano następujące grupy zabezpieczające łańcuch boczny: Arg(Tos), Asn(Xan), Asp(OcHex), Cys(pMeBzl), His(Bom), Lys(2C1-Z), Ser(OBzl), Thr(OBzl), Tyr(2Br-Z), i Trp(CHO). Grupą zabezpieczającą łańcuch boczny na N-końcowej His był Boc. [0329] Każdą ukończoną peptydylożywicę poddawano działaniu roztworu % piperydyny w dimetyloformamidzie w celu usunięcia grupy formylowej z tryptofanu. Przeprowadzono trawienia ciekłym fluorowodorem w
176 obecności p-krezolu i siarczku dimetylu. Trawienie prowadzono przez 1 godzinę na łaźni lodowej stosując aparat do HF (Penninsula Labs). Po odparowaniu HF pozostałość zawieszano w eterze dietylowym i materiały stałe odsączano. Każdy peptyd ekstrahowano do ml wodnego roztworu kwasu octowego i rozcieńczoną próbkę analizowano metodą HPLC [Beckman System Gold, 0,46x cm Zorbax C8, 1 ml/min, 4C, 214 nm, bufor A = 0,1% TFA, B = 0,1% TFA/90% acetonitrylu, gradient % do 80% B w ciągu min]. [0330] Oczyszczanie prowadzono metodą FPLC stosując kolumnę Kromasil C18 2,2 x 2 cm z monitorowaniem w paśmie UV przy 214 nm i zbieraniem frakcji w odstępie minut. Frakcje jednorodne połączono i poddano liofilizacji uzyskując produkt o czystości >9%. Prawidłową masę cząsteczkową i czystość potwierdzano stosując analizę widma masowego MALDI. PRZYKŁAD 2 Ogólny protokół pegylowania: (Cys-maleimid) [0331] Zazwyczaj analog Cys glukagonu rozpuszcza się w roztworze soli fizjologicznej buforowanej fosforanem (- mg/ml) i dodaje się 0,01 M kwas etylenodiaminotetraoctowy (-1% objętości całkowitej). Dodaje się nadmiar (2-krotny) odczynnika maleimidometoksy-peg (Nektar) i mieszaninę reakcyjną miesza się w temperaturze pokojowej monitorując postęp reakcji metodą HPLC. Po 8-24 godz. mieszaninę reakcyjną zakwasza się i wprowadza na preparatywną kolumnę w układzie faz odwróconych w celu oczyszczania stosując gradient 0,1% TFA/acetonitrylu. Odpowiednie frakcje połączono i poddano liofilizacji uzyskując pożądane
177 analogi pegylowane. PRZYKŁAD 3 Synteza glukagonu Cys 17 (1-29) i podobnych analogów MonoCys [0332] 0,2 mmol żywicy Boc Thr(OBzl) Pam (SynChem Inc) wprowadzono do naczynia reakcyjnego o objętości 60 ml, wprowadzono następującą sekwencję i przeprowadzono syntezę stosując zmodyfikowany syntezator peptydów Applied Biosystems 430A z wykorzystaniem sprzęgań pojedynczych FastBoc z aktywacją HBTU. HSQGTFTSDYSKYLDSCRAQDFVQWLMNT Zastosowano następujące grupy zabezpieczające łańcuch boczny: Arg(Tos), Asp(OcHex), Asn(Xan), Cys(pMeBzl), Glu(OcHex), His(Boc), Lys(2C1-Z), Ser(Bzl), Thr(Bzl), Trp(CHO) i Tyr(Br-Z). Ukończoną peptydylożywicę poddawano działaniu roztworu % piperydyny w dimetyloformamidzie w celu usunięcia zabezpieczającej grupy formylowej z Trp, a następnie przenoszono do naczynia do reakcji z HF i suszono pod próżnią. Dodawano 1,0 ml p-krezolu i 0, ml siarczku dimetylu oraz wrzucano mieszadełko magnetyczne. Naczynie podłączano do aparatu do HF (Pennisula Labs), chłodzono na łaźni z suchym lodem/metanolem, usuwano powietrze i skraplano ok. ml ciekłego fluorowodoru. Reakcję mieszano na łaźni lodowej przez 1 godz., a następnie HF usuwano pod próżnią. Pozostałość zawieszano w eterze etylowym, ciała stałe odsączano, przemywano eterem i peptyd ekstrahowano do 0 ml wodnego roztworu kwasu octowego. Stosowano analityczną metodę HPLC [0,46 x cm Zorbax C8, 1 ml/min, 4C, 214 nm, bufor A 0,1% TFA, bufor B 0,1% TFA/90% ACN, gradient = % B do 80% B w
178 ciągu min] wykorzystując małą próbkę ekstraktu po trawieniu. Pozostały ekstrakt nałożono na preparatywną kolumnę w układzie faz odwróconych Kromasil C18 2,2 x 2 cm i stosowano gradient acetonitrylu wykorzystując system do FPLC Pharmacia. Zbierano frakcje w odstępach min z monitorowaniem w paśmie UV przy 214 nm (2,0 A). A = 0,1% TFA, B = 0,1% TFA/0% acetonitryl. Gradient = 30% B do 0% B w ciągu 40 min. [0333] Frakcje zawierające produkt o największej czystości (48-2) połączono, zamrożono i poddano liofilizacji uzyskując 30,1 mg. Analiza produktu metodą HPLC wykazała czystość wynoszącą >90%, a analiza widma masowego metodą MALDI wykazała pożądaną masę wynoszącą 3429,7. Glukagon Cys 21, glukagon Cys 24 i glukagon Cys 29 otrzymano w podobny sposób. PRZYKŁAD 4 Synteza glukagonu-cex i innych analogów z przedłużeniem C-końcowym [0334] 28 mg (0,2 mmol) żywicy metoksybenzhydrylaminowej (Midwest Biotech) wprowadzono do naczynia reakcyjnego o objętości 60 ml, wprowadzono następującą sekwencję i przeprowadzono syntezę stosując zmodyfikowany syntezator peptydów Applied Biosystems 430A z wykorzystaniem sprzęgań pojedynczych FastBoc z aktywacją HBTU. HSQGTFTSDYSKYLDSRRAQDFVQWLMNTGPSSGAPPPS [033] Zastosowano następujące grupy zabezpieczające łańcuch boczny: Arg(Tos), Asp(OcHex), Asn(Xan), Cys(pMeBzl), Glu(OcHex), His(Boc), Lys(2C1-Z), Ser(Bzl), Thr(Bzl), Trp(CHO) i Tyr(Br-Z). Ukończoną peptydylożywicę poddawano działaniu roztworu %
179 piperydyny w dimetyloformamidzie w celu usunięcia zabezpieczającej grupy formylowej z Trp, a następnie przenoszono do naczynia do reakcji z HF i suszono pod próżnią. Dodawano 1,0 ml p-krezolu i 0, ml siarczku dimetylu oraz wrzucano mieszadełko magnetyczne. Naczynie podłączano do aparatu do HF (Pennisula Labs), chłodzono na łaźni z suchym lodem/metanolem, usuwano powietrze i skraplano ok. ml ciekłego fluorowodoru. Reakcję mieszano na łaźni lodowej przez 1 godz., a następnie HF usuwano pod próżnią. Pozostałość zawieszano w eterze etylowym, ciała stałe odsączano, przemywano eterem i peptyd ekstrahowano do 0 ml wodnego roztworu kwasu octowego. Stosowano analityczną metodę HPLC [0,46 x cm Zorbax C8, 1 ml/min, 4C, 214 nm, bufor A 0,1% TFA, bufor B 0,1% TFA/90% ACN, gradient = % B do 80% B w ciągu min] wykorzystując próbkę ekstraktu po trawieniu. Ekstrakt nałożono na preparatywną kolumnę w układzie faz odwróconych Kromasil C18 2,2 x 2 cm i do elucji stosowano gradient acetonitrylu wykorzystując system do FPLC Pharmacia. Zbierano frakcje w odstępach min z monitorowaniem w paśmie UV przy 214 nm (2,0 A). A = 0,1% TFA, B = 0,1% TFA/0% acetonitryl. Gradient = 30% B do 0% B w ciągu 40 min. Frakcje 8-6 połączono, zamrożono i poddano liofilizacji otrzymując 198,1 mg. [0336] Analiza produktu metodą HPLC wykazała czystość przekraczającą 9%. Analiza widma masowego metodą MALDI wykazała obecność pożądanej masy teoretycznej wynoszącej 4316,7, przy czym produkt stanowił amid C- końcowy. Oksyntomodulinę i oksyntomodulinę-krnr otrzymano w podobny sposób w postaci C-końcowych kwasów
180 karboksylowych stosując odpowiednio nałożoną żywicę PAM. PRZYKŁAD Glukagon Cyst Mal-PEG-K [0337] 1,1 mg glukagonu Cys 17 (1-29) i 27,3 mg metoksypoli(etyleno glikolo)maleimidu o śr. m.cz. 000 (mpeg-mal-000, Nektar Therapeutics) rozpuszczono w 3, ml roztworu soli fizjolgicznej buforowanej fosforanem (PBS) i dodano 0, ml 0,01 M kwasu etylenodiaminotetraoctowego (EDTA). Reakcję mieszano w temperaturze pokojowej, a postęp reakcji monitorowano za pomocą analizy metodą HPLC [0,46 x cm Zorbax C8, 1 ml/min, 4C, 214 nm (0, A), A = 0,1% TFA, B = 0,1% TFA/90% ACN, gradient = % B do 80% B w ciągu min]. Po godzinach mieszaninę reakcyjną nałożono na preparatywną kolumnę w układzie faz odwróconych Kromasil C18 2,2 x 2 cm. W urządzeniu do FPLC Pharmacia zastosowano gradient acetonitrylu z monitorowaniem w paśmie UV przy długości fali 214 nm i zbierano frakcje w odstępach min. A = 0,1% TFA, B = 0,1% TFA/0% acetonitryl, gradient = 30% B do 0% B w ciągu 40 min. Frakcje odpowiadające produktowi połączono, zamrożono i poddano liofilizacji otrzymując 2,9 mg. [0338] Produkt ten poddano analizie metodą HPLC [0,46 x cm Zorbax C8, 1 ml/min, 4C, 214 nm (0,A), A = 0,1% TFA, B = 0,1% TFA/90% ACN, gradient = % B do 80% B w ciągu min], która wykazała czystość ok. 90%. Analiza widma masowego metodą MALDI (jonizacja z desorpcją laserową wspomaganą matrycą) wykazała szeroki zakres mas (typowy dla pochodnych PEG) wynoszący 8700 do 900.
181 Potwierdza to zwiększenie masy wyjściowego peptydu glukagonu (3429) o około 000 jednostek masy atomowej (amu). PRZYKŁAD 6 Glukagon Cys 21 Mal-PEG-K [0339] 21,6 mg glukagonu Cys 21 (1-29) i 24 mg mpeg-mal- 000 (Nektar Therapeutics) rozpuszczono w 3, ml roztworu soli fizjologicznej buforowanej fosforanem (PBS) i dodano 0, ml 0,01 M kwasu etylenodiaminotetraoctowego (EDTA). Mieszaninę reakcyjną mieszano w temperaturze pokojowej. Po 2 godz. dodano jeszcze 12,7 mg mpeg-mal-000. Po 8 godz. mieszaninę reakcyjną nałożono na preparatywną kolumnę w układzie faz odwróconych Vydac C18 2,2 x 2 cm i stosowano gradient acetonitrylu wykorzystując system do FPLC Pharmacia przy natężeniu przepływu 4 ml/min zbierając frakcje w odstępach min. A = 0,1% TFA, B = 0,1% TFA/0% ACN. Gradient = % do 80% B w ciągu 40 min. [0340] Frakcje odpowiadające wyglądem produktowi połączono, zamrożono i poddano liofilizacji otrzymując 34 mg. Analiza produktu analityczną metodą HPLC [0,46 x cm Zorbax C8, 1 ml/min, 4C, 214 nm (0,A), A = 0,1% TFA, B = 0,1% TFA/90% ACN, gradient = % B do 80% B w ciągu min] wykazała produkt jednorodny odmienny od wyjściowego peptydu glukagonu. Analiza widma masowego metodą MALDI (jonizacja z desorpcją laserową wspomaganą matrycą) wykazała szeroki zakres mas (typowy dla analogów PEG) wynoszący 8700 do Potwierdza to zwiększenie masy wyjściowego peptydu glukagonu (3470) o około 000 amu.
182 PRZYKŁAD 7 Glukagon Cys 24 Mal-PEG-K [0341],1 mg glukagonu C 24 (1-29) i 39, mg mpeg-mal- 000 (Nektar Therapeutics) rozpuszczono w 3, ml PBS mieszając i dodano 0, ml 0,01 M EDTA. Mieszaninę reakcyjną mieszano w temperaturze pokojowej przez 7 godz., a następnie dodano jeszcze 40 mg mpeg-mal-000. Po około 1 godz. mieszaninę reakcyjną nałożono na preparatywną kolumnę w układzie faz odwróconych Vydac C18 2,2 x 2 cm i stosowano gradient acetonitrylu wykorzystując system do FPLC Pharmacia. Zbierano frakcje w odstępach min z monitorowaniem w paśmie UV przy 214 nm (2,0A). Bufor A = 0,1% TFA, bufor B = 0,1% TFA/0% ACN, gradient = 30% B do 0% B w ciągu 40 min. Frakcje odpowiadające produktowi połączono, zamrożono i poddano liofilizacji otrzymując 4,8 mg. Analiza widma masowego metodą MALDI wykazała typowy szeroki sygnał PEG z maksimum przy 917,2 oznaczający w przybliżeniu 000 amu więcej niż dla glukagonu C 24 (347,8). PRZYKŁAD 8 Glukagon Cys 24 Mal-PEG-K [0342] 2,7 mg glukagonu C 24 (1-29) i 40,7 mg mpeg-mal- K (Nektar Therapeutics) rozpuszczono w 3, ml PBS mieszając w temperaturze pokojowej i dodano 0, ml 0,01 M EDTA. Po 6 godz. stosunek materiału wyjściowego do produktu wynosił ok. 60:40 na podstawie analizy metodą HPLC. Dodano jeszcze 2,1 mg mpeg-mal-k i mieszaninę reakcyjną mieszano jeszcze przez 16 godz. Stosunek produktu nie zwiększył się znacząco, dlatego mieszaninę reakcyjną nałożono na preparatywną kolumnę w układzie
183 faz odwróconych Kromasil C18 2,2 x 2 cm i oczyszczano wykorzystując system do FPLC Pharmacia stosując gradient 30% B do 0% B w ciągu 40 min, bufor A = 0,1% TFA, bufor B = 0,1% TFA/0% ACN, przepływ = 4 ml/min i zbierano frakcje w odstępach min z monitorowaniem w paśmie UV przy 214 nm (2,0 A). Frakcje zawierające produkt jednorodny połączono, zamrożono i poddano liofilizacji otrzymując 2,7 mg. Czystość wyznaczona analityczną metodą HPLC wynosiła ~90%. Analiza widma masowego metodą MALDI wykazała szeroki pik od do oznaczający w przybliżeniu 000 amu więcej niż dla wyjściowego glukagonu C 24 (347,8). PRZYKŁAD 9 Glukagon Cys 29 Mal-PEG-K [0343],0 mg glukagonu Cys 29 (1-29) i 24,7 mg mpeg- Mal-000 (Nektar Therapeutics) rozpuszczono w 3, ml PBS mieszając w temperaturze pokojowej i dodano 0, ml 0,01 M EDTA. Po 4 godz. dodano jeszcze 1,6 mg mpeg- Mal-000, aby doprowadzić reakcję do końca. Po 8 godz. mieszaninę reakcyjną nałożono na preparatywną kolumnę w układzie faz odwróconych Vydac C18 2,2 x 2 cm i stosowano gradient acetonitrylu wykorzystując system do FPLC Pharmacia. Zbierano frakcje w odstępach min z monitorowaniem w paśmie UV przy 214 nm (2,0 A). A = 0,1% TFA, B = 0,1% TFA/0% ACN. Frakcje 7-97 połączono, zamrożono i poddano liofilizacji otrzymując 40,0 mg produktu różniącego się od wyodrębnionego materiału wyjściowego w analizie metodą HPLC (frakcje 8-63). Analiza produktu analityczną metodą HPLC [0,46 x cm Zorbax C8, 1 ml/min, 4C, 214 nm (0,A), A =
184 ,1% TFA, B = 0,1% TFA/90% ACN, gradient = % B do 80% B w ciągu min] wykazała czystość przekraczającą 9%. Analiza widma masowego metodą MALDI wykazała występowanie składnika PEG w zakresie masy 8000 do 000 (maksimum przy 902,3) oznaczającym 40 amu więcej niż w przypadku materiału wyjściowego (3484,8). PRZYKŁAD Glukagon Cys 24 (2-butyrolakton) [0344] Do 24,7 mg glukagonu Cys 24 (1-29) dodano 4 ml 0,0 M wodorowęglanu amonu/0% acetonitrylu i, μl roztworu -laktonu kwasu 2-bromo-4-hydroksymasłowego (0 μl w 900 μl acetonitrylu). Po 3 godz. mieszania w temperaturze pokojowej do mieszaniny reakcyjnej dodano jeszcze μl roztworu laktonu i mieszano ją jeszcze przez 1 godz. Mieszaninę reakcyjną rozcieńczono do ml za pomocą % roztworu wodnego kwasu octowego i nałożono na preparatywną kolumnę w układzie faz odwróconych Kromasil C18 2,2 x 2 cm. W urządzeniu do FPLC firmy Pharmacia zastosowano gradient acetonitrylu (% B do 80% B w ciągu 40 min) zbierając frakcje w odstępach min z monitorowaniem w paśmie UV przy 214 nm (2,0 A). Przepływ = 4 ml/min, A = 0,1% TFA, B = 0,1% TFA/0% ACN. Frakcje połączono, zamrożono i poddano liofilizacji otrzymując 7, mg. Analiza metodą HPLC wykazała czystość wynoszącą 9%, a analiza widma masowego metodą MALDI wykazała masę 340,7, czyli 84 jednostek masy więcej niż w przypadku materiału wyjściowego. Wynik był zgody z addycją pojedynczego ugrupowania butyrolaktonu. 30
185 PRZYKŁAD 11 Glukagon Cys 24 (S-karboksymetyl) [034] 18,1 mg glukagonu Cys 24 (1-29) rozpuszczono w 9,4 ml 0,1 M buforu fosforanu sodu (ph = 9,2) i dodano 0,6 ml roztworu kwasu bromooctowego (1,3 mg/ml w acetonitrylu). Reakcję mieszano w temperaturze pokojowej i postęp reakcji kontrolowano analityczną metodą HPLC. Po 1 godz. dodano jeszcze 0,1 ml roztworu kwasu bromooctowego. Reakcję mieszano jeszcze przez 60 min, a następnie zakwaszono za pomocą roztworu wodnego kwasu octowego i nałożono na preparatywną kolumnę w układzie faz odwróconych Kromasil C18 2,2 x 2 cm w celu oczyszczenia. Do FPLC Pharmacia zastosowano gradient acetonitrylu (przepływ = 4 ml/min) zbierając frakcje w odstępach min z monitorowaniem w paśmie UV przy 214 nm (2,0 A). A = 0,1% TFA, B = 0,1% TFA/0% ACN. Frakcje połączono, zamrożono i poddano liofilizacji otrzymując kilka mg produktu. Analityczna metoda HPLC wykazała czystość wynoszącą 90%, a analiza widma masowego metodą MALDI potwierdziła masę 31 pożądanego produktu. PRZYKŁAD 12 Glukagon Cys 24 maleimido, PEG-3.4K-dimer [0346] 16 mg glukagonu Cys 24 i 1,02 mg Mal-PEG-Mal- 3400, poli(etylenoglikolo)-bis-maleimidu, śr. m.cz (Nektar Therapeutics) rozpuszczono w 3, ml roztworu soli fizjologicznej buforowanej fosforanem i dodano 0, ml 0,01 M EDTA oraz reakcję mieszano w temperaturze pokojowej. Po 16 godz. dodano jeszcze 16 mg glukagonu Cys 24 i dalej mieszano. Po około 40 godz. mieszaninę reakcyjną nałożono na kolumnę Pharmacia

186 18 PepRPC 16/ i stosowano gradient acetonitrylu wykorzystując system do FPLC Pharmacia i zbierając frakcje w odstępach 2 min z monitorowaniem w paśmie UV przy 214 nm (2,0 A). Przepływ = 2 ml/min, A = 0,1% TFA, B = 0,1% TFA/0% ACN. Frakcje połączono, zamrożono i poddano liofilizacji otrzymując,4 mg. Analityczna metoda HPLC wykazuje czystość wynoszącą 90%, a analiza widma masowego metodą MALDI wykazała składnik w zakresie zgodny z pożądanym dimerem. 1 2 PRZYKŁAD 13 Synteza laktamów glukagonu [0347] 28 mg (0,2 mmol) żywicy metoksybenzhydrylaminowej (Midwest Biotech) nałożono do naczyń reakcyjnych o objętości 60 ml i złożono następującą sekwencję stosując zmodyfikowany syntezator peptydów Applied Biosystems 430A z wykorzystaniem sprzęgań pojedynczych Boc z aktywacją DEPBT. HSQGTFTSDYSKYLDERRAQDFVQWLMNT-NH2 (12-16 Laktam) [0348] Zastosowano następujące grupy zabezpieczające łańcuch boczny: Arg(Tos), Asp(OcHx), Asn(Xan), Glu(OFm), His(BOM), Lys(Fmoc), Ser(Bzl), Thr(Bzl), Trp(CHO) Tyr(Br-Z). Lys(Cl-Z) zastosowano w położeniu 12, jeśli laktamy składano z reszt 16-, -24 lub 24-
187 Ukończoną peptydylożywicę poddawano działaniu roztworu % piperydyny w dimetyloformamidzie przez jedną godzinę z obracaniem w celu usunięcia grupy formylowej z Trp, a także grup blokujących Fmoc i OFm z Lys12 i Glu16. Po potwierdzeniu usunięcia na podstawie dodatniego wyniku próby ninhydrynowej żywicę przemywano dimetyloformamidem, a następnie dichlorometanem i ponownie dimetyloformamidem. Żywicę poddawano działaniu mg (1 mmol) heksafluorofosforanu benzotriazol-1- iloksy-tris-pirolidynofosfoniowego (PyBOP) w dimetyloformamidzie i diizopropyloetyloaminie (DIEA). Reakcję prowadzono przez 8- godzin i cyklizację potwierdzano na podstawie ujemnego wyniku próby ninhydrynowej. Żywicę przemywano dimetyloformamidem, a następnie dichlorometanem, a później poddawano działaniu kwasem trifluorooctowym przez minut. Usunięcie grupy Boc potwierdzano na podstawie dodatniego wyniku próby ninhydrynowej. Żywicę przemywano dimetyloformamidem i dichlorometanem oraz suszono, a następnie przenoszono do naczynia reakcyjnego z kwasem fluorowodorowym (HF). Dodawano 00 μl p-krezolu i wrzucano mieszadełko magnetyczne. Naczynie podłączano do aparatu do HF (Penisula Labs), chłodzono na łaźni z suchym lodem/metanolem, usuwano powietrze i w naczyniu skraplano około ml ciekłego kwasu fluorowodorowego. Reakcję mieszano na łaźni lodowej przez 1 godzinę, a następnie HF usuwano pod próżnią. Pozostałość zawieszano w eterze etylowym, ciała stałe odsączano, przemywano eterem i peptyd solubilizowano za pomocą 10 ml % acetonitrylu/1% kwasu octowego.
188 [0349] Analityczną metodę HPLC nieoczyszczonego solubilizowanego peptydu przeprowadzono w następujących warunkach [4,6 X 30 mm Xterra C8, 1,0 ml/min, 2 nm, bufor A 0,1% TFA/% ACN, bufor B 0,1% TFA/0% ACN, gradient -9% B w ciągu 1 minut]. Ekstrakt rozcieńczono dwukrotnie wodą i nałożono na preparatywną kolumnę w układzie faz odwróconych Vydac C4 2,2 x 2 cm oraz eluowano stosując gradient acetonitrylu i wykorzystując system do HPLC Waters (bufor A 0,1% TFA/% ACN, bufor B 0,1% TFA/% ACN oraz gradient 0-0% B w ciągu 1 minut przy przepływie 1,00 ml/min. Analiza oczyszczonego peptydu metodą HPLC wykazała czystość przekraczającą 9%, a analiza widma masowego metodą jonizacji z elektrorozpylaniem potwierdziła masę 306 Da dla laktamu. W podobny sposób otrzymano laktamy z reszt 16-, -24 i PRZYKŁAD 14 Testy rozpuszczalności glukagonu: [030] Otrzymano roztwór (1 mg/ml lub 3 mg/ml) glukagonu (lub analogu) w 0,01 N HCl. 0 μl roztworu podstawowego rozcieńczono do 1 ml za pomocą 0,01 N HCl i wyznaczono absorbancję w paśmie UV (276 nm). ph pozostałego roztworu podstawowego doprowadzono do wartości ph 7 za pomocą 0- μl 0,1 M Na2HPO4 (ph 9,2). Roztwór pozostawiono na noc w 4 C, a następnie odwirowano. Następnie 0 μl supernatantu rozcieńczono do 1 ml za pomocą 0,01 N HCl i wyznaczono absorbancję w paśmie UV (w dwóch powtórzeniach). [031] Początkowy odczyt absorbancji skorygowano względem wzrostu objętości i do wyznaczenia rozpuszczalności procentowej zastosowano następujący
189 188 wzór: Absorbancja końcowa / Absorbancja początkowa X 0 = rozpuszczalność procentowa PRZYKŁAD 1 Test wiązania receptora glukagonu [032] Powinowactwo peptydów do receptora glukagonu mierzono za pomocą testu wiązania kompetycyjnego wykorzystując metodę zbliżeniowego testu scyntylacyjnego. Seryjne 3-krotne rozcieńczenia peptydów otrzymane w buforze do zbliżeniowego testu scyntylacyjnego (0,0 M Tris-HCl, ph 7,, 0,1 M NaCl, 0,1% wag./obj. albuminy surowicy bydlęcej) wymieszano na 96-dołkowej płytce białej z przezroczystym dnem (Corning Inc., Acton, MA) z 0,0 nm (3-[ 12 I]- jodotyrozylo) Tyr glukagonu (Amersham Biosciences, Piscataway, NJ), 1-6 mikrogramów na dołek, fragmentami błony plazmatycznej otrzymanymi z komórek z nadekspresją ludzkiego receptora glukagonu i 1 mg/dołek kulek do zbliżeniowego testu scyntylacyjnego z aglutyniną typu A z zarodków pszennych poddaną działaniu polietylenoiminy (Amersham Biosciences, Piscataway, NJ). Po min wytrząsania przy 800 obr/min na wytrząsarce obrotowej płytkę inkubowano przez 12 godz. w temperaturze pokojowej, a następnie dokonano odczytu stosując licznik scyntylacyjny do cieczy MicroBeta140 (Perkin-Elmer, Wellesley, MA). Zmierzono radioaktywność wynikającą z wiązania nieswoistego (NSB) w dołkach z 4-krotnie większym stężeniem zimnego liganda natywnego od najwyższego stężenia w próbkach badanych i w dołkach bez związku kompetycyjnego wykrywano całkowitą związaną radioaktywność. Procentowe
190 wiązanie swoiste obliczano w następujący sposób: wiązanie swoiste (%) = ((związana NSB)/(całkowita związana NSB)) X 0. Wartości IC0 wyznaczono za pomocą oprogramowania Origin (OriginLab, Northampton, MA). PRZYKŁAD 16 Test czynnościowy synteza camp [033] Zdolność analogów glukagonu do indukowania camp mierzono w teście reporterowym z użyciem lucyferazy świetlika. Komórki HEK293 poddane kotransfekcji receptorem (receptor glukagonu, receptor GLP-1 lub receptor GIP) i genem lucyferazy połączonym z elementem reagującym na camp pozbawiono surowicy w wyniku hodowli przez 16 godz. w DMEM (Invitrogen, Carlsbad, CA), do której dodano 0,2% bydlęcej surowicy wzrostowej (HyClone, Logan, UT), a następnie inkubowano z seryjnymi rozcieńczeniami glukagonu, GLP-1, GIP lub nowymi analogami glukagonu przez godz. w 37 C, % CO2 w 96-dołkowych płytkach Biocoat opłaszczonych poli-dlizyną (BD Biosciences, San Jose, CA). Po zakończeniu inkubacji do każdego dołka dodano 0 mikrolitrów odczynnika substratu luminescencji LucLite (Perkin- Elmer, Wellesley, MA). Płytkę krótko wytrząsano, inkubowano przez min w ciemności i emisję światła mierzono stosując licznik scyntylacyjny do cieczy MicroBeta140 (Perkin-Elmer, Wellesley, MA). Wartości 0% stężenia efektywnego wyznaczono za pomocą oprogramowania Origin (OriginLab, Northampton, MA). 30
191 PRZYKŁAD 17 Test stabilności analogów glukagonu Cys-maleimido PEG [034] Każdy analog glukagonu rozpuszczono w wodzie lub PBS i przeprowadzono początkową analizę metodą HPLC. Po skorygowaniu ph (4,, 6, 7) próbki inkubowano przez określony czas w 37 C i ponownie poddano analizie metodą HPLC w celu określenia integralności peptydu. Wyznaczono stężenie odpowiedniego konkretnego peptydu i obliczono pozostały procent niezmienionego peptydu względem wartości z analizy początkowej. PRZYKŁAD 18 Otrzymywanie peptydów acylowanych i(lub) PEGylowanych [03] Peptydy acylowane i(lub) PEGylowane otrzymano w następujący sposób. Peptydy syntetyzowano na żywicy stanowiącej podłoże stałe stosując syntezator peptydów CS Bio 4886 lub syntezator peptydów Applied Biosystems 430A. Zastosowano neutralizację chemiczną in situ zgodnie z opisem w Schnolzer i wsp., Int. J. Peptide Protein Res. 40: (1992). W przypadku peptydów acylowanych docelowa reszta aminokwasu poddawana acylowaniu (np. położenie dziesiąte) jest podstawiona przez resztę Nε-FMOC-lizyny. W wyniku działania na ukończony peptyd zabezpieczony na końcu N przez BOC za pomocą % piperydyny w DMF przez 30 minut następuje usuwanie grup FMOC/formylowych. Sprzęganie z wolną resztą ε-amino-lys uzyskuje się w wyniku sprzęgania dziesięciokrotnego nadmiaru molowego aminokwasu zabezpieczonego wstawką FMOC (z FMOC-(N-BOC)-tryptofan- OH) lub łańcucha acylowego (z C17-COOH) oraz odczynnikiem sprzęgającym PyBOP lub DEPBT w DMF/DIEA. Następnie po usunięciu grupy FMOC z aminokwasu we
192 wstawce przeprowadza się powtórne sprzęganie z łańcuchem acylowym. Ostateczne działanie za pomocą 0% TFA powoduje usunięcie wszystkich grup zabezpieczających łańcuch boczny i N-końcowej grupy BOC. Peptydożywice neutralizuje się za pomocą % DIEA/DMF, suszy, a następnie odcina od podłoża stosując HF/p-krezol, 9:, w 0 C przez jedną godzinę. Po ekstrakcji eterem do solwatowania nieoczyszczonego peptydu stosuje się % roztwór HOAc. Następnie metodą ESI-MS sprawdza się próbkę roztworu, czy zawiera peptyd o odpowiedniej masie cząsteczkowej. Prawidłowe peptydy oczyszcza się metodą RP-HPLC stosując liniowy gradient % CH3CN/0,1% TFA do 0,1% TFA w 0% CH3CN. Do oczyszczania stosuje się kolumnę do białek Vydac C18 22 mm x mm. Elucja acylowanych analogów peptydów kończy się zazwyczaj przy stosunku buforu wynoszącym :80. Frakcje łączy się ze sobą, a czystość sprawdza analityczną metodą RP-HPLC. Czyste frakcje poddaje się liofilizacji otrzymując białe, stałe peptydy. [036] Jeśli peptyd zawiera mostek laktamowy i reszty docelowe przeznaczone do acylowania, przeprowadza się acylowanie zgodnie z opisem powyżej po dodaniu danego aminokwasu do szkieletu peptydowego. [037] W przypadku pegylowania peptydu 40 kda metoksypoli(etylenoglikolo)maleimidopropionamid (Chirotech Technology Ltd.) poddaje się reakcji z równoważnikiem molowym peptydu w 7 M moczniku, 0 mm buforze Tris-HCl stosując minimalną ilość rozpuszczalnika niezbędną do rozpuszczenia zarówno peptydu, jak i PEG i uzyskania przezroczystego roztworu (zazwyczaj poniżej 2 ml w przypadku reakcji z użyciem
193 mg peptydu). Rozpoczyna się silne mieszanie w temperaturze pokojowej przez 4-6 godzin i reakcję analizuje się analityczną metodą RP-HPLC. Produkty PEGylowane wyraźnie odróżniają się od materiału wyjściowego (mają krótsze czasy retencji). Oczyszczanie prowadzi się na kolumnie Vydac C4 w warunkach podobnych do zastosowanych do początkowego oczyszczania peptydów. Elucja następuje zazwyczaj przy przybliżonych stosunkach buforu 0:0. Frakcje czystego peptydu PEGylowanego zbiera się i poddaje liofilizacji. [038] Bada się aktywność biologiczną peptydów zgodnie z opisem powyżej w przykładzie 16. Peptydy acylowane mogą wykazywać zwiększoną siłę działania wobec receptora GLP-1. Zastosowanie wstawki tryptofanowej może zapewnić zwiększoną siłę działania wobec receptora glukagonu. [039] Mimo że acylowanie może zwiększyć okres półtrwania peptydu do kilku godzin lub więcej, PEGylowanie z powtórzeniami w zakresie dziesiątek kda może być jeszcze skuteczniejsze. Otrzymuje się peptydy zawierające oba rodzaje modyfikacji. Oczekuje się, że te peptydy będą miały wydłużony okres półtrwania w krążeniu, a także odporność na DPP-IV i inne proteazy. PRZYKŁAD Wpływ in vivo na wzrost masy ciała, łaknienie i poziomy glukozy we krwi [0360] Następujące peptydy zsyntetyzowano zasadniczo zgodnie z opisem powyżej. (A) Pegylowany peptyd o działaniu koagonistycznym wobec glukagonu/glp-1 (chimera 2 AIB2 C24 40K PEG, będąca peptydem chimery 2 (patrz przykład 21) zmodyfikowanym
194 dodatkowo przez AIB w położeniu 2, Cys w położeniu 24, dołączoną do grupy 40K PEG); (B) pegylowany antagonista GIP (Pro3 C24 GIP NH2 (1-42) 40K PEG, obejmujący aminokwasy 1-42 w GIP (sekwencja natywnego GIP to NR. ID. SEKW.: 4) zmodyfikowany przez Pro w położeniu 3, Cys w położeniu 24, dołączoną do grupy 40K PEG i amid zamiast C-końcowego karboksylanu); (C) agonista GIP (AIB2 C24 GIP (1-42) 40K PEG, obejmujący aminokwasy 1-42 w GIP (sekwencja natywnego GIP to NR. ID. SEKW.: 4) zmodyfikowany przez AIB w położeniu 2, Cys w położeniu 24, dołączoną do grupy 40K PEG); oraz [0361] Peptydy badano in vivo za pomocą iniekcji podskórnych różnych peptydów lub samego nośnika u myszy z otyłością wywołaną dietą (DIO)(70 lub 2 nmol/kg/tydzień). Każda grupa zawierała 8 myszy, a ich początkowa średnia masa ciała wynosiła 0 g. Okresowo mierzono masę ciała, skład ciała, przyjmowanie pokarmu i poziomy glukozy we krwi. [0362] Jak pokazano na figurach 1 do 3, ani peptyd o działaniu antagonistycznym wobec GIP, ani agonistycznym wobec GIP nie był skuteczny w zmniejszaniu masy ciała, skumulowanego przyjmowania pokarmu i poziomów glukozy we krwi u myszy w porównaniu do pegylowanego peptydu działającego koagonistycznie wobec glukagonu/glp-1. PRZYKŁAD 21 Aktywność GIP, GLP-1 i glukagonu w przypadku peptydów [0363] Peptydy według NR. ID. SEKW.: -94 (z których każdy zawierał amid zamiast C-końcowego karboksylanu) zsyntetyzowano zasadniczo zgodnie z opisem powyżej i badano in vitro pod kątem aktywności wobec receptora

195 194 GIP, receptora GLP-1 i receptorów glukagonu zgodnie z przykładem 16. Wartość EC0 każdego peptydu przedstawiono w tabeli 1.
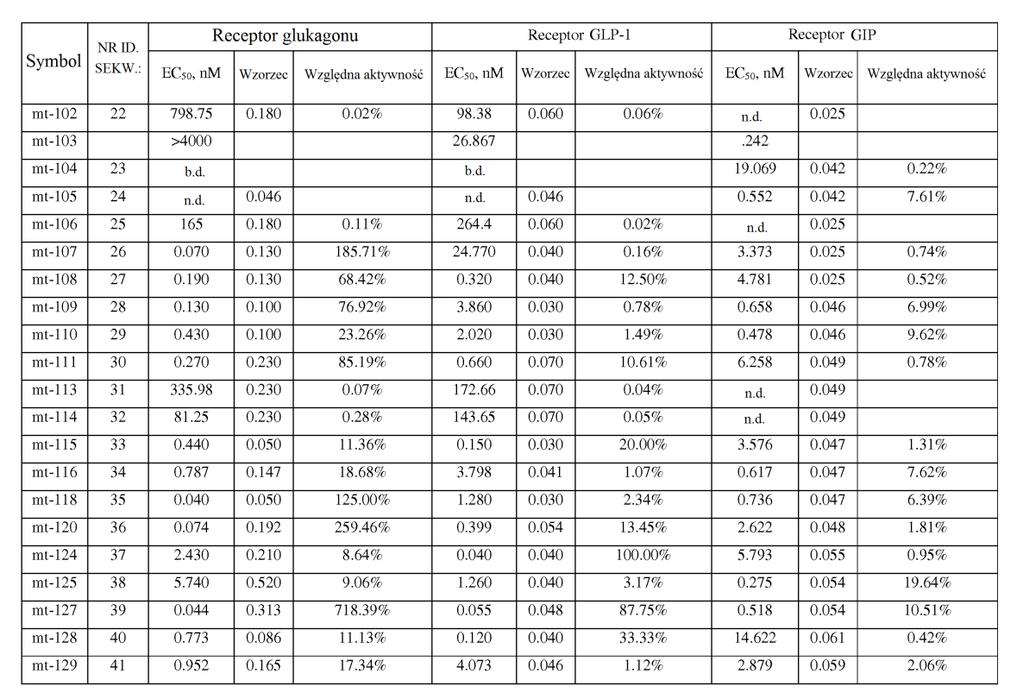
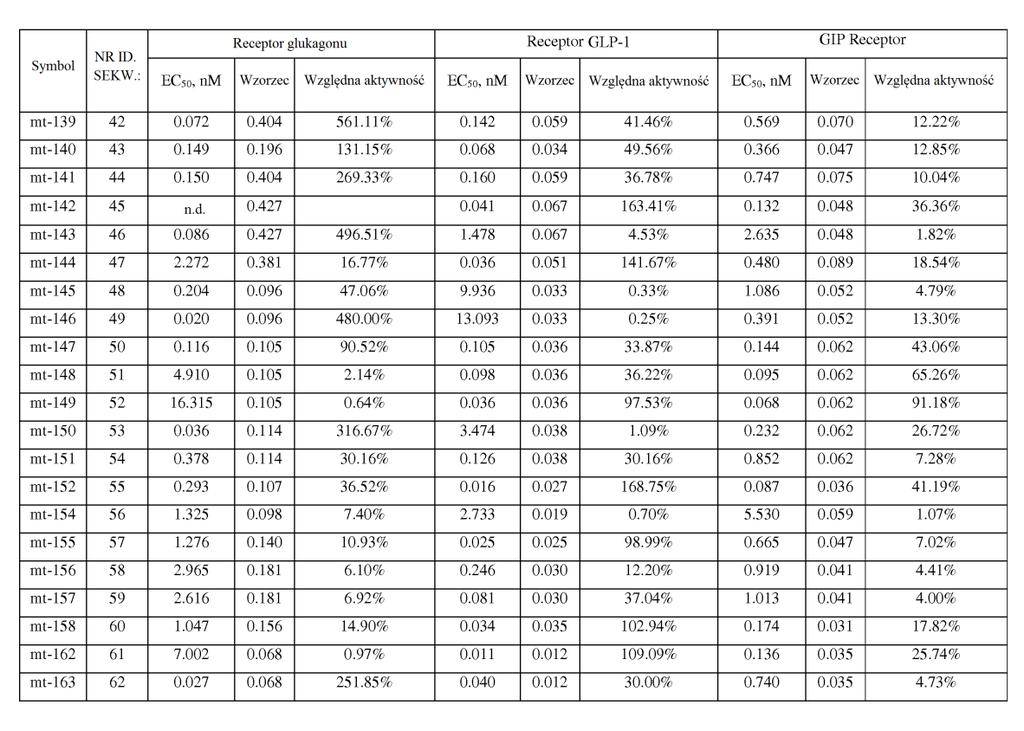
196 19

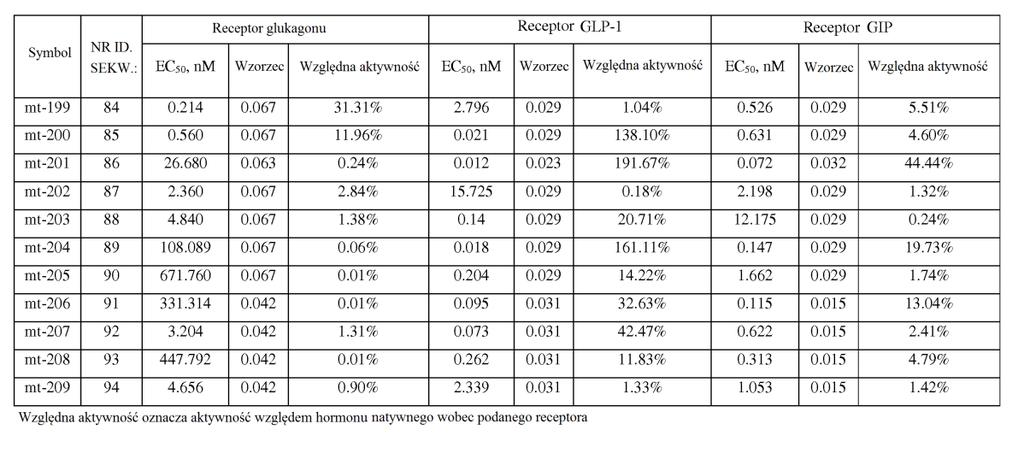
197 196
198 [0364] Na podstawie tych danych stwierdzono, że peptydy mt-140, mt-147, mt-11, mt-12, mt-18, mt-164, mt-16, mt-166, mt-169, mt-170, mt-172, mt-17 i mt-179 stanowiły przykładowe peptydy działające triagonistycznie wobec GLP-1/GIP/glukagonu, natomiast peptydy mt-148, mt-149, mt-162, mt-174, mt-178, mt-1 i mt-4 stanowiły przykładowe peptydy działające koagonistycznie wobec GLP-1/GIP, a peptydy mt-116, mt- 176, mt-177 i mt-182 stanowiły przykładowe peptydy działające koagonistycznie wobec GIP/glukagonu. PRZYKŁAD 22 [036] Peptyd działający triagonistycznie wobec GLP- 1/GIP/glukagonu (mt-170), peptyd działający koagonistycznie wobec GLP-1/GIP (mt-178) i dwa peptydy działające koagonistycznie wobec GIP/glukagonu (mt-182 i mt-179) według przykładu 21 zbadano in vivo za pomocą iniekcji podskórnych tych peptydów u myszy z otyłością wywołaną dietą (DIO), a także peptyd działający koagonistycznie wobec glukagonu/glp-1 (chimera 2 AIB2 (natywna sekwencja aminokwasowa glukagonu (NR. ID. SEKW.: 1) zawierająca następujące modyfikacje: Gln w położeniu 17, Ala w położeniu 18, Lys w położeniu, Glu w położeniu 21, Ile w położeniu 23 i Ala w położeniu 24 i C-końcowy amid ( chimera 2 ) z dodatkową modyfikacją AIB w położeniu 2) lub chimerę 2 AIB2 laktam (taki sam jak chimera 2 AIB2 bez dodatkowych modyfikacji Glu w położeniu 16 i Lys w położeniu, przy czym mostek laktamowy obejmuje łańcuchy boczne Glu16 i Lys) lub sam nośnik raz w tygodniu (70 lub 2 nmol/kg/tydzień). Każda grupa zawierała 8 myszy, a ich początkowa średnia masa ciała wynosiła 0 g.
199 Okresowo mierzono masę ciała. [0366] Jak pokazano na figurze 4, peptyd o działaniu triagonistycznym i peptyd o działaniu koagonistycznym wobec GLP-1/GIP były nieco bardziej skuteczne w zmniejszeniu masy ciała u myszy w porównaniu z chimerą 2 AIB2, ale mniej skuteczne niż chimera 2 AIB2 laktam, co potwierdza największą zdolność do zmniejszania masy ciała. W przeciwieństwie do tego obydwa peptydy o działaniu koagonistycznym wobec GIP/glukagonu, w szczególności mt-182, były mniej skuteczne w zmniejszaniu masy ciała. PRZYKŁAD 23 [0367] Peptyd działający triagonistycznie wobec GLP- 1/GIP/glukagonu (mt-170) i peptyd działający koagonistycznie wobec GLP-1/GIP (mt-178), agonistę GLP (zawierającego NR. ID. SEKW.: 3 z Glu w położeniu 16) lub sam nośnik badano in vivo przez wstrzyknięcie podskórne tych peptydów u myszy z otyłością wywołaną dietą (DIO) raz w tygodniu ( nmol/kg/tydzień przez 4 tygodnie lub 3 nmol/kg/tydzień przez 2 tygodnie). Każda grupa zawierała 8 myszy, a początkowa średnia masa ciała każdej myszy wynosiła 49 g. Okresowo mierzono masę ciała i poziomy glukozy we krwi. Jak pokazano na figurach i 6, zarówno peptyd o działaniu koagonistycznym wobec GLP-1/GIP, jak i o działaniu triagonistycznym były skuteczniejsze w zmniejszaniu masy ciała i poziomów glukozy we krwi w porównaniu do agonisty GLP-1. PRZYKŁAD 24 [0368] Wpływ stabilizacji helisy alfa analogów na bazie glukagonu za pomocą aminokwasu alfa,alfa-
200 199 1 dipodstawionego zamiast laktamu badano zastępując laktam w mt-16 (NR. ID. SEKW.: 64) i w mt-170 (NR. ID. SEKW.: 69) przez AIB w położeniu 16. Peptyd zawierający sekwencję mt-16 z AIB w położeniu 16 zamiast laktamu oznaczono jako mt-241 i miał on sekwencję aminokwasową według NR. ID. SEKW.: 167, a peptyd zawierający sekwencję mt-170 z AIB w położeniu 16 zamiast laktamu oznaczono jako mt-248 i miał on sekwencję aminokwasową według NR. ID. SEKW.: 173. [0369] Otrzymano ponadto dodatkowe peptydy liniowe niezawierające mostka laktamowego i zawierające AIB w położeniu 16 i(lub) zasadniczo zgodnie z powyższym opisem. Te peptydy oznaczono jako mt-242, mt-249, mt-, mt-21, mt-22, mt-2, mt-28 i mt- 29 oraz miały one odpowiednio sekwencje aminokwasowe według NR. ID. SEKW.: 168, , 7, 8, 177 i 28. Aktywność biologiczną in vitro każdego z tych peptydów wobec receptorów glukagonu, GLP-1 i GIP badano zasadniczo zgodnie z opisem w przykładzie 16. Wyniki podano w tabeli 2.

201 0
202 1 1 [0370] Jak potwierdzają wyniki podane w tabeli 2, peptydy liniowe niezawierające laktamu zapewniały aktywność wobec receptora GIP, a także wobec receptorów glukagonu i(lub) GLP-1. Dokładniej mt-242, mt-248, mt- 249, mt-, mt-22, mt-2, mt-28 i mt-29 wykazywały aktywności peptydów działających triagonistycznie wobec glukagonu/glp-1/gip, przy czym mt-21 wykazywał aktywność peptydu działającego koagonistycznie wobec GLP-1/GIP. Peptyd mt-22, zawierający Lys w położeniu 16 i AIB w położeniu, wykazywał siłę działania wobec receptorów glukagonu i GLP-1 oraz wykazywał zwiększoną aktywność wobec receptora GIP. PRZYKŁAD 2 [0371] Otrzymano peptydy liniowe niezawierające pierścienia laktamowego i zawierające Lys lub podobną resztę w położeniu 16 i AIB w położeniu zasadniczo zgodnie z powyższym opisem. Peptydy miały sekwencje aminokwasowe według NR. ID. SEKW.: , i 166. Aktywność biologiczną in vitro peptydów wobec receptorów glukagonu, GLP-1 i GIP badano zasadniczo zgodnie z opisem w przykładzie 16. Wyniki podano w tabeli 3.
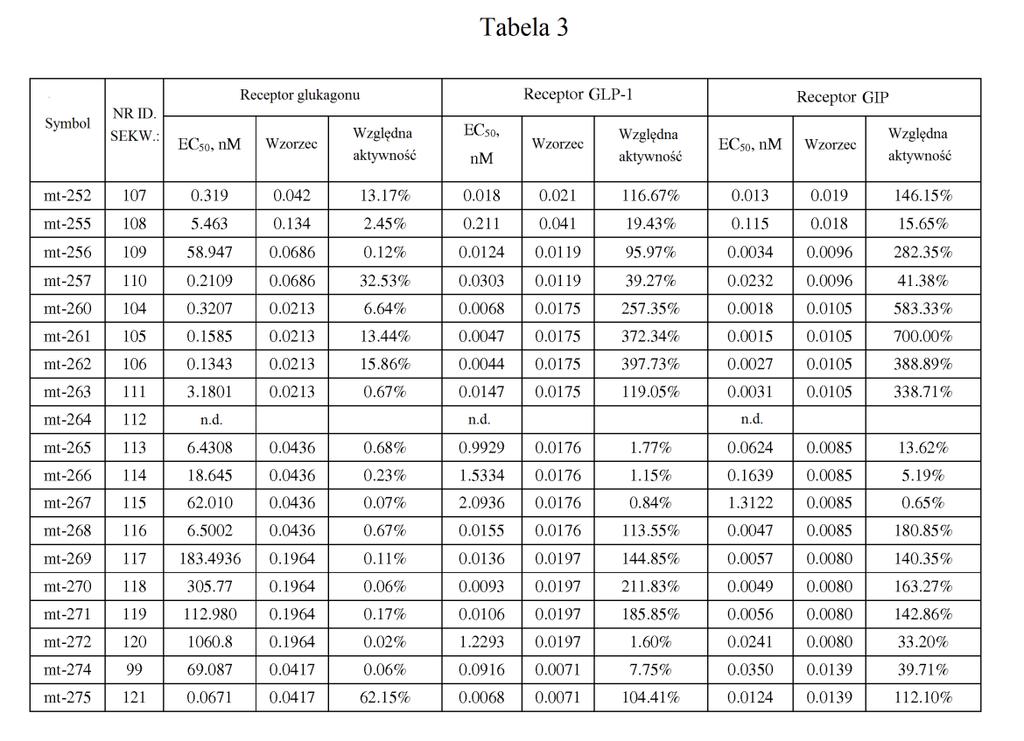

203 2
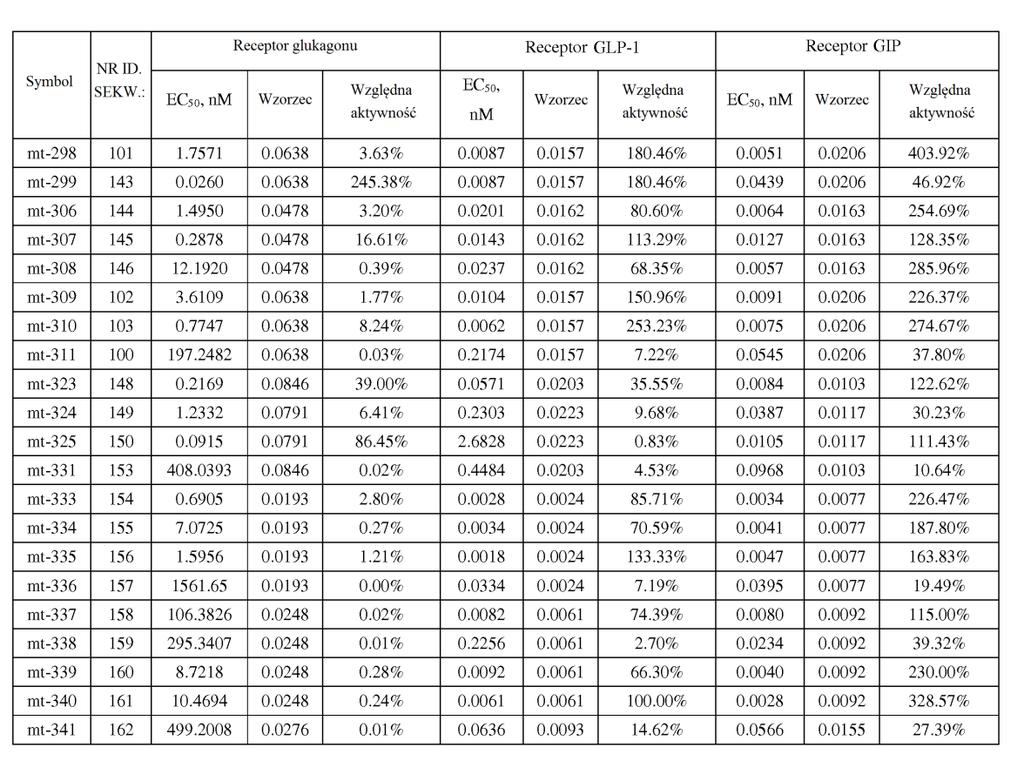
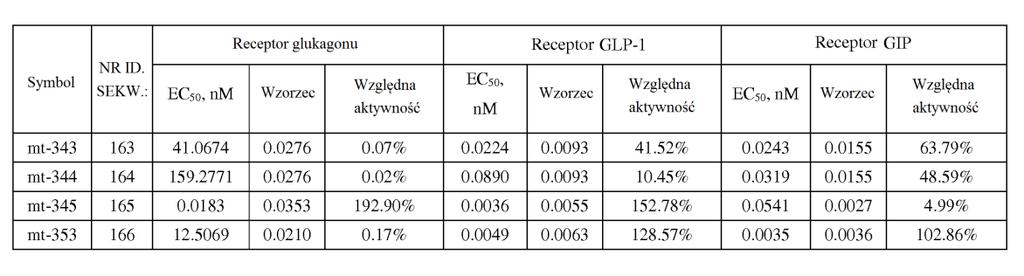
204 3
205 [0372] Jak pokazano w tabeli 3, peptydy liniowe były aktywne wobec receptora GIP oraz w wielu przypadkach peptyd był ponadto aktywny wobec receptora glukagonu i(lub) receptora GLP-1. Dokładniej wszystkie spośród mt-22, mt-2, mt-27, mt-260, mt-261, mt-262, mt-26, mt-266, mt-267, mt-27, mt-276, mt-277, mt-278, mt-279, mt-280, mt-286, mt-287, mt-288, mt-289, mt-290, mt-291, mt-292, mt-293, mt-29, mt-296, mt-297, mt-299, mt-306, mt-307, mt-3, mt-323, mt-324 i mt-34 wykazywały aktywność wobec receptorów GIP, GLP-1 i glukagonu, a pozostałe peptydy z tabeli 3 wykazywały aktywność wobec receptorów GIP i GLP-1 (oprócz mt-28, który wykazywał aktywność wyłącznie wobec receptora GIP, i mt-32, który wykazywał aktywność wobec receptorów glukagonu i GIP, ale nie wobec receptora GLP-1). [0373] Jeśli porównuje się dane dotyczące mt-22, który zawierał przedłużenie C-końcowe, z danymi dotyczącymi mt-27, mt-28 i mt-29, które to peptydy nie zawierały przedłużenia C-końcowego, było oczywiste, że przedłużenie C-końcowe zwiększało aktywności wobec wszystkich spośród receptorów glukagonu, GLP-1 i GIP. [0374] Jeśli porównuje się dane dotyczące mt-22, który zawierał Lys w położeniu 16, z danymi dotyczącymi mt- 27 i mt-276, które to peptydy zawierały odpowiednio reszty Orn i Dab w położeniu 16, było oczywiste, że Lys można zastąpić resztą podobną do Lys. [037] Ponadto, jeśli porównuje się dane dotyczące mt- 22, który zawierał Gln w położeniu 3, z danymi dotyczącymi mt-26 i mt-274, które to peptydy zawierają Glu w położeniu 3, jest oczywiste, że substytucja Gln w położeniu 3 przez resztę Glu pozwalała uzyskać
206 selektywność wobec receptorów GLP-1 i GIP względem receptora glukagonu. [0376] Wpływ acylowania za pomocą kwasów tłuszczowych C14, C16 i C18 był oczywisty na podstawie danych dotyczących mt-260 do mt-263 i mt-26 do mt-272. Na podstawie tych danych było oczywiste, że acylowanie za pomocą kwasów tłuszczowych C16 i C18 zapewniało zwiększoną aktywność wobec receptorów GLP-1 i GIP. Acylowanie tych peptydów pozwalało nawet uzyskać zwiększoną aktywność wobec receptora glukagonu, mimo że te peptydy zawierały substytucję Gln3Glu. Zwiększoną aktywność wobec receptora glukagonu można ponadto stwierdzić na podstawie danych o peptydach działających triagonistycznie mt-277 do mt-280. PRZYKŁAD 26 [0377] Aktywności in vivo pegylowanego, cyklicznego, zawierającego laktam peptydu mt-178 (NR. ID. SEKW.: 7) i pegylowanego peptydu liniowego niezawierającego laktamu mt-274 (NR. ID. SEKW.: 99) badano u myszy DIO i porównywano z aktywnością in vivo czystego peptydu będącego agonistą GLP-1 zawierającego strukturę na bazie GLP-1 według NR. ID. SEKW.: 179. Peptydy lub kontrolę (nośnik) wstrzykiwano dootrzewnowo myszom w dniu 0 w dawce 1, 3 lub nmol/kg/tydzień. [0378] U myszy przeprowadzono 1-godzinny test tolerancji glukozy (GTT) wstrzykując myszom dootrzewnowo 2% (wag./obj.) roztwór glukozy w soli fizjologicznej jedną godzinę po wstrzyknięciu jednego z peptydów lub kontrolę (nośnik). Roztwór glukozy w soli fizjologicznej podawano myszom w dawce 1, g na kg masy ciała myszy. Poziomy glukozy we krwi mierzono w
207 momencie wstrzyknięcia peptydu lub kontroli (nośnik) (- 60 min), w momencie wstrzyknięcia roztworu glukozy w soli fizjologicznej (0 min) lub 1, 30, 60 lub 1 min po wstrzyknięciu roztworu glukozy w soli fizjologicznej. Wyniki 1-godzinnego testu GTT przedstawiono na figurze 7. [0379] U myszy przeprowadzono ponadto 24-godzinny test GTT w ten sam sposób co 1-godzinny GTT, oprócz tego, że roztwór glukozy w soli fizjologicznej podano myszom 24 godziny po wstrzyknięciu peptydu lub kontroli (nośnik). Wyniki 24-godzinnego testu GTT przedstawiono na figurze 8. [0380] Całkowite poziomy glukozy we krwi u każdej myszy mierzono 0 i 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik) oraz przedstawiono na figurze 9. [0381] Ponadto masę ciała każdej myszy mierzono 0, 1, 3, i 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik). Zmianę procentową masy ciała myszy przedstawiono na figurze. [0382] Jak pokazano na figurze, u myszy, którym wstrzyknięto mt-178 i mt-274, w dawce 3 lub nmol/kg/tydzień, nastąpiła utrata masy w tym samym lub większym stopniu co u myszy, którym wstrzyknięto peptyd kontrolny (agonistę GLP) (w dawce nmol/kg/tydzień). [0383] Jak pokazano na figurach 7 i 8, u myszy, którym wstrzyknięto mt-178, mt-274 lub peptyd kontrolny (agonistę GLP-1), stwierdzono zmniejszone poziomy glukozy we krwi w porównaniu do myszy, którym wstrzyknięto kontrolę (nośnik). Stwierdzono, że siła działania mt-178 i mt-274 jest większa niż w przypadku peptydu kontrolnego (agonisty GLP-1), ponieważ mt-178 i
208 mt-274 w dawce 3 nmol/kg/tydzień osiągnęły to samo działanie co peptyd kontrolny (agonista GLP-1) w dawce nmol/kg/tydzień. Stwierdzono, że siła działania mt- 274 była największa, ponieważ ten peptyd w dawce 1 nmol/kg/tydzień osiągnął podobne wyniki co peptyd kontrolny (agonista GLP-1) w dawce nmol/kg/tydzień. PRZYKŁAD 27 [0384] Te same peptydy, które opisano w przykładzie 26, badano ponownie u myszy, oprócz tego, że myszom wstrzyknięto podskórnie większe dawki ( lub 3 nmol/kg/tydzień) peptydów. W tych dawkach badano ponadto dwa dodatkowe peptydy: jeden peptyd mający tę samą strukturę co mt-178, ale zawierający grupę PEG dołączoną do Cys w położeniu 40 poprzez bardziej stabilne wiązanie tioeterowe (-SCH2CO-) utworzone w wyniku substytucji nukleofilowej w przeciwieństwie do tradycyjnego wiązania tioeterowego tworzonego przez maleimid-peg w reakcji addycji Michaela jak w mt-178) i drugi peptyd mający tę samą strukturę co mt-274, ale zawierający grupę PEG dołączoną do Cys w położeniu 40 poprzez wiązanie tioeterowe (-SCH2CO-) utworzone w wyniku substytucji nukleofilowej. Te peptydy określa się w niniejszym dokumencie odpowiednio jako mt-178(te) i mt-274(te). [038] Całkowite poziomy glukozy we krwi u myszy mierzono 0 i 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik) oraz przedstawiono na figurze 11. Całkowite zmiany poziomu glukozy we krwi u myszy mierzono 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik) oraz przedstawiono na figurze 12. [0386] Masę ciała każdej myszy mierzono 0, 1, 3,, 7 i
209 dni po wstrzyknięciu peptydu lub kontroli (nośnik). Zmiany procentowe masy ciała u myszy w zależności od czasu przedstawiono na figurze 13, a całkowite zmiany masy u myszy mierzonej 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik) przedstawiono na figurze 14. [0387] Jak pokazano na figurze 12, całkowita zmiana poziomu glukozy we krwi zmniejszyła się u wszystkich myszy, którym wstrzyknięto którykolwiek z badanych peptydów, w porównaniu do myszy, którym wstrzyknięto kontrolę (nośnik). Najbardziej znaczące zmiany poziomu glukozy we krwi stwierdzono u myszy, którym wstrzyknięto mt-274 lub mt-178(te) w dawce 3 nmol/kg/tydzień. [0388] Jak pokazano na figurze 14, całkowita zmiana masy ciała zmniejszyła się u wszystkich myszy, którym wstrzyknięto którykolwiek z badanych peptydów, w porównaniu do myszy, którym wstrzyknięto kontrolę (nośnik). Podobnie jak w przypadku całkowitej zmiany poziomu glukozy, najbardziej znaczące zmiany masy ciała stwierdzono u myszy, którym wstrzyknięto mt-274 lub mt- 178(TE) w dawce 3 nmol/kg/tydzień. PRZYKŁAD 28 [0389] Aktywności in vivo peptydów, które opisano w przykładzie 26, porównano z aktywnościami in vivo odmian acylowanych liniowego peptydu mt-274. Dokładniej otrzymano i zbadano trzy odmiany acylowane mt-274, w których aminokwas C-końcowy (reszta Lys) był połączony kowalencyjnie z tłuszczową grupą acylową C16, tłuszczową grupą acylową C14 lub tłuszczową grupą acylową C18. Te peptydy określa się w niniejszym dokumencie odpowiednio jako mt-298, mt-309 i mt-3.
210 Podobnie jak peptyd macierzysty mt-274 peptydy acylowane także zawierały grupę PEG o masie 40 kda. Jednak grupa PEG w peptydach acylowanych była dołączona kowalencyjnie do łańcucha bocznego reszty Cys w położeniu 24 peptydów. Sekwencje aminokwasowe peptydów acylowanych mt-298, mt-309 i mt-3 podano w niniejszym dokumencie odpowiednio jako NR. ID. SEKW.: 1-3. [0390] Otrzymano i zbadano odmianę nieacylowaną mt-298, mt-9 i mt-3 (określaną poniżej jako mt-311). Peptyd mt-311 różnił się od mt-274 tym, że mt-311 był dołączony kowalencyjnie do grupy PEG poprzez łańcuch boczny reszty Cys w położeniu 24 (w przeciwieństwie do reszty Cys na końcu C występującej w mt-274), a reszta C-końcowa w mt-311 stanowiła resztę Lys, a nie resztę Cys, która występowała w mt-274. [0391] Peptydy lub kontrolę (nośnik) wstrzykiwano podskórnie myszom DIO w dniu 0 w dawce nmol/kg przez jeden tydzień. [0392] Poziomy glukozy we krwi u myszy mierzono 0 i 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik) oraz przedstawiono na figurze 1. Masę ciała każdej myszy mierzono 0, 1, 3, i 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik). Zmiany procentowe masy ciała u myszy w zależności od czasu przedstawiono na figurze 16, a całkowite zmiany masy ciała u myszy mierzonej 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik) przedstawiono na figurze 17. [0393] Jak pokazano na figurze 17, w przypadku myszy, którym wstrzyknięto którykolwiek z peptydów, stwierdzono spadek masy ciała w porównaniu do myszy, którym wstrzyknięto kontrolę (nośnik). U myszy, którym
211 wstrzyknięto pegylowany, cykliczny peptyd zawierający laktam (mt-178), stwierdzono największy spadek masy ciała 7 dni po wstrzyknięciu. [0394] Jak pokazano na figurze 1, poziomy glukozy we krwi u myszy, którym wstrzyknięto mt-178, mt-274, mt- 311 lub ich odmiany acylowane C14 lub C16, zmniejszył się. Stwierdzono, że acylowanie za pomocą tłuszczowej grupy acylowej C18 nie powoduje zmniejszenia poziomów glukozy we krwi, co wskazuje na to, że wielkość grupy acylowej może mieć istotne znaczenie dla działań peptydów polegającego na obniżeniu poziomu glukozy. PRZYKŁAD 29 [039] Otrzymano liniowe peptydy na bazie glukagonu i niezawierające laktamu, acylowane, ale nie pegylowane, zasadniczo zgodnie z powyższym opisem. Dokładniej otrzymano mt-260 zawierający tłuszczową grupę acylową C14 na aminokwasie C-końcowym (NR. ID. SEKW.: 4), mt- 261 zawierający tłuszczową grupę acylową C16 na aminokwasie C-końcowym (NR. ID. SEKW.: ) i mt-262 zawierający tłuszczową grupę acylową C18 na aminokwasie C-końcowym (NR. ID. SEKW.: 6). Struktury każdego z tych peptydów były podobne do struktur mt-298, mt-309 i mt-3, ale różniły się tym, że mt-260, mt-261 i mt-262 zawierały resztę Asn zamiast pegylowanej reszty Cys w położeniu 24. [0396] Peptydy mt-260, mt-261 lub mt-262, peptyd kontrolny (liraglutyd, acylowany analog GLP-1 lub kontrolę (nośnik) wstrzykiwano myszom raz na dobę w dawce 2 lub 12 nmol/kg przez 7 dni. [0397] Poziomy glukozy we krwi u myszy mierzono 0 i 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik) oraz
212 przedstawiono na figurze 18. Masę ciała każdej myszy mierzono 0, 1, 3, i 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik). Zmiany procentowe masy ciała u myszy w zależności od czasu przedstawiono na figurze 19, a całkowite zmiany masy ciała u myszy mierzonej 7 dni po wstrzyknięciu peptydu lub kontroli (nośnik) przedstawiono na figurze. [0398] Jak pokazano na figurze 18, stwierdzono bardzo znaczne wpływy acylowanych, niepegylowanych peptydów linowych (mt-260, mt-261 i mt-262) na poziomy glukozy we krwi. W dawce 2 nmol/kg te peptydy spowodowały około 0% spadek poziomów glukozy we krwi, a w dawce wyższej peptydy te spowodowały spadek poziomów glukozy we krwi przekraczający 0%. [0399] Jak pokazano na figurze 19, każdy spośród acylowanych, niepegylowanych peptydów linowych (w dawce niskiej lub wysokiej) spowodował zmniejszenie masy ciała o większej wartości niż zmniejszenie masy ciała uzyskane dzięki liraglutydowi w niskiej dawce. Masa ciała nadal zmniejszała się w okresie 7 dni badania. [0400] Wyniki te sugerują, że acylowane, niepegylowane linowe peptydy na bazie glukagonu, mające aktywność wobec receptorów GIP i GLP-1, mogą w bardzo znaczący sposób zmniejszyć poziomy glukozy we krwi i masę ciała, co oznacza, że te peptydy można stosować w leczeniu zaburzeń metabolicznych, w tym cukrzycy, i w leczeniu otyłości. PRZYKŁAD 30 [0401] Linowy peptyd na bazie glukagonu mt-261 (NR. ID. SEKW.: ) badano w różnych dawkach u myszy DIO (N = 8 w grupie; średnia początkowa masa ciała = 48 g). Myszom
213 wstrzykiwano podskórnie raz na dobę przez jeden tydzień wyłącznie nośnik, liraglutyd (30 nmol/kg masy ciała) lub mt-261 (0,3, 1, 3, lub 30 nmol/kg masy ciała). [0402] Masę ciała myszy mierzono 0, 1, 3, i 7 dni po pierwszym wstrzyknięciu. Jak pokazano na figurze 21, wstrzyknięcie mt-261 lub liraglutydu spowodowało spadek masy u myszy. Peptyd mt-261 wykazywał większą siłę działania niż liraglutyd, ponieważ dawka 3 nmol/kg mt- 261 pozwalała uzyskać zasadniczo to samo działanie co dawka 30 nmol/kg liraglutydu (figura 21). [0403] Masę tłuszczu u myszy mierzono 7 dni po pierwszym wstrzyknięciu metodą obrazowania za pomocą jądrowego rezonansu magnetycznego. Jak pokazano na figurze 22, zwiększenie dawek mt-261 było zasadniczo skorelowane ze zmniejszeniem masy tłuszczu. Masa tłuszczu u myszy, którym wstrzyknięto 3 nmol/kg mt-261, była w przybliżeniu taka sama jak masa tłuszczu u myszy, którym wstrzyknięto 30 nmol/kg liraglutydu, co pokazuje większą siłę działania mt-261 w porównaniu do liraglutydu. [0404] Poziomy glukozy we krwi u myszy mierzono 0 i 7 dni po pierwszym wstrzyknięciu. Jak pokazano na figurze 23, dawki zaledwie 3 nmol/kg mt-261 doprowadziły do znacznego zmniejszenia poziomów glukozy we krwi. Zgodnie z wynikami badań dotyczących masy tłuszczu i masy ciała, zmniejszenie poziomów glukozy we krwi u myszy, którym wstrzyknięto 3 nmol/kg mt-261, było podobne do zmniejszenia poziomów glukozy we krwi u myszy, którym wstrzyknięto 30 nmol/kg liraglutydu, co pokazuje większą siłę działania mt-261 w porównaniu do liraglutydu.
214 [040] W oddzielnym doświadczeniu nieacylowaną odmianę mt-261, czyli mt-263 (NR. ID. SEKW.: 111), badano na dziewięciu grupach myszy C7BI/6 (8 myszy w grupie) pod kątem jej wpływów in vivo na masę ciała, przyjmowanie pokarmu, poziomy glukozy we krwi i masę tłuszczu. Wiek myszy wynosił 11 miesięcy i pozostawały one na diecie diabetogennej przez 9 miesięcy w okresie badania. Średnia masa ciała myszy wynosiła 7 g. Myszom wstrzykiwano podskórnie raz na dobę 3, lub 30 nmol/kg mt-263 przez jeden tydzień. Grupom kontrolnym podano kontrolę (nośnik) lub eksendynę 4 w dawce lub 30 nmol/kg/dobę. [0406] Aby ocenić wpływy in vivo na masę ciała, w dniach 0, 1, 3, i 7 mierzono masę ciała myszy, przy czym dzień 0 oznaczał pierwszy dzień wstrzykiwania. Jak pokazano na figurze 24, wstrzykiwanie mt-263 w dowolnej z trzech dawek doprowadziło do stałego zmniejszenia masy ciała w okresie badania trwającego 7 dni. Ponadto wpływ na masę ciała wydawał się zależeć od dawki, ponieważ zmiana całkowita masy ciała (pokazana na figurze 2) zwiększyła się po zwiększeniu dawki peptydu mt-263. Ponadto, jak pokazano na figurze 2, całkowite zmiany masy ciała uzyskane dzięki którejkolwiek z trzech dawek mt-263 były znacznie większe od całkowitej zmiany masy ciała uzyskanej dzięki wstrzyknięciu eksendyny 4 (niezależnie od dawki). [0407] Określono ponadto wpływy in vivo na przyjmowanie pokarmu, masę tłuszczu i poziom glukozy we krwi. Całkowite przyjmowanie pokarmu i masa tłuszczu mierzone w dniu 7 u myszy, którym wstrzyknięto mt-263, zmniejszyły się w porównaniu do kontroli (nośnik) i
215 eksendyny 4. Ponadto całkowite zmiany poziomów glukozy we krwi (mierzone w dniu 7 w porównaniu do poziomów zmierzonych w dniu 0) u myszy, którym wstrzyknięto mt- 263, znacznie zmniejszyły się w porównaniu do myszy, którym wstrzyknięto kontrolę (nośnik) lub eksendynę 4 (figura 26). Stwierdzono, że dawka nmol/kg peptydu jest optymalna, ponieważ pozwalała uzyskać największy spadek poziomów glukozy we krwi (prawie -80 mg/dl). [0408] Ponadto porównano wpływy in vivo na masę ciała, przyjmowanie pokarmu i poziomy glukozy we krwi peptydu mt-263 w porównaniu do peptydów mt-349 (NR. ID. SEKW.: 262), mt-280, mt-36 i mt-37 oraz kontroli (nośnik). Myszom podawano 30 nmol/kg/dobę jednego z peptydów przez 1 tydzień. Wszystkie peptydy były skuteczne w zmniejszaniu masy ciała u myszy w porównaniu do kontroli (nośnik). PRZYKŁAD 31 [0409] Zbadano wpływy in vivo liniowych, acylowanych peptydów stanowiących pochodne glukagonu mt-277, mt-278 i mt-279 oraz porównano do tych dla liraglutydu. Myszom DIO (8 myszy w grupie; średnia początkowa masa ciała = 1,4 g) wstrzykiwano podskórnie raz na dobę przez 1 tydzień kontrolę (nośnik) lub nmol/kg liraglutydu, mt-277, mt-278 lub mt-279. [04] Masę ciała myszy mierzono 0, 1, 3, i 7 dni po pierwszym wstrzyknięciu. Jak pokazano na figurze 27, wstrzyknięcie mt-277, mt-278 lub mt-279 spowodowało znaczący spadek masy u myszy. Ponadto wszystkie te peptydy wykazywały większą siłę działania niż liraglutyd. [0411] Poziomy glukozy we krwi u myszy mierzono 0 i 7
216 dni po pierwszym wstrzyknięciu. Jak pokazano na figurze 28, każdy spośród mt-277, mt-278 i mt-279 doprowadził do znacznego zmniejszenia poziomów glukozy we krwi, który był znacznie większy od stwierdzonego u myszy, którym wstrzyknięto liraglutyd. PRZYKŁAD 32 [0412] Analog na bazie glukagonu aktywny wobec receptora GLP-1 zmodyfikowano tak, żeby zawierał przedłużenie C-końcowe zgodnie z sekwencją aminokwasową według NR. ID. SEKW.: 9 oraz dodatkowo zmodyfikowano tak, żeby zawierał Lys na końcu C według NR. ID. SEKW.: 9. Resztę Lys znajdującą się w położeniu 40 w analogu poddano acylowaniu za pomocą tłuszczowej grupy acylowej C14. Aktywność in vitro tego analogu acylowanego wobec receptorów glukagonu, GLP-1 i GIP badano zasadniczo zgodnie z opisem w przykładzie 16. Aktywność in vitro porównywano z aktywnością macierzystego analogu na bazie glukagonu, aktywnego wobec receptora GLP-1 niezawierającego przedłużenia C-końcowego i acylowania w położeniu 40. Analog acylowany z przedłużeniem C- końcowym wykazywał w przybliżeniu 1% wzrost aktywności wobec receptora GIP i w przybliżeniu 2% wzrost wobec receptora glukagonu. Stwierdzono, że aktywność wobec receptora GLP-1 faktycznie zmniejszyła się w wyniku stymulacji przez acylowany analog z przedłużeniem C- końcowym. Jednak aktywność ta była nadal większa niż 0% aktywności uzyskanej przez natywny GLP-1 wobec receptora GLP-1. PRZYKŁAD 33 [0413] Acylowane peptydy stanowiące analogi glukagonu (z których każdy zawierał amid zamiast C-końcowego
217 216 1 karboksylanu) zsyntetyzowano zasadniczo zgodnie z opisem powyżej. Peptydy mt-38, mt-367, mt-368 i mt-369 stanowiły acylowane monomery, a mt-34, mt-376 i mt-377 stanowiły acylowane dimery, przy czym każdy dimer zawierał dwa monomery połączone poprzez C-końcowe reszty Cys. Peptydy mt-367, mt-368 i mt-369 zawierały wstawkę dipeptydową γglu-γglu w celu umożliwienia dołączenia grupy acylowej, przy czym mt-38 był acylowany i nie zawierał wstawki. Peptydy mt-22, mt- 227 i mt-294 stanowiły pegylowane monomery zawierające mostek laktamowy między kwasem glutaminowym w położeniu 16 a lizyną w położeniu. Peptydy mt-22 i mt-227 zawierały wstawkę dipeptydową w celu umożliwienia dołączenia PEG, zaś mt-294 był acylowany poprzez tioeter otrzymany w wyniku reakcji z haloacetylem. Peptydy mt-36 i mt-37 służyły jako nieacylowane peptydy kontrolne, przy czym mt-37 zawierał Ile w położeniu 7, a mt-36 zawierał Thr. Aktywność in vitro wszystkich peptydów wobec receptora GIP, receptora GLP- 1 i receptorów glukagonu badano zasadniczo zgodnie z opisem w przykładzie 16. Wartości EC0 (nm) i aktywność każdego peptydu względem hormonu natywnego przedstawiono w tabeli 4.
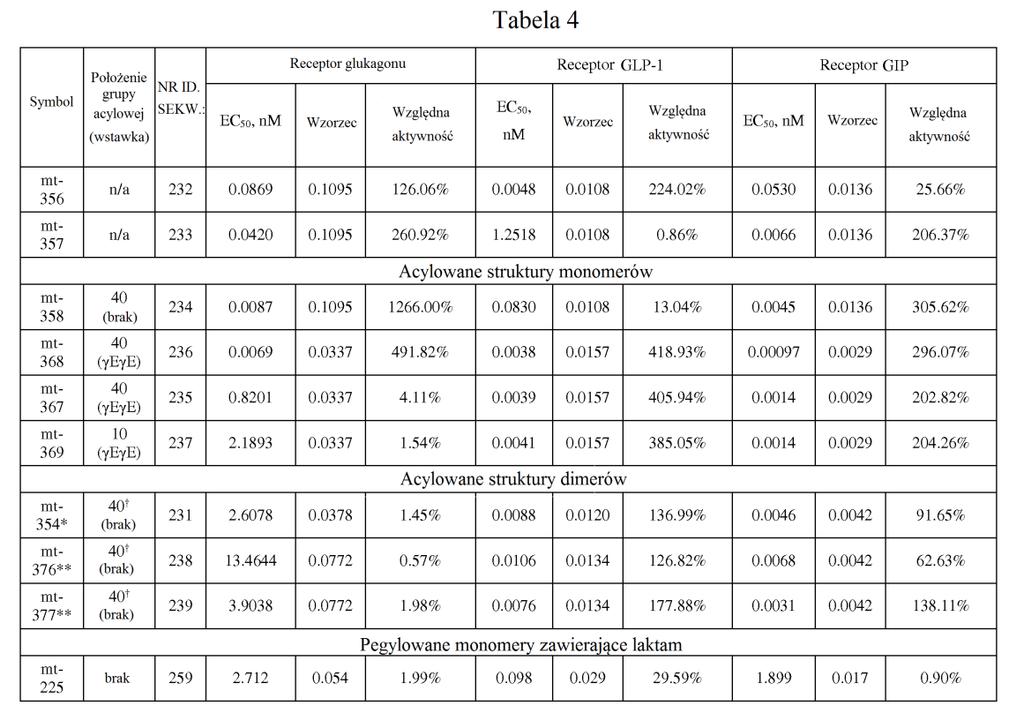

218 217
![218 [0414] Jak pokazano](/docs-images/65/53262204/images/219-0.jpg "w tabeli 4, wszystkie")
219 218 [0414] Jak pokazano w tabeli 4, wszystkie trzy

220 219 1 acylowane dimery wykazywały silne działanie wobec receptorów GLP-1 i GIP. Ponadto aktywność wobec receptora GLP-1 stwierdzona w przypadku mt-368 bardzo znacznie zwiększyła się w porównaniu do aktywności stwierdzonej w przypadku mt-38, a aktywność wobec receptora GIP stwierdzona w przypadku mt-368 zasadniczo utrzymała się w porównaniu mt-38, co wskazuje na to, że acylowanie peptydu glukagonu poprzez wstawkę pozwala zwiększyć aktywność wobec receptora GLP-1 przy utrzymaniu znacznej aktywności wobec receptora GIP. Stwierdzono, że acylowanie poprzez wstawkę w położeniu analogu glukagonu wydaje się mieć tak samo dobry wpływ jak położenie 40 w analogu glukagonu, ponieważ aktywności względne wobec receptorów GLP-1 i GIP były w przybliżeniu takie same w przypadku mt-367 i mt-369. PRZYKŁAD 34 [041] Otrzymano dwa peptydy acylowane będące analogami glukagonu i mające tę samą sekwencję aminokwasową, ale różniące się łącznikiem do pegylowania, zasadniczo zgodnie z opisem w niniejszym dokumencie: mt-331 (NR. ID. SEKW.: 13) zawierał połączenie z PEG według struktury: 2 przy czym mt-311 (NR. ID. SEKW.: 0) zawierał połączenie z PEG według struktury:
![2 1 2 [0416] Te dwa peptydy lub kontrolę (nośnik) podawano we wstrzyknięciu podskórnym raz na dobę przez jeden tydzień kilku grupom myszy DIO (6 myszy w grupie; średnia masa ciała = 64,6 g).](/docs-images/65/53262204/images/221-0.jpg "Peptydy podawano w dawce lub 3 nmol/kg. [0417] Masy ciała myszy mierzono 0, 1, 3, i 7 dni po podaniu peptydów lub kontroli (nośnika).")
221 2 1 2 [0416] Te dwa peptydy lub kontrolę (nośnik) podawano we wstrzyknięciu podskórnym raz na dobę przez jeden tydzień kilku grupom myszy DIO (6 myszy w grupie; średnia masa ciała = 64,6 g). Peptydy podawano w dawce lub 3 nmol/kg. [0417] Masy ciała myszy mierzono 0, 1, 3, i 7 dni po podaniu peptydów lub kontroli (nośnika). Masa ciała u myszy, którym wstrzyknięto wyższą dawkę mt-311 lub mt- 331, stale się zmniejszała w okresie tego tygodnia. Całkowitą zmianę masy ciała (%)przedstawiono na figurze 29. Jak pokazano na tej figurze, całkowita zmiana masy ciała była największa w przypadku myszy, którym wstrzyknięto mt-311. [0418] Całkowite przyjmowanie pokarmu w każdej grupie myszy mierzono ponadto 0, 1, 3, i 7 dni po podaniu peptydów lub kontroli (nośnika). Jak pokazano na figurze 30, całkowite przyjmowanie pokarmu w grupach myszy, którym wstrzyknięto jedną z dawek mt-311 lub mt- 331, zmniejszyło się w porównaniu do myszy, którym wstrzyknięto kontrolę (nośnik). [0419] Poziomy glukozy we krwi w każdej grupie myszy mierzono 0 i 7 dni po podaniu peptydów lub kontroli (nośnika). Poziomy glukozy we krwi u myszy zmniejszyły się po podaniu wyższej dawki mt-311 lub mt-331. Jak pokazano na figurze 31, zmniejszenie poziomów glukozy we krwi było większe u myszy, którym wstrzyknięto 3 nmol/kg mt-331.
222 [04] Zmierzono masę tłuszczu w każdej grupie myszy. Wydaje się jednak, że podawanie peptydów nie ma wpływu na masę tłuszczu. PRZYKŁAD 3 [0421] Zbadano wpływy in vivo dwóch peptydów mających tę samą sekwencję aminokwasową, ale różniących się brakiem grupy acylowej dołączonej do Lys w położeniu 40 (mt-331 (NR. ID. SEKW.: 13)) lub obecnością tłuszczowej grupy acylowej C14 (mt-33 (NR. ID. SEKW.: 166)) na masę ciała, przyjmowanie pokarmu, poziomy glukozy we krwi i masę tłuszczu u myszy C7BI/6 w wieku 7 miesięcy. Myszy pozostawały na diecie diabetogennej przez miesięcy, a średnia początkowa masa ciała wynosiła 3 g. Peptydy lub kontrolę (nośnik) podawano myszom we wstrzyknięciu podskórnym przez jeden tydzień w dawce 0,1, 0,3, 3 lub nmol/kg. [0422] Masę ciała mierzono 0, 1, 3, i 7 dni po podaniu peptydu lub kontroli (nośnika). Jak pokazano na figurze 32, całkowita zmiana masy ciała była najbardziej znacząca w przypadku myszy, którym wstrzyknięto nmol/kg mt-331 lub mt-33. [0423] Przyjmowanie pokarmu przez myszy mierzono 0, 1, 3, i 7 dni po podaniu peptydu lub kontroli (nośnik). Jak pokazano na figurze 33, całkowite przyjmowanie pokarmu przez myszy, którym wstrzyknięto 3 lub nmol/kg mt-331 lub mt-33, zmniejszyło się w porównaniu do myszy, którym podano kontrolę (nośnik). [0424] Ponadto kontrolowano poziomy glukozy we krwi u myszy. Jak pokazano na figurze 34, mt-331 doprowadził do zmniejszenia poziomów glukozy we krwi w sposób zależny od dawki. Dwie dawki mt-33 doprowadziły

223 222 1 ponadto do zmniejszenia poziomów glukozy we krwi. Nie stwierdzono znaczącego wpływu na poziomy masy tłuszczu w wyniku podawania któregokolwiek peptydu. PRZYKŁAD 36 [042] Zbadano wpływy in vivo trzech acylowanych peptydów o działaniu triagonistycznym mt-277, mt-278 i mt-279, mających strukturę odpowiednio o NR. ID. SEKW.: 123, 124 i 12, na masę ciała, poziomy glukozy we krwi i przyjmowanie pokarmu w 8 grupach myszy DIO (8 myszy w grupie). Peptydy miały tę samą sekwencję aminokwasową, ale różniły się wielkością tłuszczowej grupy acylowej, do której były dołączone. Jako kontrolę zastosowano liraglutyd w stężeniu nm/kg. Peptydy lub kontrolę (nośnik) podawano we wstrzyknięciu podskórnym raz na dobę przez jeden tydzień. [0426] Zbadano aktywności in vitro wobec receptorów glukagonu, GLP-1 i GIP, a aktywność procentową każdego peptydu względem hormonu natywnego przedstawiono poniżej w tabeli. [0427] Masę ciała myszy mierzono 0, 1, 3, i 7 dni po podaniu peptydów lub kontroli (nośnik). W okresie tygodnia masa ciała myszy, którym wstrzyknięto jeden z acylowanych peptydów o działaniu triagonistycznym,
224 bardzo znacznie się zmniejszyła w porównaniu do kontroli (nośnika). Jak pokazano na figurze 3, całkowita zmiana masy ciała (%) u myszy, którym wstrzyknięto jeden z acylowanych peptydów o działaniu triagonistycznym, wynosiła w przybliżeniu -1%, przy czym liraglutyd pozwolił na zmniejszenie masy ciała o poniżej %. PRZYKŁAD 37 [0428] Zbadano wpływ częstości dawkowania na skuteczność peptydu pegylowanego, acylowanego (mt-309; NR. ID. SEKW.: 2) i peptydu niepegylowanego, acylowanego (mt-261; NR. ID. SEKW.: ) w 7 grupach myszy DIO (8 myszy w grupie), których średnia masa ciała wynosiła 8 g. Peptydy wstrzykiwano myszom podskórnie raz na dobę w dawce nmol/kg, co drugi dzień w dawce nmol/kg lub raz na tydzień w dawce 30 nmol/kg. Badanie trwało 6 dni, dzięki czemu każda grupa myszy otrzymała do końca trwania badania dawkę 30 nmol/kg. Masę ciała i poziomy glukozy we krwi mierzono 0 i 6 dni po pierwszym podaniu. [0429] Jak pokazano na figurze 36, całkowita zmiana masy ciała (%) u myszy, którym mt-309 wstrzykiwano raz na tydzień, była w przybliżeniu identyczna jak całkowita zmiana masy ciała u myszy, którym ten sam peptyd wstrzykiwano raz na dobę. Ponadto, jak pokazano na figurze 36, całkowita zmiana masy ciała u myszy, którym mt-261 wstrzykiwano raz na dobę, była w przybliżeniu identyczna jak całkowita zmiana masy ciała u myszy, którym ten peptyd wstrzykiwano co drugi dzień. [0430] Te same tendencje co w przypadku masy ciała stwierdzono ponadto w odniesieniu do poziomów glukozy
225 we krwi (figura 37): wstrzykiwanie mt-309 raz na tydzień pozwoliło uzyskać ten sam spadek poziomów glukozy we krwi co wstrzykiwanie tego peptydu raz na dobę, a wstrzykiwanie mt-261 raz na dobę pozwoliło uzyskać ten sam spadek poziomów glukozy we krwi co wstrzykiwanie tego peptydu raz na dwa dni. PRZYKŁAD 38 [0431] Zbadano wpływ częstości dawkowania na skuteczność w przypadku peptydu acylowanego o działaniu agonistycznym wobec glukagonu, mt-261 (NR. ID. SEKW.: ) przez wstrzyknięcie podskórne 8 grupom myszy DIO (8 myszy w grupie) o początkowej masie ciała 6 g w dawce nmol/kg na dobę, nmol/kg co dwa dni, 1 nmol/kg co trzy dni lub 30 nmol/kg przez jeden dzień, tak że każda grupa otrzymała dawkę całkowitą 30 nmol/kg na tydzień. Wiek myszy wynosił 8 miesięcy i pozostawały one na diecie diabetogennej przez 6 miesięcy. Mierzono masę ciała, przyjmowanie pokarmu, poziomy glukozy we krwi i masę tłuszczu w każdej grupie. Jak pokazano na figurze 38, w przypadku myszy, którym peptyd wstrzykiwano co trzy dni, stwierdzono największy spadek masy ciała. Co ciekawe, u myszy, którym peptyd wstrzykiwano raz na dobę, i u myszy, którym peptyd wstrzykiwano raz na dwa dni, stwierdzono niemal identyczny spadek masy ciała. PRZYKŁAD 39 [0432] Następujące peptydy stanowiące analogi glukagonu i mające znaczącą aktywność agonistyczną wyłącznie wobec receptora glukagonu, a nie wobec receptora GIP oraz zawierające szkielet stanowiący peptyd J
 lub peptyd K HS-X-GTFTSDYSKYLD(Aib)RRAADFVAWLMDE (NR. ID. SEKW.")
226 22 HS-X-GTFTSDYSKYLDTRRAAEFVAWL(Nle)DE (NR. ID. SEKW.: 240) lub peptyd K HS-X-GTFTSDYSKYLD(Aib)RRAADFVAWLMDE (NR. ID. SEKW.: 241) z dodatkową modyfikacją w położeniu 3 otrzymano w wyniku syntezy peptydów w fazie stałej zasadniczo zgodnie z opisem w niniejszym dokumencie. Aktywność in vitro peptydów wobec receptora glukagonu badano zasadniczo zgodnie z opisem w przykładzie 16. Wartość EC0 (nm) każdego peptydu przedstawiono w tabeli 6.
![226 1 [0433] Jak pokazuje tabela 6, wiele aminokwasów może zastąpić Gln w położeniu 3 bez znaczącej utraty aktywności wobec receptora glukagonu, a w niektórych przypadkach modyfikacja doprowadziła w](/docs-images/65/53262204/images/227-0.jpg "istocie do zwiększenia aktywności, np. Dab(Ac) i Q(Me) w szkielecie peptydu K.")
227 226 1 [0433] Jak pokazuje tabela 6, wiele aminokwasów może zastąpić Gln w położeniu 3 bez znaczącej utraty aktywności wobec receptora glukagonu, a w niektórych przypadkach modyfikacja doprowadziła w istocie do zwiększenia aktywności, np. Dab(Ac) i Q(Me) w szkielecie peptydu K. PRZYKŁAD 40 [0434] Otrzymano peptydy stanowiące analogi glukagonu mające istotną aktywność wobec receptora glukagonu, a nie wobec receptora GIP i zawierające Dab(Ac) w położeniu 3 w różnych szkieletach analogów glukagonu zasadniczo zgodnie z opisem w niniejszym dokumencie oraz zbadano ich aktywność in vitro wobec receptora glukagonu. Strukturę i aktywności każdego peptydu przedstawiono w tabeli 7.
228 227 1 PRZYKŁAD 41 [043] Analogi glukagonu zawierające amid C-końcowy zamiast C-końcowego alfa-karboksylanu otrzymano zasadniczo zgodnie z opisem w niniejszym dokumencie. Peptydy mt-367, mt-368 i mt-369 zawierały odpowiednio struktury według NR. ID. SEKW.: 23, 236 i 237. Peptyd mt-384 zawierał aminokwas według NR. ID. SEKW.: 1, z następującymi modyfikacjami aminokwasów: Tyr w położeniu 1, AIB w położeniu 2, Lys w położeniu, przy czym Lys była połączona kowalencyjnie z tłuszczową grupą acylową C16 poprzez wstawkę dipeptydową γ-glu-γ- Glu, Ile w położeniu 12, Lys w położeniu 16, Gln w położeniu 17, Ala w położeniu 18, AIB w położeniu, Glu w położeniu 21, Asn w położeniu 24, Leu w położeniu 27, Ala w położeniu 28 i Gly w położeniu 29, a następnie aminokwas według NR. ID. SEKW.: 9 po stronie C-końcowej aminokwasu w położeniu 29. Peptyd mt-38 zawierał tę samą strukturę co peptyd mt-384 oprócz tego, że Thr w położeniu 7 zmieniono na Ile w mt-38. [0436] Aktywność in vitro analogów wobec każdego spośród receptorów glukagonu, GLP-1 i GIP badano zasadniczo zgodnie z opisem w niniejszym dokumencie. Wyniki podano w tabeli 8.
![228 1 [0437] Dziewięciu grupom po 8 myszy DIO (szczep: C7B16 WT) wstrzykiwano podskórnie raz na dobę przez 7 dni nmol/kg jednego z peptydów z tabeli 8.](/docs-images/65/53262204/images/229-0.jpg "Średnia początkowa masa ciała myszy wynosiła 7,6 g. Wiek myszy wynosił w przybliżeniu miesięcy i pozostawały one na diecie wysokotłuszczowej przez około 8 miesięcy.")
229 228 1 [0437] Dziewięciu grupom po 8 myszy DIO (szczep: C7B16 WT) wstrzykiwano podskórnie raz na dobę przez 7 dni nmol/kg jednego z peptydów z tabeli 8. Średnia początkowa masa ciała myszy wynosiła 7,6 g. Wiek myszy wynosił w przybliżeniu miesięcy i pozostawały one na diecie wysokotłuszczowej przez około 8 miesięcy. [0438] Całkowitą zmianę masy ciała mierzono w dniu 7. W przypadku wszystkich myszy, którym wstrzyknięto peptyd z tabeli 8, stwierdzono spadek masy ciała w porównaniu do kontroli (nośnika). U myszy, którym wstrzyknięto mt- 369, stwierdzono największy spadek masy ciała (spadek o ~2%), a następnie u myszy, którym wstrzyknięto mt-368 (spadek o ~22%), i myszy, którym wstrzyknięto mt-384 (spadek o ~21%). U myszy, którym wstrzyknięto mt-367 lub mt-38, stwierdzono mniejszy, ale nadal znaczący spadek masy ciała (mt-367: ~18% spadek i mt-38: ~1% spadek). [0439] Określenia w liczbie pojedynczej podobne określenia wykorzystywane w kontekście opisu wynalazku (w szczególności w kontekście poniższych zastrzeżeń) należy interpretować w taki sposób, że obejmują zarówno liczbę pojedynczą, jak i mnogą, chyba że w niniejszym
230 dokumencie podano inaczej lub kontekst stanowi wyraźnie inaczej. Określenia zawierający, mający, obejmujący i w tym należy interpretować jako określenia otwarte (tzn. oznaczają one w tym ale nieograniczająco ), chyba że podano inaczej. [0440] Jeśli w niniejszym dokumencie wymieniane są zakresy wartości, ich celem jest jedynie zastosowanie skróconego zapisu, zamiast podawania pojedynczo każdej odrębnej wartości znajdującej się w danym zakresie i w każdym punkcie końcowym, chyba że w niniejszym dokumencie podano inaczej, przy czym każda odrębna wartość i punkt końcowy zostają uwzględnione w opisie, tak jakby zostały indywidualnie wymienione w niniejszym dokumencie. [0441] Wszystkie sposoby opisywane w niniejszym dokumencie można zrealizować w dowolnej odpowiedniej kolejności, chyba że w niniejszym dokumencie podano inaczej lub kontekst stanowi wyraźnie inaczej. Zastosowanie wszelkich przykładów lub określeń przykładowych (np. taki jak ) w niniejszym dokumencie ma jedynie lepiej objaśnić wynalazek i nie stanowi ograniczenia zakresu wynalazku, chyba że zastrzeżono inaczej. Żadnego sformułowania w opisie nie należy interpretować w taki sposób, że wskazuje ono, iż którykolwiek element niezastrzegany jest niezbędny w praktyce wynalazku. [0442] W niniejszym dokumencie opisano korzystne przykłady wykonania niniejszego wynalazku, w tym najlepszy znany twórcom niniejszego wynalazku sposób realizacji wynalazku. Warianty tych korzystnych przykładów wykonania mogą być oczywiste dla
231 230 1 specjalistów w dziedzinie po zapoznaniu się z powyższym opisem. Twórcy niniejszego wynalazku oczekują, że specjaliści w dziedzinie będą stosować takie warianty w zależności od potrzeb, ponadto twórcy niniejszego wynalazku przewidują, że wynalazek może być realizowany w inny sposób niż opisany konkretnie w niniejszym dokumencie. W związku z tym niniejszy wynalazek obejmuje wszystkie modyfikacje i odpowiedniki przedmiotu, o którym mowa w zastrzeżeniach dołączonych do niniejszego dokumentu, w zakresie dopuszczalnym stosownymi przepisami prawa. Ponadto wynalazek obejmuje dowolną kombinację elementów, o których mowa powyżej, we wszystkich możliwych odmianach, chyba że w niniejszym dokumencie podano inaczej lub kontekst stanowi wyraźnie inaczej.
232 231 1 LISTA SEKWENCJI [0443] <1> Dimarchi i wsp. <1> POCHODNE GIP MAJĄCE MIESZANE DZIAŁANIE AGONISTYCZNE W LECZENIU ZABURZEŃ METABOLIZMU I OTYŁOŚCI <130> 3113/43978C <160> 262 <170> Patent In wersja 3. <2> 1 <211> 29 <213> Homo sapiens <223> Glukagon typu dzikiego <400> 1 2 <2> 2 <211> 31 <213> Homo sapiens <222> (1)..(31) <223> GLP-1(7-37) <400> 2
233 232 1 <2> 3 <211> 30 <222> (1)..(30) <223> GLP-1(7-36) amidowany <222> (30)..(30) <223> amidowanie na końcu C <400> 3 2 <2> 4 <211> 42 <213> Homo sapiens <221> mat_peptide <222> (1)..(42) <223> polipeptyd hamujący działający w żołądku <400> 4
234 233 1 <2> <211> 29 <223> Analog 61 <222> (16)..() <223> Mostek laktamowy między resztami 16 i. <400> 2 <2> 6 <211> 29 <223> Analog 62
235 234 <222> (16)..() <223> Mostek laktamowy między resztami 16 i. <400> 6 1 <2> 7 <211> 29 <223> Analog 63 <222> (12)..(16) <223> Mostek laktamowy między resztami 12 i 16. <400> 7 2 <2> 8 <211> 29
236 23 <223> Analog 66 <400> 8 1 <2> 9 <211> 29 <223> Analog 68 <400> 9 2 <2> <211> 29 <223> Analog 69
237 236 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 1 <2> 11 <211> 29 <223> Analog 84 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 11 2 <2> 12 <211> 29
238 237 <223> Analog 8 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (24)..(24) <223> PEG (kda) dołączony kowalencyjnie do cysteiny <400> 12 1 <2> 13 <211> 29 <223> Analog 92 <400> 13 2 <2> 14 <211> 29
239 238 <223> Analog 93 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (17)..(21) <223> Mostek laktamowy między resztami 17 i 21 <400> <2> 1 <211> 29 <223> Analog 9 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 1
 <223> Mostek laktamowy między resztami 16 i <400> 16 2 <2> 17")
240 239 1 <2> 16 <211> 29 <223> Analog 96 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 16 2 <2> 17 <211> 29 <223> Analog 97
241 240 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 17 1 <2> 18 <211> 29 <223> Analog 98 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 18 2 <2> 19 <211> 42
242 241 <223> Analog 99 <400> 19 1 <2> <211> 42 <223> Analog 0 <222> (2)..(2) <400> <2> 21 <211> 42
243 242 <223> Analog 1 <400> 21 1 <2> 22 <211> 42 <223> Analog 2 <222> (2)..(2) <400> 22 <2> 23 <211> 42
.")
244 243 <223> Analog 4 <400> 23 1 <2> 24 <211> 42 <223> Analog <222> (24)..(24) <223> PEG (40kDa) dołączony do cysteiny <400> 24
245 244 <2> 2 <211> 29 <223> Analog 6 <400> <2> 26 <211> 29 <223> Analog 7 <222> (16)..() <223> Mostek laktamowy między resztami 16 i
246 24 <400> 26 1 <2> 27 <211> 29 <223> Analog 8 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 27 2 <2> 28 <211> 29 <223> Analog 9
247 246 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 28 1 <2> 29 <211> 29 <223> Analog 1 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 29 2 <2> 30 <211> 29
248 247 <223> Analog 111 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> <2> 31 <211> 24 <223> Analog 113 <222> (1)..(1) <223> Xaa oznacza kwas 3-fenylomlekowy <222> (11)..(1) <223> Mostek laktamowy między resztami 11 i 1 <400> 31
249 248 1 <2> 32 <211> 24 <223> Analog 114 <222> (1)..(1) <223> Xaa oznacza kwas 3-fenylomlekowy <222> (11)..(1) <223> Mostek laktamowy między resztami 11 i 1 <400> 32 2 <2> 33 <211> 39 <223> Analog 11
250 249 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 33 1 <2> 34 <211> 39 <223> Analog 116 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 34 <2> 3 <211> 29
251 <223> Analog 118 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> <2> 36 <211> 39 <223> Analog 1 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 36
252 21 1 <2> 37 <211> 29 <223> Analog 124 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 37 2 <2> 38 <211> 29 <223> Analog 12
253 22 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 38 1 <2> 39 <211> 39 <223> Analog 127 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 39 2 <2> 40 <211> 39
 <223> Mostek laktamowy między resztami 16 i <400> 40 1 2 <2> 41 <211> 39 <223> Analog 129")
254 23 <223> Analog 128 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> <2> 41 <211> 39 <223> Analog 129 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 41
255 24 1 <2> 42 <211> 39 <223> Analog 139 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 42 <2> 43 <211> 39
256 2 <223> Analog 140 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 43 1 <2> 44 <211> 39 <223> Analog 141 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 44
257 26 1 <2> 4 <211> 39 <223> Analog 142 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 4 <2> 46 <211> 39
258 27 <223> Analog 143 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 46 1 <2> 47 <211> 39 <223> Analog 144 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 47
259 28 1 <2> 48 <211> 39 <223> Analog 14 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 48 <2> 49 <211> 39
260 29 <223> Analog 146 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 49 1 <2> 0 <211> 39 <223> Analog 147 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 0
261 260 1 <2> 1 <211> 39 <223> Analog 148 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 1 <2> 2 <211> 39
262 261 <223> Analog 149 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 2 1 <2> 3 <211> 39 <223> Analog 10 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 3
263 262 1 <2> 4 <211> 39 <223> Analog 11 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 4 <2> <211> 39
264 263 <223> Analog 12 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 1 2 <2> 6 <211> 39 <223> Analog 14 <222> (1)..(1) <223> Xaa oznacza kwas hipurowy
265 264 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 6 1 <2> 7 <211> 39 <223> Analog 1 <222> (12)..(16) <223> Mostek laktamowy między resztami 12 i 16 <400> 7 <2> 8 <211> 39
266 26 <223> Analog 16 <222> ().. (24) <223> Mostek laktamowy między resztami i 24 <400> <2> 9 <211> 39 <223> Analog 17 <222> (24).. (28) <223> Mostek laktamowy między resztami 24 i 28 <400> 9
267 266 1 <2> 60 <211> 39 <223> Analog 18 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 60 <2> 61 <211> 39
268 267 1 <223> Analog 162 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 61 2 <2> 62 <211> 29 <223> Analog 163
269 268 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 62 1 <2> 63 <211> 29 <223> Analog 164 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 63 2 <2> 64 <211> 40
270 269 <223> Analog 16 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> <2> 6 <211> 40 <223> Analog 166 <222> (2).. (2)
271 270 <223> Xaa oznacza d-alaninę <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 6 1 <2> 66 <211> 39 <223> Analog 167 <222> (17)..() <223> Mostek laktamowy między resztami 17 i <400> 66 <2> 67
272 271 <211> 39 <223> Analog 168 <222> (18)..(21) <223> Mostek laktamowy między resztami 18 i 21 <400> <2> 68 <211> 29 <223> Analog 169 <222> (2).. (2)
273 272 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> <2> 69 <211> 40 <223> Analog 170 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> 69
274 273 1 <2> 70 <211> 29 <223> Analog 172 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (24).. (24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> 70 2 <2> 71
275 274 1 <211> 40 <223> Analog 174 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 71 2 <2> 72 <211> 39 <223> Analog 17
 <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> 72 1 2 <2> 73 <211> 40 <223>")
276 27 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (24).. (24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> <2> 73 <211> 40 <223> Analog 176
277 276 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> <2> 74 <211> 29 <223> Analog 177 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 74
278 277 1 <2> 7 <211> 40 <223> Analog 178 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> 7 2 <2> 76
279 278 1 <211> 29 <223> Analog 179 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (24).. (24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> 76 2 <2> 77 <211> 40
280 279 1 <223> Analog 182 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> 77 2 <2> 78 <211> 29 <223> Analog 186
281 280 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (24).. (24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> <2> 79 <211> 39 <223> Analog 191 <222> (2).. (2) <400> 79

282 281 <2> 80 <211> 34 <223> Analog 192 <400> 80 1 <2> 81 <211> 38 <223> Analog 194
283 282 <222> (1)..(1) <400> 81 1 <2> 82 <211> 26 <223> Analog 197 <222> (13)..(17) <223> Mostek laktamowy między resztami 13 i 17 <400> 82 <2> 83 <211> 28
284 283 <223> Analog 198 <222> (1)..(1) <222> (1)..(19) <223> Mostek laktamowy między resztami 1 i 19 <400> <2> 84 <211> 40 <223> Analog 199 <222> (2).. (2)
285 284 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> <2> 8 <211> 40 <223> Analog 0 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (27).. (27) <223> Xaa oznacza Nle <400> 8
286 28 1 <2> 86 <211> 39 <223> Analog 1 <222> (2).. (2) <400> 86 <2> 87 <211> 40
287 286 1 <223> Analog 2 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> 87 2 <2> 88 <211> 40 <223> Analog 3
288 287 1 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (27).. (27) <223> Xaa oznacza Nle <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> 88 2 <2> 89 <211> 40 <223> Analog 4
289 288 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> <2> 90 <211> 40 <223> Analog <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i
290 289 <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> 90 1 <2> 91 <211> 43 <223> Analog 6 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 91
291 290 1 <2> 92 <211> 43 <223> Analog 7 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 92 2 <2> 93 <211> 43
292 291 <223> Analog 8 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (43)..(43) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> <2> 94 <211> 40 <223> Analog 9 <222> (2).. (2)
293 292 <222> (9)..(12) <223> Mostek laktamowy między resztami 9 i 12 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <400> 94 1 <2> 9 <211> <223> CEX <400> 9 2 <2> 96 <211> 11
294 293 <222> (1)..(1) <223> Xaa oznacza dowolny aminokwas <400> 96 <2> 97 <211> 8 <400> 97 1 <2> 98 <211> 4 <400> <2> 99 <211> 40
295 294 1 <223> mt-274 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Amidowanie na końcu C <400> 99 2 <2> 0 <211> 40
296 29 1 <223> mt-311 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Amidowanie na końcu C <400> 0 2 <2> 1 <211> 40
297 296 1 <223> mt-298 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> 1 2 <2> 2 <211> 40
298 <223> mt-309 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> 2
299 <2> 3 <211> 40 <223> mt-3 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 18 atomów węgla dołączona kowalencyjnie do lizyny
300 299 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 4 <211> 40 <223> mt-260 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów
301 300 węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> <211> 40 <223> mt-261 <222> (2).. (2) <222> ().. () <222> (40)..(40)
302 301 <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> 1 2 <2> 6 <211> 40 <223> mt-262 <222> (2).. (2) <222> ().. ()
303 302 <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 18 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 7 <211> 40 <223> mt-22 <222> (2).. (2) <222> ()..()
304 303 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 8 <211> 40 <223> mt-2 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> PEG (40kDa) dołączony do cysteiny
305 304 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 9 <211> 40 <223> mt-26 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Amidowanie na końcu C <400> 9
306 30 1 <2> 1 <211> 29 <223> mt-27 <222> (2).. (2) <222> ().. () <222> (29).. (29) <223> Amidowanie na końcu C <400> 1 2 <2> 111
307 306 1 <211> 40 <223> mt-263 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 112 <211> 29
308 307 <223> mt-264 <222> (2).. (2) <222> ().. () <400> <2> 113 <211> 39 <223> mt-26 <222> (2).. (2)
309 308 <222> ()..() <222> (24).. (24) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (39).. (39) <223> Amidowanie na końcu C <400> <2> 114 <211> 39 <223> mt-266 <222> (2).. (2)
310 309 <222> ()..() <222> (24).. (24) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (39).. (39) <223> Amidowanie na końcu C <400> <2> 11 <211> 39 <223> mt-267 <222> (2).. (2)
311 3 <222> ().. () <222> (24).. (24) <223> Tłuszczowa grupa acylowa o długości 18 atomów węgla dołączona kowalencyjnie do lizyny <222> (39).. (39) <223> Amidowanie na końcu C <400> <2> 116 <211> 39 <223> mt-268 <222> (2).. (2)
312 311 <222> ().. () <222> (39).. (39) <223> Amidowanie na końcu C <400> <2> 117 <211> 39 <223> mt-269 <222> (2).. (2) <222> ()..()
313 312 <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> ().. () <222> (39)..(39) <223> Amidowanie na końcu C <400> <2> 118 <211> 39 <223> mt-270 <222> (2).. (2)
314 313 <222> ()..() <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> ().. () <222> (39).. (39) <223> Amidowanie na końcu C <400> <2> 119 <211> 39 <223> mt-271 <222> (2).. (2)
315 314 <222> ()..() <223> Tłuszczowa grupa acylowa o długości 18 atomów węgla dołączona kowalencyjnie do lizyny <222> ().. () <222> (39).. (39) <223> Amidowanie na końcu C <400> <2> 1 <211> 39 <223> mt-272 <222> (2).. (2)
316 31 <222> ().. () <222> (39).. (39) <223> Amidowanie na końcu C <400> <2> 121 <211> 40 <223> mt-27 <222> (2)..(2) <222> (16).. (16) <223> Xaa oznacza Orn
317 316 <222> ().. () <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 122 <211> 40 <223> mt-276 <222> (2).. (2) <222> (16)..(16) <223> Xaa oznacza kwas 2,4-diaminomasłowy
318 317 <222> ().. () <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 123 <211> 40 <223> mt-277 <222> (2).. (2) <222> ()..()
319 318 <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 124 <211> 40 <223> mt-278 <222> (2).. (2) <222> ().. ()
320 319 <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 12 <211> 40 <223> mt-279 <222> (2).. (2)
321 3 <222> ().. () <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 18 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 126 <211> 40 <223> mt-280 <222> (2).. (2)
322 321 <222> ().. () <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 127 <211> 29 <223> mt-281 <222> (2).. (2) <222> ().. ()
323 322 <222> (24).. (24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (29).. (29) <223> Amidowanie na końcu C <400> <2> 128 <211> 29 <223> mt-282 <222> (2)..(2) <222> ()..() <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny
324 323 <222> ()..() <222> (29)..(29) <223> Amidowanie na końcu C <400> <2> 129 <211> 29 <223> mt-283 <222> (2)..(2) <222> ()..() <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> ()..()
325 324 <222> (29)..(29) <223> Amidowanie na końcu C <400> <2> 130 <211> 29 <223> mt-284 <222> (2)..(2) <222> ()..() <223> Tłuszczowa grupa acylowa o długości 18 atomów węgla dołączona kowalencyjnie do lizyny <222> ()..()
326 32 <222> (29)..(29) <223> Amidowanie na końcu C <400> <2> 131 <211> 29 <223> mt-28 <222> (2)..(2) <222> ()..() <222> (29)..(29) <223> Amidowanie na końcu C <400> 131
327 <2> 132 <211> 40 <223> mt-286 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> 132
328 <2> 133 <211> 40 <223> mt-287 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C
329 328 <400> <2> 134 <211> 40 <223> mt-288 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 18 atomów węgla dołączona kowalencyjnie do lizyny
330 329 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 13 <211> 40 <223> mt-289 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Amidowanie na końcu C <400> 13
331 <2> 136 <211> 40 <223> mt-290 <222> (2)..(2) <222> ()..() <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny
332 331 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 137 <211> 40 <223> mt-291 <222> (2)..(2) <222> ()..() <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny
 <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40).")
333 332 <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 138 <211> 40 <223> mt-292 <222> (2)..(2)
334 333 1 <222> ()..() <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 18 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 139 <211> 40 <223> mt-293
335 334 1 <222> (2)..(2) <222> ()..() <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 140 <211> 40 <223> mt-29
336 33 1 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 141 <211> 40 <223> mt-296
 <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40).")
337 336 1 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 142 <211> 40 <223> mt-297
338 337 1 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 143 <211> 40
339 338 1 <223> mt-299 <222> (2)..(2) <222> ()..() <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 144 <211> 40
340 339 1 <223> mt-306 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 14 <211> 40
341 <223> mt-307 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 18 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> 14 <2> 146
342 341 1 <211> 40 <223> mt-308 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 147 <211> 40
343 342 1 <223> mt-322 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Grupa acylowa 3-SH-propionowa dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 148 <211> 40
344 343 1 <223> mt-323 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 8 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 149 <211> 40
345 344 1 <223> mt-324 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 10 <211> 40
346 34 1 <223> mt-32 <222> (2)..(2) <222> (7)..(7) <223> Xaa oznacza kwas 2-aminomasłowy <222> ()..() <222> (40)..(40) <223> Amidowanie na końcu C <400> 10 2 <2> 11 <211> 40
347 346 1 <223> mt-329 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 8 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> 11 2 <2> 12 <211> 40
348 347 1 <223> mt-330 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 8 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> 12 2 <2> 13 <211> 40
349 348 1 <223> mt-331 <222> (2)..(2) <222> ()..() <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Amidowanie na końcu C <400> 13 2 <2> 14 <211> 40
350 <223> mt-333 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Xaa oznacza d-lizynę <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do d-lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> 14
351 <2> 1 <211> 40 <223> mt-334 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Xaa oznacza d-lizynę <222> (40)..(40) <223> Amidowanie na końcu C <400> 1
352 <2> 16 <211> 40 <223> mt-33 <222> (2)..(2) <222> ()..() <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Xaa oznacza d-lizynę <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny
353 32 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 17 <211> 40 <223> mt-336 <222> (2)..(2) <222> ()..() <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny
354 33 <222> (40)..(40) <223> Xaa oznacza d-lizynę <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 18 <211> 39 <223> mt-337 <222> (2)..(2) <222> ()..() <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny
355 34 <222> ()..() <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 19 <211> 39 <223> mt-338 <222> (2)..(2) <222> ().. ()
356 3 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 160 <211> 41 <223> mt-339 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów
357 36 węgla dołączona kowalencyjnie do lizyny <222> (41)..(41) <223> Amidowanie na końcu C <400> <2> 161 <211> 41 <223> mt-340 <222> (2)..(2) <222> ()..() <222> (41)..(41)
358 37 <223> Amidowanie na końcu C <400> <2> 162 <211> 39 <223> mt-341 <222> (2)..(2) <222> ()..() <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> ()..()
359 38 <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (39)..(39) <223> Amidowanie na końcu C <400> <2> 163 <211> 41 <223> mt-343 <222> (2)..(2) <222> ()..()
360 39 <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (41)..(41) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (41)..(41) <223> Amidowanie na końcu C <400> <2> 164 <211> 41 <223> mt-344 <222> (2)..(2)
361 360 <222> ().. () <222> (41)..(41) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (41)..(41) <223> Amidowanie na końcu C <400> <2> 16 <211> 40 <223> mt-34 <222> (2)..(2)
362 361 <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 166 <211> 40 <223> mt-33 <222> (2)..(2) <222> ()..()
363 362 1 <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 167 <211> 40 <223> mt-241
364 363 <222> (2)..(2) <222> (16)..(16) <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 168 <211> 40 <223> mt-242 <222> (2)..(2)
365 364 <222> (16)..(16) <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 169 <211> 40 <223> mt-273 <222> (2)..(2) <222> (12)..(16) <223> Mostek laktamowy między resztami 12 i 16
366 36 <222> ().. () <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 170 <211> <400> <2> 171 <211> 11 <222> (1)..(1) <223> Xaa oznacza Gly lub mały aminokwas alifatyczny lub niepolarny lub nieco polarny
367 366 <400> 171 <2> 172 <211> 12 <222> (1)..(1) <223> Xaa oznacza Gly lub mały aminokwas alifatyczny lub niepolarny lub nieco polarny <400> <2> 173 <211> 40 <223> mt-248 <222> (2).. (2) <222> (16)..(16)
368 367 <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 174 <211> 40 <223> mt-249 <222> (2)..(2) <222> (16)..(16)
 <223> Amidowanie na końcu C <400> 174 1 2 <2> 17 <211> 40 <223> mt- <222> (2).")
369 368 <222> (40)..(40) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 17 <211> 40 <223> mt- <222> (2)..(2)
 <223> Amidowanie na końcu C <400> 17 1 2 <2> 176 <211> 40")
370 369 <222> (16)..(16) <222> ()..() <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 176 <211> 40 <223> mt-21 <222> (2)..(2)
371 370 <222> ()..() <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 177 <211> 29 <223> mt-28 <222> (2)..(2) <222> ()..()
372 371 <222> (29)..(29) <223> Amidowanie na końcu C <400> <2> 178 <211> 29 <223> mt-29 <223> Amidowanie na końcu C <222> (2)..(2) <222> ()..() <400> 178
373 372 1 <2> 179 <211> 30 <222> (2)..(2) <222> (24)..(24) <223> PEG (40kDa) dołączony kowalencyjnie do cysteiny <400> <2> 180 <211> 42 <223> NR. ID. SEKW.: 24
374 373 <222> (24)..(24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> <2> 181 <211> 40 <223> NR. ID. SEKW.: 69 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z
375 374 grupą hydrofilową <400> <2> 182 <211> 29 <223> NR. ID. SEKW.: 70 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (24)..(24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <221> misc_feature
376 37 <222> (29)..(29) <223> Xaa może stanowić dowolny aminokwas występujący naturalnie <400> <2> 183 <211> 39 <223> NR. ID. SEKW.: 72 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (24)..(24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> 183
377 376 1 <2> 184 <211> 40 <223> NR. ID. SEKW.: 7 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> 184
378 377 1 <2> 18 <211> 29 <223> NR. ID. SEKW.: 76 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (24)..(24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> 18 2
379 378 1 <2> 186 <211> 40 <223> NR. ID. SEKW.: 77 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> <2> 187 <211> 29
380 379 1 <223> NR. ID. SEKW.: 78 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> <2> 188 <211> 40
381 380 1 <223> NR. ID. SEKW.: 87 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> <2> 189 <211> 40 <223> NR. ID. SEKW.: 88
382 381 1 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (27)..(27) <223> Xaa oznacza Nle <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> <2> 190 <211> 40 <223> NR. ID. SEKW.: 90
383 382 <222> (2)..(2) <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> <2> 191 <211> 43 <223> NR. ID. SEKW.: 93 <222> (2)..(2)
384 383 <222> (16)..() <223> Mostek laktamowy między resztami 16 i <222> (43)..(43) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> <2> 192 <211> 40 <223> NR. ID. SEKW.: 99 <222> (2)..(2) <222> ()..()
385 384 <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 193 <211> 40 <223> NR. ID. SEKW.: 0 <222> (2)..(2)
386 38 <222> ()..() <222> (24)..(24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 194 <211> 40 <223> NR. ID. SEKW.: 1, 2, 3 <222> (2)..(2)
387 386 1 <222> ()..() <222> (24)..(24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 19 <211> 40
388 387 1 <223> NR. ID. SEKW.: 4,, 6 <222> (2)..(2) <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> 19 2 <2> 196 <211> 40
389 388 1 <223> NR. ID. SEKW.: 8 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 197 <211> 39
390 389 1 <223> NR. ID. SEKW.: 113, 114, 11 <222> (2)..(2) <222> ()..() <222> (24)..(24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (39).. (39) <223> Amidowanie na końcu C <400> <2> 198 <211> 39
391 390 1 <223> NR. ID. SEKW.: 117, 118, 119 <222> (2)..(2) <222> ()..() <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> ()..() <222> (39)..(39) <223> Amidowanie na końcu C <400> <2> 199 <211> 40
392 391 1 <223> NR. ID. SEKW.: 123, 124, 12 <222> (2)..(2) <222> ()..() <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 0 <211> 29
393 392 1 <223> NR. ID. SEKW.: 127 <222> (2)..(2) <222> ()..() <222> (24)..(24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (29)..(29) <223> Amidowanie na końcu C <400> 0 2 <2> 1 <211> 29
394 393 1 <223> NR. ID. SEKW.: 128, 129, 130 <222> (2).. (2) <222> ()..() <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> ().. () <222> (29).. (29) <223> Amidowanie na końcu C <400> 1 2 <2> 2 <211> 40
395 394 1 <223> NR. ID. SEKW.: 132 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> 2 2 <2> 3 <211> 40
396 <223> NR. ID. SEKW.: 136, 137, 138 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> 3
397 <2> 4 <211> 40 <223> NR. ID. SEKW.: 139 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Amidowanie na końcu C
398 397 <400> <2> <211> 40 <223> NR. ID. SEKW.: 140 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40)
399 398 <223> Amidowanie na końcu C <400> 1 2 <2> 6 <211> 40 <223> NR. ID. SEKW.: 141 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową
400 399 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 7 <211> 40 <223> NR. ID. SEKW.: 142 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową
401 400 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 8 <211> 40 <223> NR. ID. SEKW.: 143 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z
402 401 grupą hydrofilową <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 9 <211> 40 <223> NR. ID. SEKW.: 144, 14 <222> (2).. (2)
403 402 <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 2 <211> 40 <223> NR. ID. SEKW.: 148 <222> (2).. (2)
404 403 <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 211 <211> 40 <223> NR. ID. SEKW.: 12 <222> (2).. (2)
405 404 <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 212 <211> 40 <223> NR. ID. SEKW.: 12 <222> (2).. (2)
406 40 1 <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 213 <211> 40 <223> NR. ID. SEKW.:: 13
407 406 1 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 214 <211> 40 <223> NR. ID. SEKW.: 14
408 407 1 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 21 <211> 40 <223> NR. ID. SEKW.: 16
409 408 1 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> 21 2 <2> 216 <211> 40
410 409 1 <223> NR. ID. SEKW.: 17 <222> (2).. (2) <222> ().. () <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 217 <211> 39
411 4 1 2 <223> NR. ID. SEKW.: 23 <222> (2).. (2) <222> ()..() <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> ().. () <222> (40)..(40) <223> Amidowanie na końcu C <400> 217 <2> 218
412 <211> 41 <223> NR. ID. SEKW.: 160 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (41)..(41) <223> Amidowanie na końcu C <400> 218
413 <2> 219 <211> 39 <223> NR. ID. SEKW.: 162 <222> (2).. (2) <222> ()..() <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> ().. () <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (39).. (39) <223> Amidowanie na końcu C
414 413 <400> <2> 2 <211> 41 <223> NR. ID. SEKW.: 163 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (41)..(41)
 <223> Amidowanie na końcu C <400> 2 1 2 <2> 221 <211> 41 <223> NR. ID. SEKW.: 164 <222> (2).")
415 414 <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (41)..(41) <223> Amidowanie na końcu C <400> <2> 221 <211> 41 <223> NR. ID. SEKW.: 164 <222> (2).. (2) <222> ().. ()
416 41 <222> (41)..(41) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (41)..(41) <223> Amidowanie na końcu C <400> <2> 222 <211> 40 <223> NR. ID. SEKW.: 16 <222> (2).. (2) <222> (40)..(40)
417 416 <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 223 <211> 40 <223> NR. ID. SEKW.: 166 <222> (2).. (2) <222> ().. ()
418 417 <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą acylową lub alkilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 224 <211> 40 <223> NR. ID. SEKW.:: 173 <222> (2).. (2)
419 418 <222> (16).. (16) <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 22 <211> 40 <223> NR. ID. SEKW.: 174 <222> (2).. (2)
420 419 1 <222> (16).. (16) <222> (40)..(40) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <222> (40)..(40) <223> Amidowanie na końcu C <400> 22 2 <2> 226 <211> 30 <222> (2).. (2)
421 4 <222> (24).. (24) <223> Podstawiony aminokwas połączony kowalencyjnie z grupą hydrofilową <400> <2> 227 <211> 29 <222> (2).. (2) <222> ().. () <222> (29).. (29) <223> Amidowanie na końcu C <400> 227 <2> 228
422 <211> 29 <222> (2).. (2) <223> Xaa oznacza D-Ser, D-Ala, Val, Gly, N-metylo Ser, N-metylo Ala, lub Aib <222> (16).. (16) <223> Xaa oznacza Lys, Glu, Ser, Orn, Dab, Aib lub dowolny aminokwas alfa,alfa-dipodstawiony <222> ().. () <223> Xaa oznacza dowolny aminokwas alfa,alfadipodstawiony <222> (27).. (27) <223> Xaa oznacza Ile, Leu, lub Nle <222> (28).. (28) <223> Xaa oznacza Ala lub Gly <222> (29).. (29)
423 422 <223> Xaa oznacza Thr, Ala lub Gly <222> (29).. (29) <223> Amidowanie na końcu C <400> <2> 229 <211> 29 <222> (2).. (2) <222> ().. () <222> (29).. (29) <223> Amidowanie na końcu C <400> 229
424 <2> 230 <211> 29 <222> (2).. (2) <223> Xaa oznacza D-Ser, D-Ala, Val, Gly, N-metylo Ser, N-metylo Ala lub Aib <222> (16).. (16) <223> Xaa oznacza Lys, Glu, Ser, Orn, Dab, Aib lub dowolny aminokwas alfa,alfa-dipodstawiony <222> ().. () <223> Xaa oznacza dowolny aminokwas alfa,alfadipodstawiony <222> (27).. (27) <223> Xaa oznacza Ile, Leu lub Nle <222> (28).. (28) <223> Xaa oznacza Ala lub Gly
425 424 <222> (29).. (29) <223> Xaa oznacza Thr, Ala lub Gly <222> (29).. (29) <223> Amidowanie na końcu C <400> <2> 231 <211> 41 <223> Mt-34 <223> Homodimer <222> (2).. (2) <222> ().. ()
426 42 <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <221> DISIARCZEK <222> (41)..(41) <223> Wiązanie disiarczkowe utworzone z resztą Cys drugiego peptydu mającego tę samą strukturę. <400> <2> 232 <211> 40 <223> Mt-36 <222> (2).. (2)
427 426 <222> ().. () <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 233 <211> 40 <223> Mt-37 <222> (2).. (2) <222> ().. ()
428 427 <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 234 <211> 40 <223> Mt-38 <222> (2).. (2) <222> ().. () <222> (40)..(40)
429 428 <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 23 <211> 40 <223> Mt-367 <222> (2).. (2) <222> ().. ()
430 429 <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny poprzez wstawkę gamma-glu-gamma-glu <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 236 <211> 40 <223> Mt-368 <222> (2).. (2) <222> ().. ()
431 430 <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny poprzez wstawkę gamma-glu-gamma-glu <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 237 <211> 40 <223> Mt-369 <222> (2).. (2)
432 431 <222> ()..() <223> Tłuszczowa grupa acylowa o długości 16 atomów węgla dołączona kowalencyjnie do lizyny poprzez wstawkę gamma-glu-gamma-glu <222> ().. () <222> (40)..(40) <223> Amidowanie na końcu C <400> <2> 238 <211> 41 <223> Mt-376 <223> Homodimer
433 432 1 <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (41)..(41) <223> Amidowanie na końcu C <222> (41)..(41) <223> Reszta Cys dołączona kowalencyjnie do PEG o masie kda, który z kolei jest dołączony kowalencyjnie do reszty Cys drugiego peptydu mającego tę samą strukturę <400> <2> 239 <211> 41
434 <223> Mt-377 <223> homodimer <222> (2).. (2) <222> ().. () <222> (40)..(40) <223> Tłuszczowa grupa acylowa o długości 14 atomów węgla dołączona kowalencyjnie do lizyny <222> (41)..(41) <223> PEG (kda) połączony kowalencyjnie z cysteiną <222> (41)..(41) <223> Amidowanie na końcu C
435 434 <222> (41)..(41) <223> Reszta Cys dołączona kowalencyjnie do PEG o masie kda, który z kolei jest dołączony kowalencyjnie do reszty Cys drugiego peptydu mającego tę samą strukturę <400> <2> 240 <211> 29 <223> Peptyd J <222> (3)..(3) <223> Xaa oznacza analog glutaminy <222> (27).. (27) <223> Xaa oznacza Nle <400> 240
436 43 1 <2> 241 <211> 29 <223> Peptyd K <222> (3)..(3) <223> Xaa oznacza analog glutaminy <222> (16).. (16) <400> <2> 242 <211> 29
 <223> Xaa oznacza acetamidometylocysteinę <222> (27).")
437 436 <222> (27).. (27) <223> Xaa oznacza Nle <400> <2> 243 <211> 29 <223> Peptyd syntetyczny <222> (3)..(3) <223> Xaa oznacza acetamidometylocysteinę <222> (27).. (27) <223> Xaa oznacza Nle <400> <2> 244 <211> 29 <223> Peptyd syntetyczny
438 437 <222> (3)..(3) <223> Xaa oznacza kwas acetylodiaminobutanowy <222> (27).. (27) <223> Xaa oznacza Nle <400> <2> 24 <211> 29 <222> (3)..(3) <223> Xaa oznacza kwas karbamoilodiaminopropanowy <222> (27).. (27) <223> Xaa oznacza Nle <400> 24 <2> 246
439 438 <211> 29 <222> (3)..(3) <223> Xaa oznacza metyloglutaminę <222> (27).. (27) <223> Xaa oznacza Nle <400> <2> 247 <211> 29 <223> Peptyd syntetyczny <222> (3)..(3) <223> Xaa oznacza sulfotlenek metioniny <222> (27).. (27) <223> Xaa oznacza Nle
440 439 <400> <2> 248 <211> 29 <223> Peptyd syntetyczny <222> (3)..(3) <223> Xaa oznacza acetyloornitynę <222> (27).. (27) <223> Xaa oznacza Nle <400> <2> 249 <211> 29 <222> (16).. (16)
441 440 <400> <2> <211> 29 <222> (3)..(3) <223> Xaa oznacza kwas acetylodiaminobutanowy <222> (16).. (16) <400> 2 <2> 21 <211> 29
442 441 <222> (3)..(3) <223> Xaa oznacza metyloglutaminę <222> (16).. (16) <400> 21 1 <2> 22 <211> 29 <400> 22 2 <2> 23 <211> 29 <222> (3)..(3) <223> Xaa oznacza kwas acetylodiaminobutanowy
443 442 <222> (16).. (16) <400> 23 1 <2> 24 <211> 39 <222> (3)..(3) <223> Xaa oznacza kwas acetylodiaminobutanowy <222> (16).. (16) <222> (39).. (39) <223> Amidowanie na końcu C <400> 24 2
444 443 1 <2> 2 <211> 29 <222> (3)..(3) <223> Xaa oznacza kwas acetylodiaminobutanowy <222> (16).. (16) <400> 2 2 <2> 26 <211> 29 <222> (3)..(3) <223> Xaa oznacza kwas acetylodiaminobutanowy <222> (16).. (16)
445 444 <400> <2> 27 <400> <2> 28 <400> <2> 29 <211> 40 <223> Peptyd syntetyczny <223> mt-22 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy łączy Glu16 i Lys <222> (40)..(40)
446 44 <223> Grupa epsilon-aminowa Lys jest dołączona do Ala- Ac-Cys(PEG), przy czym Ac-Cys(PEG) oznacza resztę Cys zawierającą grupę alfa-aminową z dołączoną grupą acetylową (CH3CO) i zawierającą PEG o masie 40 kda dołączony kowalencyjnie do jej łańcucha bocznego <400> <2> 260 <211> 40 <223> Peptyd syntetyczny <223> mt-227 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy łączy Glu16 i Lys <222> (40)..(40)
447 446 <223> Grupa epsilon-aminowa Lys jest dołączona do Ala- Ac-Cys(PEG), przy czym Ac-Cys(PEG) oznacza resztę Cys zawierającą grupę alfa-aminową z dołączoną grupą acetylową (CH3CO) i zawierającą PEG o masie 40 kda dołączony kowalencyjnie do jej łańcucha bocznego <400> <2> 261 <211> 40 <223> Peptyd syntetyczny <223> mt-294 <222> (2).. (2) <222> (16)..() <223> Mostek laktamowy łączy Glu16 i Lys <222> (40)..(40)
.")
448 447 <223> Cys-PEG połączony kowalencyjnie przez wiązanie tioeterowe <400> <2> 262 <211> 40 <223> Peptyd syntetyczny <222> (2).. (2) <400> 262 Indiana University Research and Technology Corporation Pełnomocnik:
449 448 Zastrzeżenia patentowe 1 1. Analog glukagonu (NR. ID. SEKW.: 1) mający działanie agonistyczne wobec GIP z następującymi modyfikacjami: (a) modyfikacja aminokwasu w położeniu 1 nadająca aktywność agonistyczną wobec GIP, przy czym ewentualnie aminokwasem w położeniu 1 jest aminokwas niezawierający imidazolowego łańcucha bocznego; (b) jeden, dwa, trzy lub wszystkie spośród aminokwasów w położeniach 16,, 21 i 24 analogu jest podstawionych przez aminokwas α,α-dipodstawiony, (c) modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29 oraz (d) 1-9 dodatkowych modyfikacji aminokwasów względem sekwencji glukagonu (NR. ID. SEKW.: 1), przy czym wartość EC0 analogu wobec aktywacji receptora GIP wynosi około nm lub mniej, i przy czym wartość EC0 analogu wobec receptora GIP różni się mniej niż około 0-krotnie względem jego wartości EC0 wobec receptora GLP-1, przy czym ewentualnie siła działania GIP analogu pozostaje w granicach około 1- krotności siły działania GLP-1 tego analogu Analog glukagonu (NR. ID. SEKW.: 1) mający działanie agonistyczne wobec GIP z następującymi modyfikacjami: (a) modyfikacja aminokwasu w położeniu 1 nadająca aktywność agonistyczną wobec GIP, przy czym ewentualnie aminokwasem w położeniu 1 jest aminokwas niezawierający imidazolowego łańcucha bocznego,
450 449 (b) substytucja aminokwasu Ser w położeniu 16 przez aminokwas o wzorze IV: [wzór IV], w którym n wynosi 1 do 7, przy czym każdy spośród R1 i R2 jest niezależnie wybrany z grupy składającej się z następujących: H, C1-C18 alkil, (C1-C18 alkilo)oh, (C1-C18 alkilo)nh2, (C1-C18 alkilo)sh, (C0-C4 alkilo)(c3- C6)cykloalkil, (C0-C4 alkilo)(c2-c heterocykl), (C0-C4 1 2 alkilo)(c6-c arylo)r7 i (C1-C4 alkilo)(c3-c9 heteroaryl), przy czym R7 oznacza H lub OH, a łańcuch boczny aminokwasu o wzorze IV zawiera wolną grupę aminową, aminokwas o wzorze IV stanowi ewentualnie homolys, Lys, Orn lub kwas 2,4-diaminomasłowy (Dab), (c) substytucja aminokwasu Gln w położeniu aminokwasem alfa,alfa-dipodstawionym, ewentualnie AIB, (d) modyfikacje aminokwasów w jednym, dwóch lub wszystkich spośród położeń 27, 28 i 29 oraz (e) 1-9 dodatkowych modyfikacji aminokwasów względem sekwencji glukagonu (NR. ID. SEKW.: 1), przy czym wartość EC0 analogu wobec aktywacji receptora GIP wynosi około nm lub mniej, i przy czym wartość EC0 analogu wobec receptora GIP różni się mniej niż około 0-krotnie względem jego wartości EC0 wobec receptora GLP-1, przy czym ewentualnie siła działania GIP analogu pozostaje w granicach około 1-
451 40 krotności siły działania GLP-1 tego analogu. 3. Analog według zastrzeżenia 1 albo 2, w którym (a) aminokwas w położeniu 1 stanowi duży aminokwas aromatyczny, ewentualnie Tyr, i (b) przy czym (i) Met w położeniu 27 jest podstawiona przez duży aminokwas alifatyczny, ewentualnie Leu, (ii) Asn w położeniu 28 jest podstawiona przez mały aminokwas alifatyczny, ewentualnie Ala, lub (iii) Thr w położeniu 29 jest podstawiona przez mały aminokwas alifatyczny, ewentualnie Gly, lub przy czym analog zawiera kombinację (i), (ii) i (iii) Analog według któregokolwiek z zastrzeżeń 1-3, zawierający ponadto sekwencję aminokwasową GPSSGAPPPS (NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.: 96) po stronie C-końcowej aminokwasu w położeniu Analog według któregokolwiek z zastrzeżeń 1-4, zawierający jedną lub więcej z następujących modyfikacji: (a) Ser w położeniu 2 podstawiona przez D-Ser, Ala, D-Ala, Gly, N-metylo-Ser, AIB, Val lub kwas α- amino-n-masłowy; (b) Gln w położeniu 3 podstawiona przez Glu; (c) substytucja aminokwasu Tyr w położeniu przez aminokwas, ewentualnie aminokwas o wzorze I:
452 41 1 w którym n = 1 do 4, zawierający łańcuch boczny połączony kowalencyjnie z grupą acylową lub grupą alkilową; (d) addycja aminokwasu, ewentualnie aminokwasu o wzorze I, zawierającego łańcuch boczny połączony kowalencyjnie z grupą acylową lub grupą alkilową, jako aminokwasu C-końcowego w analogu; (e) Lys w położeniu 12 podstawiona przez Ile; (f) Arg w położeniu 17 podstawiona przez Gln; (g) Arg w położeniu 18 podstawiona przez Ala; (h) Asp w położeniu 21 podstawiona przez Glu; (i) Gln w położeniu 24 podstawiona przez Asn; oraz (j) zastąpienie kwasu karboksylowego w aminokwasie C-końcowym grupą o ładunku obojętnym, ewentualnie amidem. 6. Analog według któregokolwiek z zastrzeżeń 1-, zawierający (a) modyfikację aminokwasu w położeniu 2 nadającą odporność na DPP-IV, i (b) aminokwas w położeniu 40 połączony kowalencyjnie z grupą acylową lub grupą alkilową Analog według któregokolwiek z zastrzeżeń 1-6, zawierający ugrupowanie hydrofilowe połączone z aminokwasem w położeniu 24.
453 42 8. Analog według zastrzeżenia 2, zawierający sekwencję aminokwasową według którejkolwiek z NR. ID. SEKW.: 227, 228, 229 lub 230 oraz przedłużenie o długości 1 do 21 aminokwasów po stronie C-końcowej aminokwasu w położeniu 29, ewentualnie zawierający ponadto do 6 dodatkowych modyfikacji aminokwasów Analog według zastrzeżenia 8, w którym przedłużenie o długości 1 do 21 aminokwasów zawiera sekwencję aminokwasową GPSSGAPPPS (NR. ID. SEKW.:NR. ID. SEKW.: 9) lub XGPSSGAPPPS (NR. ID. SEKW.:NR. ID. SEKW.: 96), w której X stanowi dowolny aminokwas, lub sekwencję aminokwasową zawierającą jedną lub większą liczbę substytucji konserwatywnych względem NR. ID. SEKW.: 9 lub 96..Analog według któregokolwiek z zastrzeżeń 7-8, w którym co najmniej jeden z aminokwasów w przedłużeniu jest acylowany lub alkilowany w położeniu odpowiadającym któremukolwiek z położeń 37-43, ewentualnie aminokwas o wzorze I w położeniu Analog według zastrzeżenia 1, zawierający sekwencję aminokwasową wybraną z grupy składającej się z NR. ID. SEKW.: , , 166, 192-7, i Peptyd składający się z sekwencji aminokwasowej według NR. ID. SEKW.: Peptyd składający się z sekwencji aminokwasowej według NR. ID. SEKW.: 7, NR. ID. SEKW.:
454 43 8, NR. ID. SEKW.: 9, NR. ID. SEKW.: 146 lub NR. ID. SEKW.: Peptyd składający się z aminokwasów według NR. ID. SEKW.: 1 z następującymi modyfikacjami aminokwasów: Tyr w położeniu 1, AIB w położeniu 2, Lys w położeniu, przy czym Lys jest połączona kowalencyjnie z tłuszczową grupą acylową C16 poprzez wstawkę dipeptydową γ-glu-γ-glu, Ile w położeniu 12, Lys w położeniu 16, Gln w położeniu 17, Ala w położeniu 18, AIB w położeniu, Glu w położeniu 21, Asn w położeniu 24, Leu w położeniu 27, Ala w położeniu 28, Gly w położeniu 29, a następnie aminokwas według NR. ID. SEKW.: 9 po stronie C-końcowej aminokwasu w położeniu 29 i C-końcowy amid zamiast C-końcowego alfakarboksylanu. 2 1.Peptyd zawierający sekwencję aminokwasową według NR. ID. SEKW.:, przy czym wartość EC0 peptydu wobec aktywacji receptora GIP wynosi około nm lub mniej, i przy czym wartość EC0 peptydu wobec receptora GIP różni się mniej niż około 0-krotnie względem jego wartości EC0 wobec receptora GLP-1, przy czym ewentualnie siła działania GIP peptydu pozostaje w granicach około 1-krotności siły działania GLP-1 tego peptydu Peptyd zawierający sekwencję aminokwasową według NR. ID. SEKW.: 7, NR. ID. SEKW.: 8, NR. ID. SEKW.: 9, NR. ID. SEKW.: 146 lub NR. ID. SEKW.: 13, przy czym wartość EC0 peptydu wobec aktywacji receptora GIP wynosi około nm lub mniej, i przy czym
455 44 wartość EC0 peptydu wobec receptora GIP różni się mniej niż około 0-krotnie względem jego wartości EC0 wobec receptora GLP-1, przy czym ewentualnie siła działania GIP peptydu pozostaje w granicach około 1- krotności siły działania GLP-1 tego peptydu Peptyd zawierający aminokwas według NR. ID. SEKW.: 1 z następującymi modyfikacjami aminokwasów: Tyr w położeniu 1, AIB w położeniu 2, Lys w położeniu, przy czym Lys jest połączona kowalencyjnie z tłuszczową grupą acylową C16 poprzez wstawkę dipeptydową γ-glu-γ- Glu, Ile w położeniu 12, Lys w położeniu 16, Gln w położeniu 17, Ala w położeniu 18, AIB w położeniu, Glu w położeniu 21, Asn w położeniu 24, Leu w położeniu 27, Ala w położeniu 28, Gly w położeniu 29, a następnie aminokwas według NR. ID. SEKW.: 9 po stronie C- końcowej aminokwasu w położeniu 29 i C-końcowy amid zamiast C-końcowego alfa-karboksylanu, przy czym wartość EC0 peptydu wobec aktywacji receptora GIP wynosi około nm lub mniej, i przy czym wartość EC0 peptydu wobec receptora GIP różni się mniej niż około 0-krotnie względem jego wartości EC0 wobec receptora GLP-1, przy czym ewentualnie siła działania GIP peptydu pozostaje w granicach około 1-krotności siły działania GLP-1 tego peptydu Analog według zastrzeżeń 1-9, zawierający grupę acylową lub grupę alkilową połączoną z łańcuchem bocznym aminokwasu przez wstawkę, przy czym ewentualnie wstawka ma długość 3 do atomów i stanowi dipeptyd, przy czym ewentualnie wstawka zawiera dwa aminokwasy naładowane ujemnie.
456 4 19.Analog według zastrzeżenia 18, w którym grupę acylową stanowi tłuszczowa grupa acylowa C12 do C18, ewentualnie tłuszczowa grupa acylowa C14 lub C16. 1.Analog według któregokolwiek z zastrzeżeń 1-11 lub lub peptyd według któregokolwiek z zastrzeżeń 12-17, przy czym analog lub peptyd jest połączony kowalencyjnie z ugrupowaniem hydrofilowym, ewentualnie poli(glikolem etylenowym) (PEG), w dowolnym spośród położeń aminokwasów 19,, 23, 24, 27, 32, 43 lub z końcem C, przy czym ewentualnie ugrupowanie hydrofilowe jest połączone kowalencyjnie z Lys, Cys, Orn, homocysteiną lub acetylofenyloalaniną. 21.Analog lub peptyd według zastrzeżenia, przy czym PEG ma masę cząsteczkową około 000 daltonów do około daltonów. 22.Koniugat, dimer lub peptyd fuzyjny zawierający analog według któregokolwiek z zastrzeżeń 1-11 lub lub peptyd według któregokolwiek z zastrzeżeń lub Kompozycja farmaceutyczna zawierająca analog według któregokolwiek z zastrzeżeń 1-11 lub lub peptyd według któregokolwiek z zastrzeżeń lub - 21, koniugat, dimer lub peptyd fuzyjny według zastrzeżenia 22, lub ich kombinację, oraz dopuszczalny farmaceutycznie nośnik. 24.Zestaw zawierający kompozycję farmaceutyczną według zastrzeżenia 23 i urządzenie do podawania wspomnianej kompozycji farmaceutycznej pacjentowi, przy
457 46 czym ewentualnie urządzenie stanowi strzykawka zawierająca kompozycję farmaceutyczną. 2.Analog według któregokolwiek z zastrzeżeń 1-11 lub 18-21, lub peptyd według któregokolwiek z zastrzeżeń lub -21 do zastosowania w zmniejszaniu wzrostu masy lub indukowania utraty masy, lub zastosowanie analogu według któregokolwiek z zastrzeżeń 1-11 lub 18-21, lub peptydu według któregokolwiek z zastrzeżeń lub -21, do wytwarzania leku do zmniejszania wzrostu masy lub indukowania utraty masy Analog według któregokolwiek z zastrzeżeń 1-11 lub 18-21, lub peptyd według któregokolwiek z zastrzeżeń lub -21, do zastosowania w leczeniu cukrzycy, lub zastosowanie analogu według któregokolwiek z zastrzeżeń 1-11 lub 18-21, lub peptydu według któregokolwiek z zastrzeżeń lub -21, do wytwarzania leku do leczenia cukrzycy. 2 Indiana University Research and Technology Corporation Pełnomocnik:

458 47

459 48
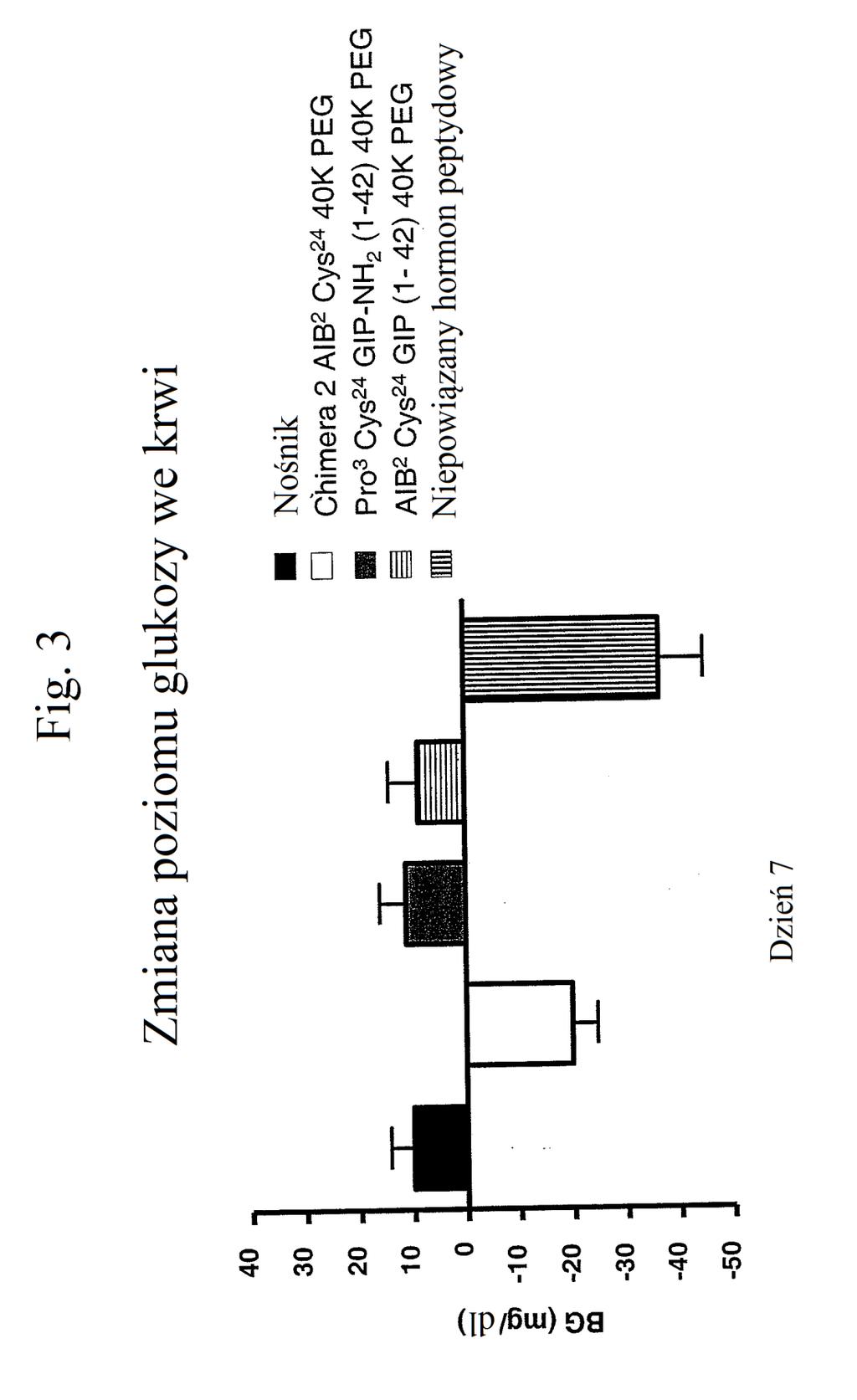
460 49

461 460
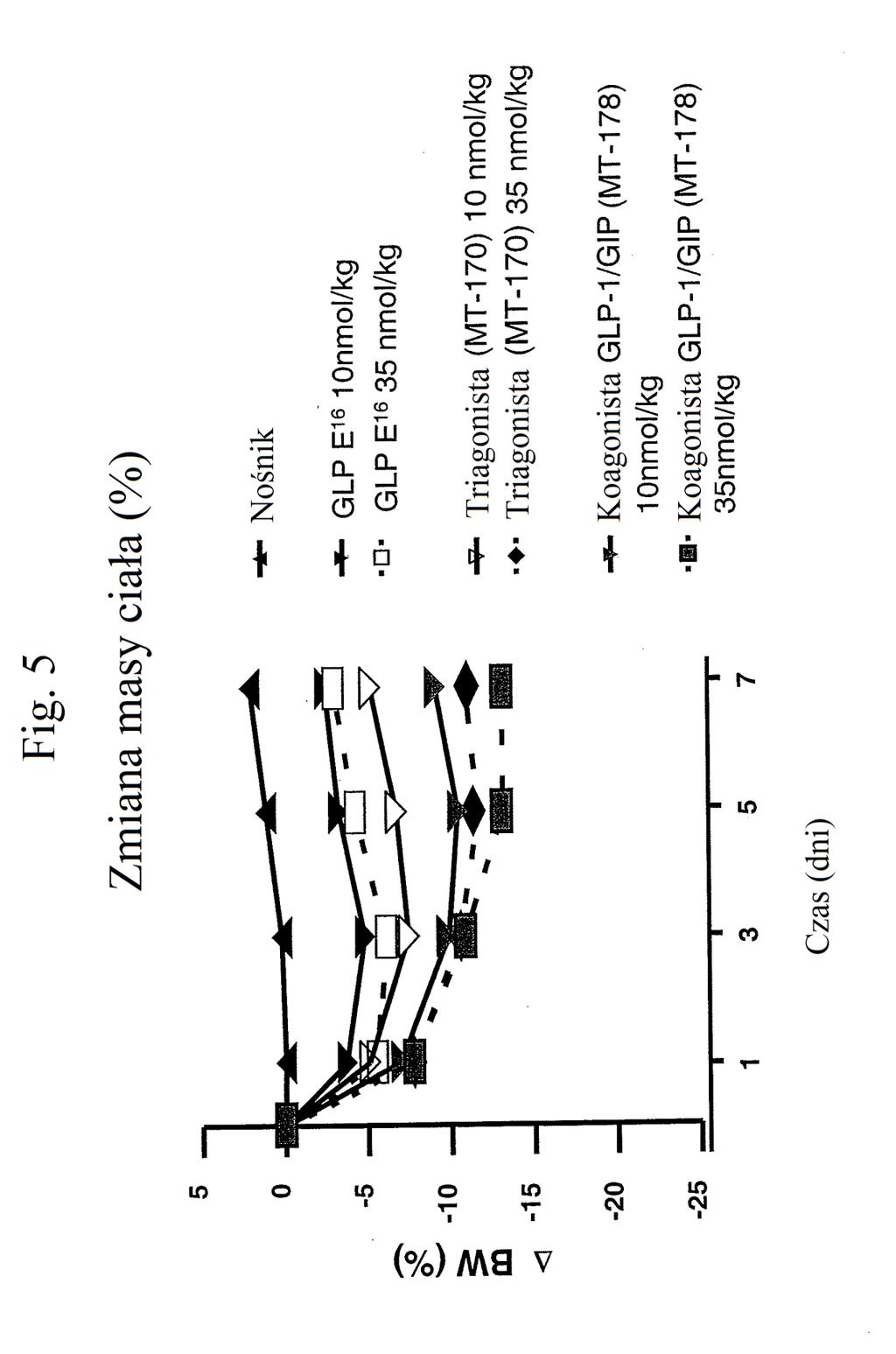
462 461
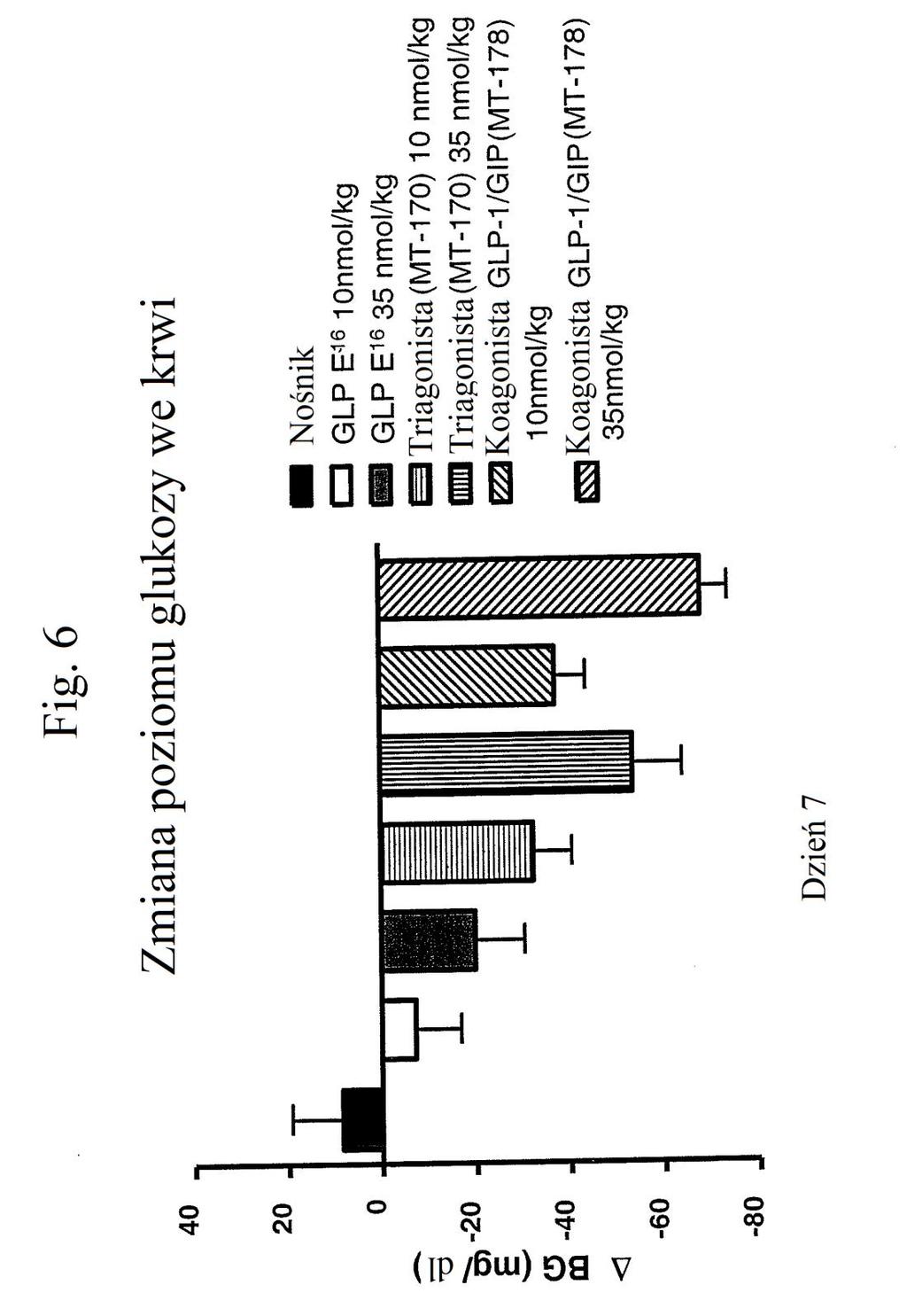
463 462
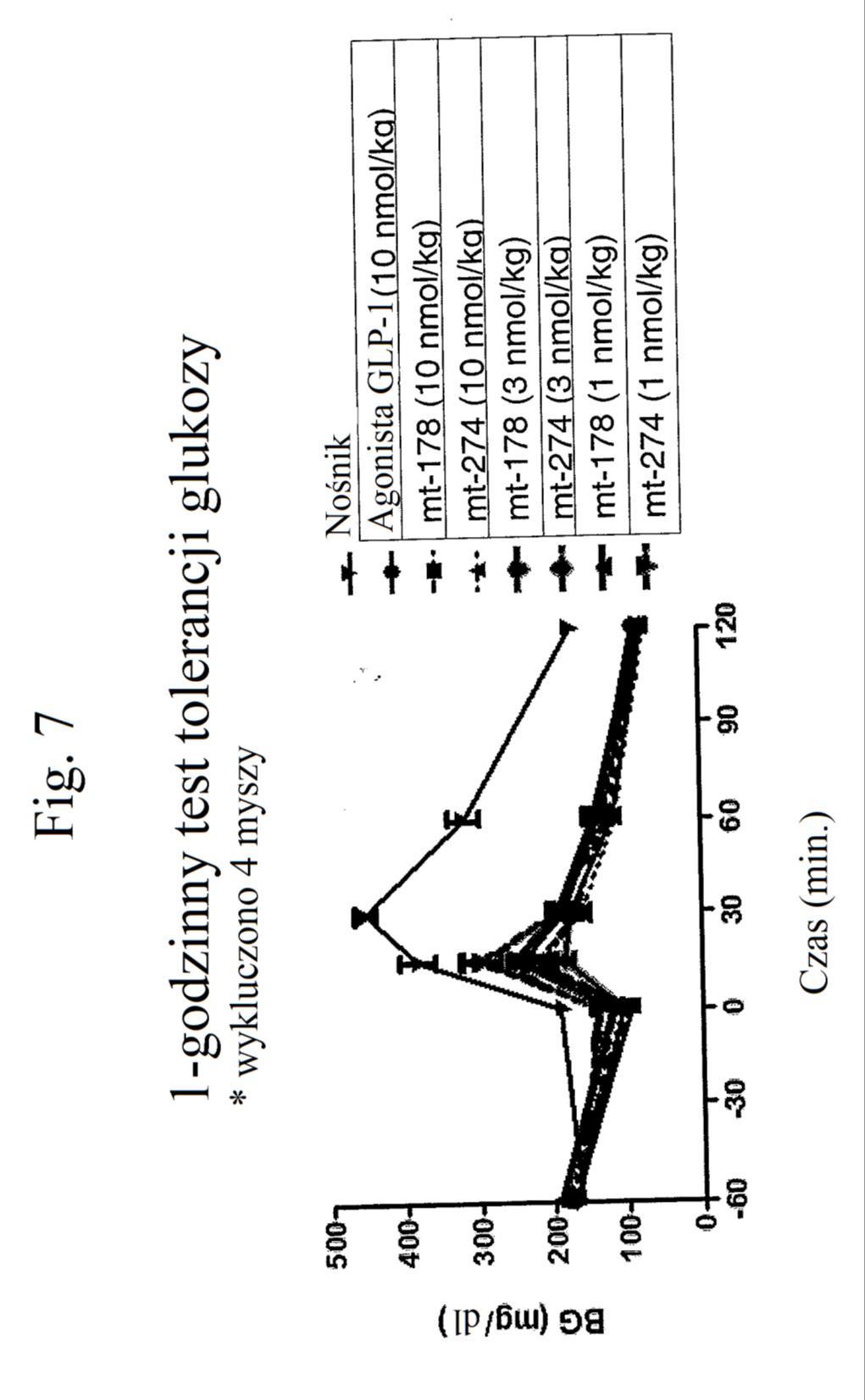
464 463
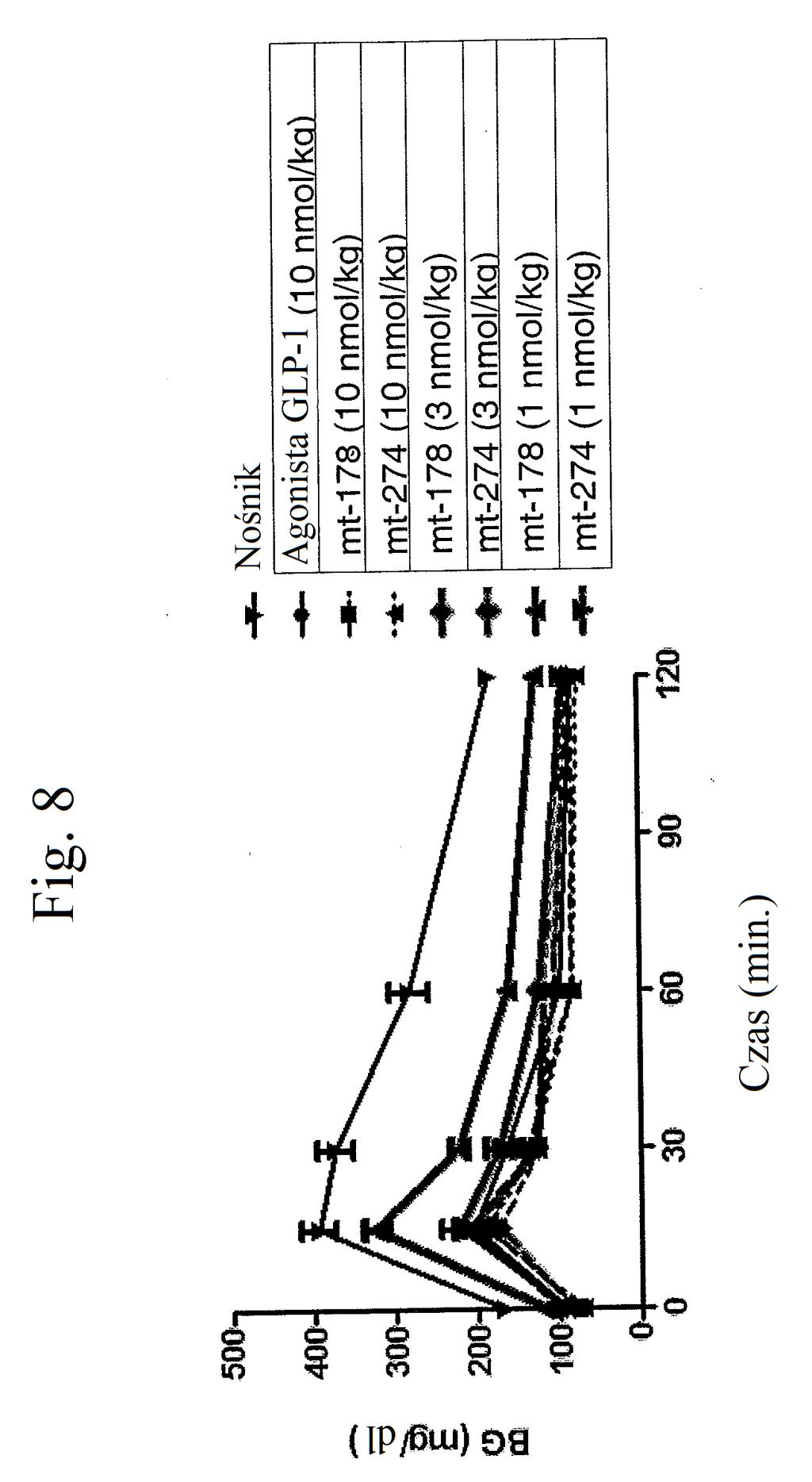
465 464

466 46
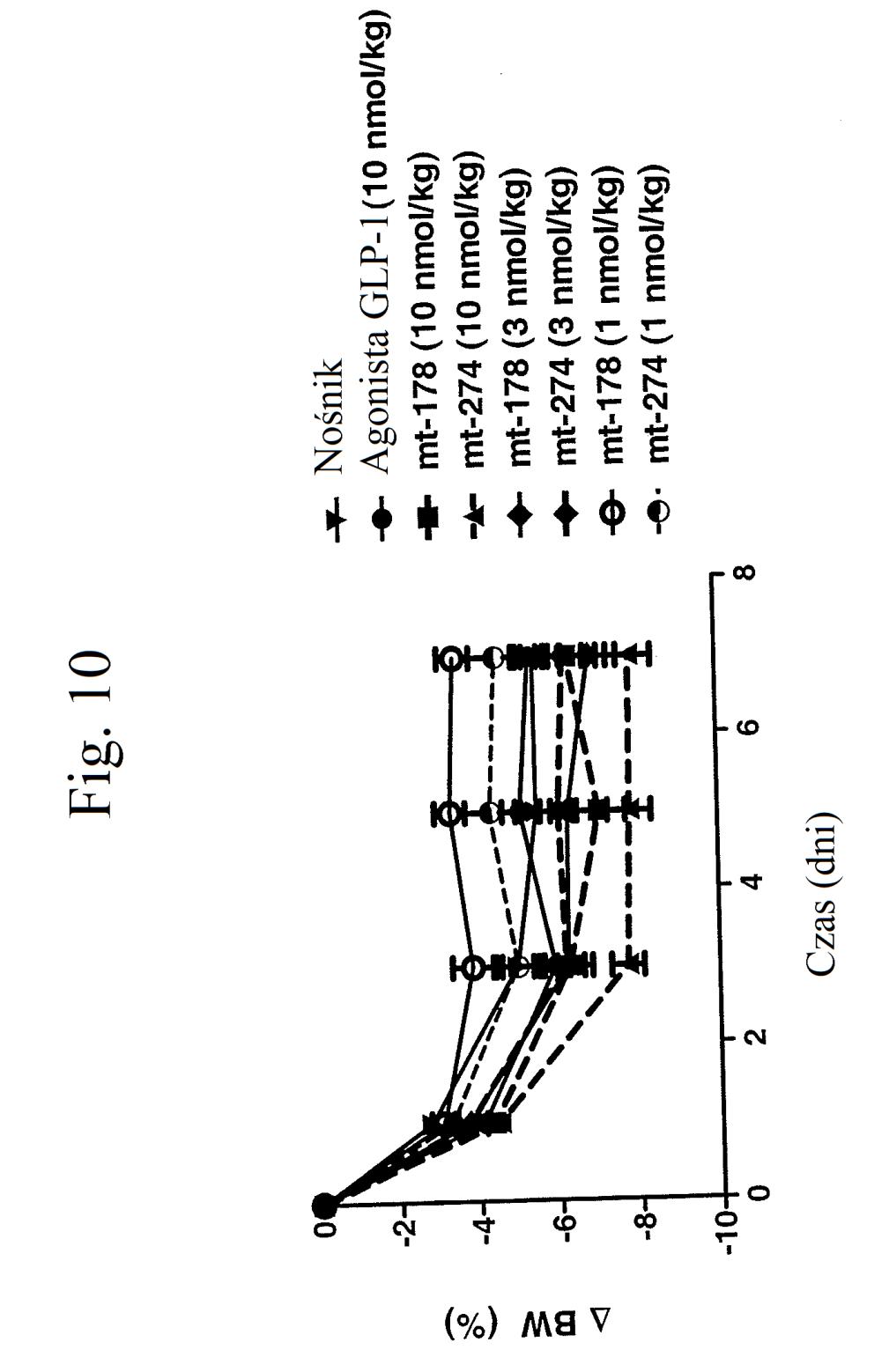
467 466

468 467
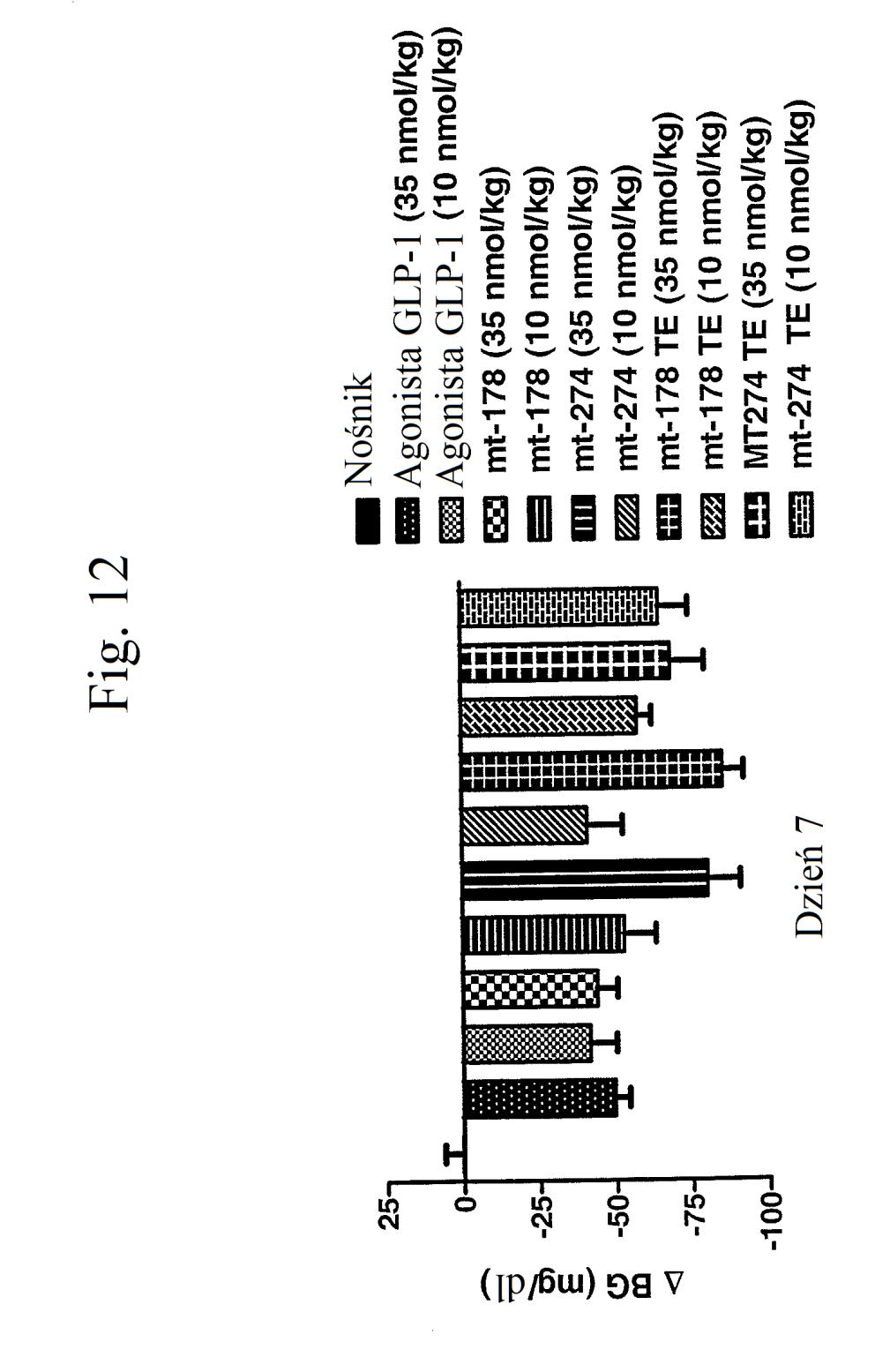
469 468
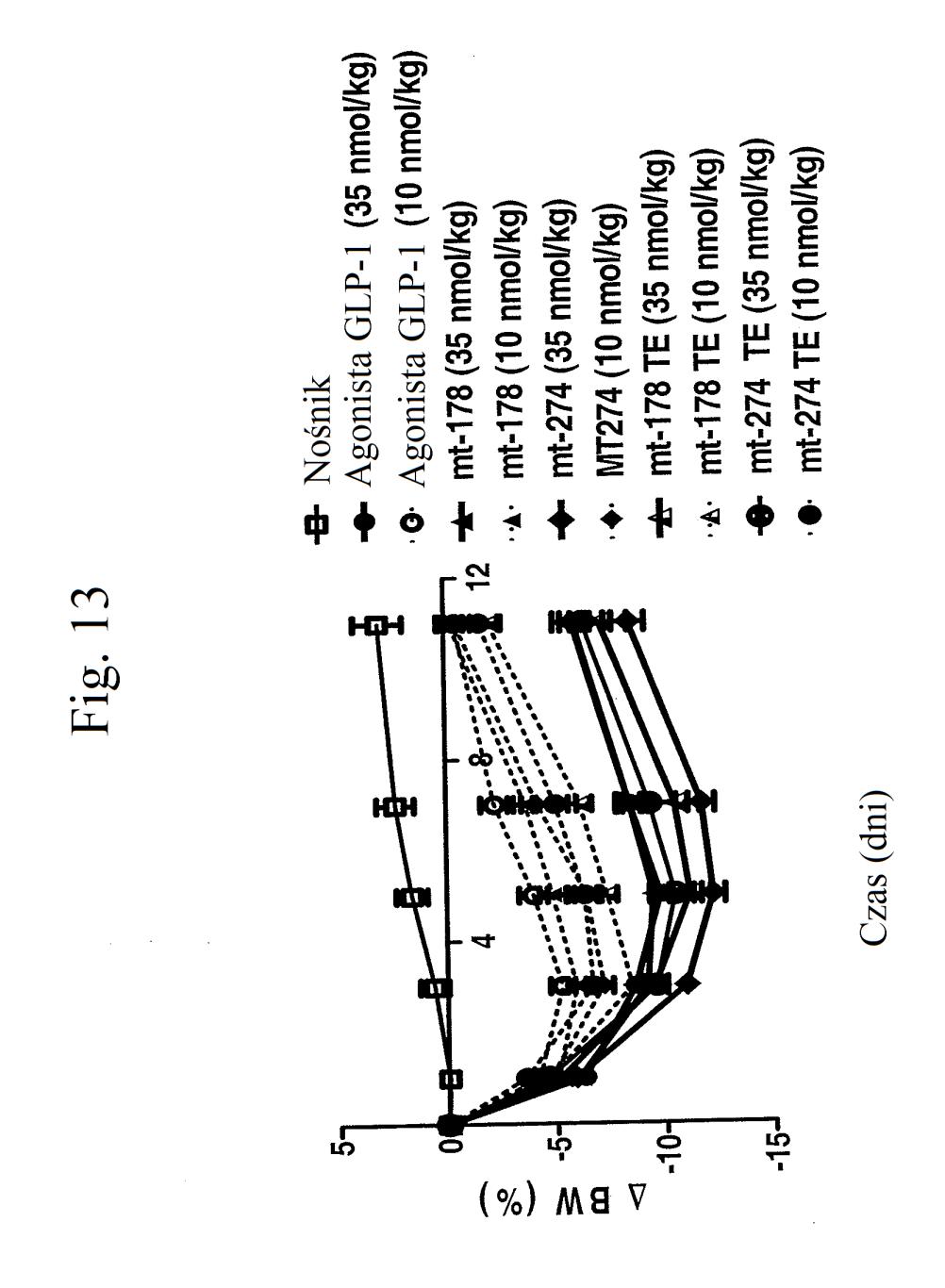
470 469
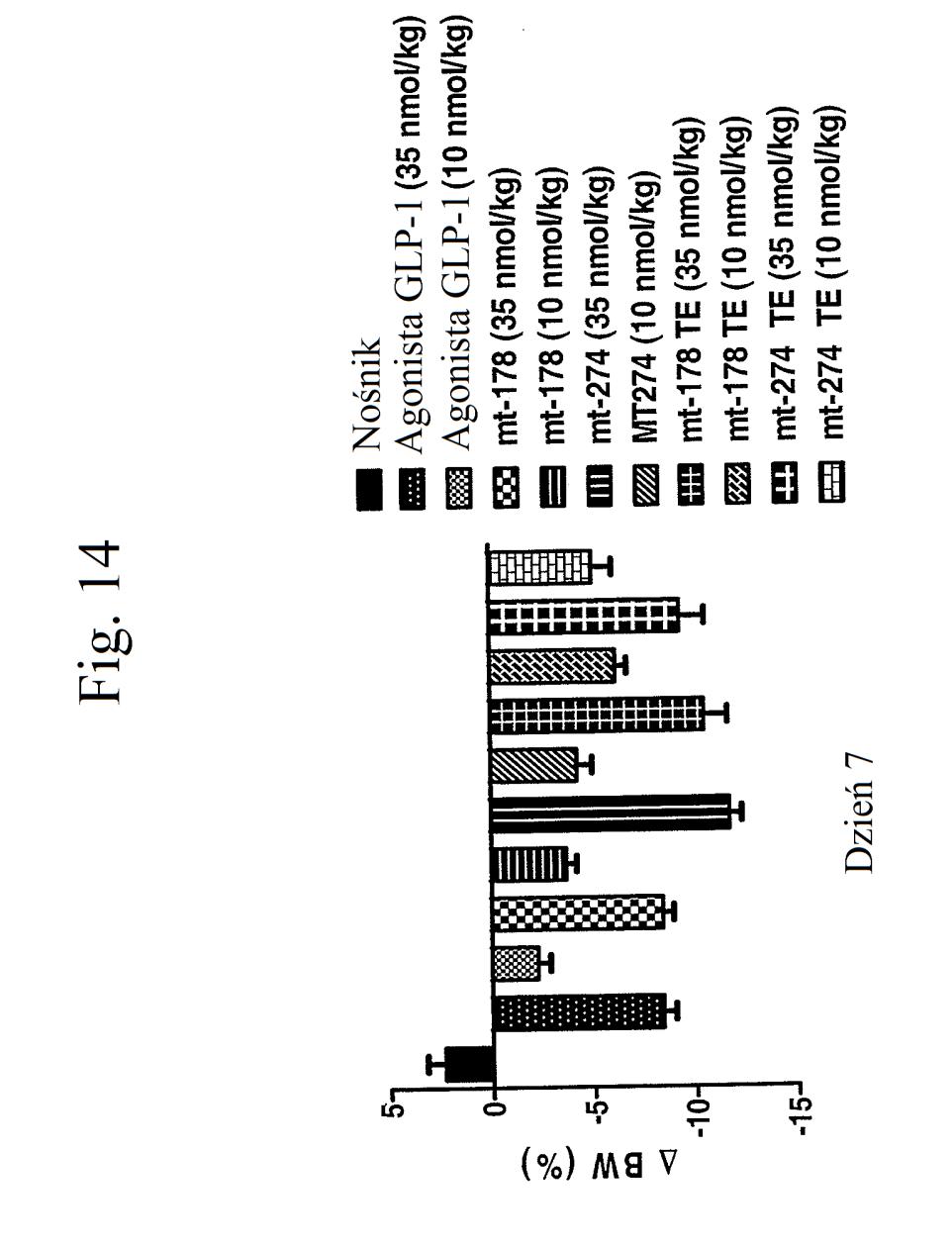
471 470
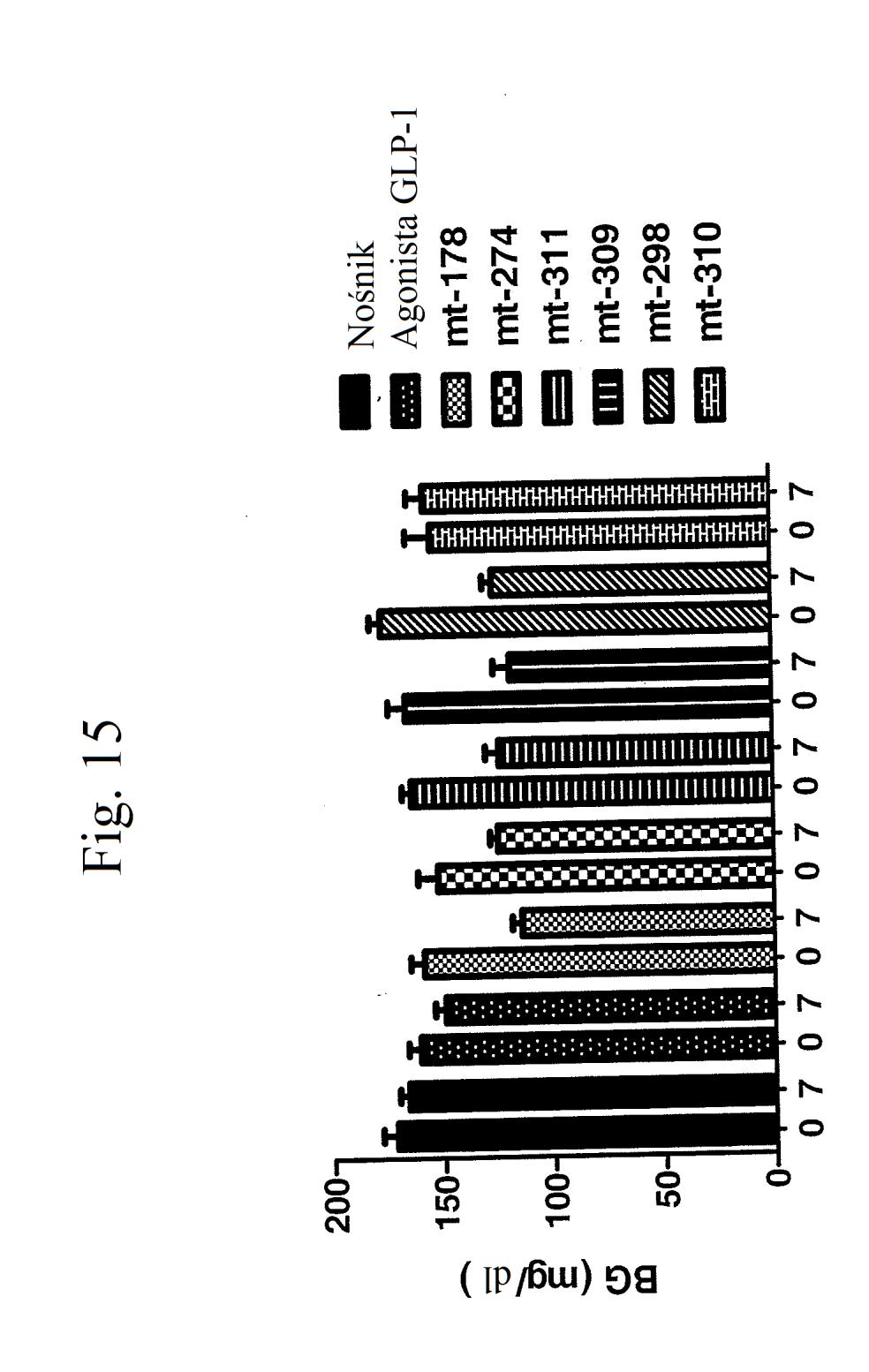
472 471
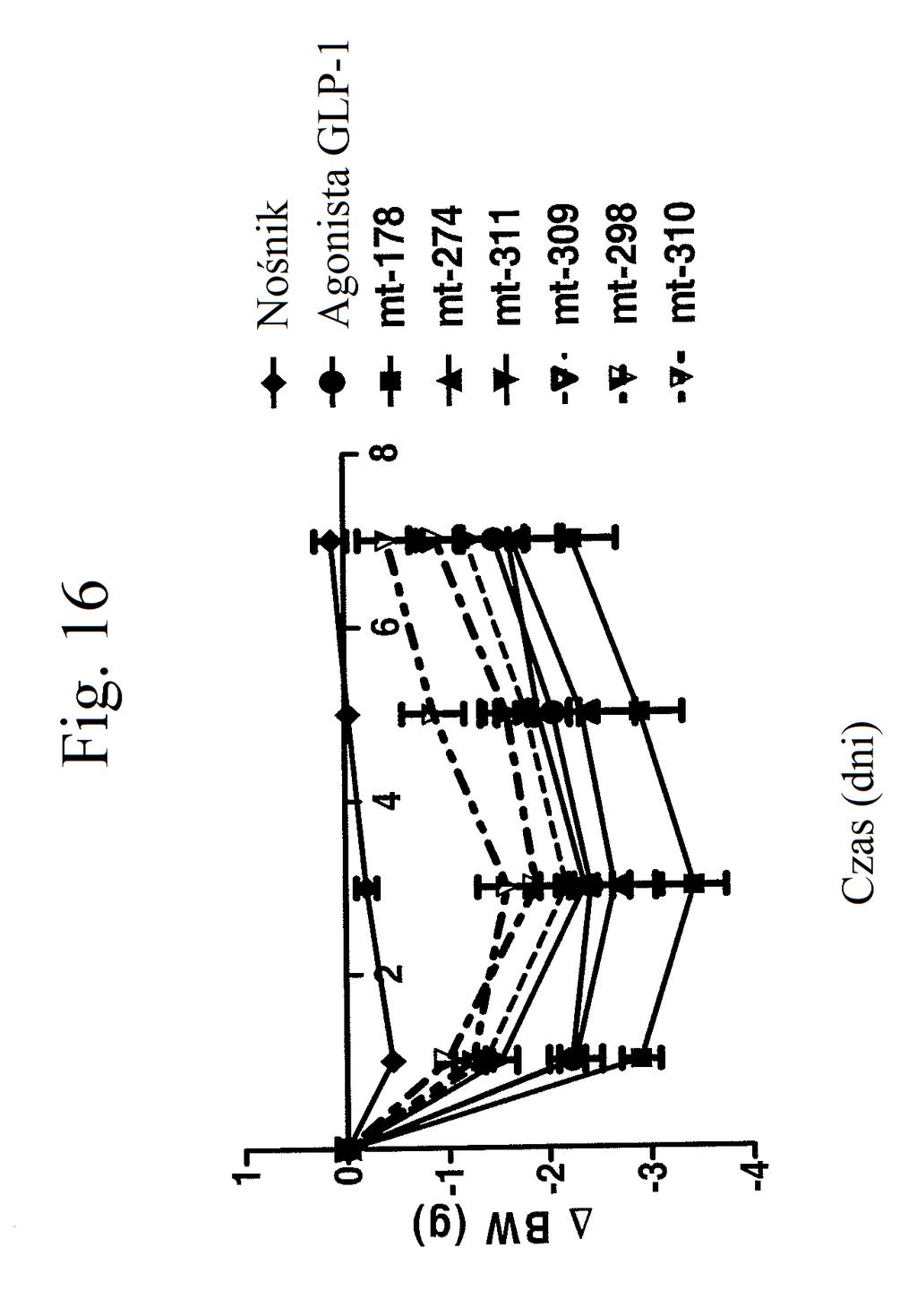
473 472
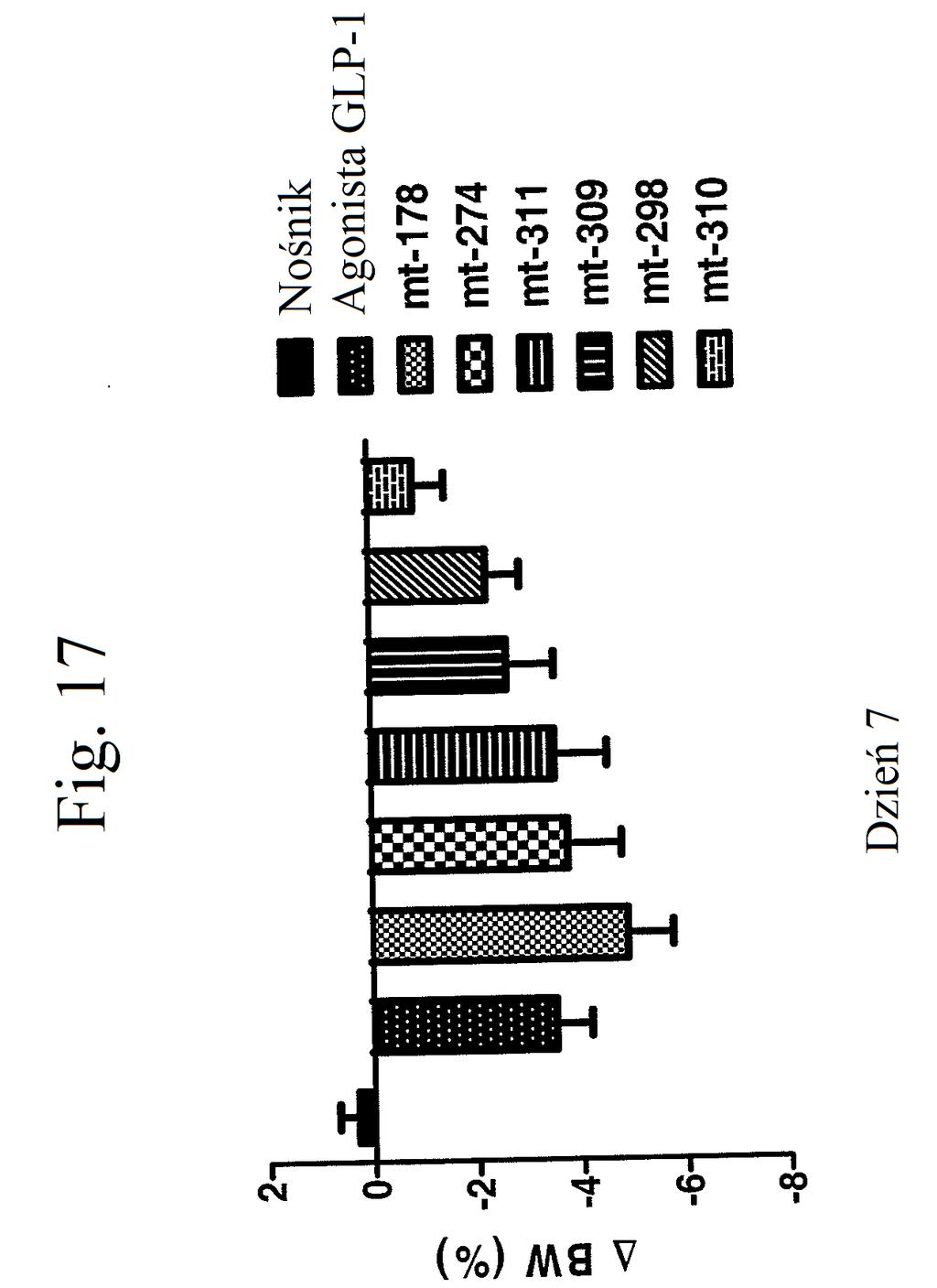
474 473
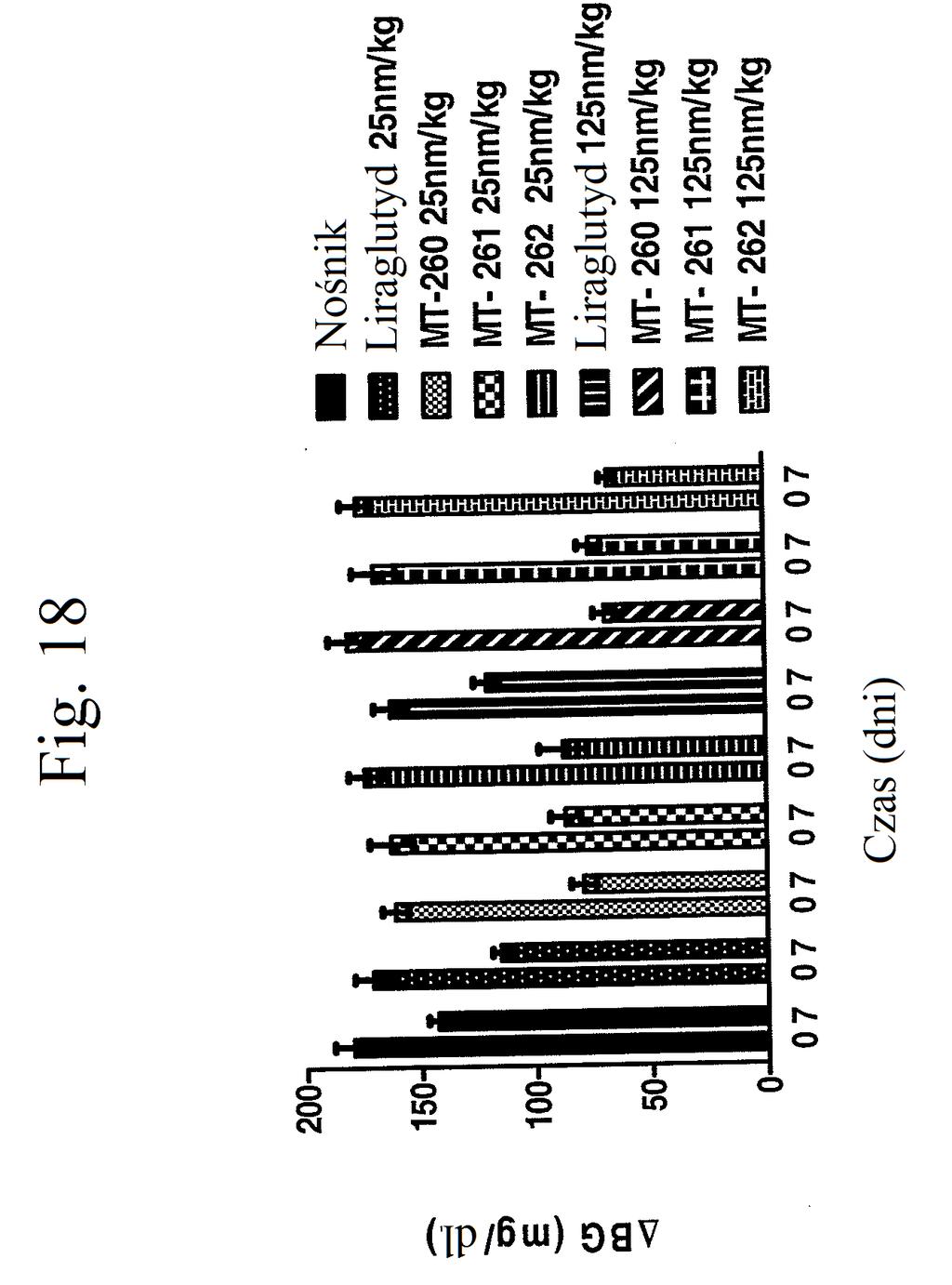
475 474

476 47

477 476
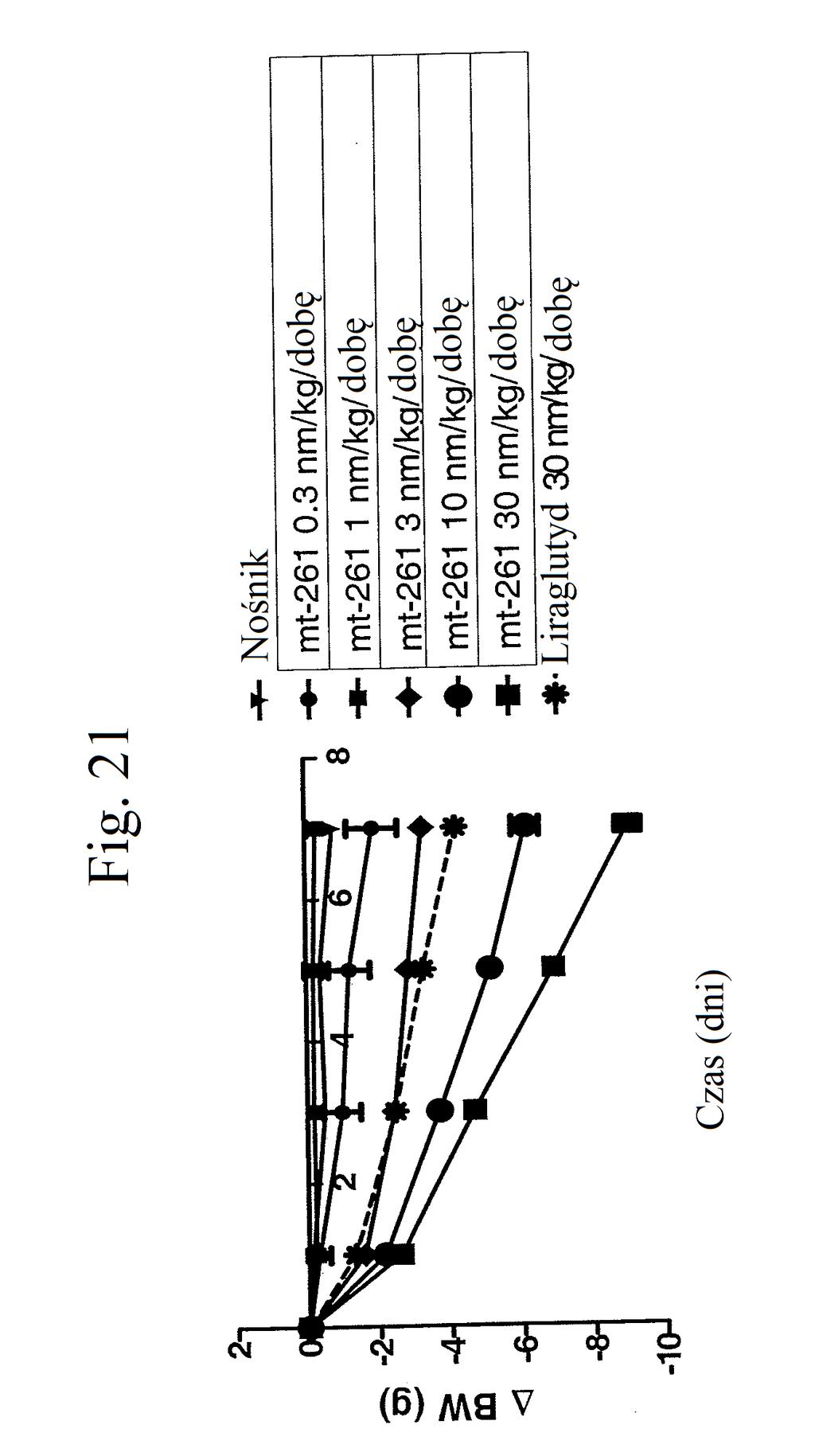
478 477
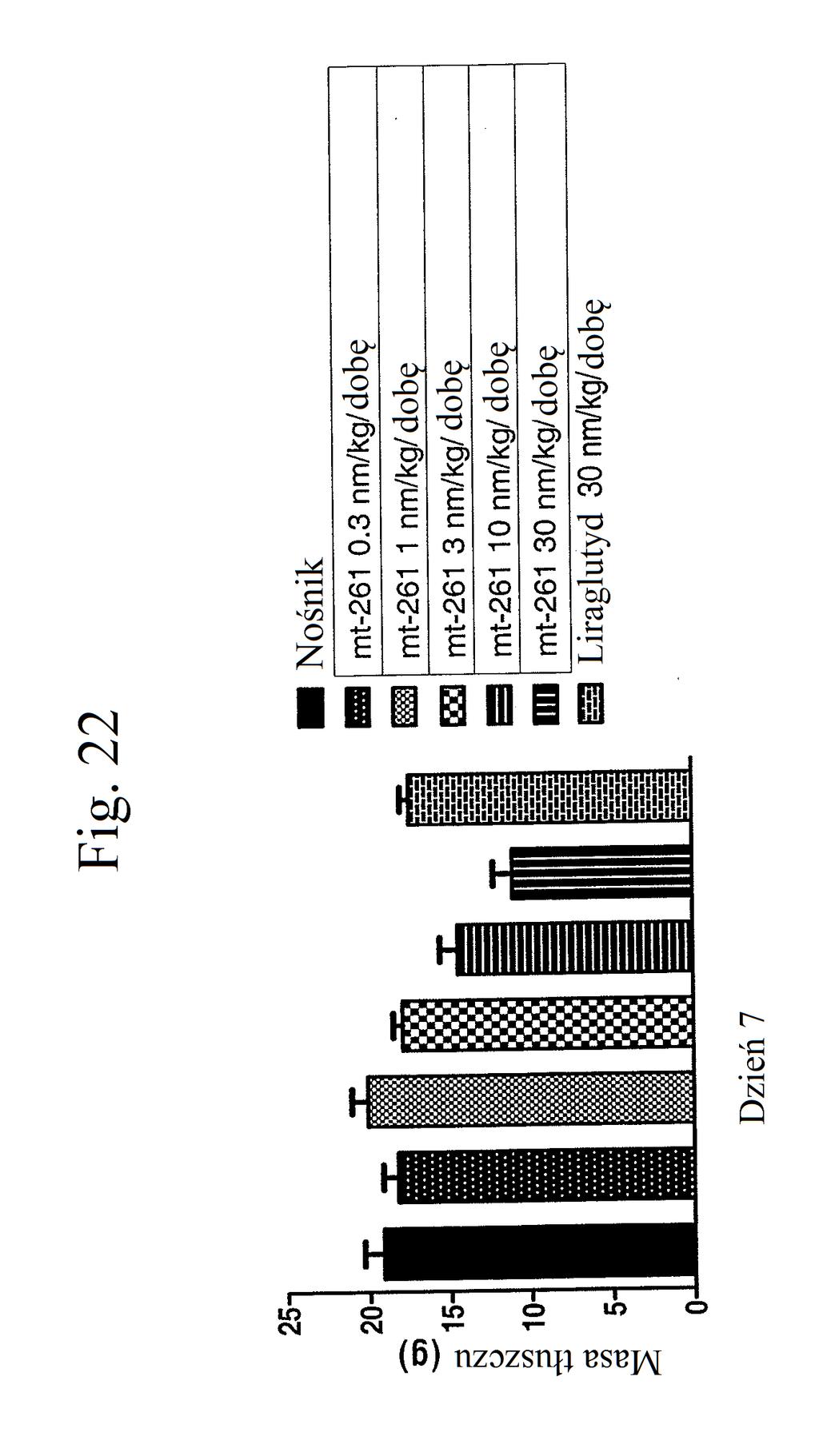
479 478
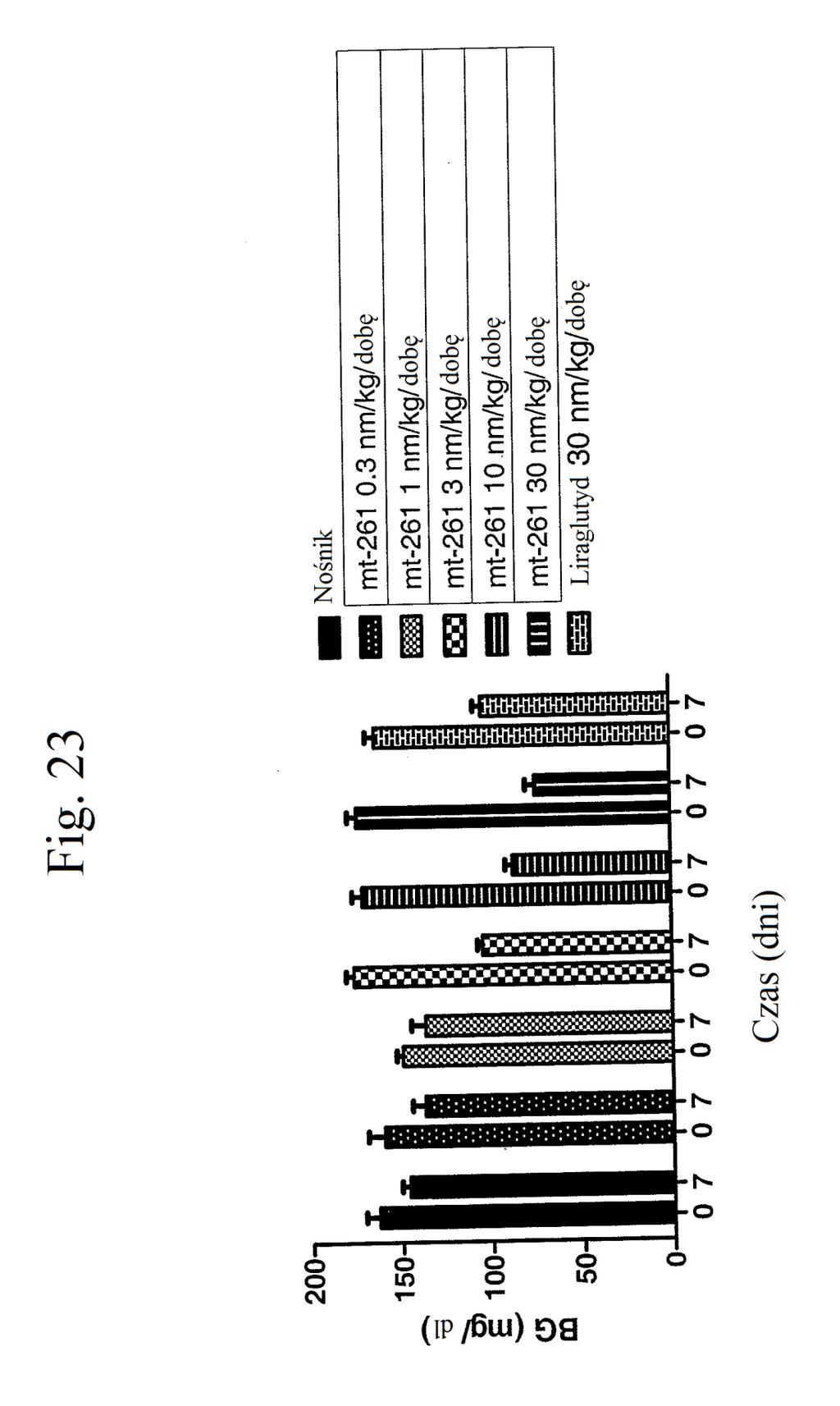
480 479
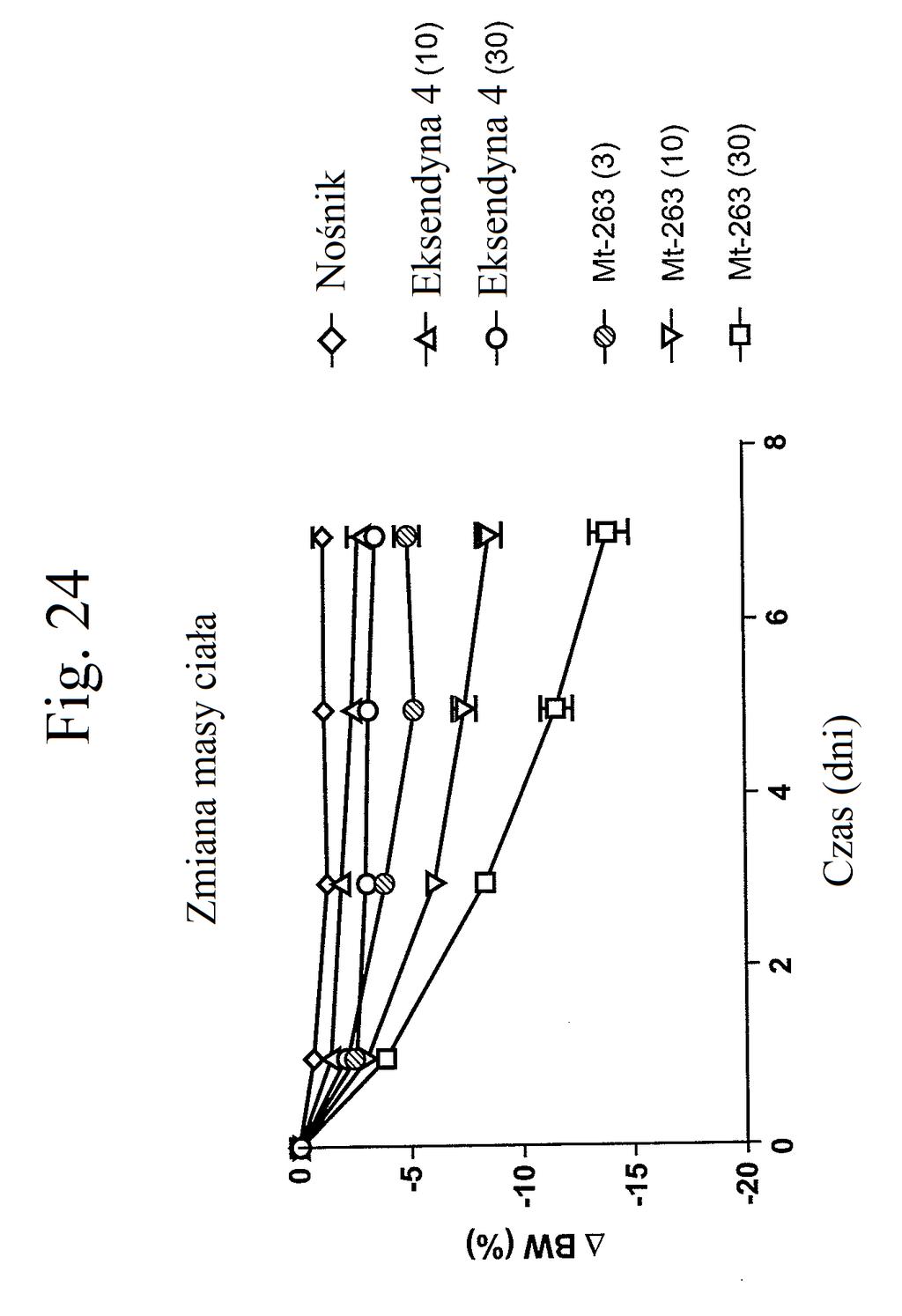
481 480
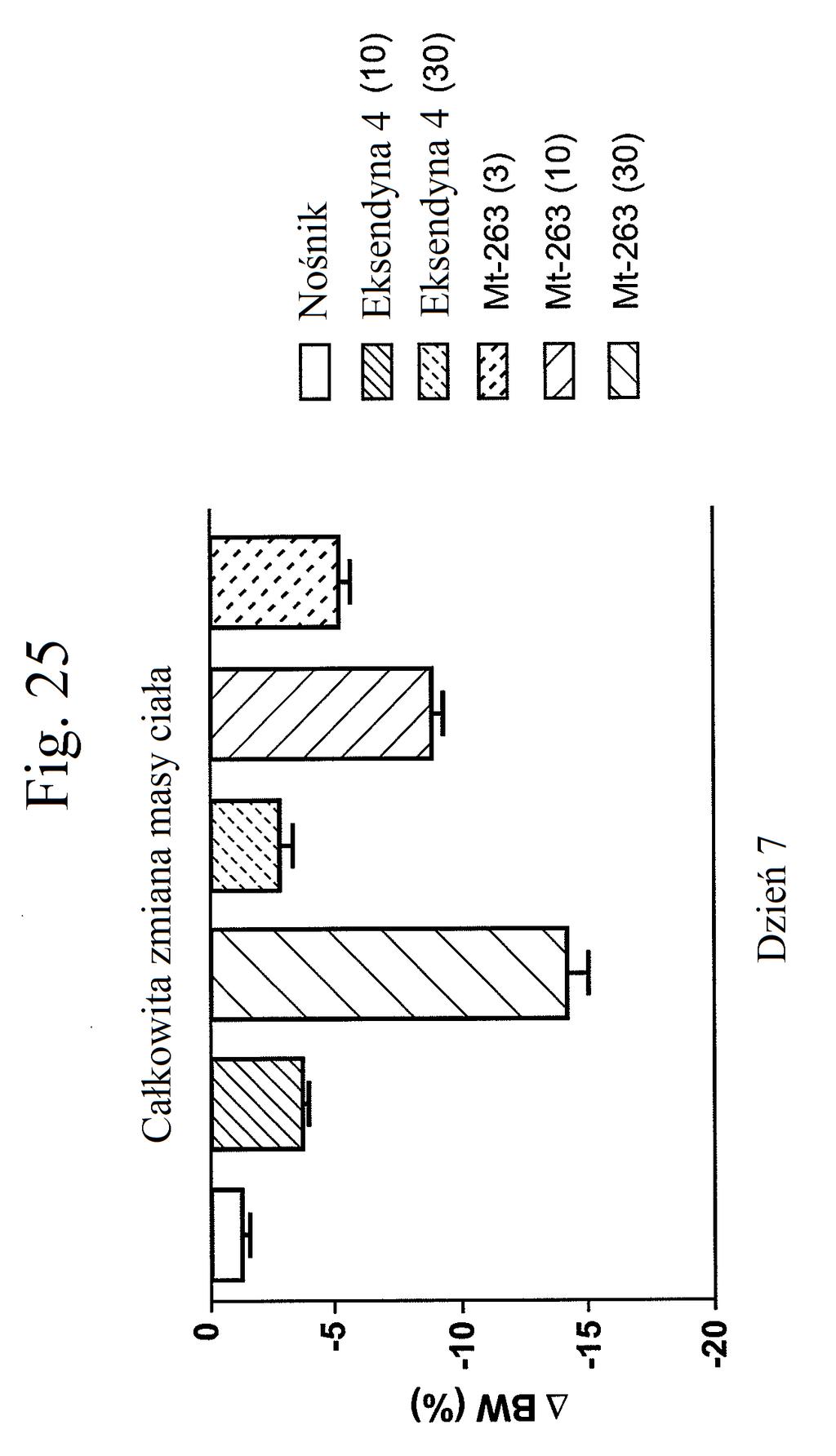
482 481

483 482
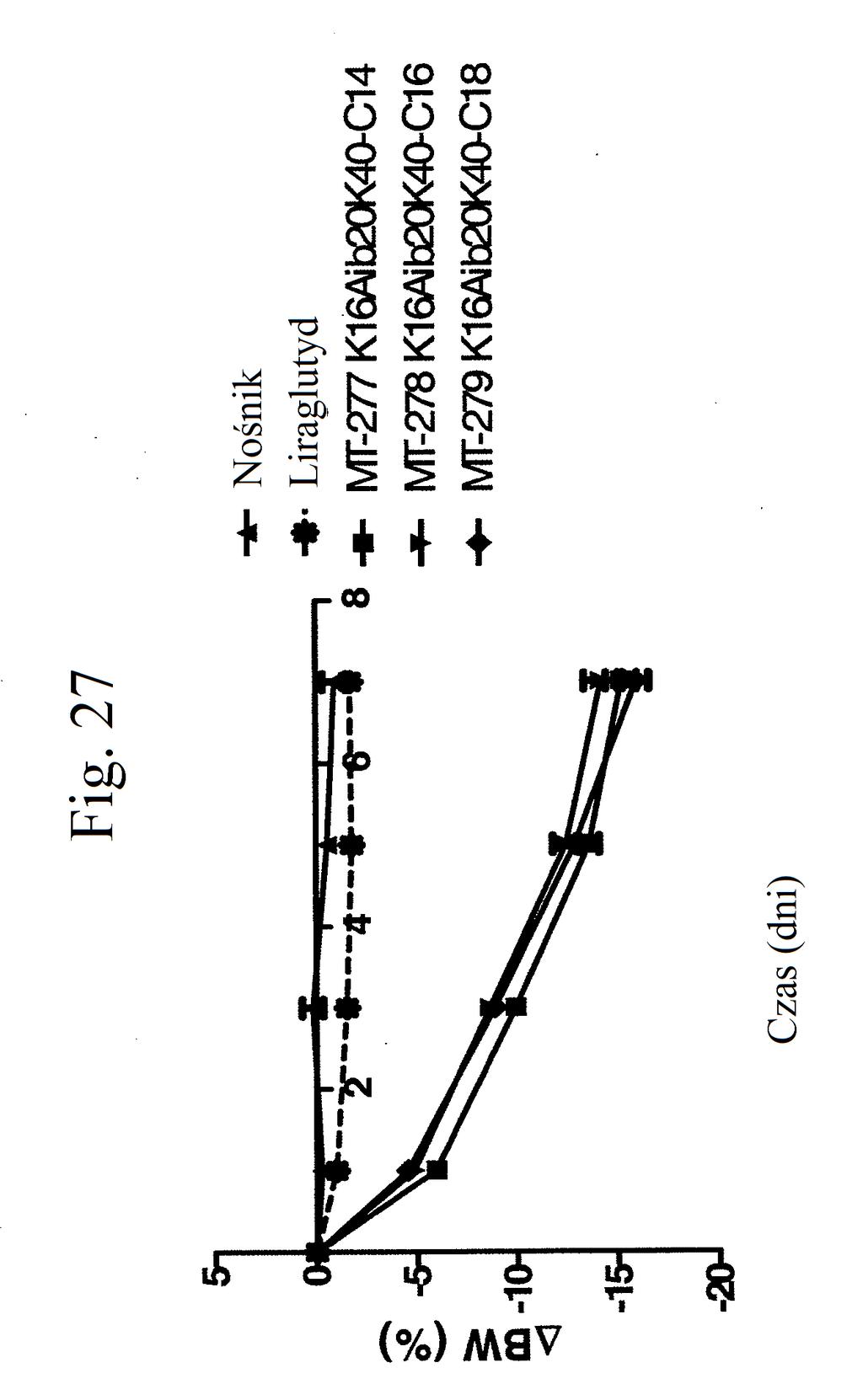
484 483
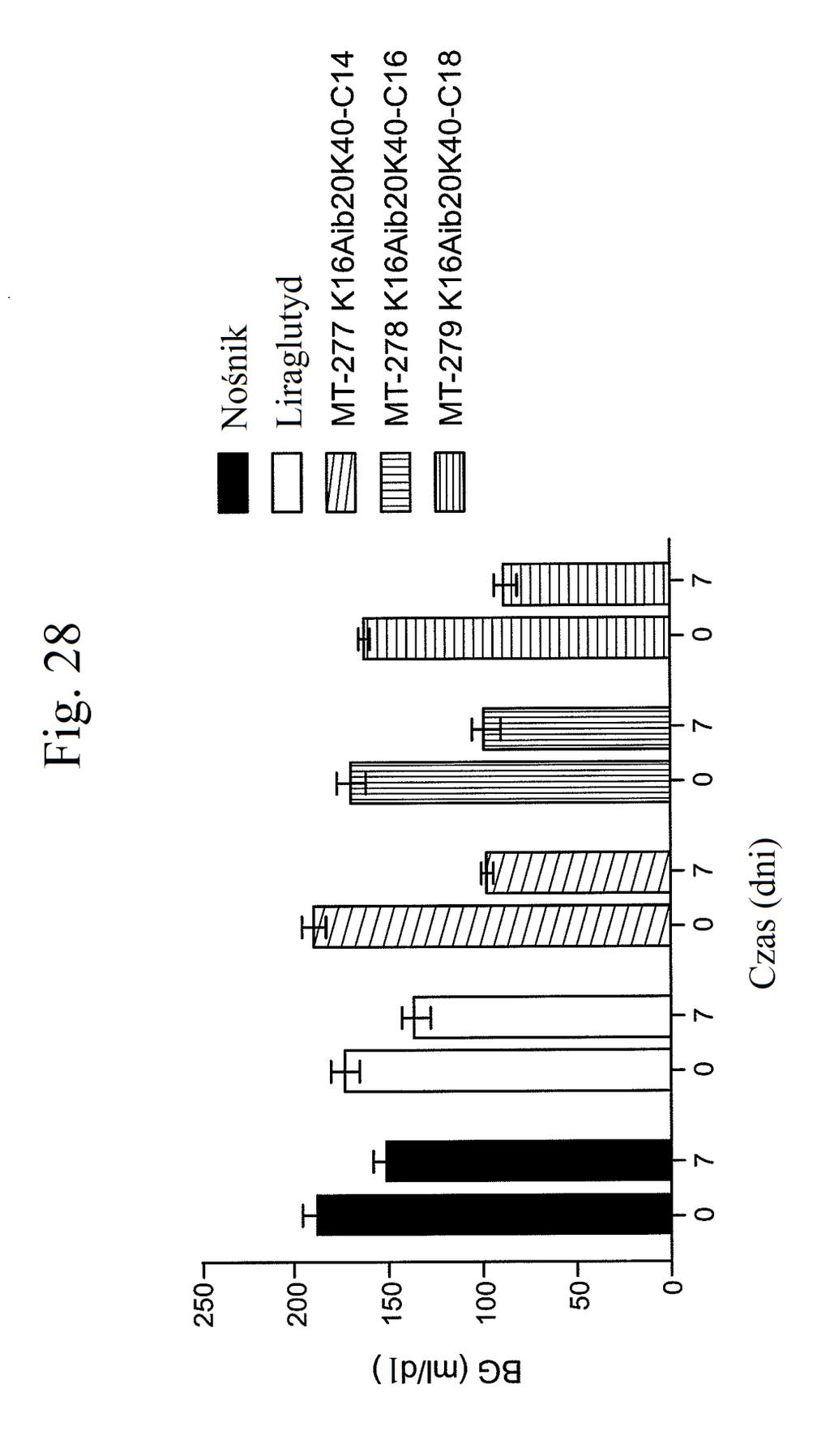
485 484
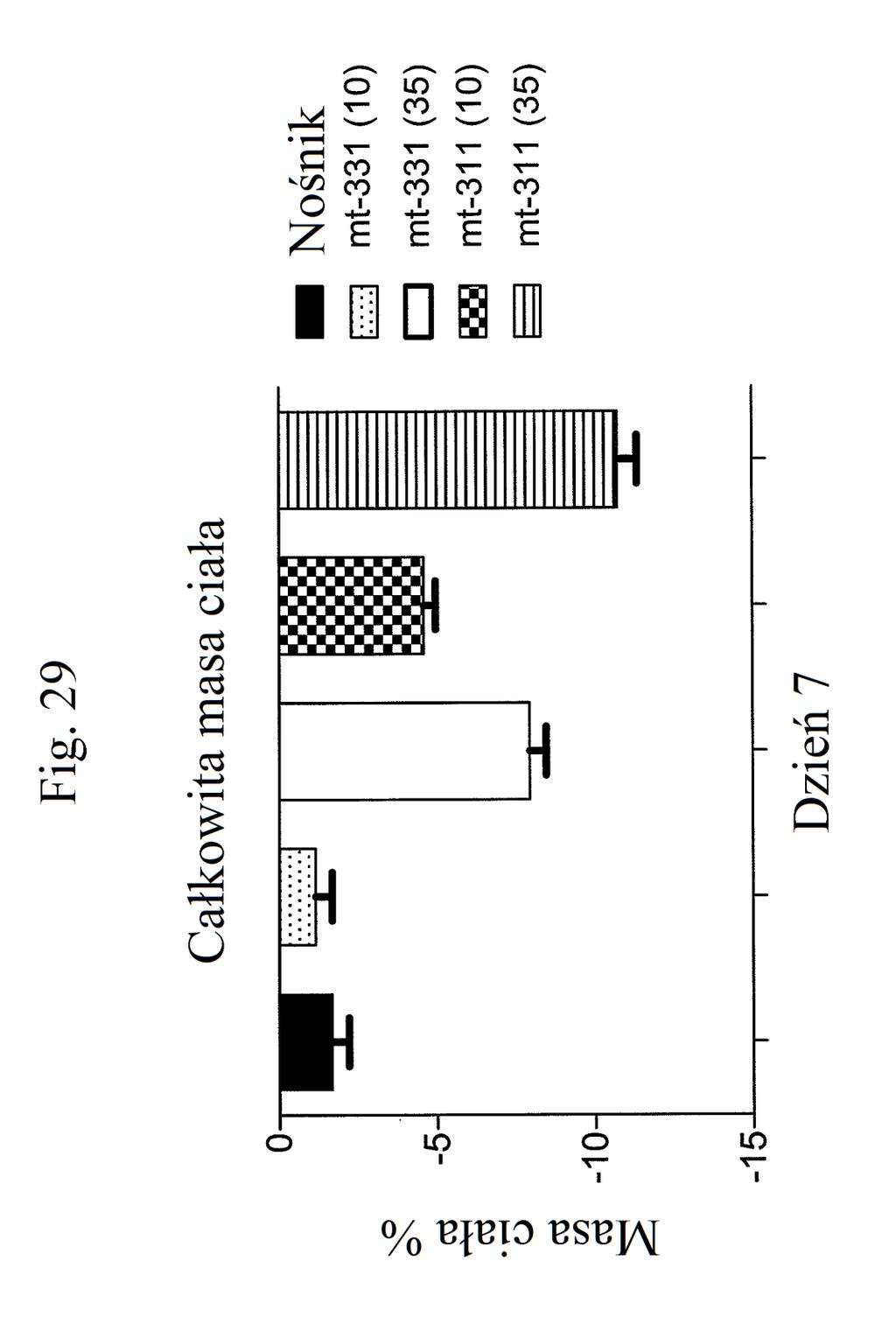
486 48

487 486
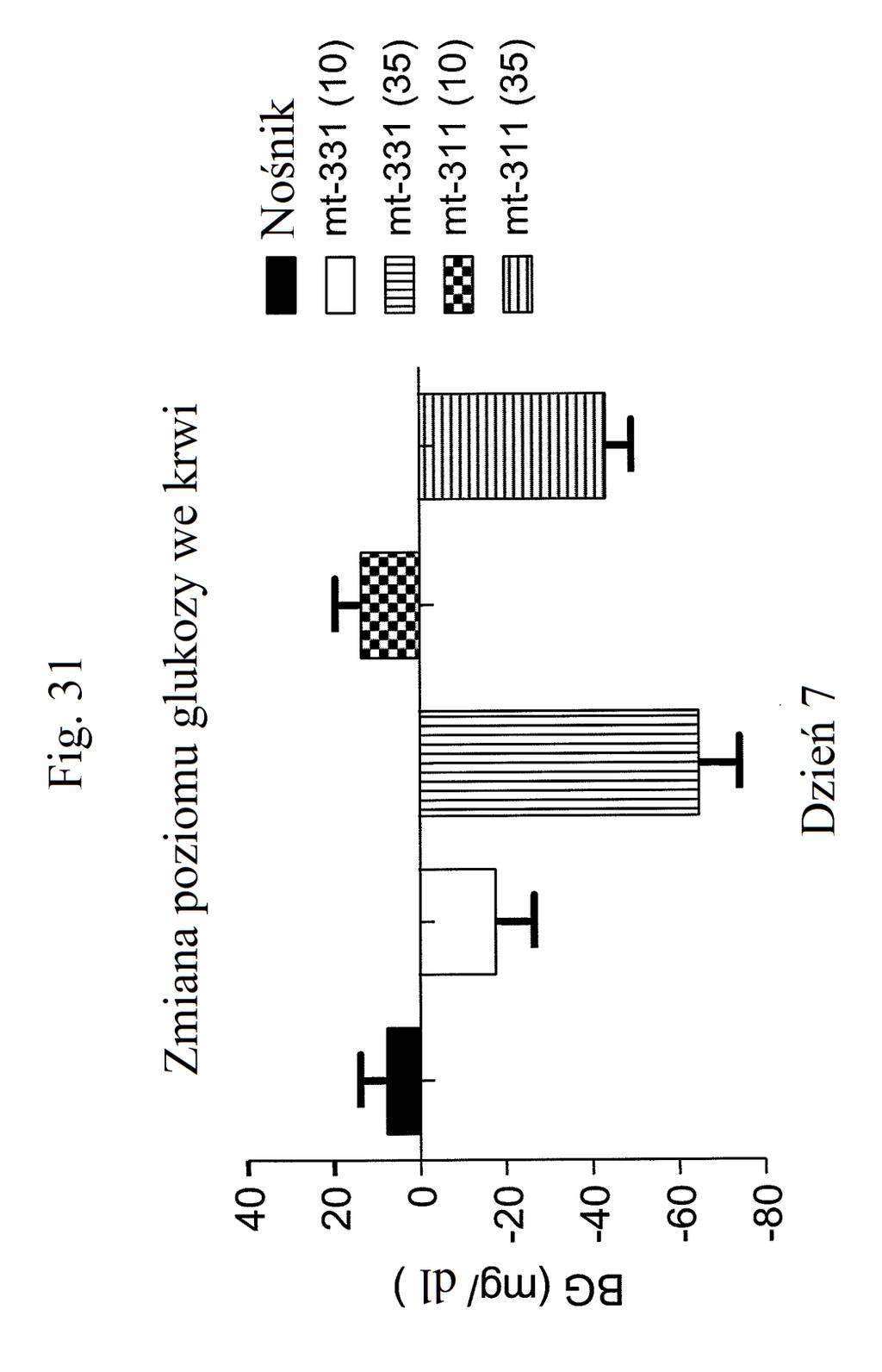
488 487
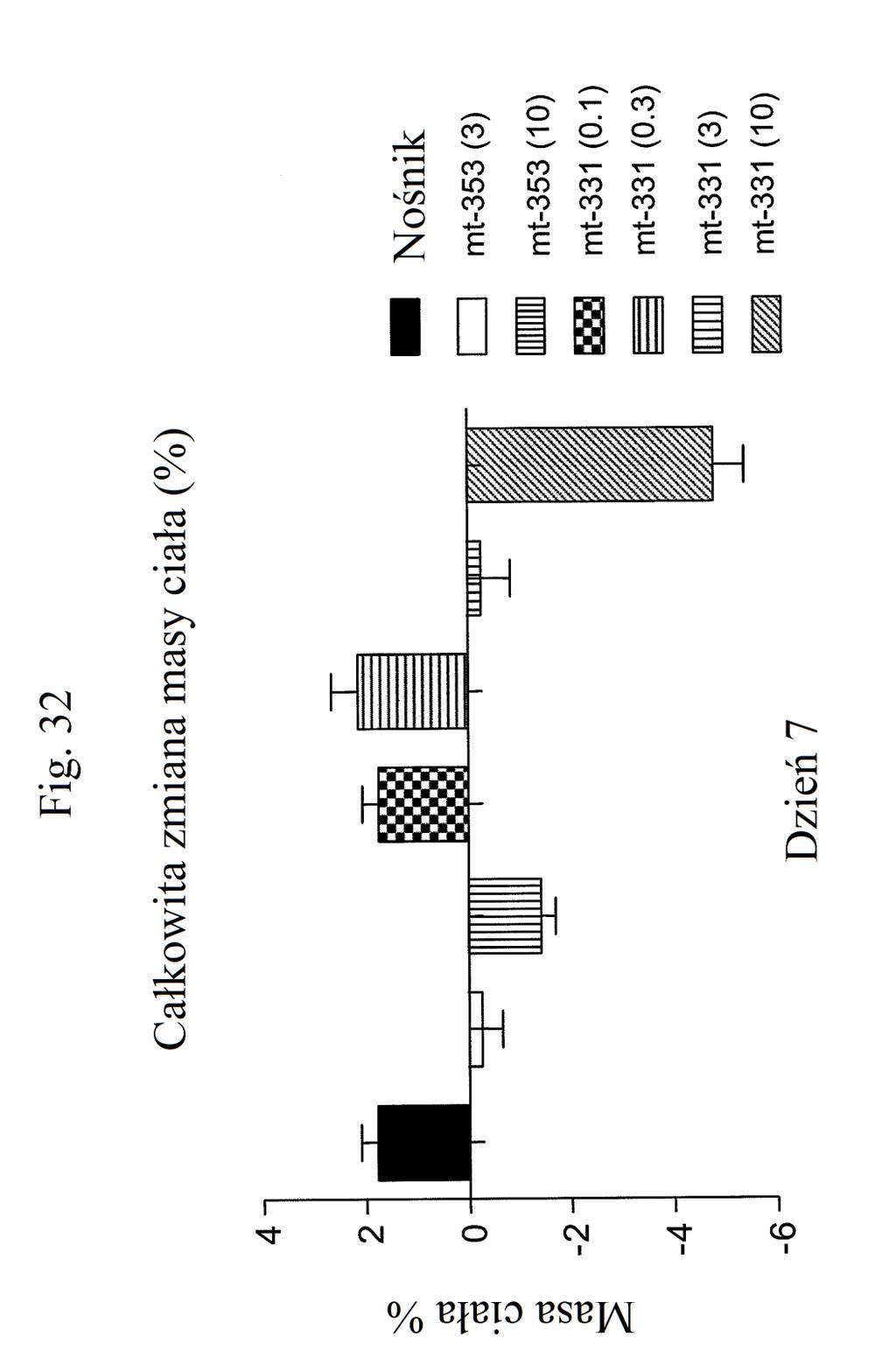
489 488
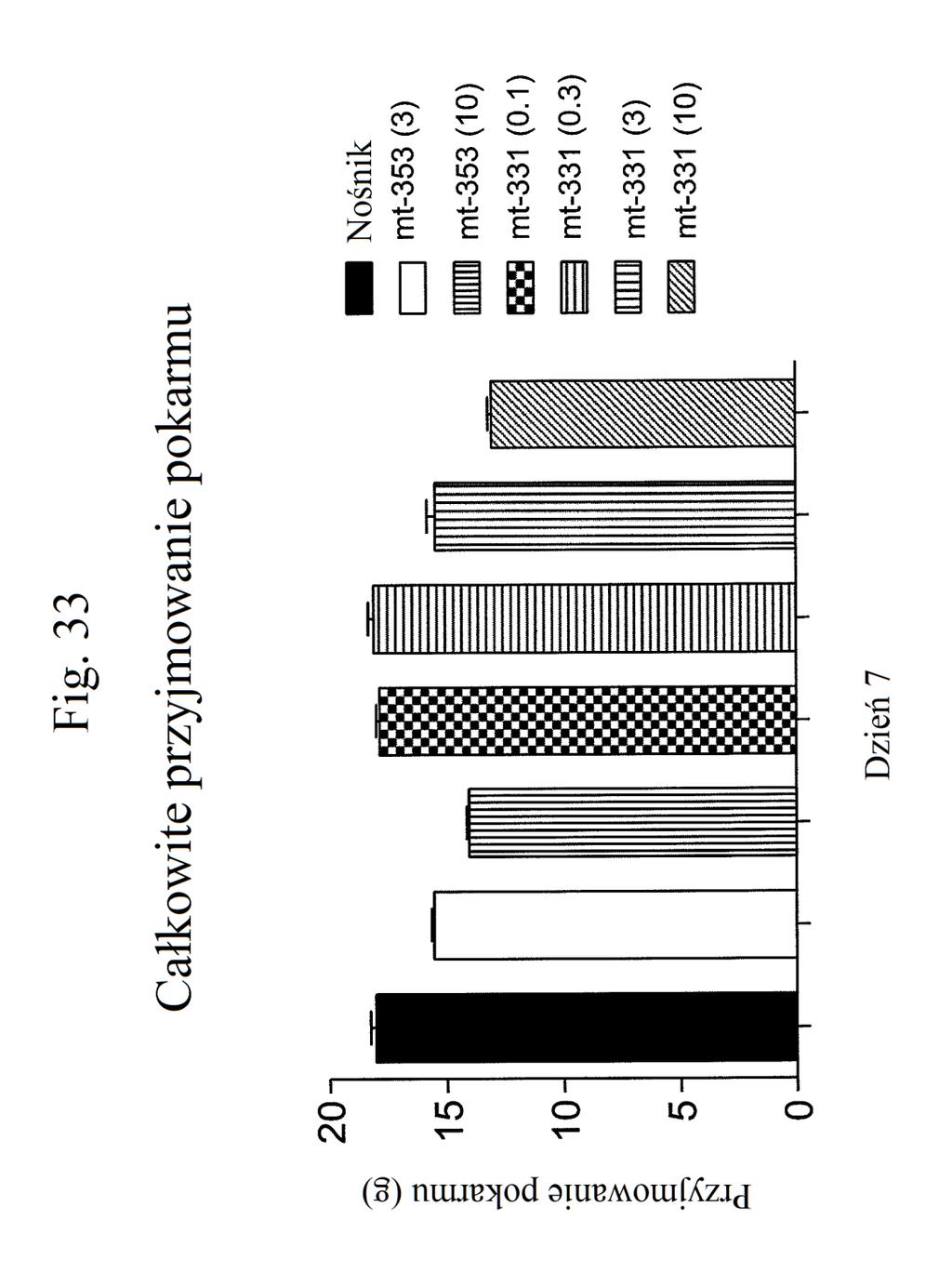
490 489
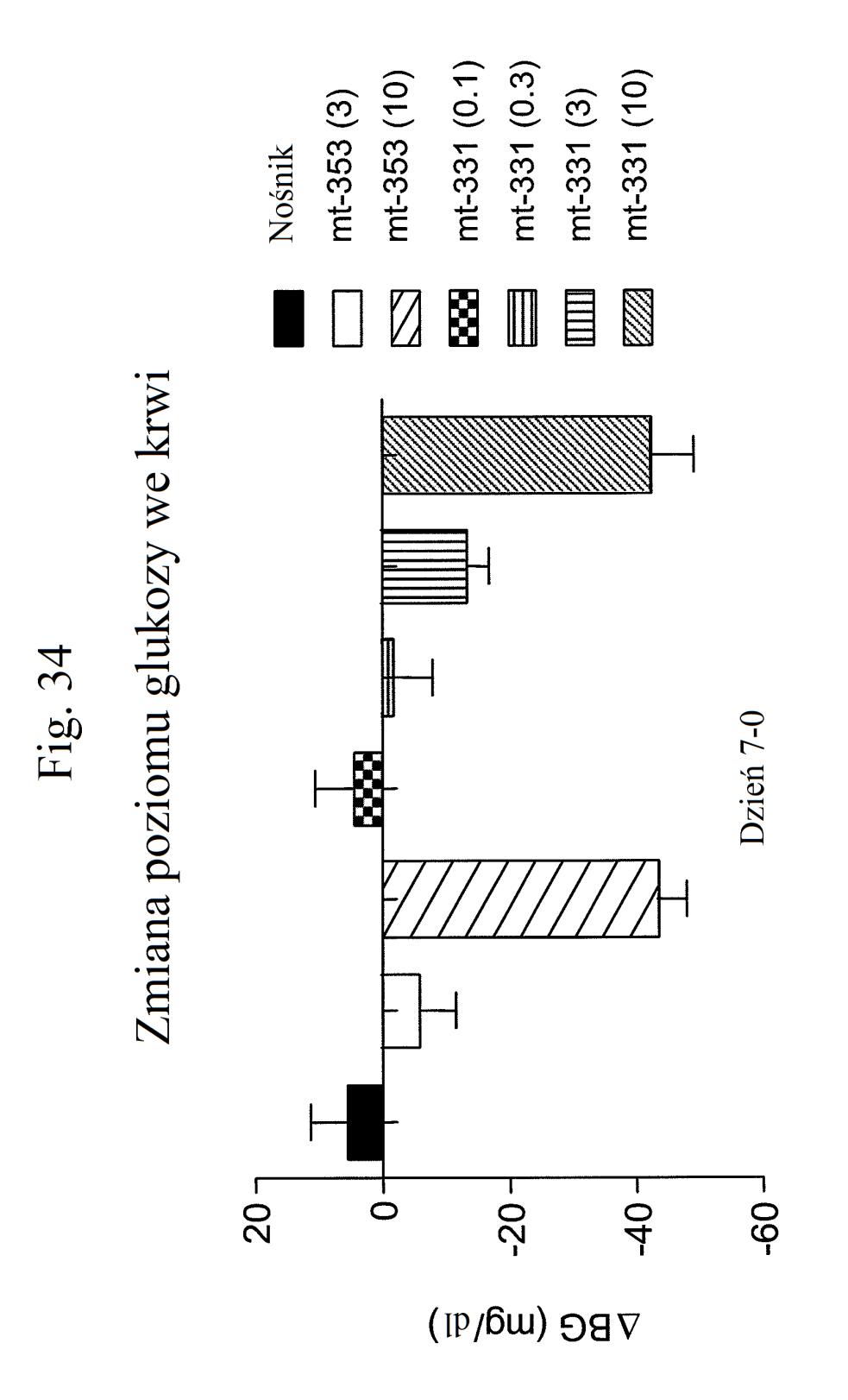
491 490
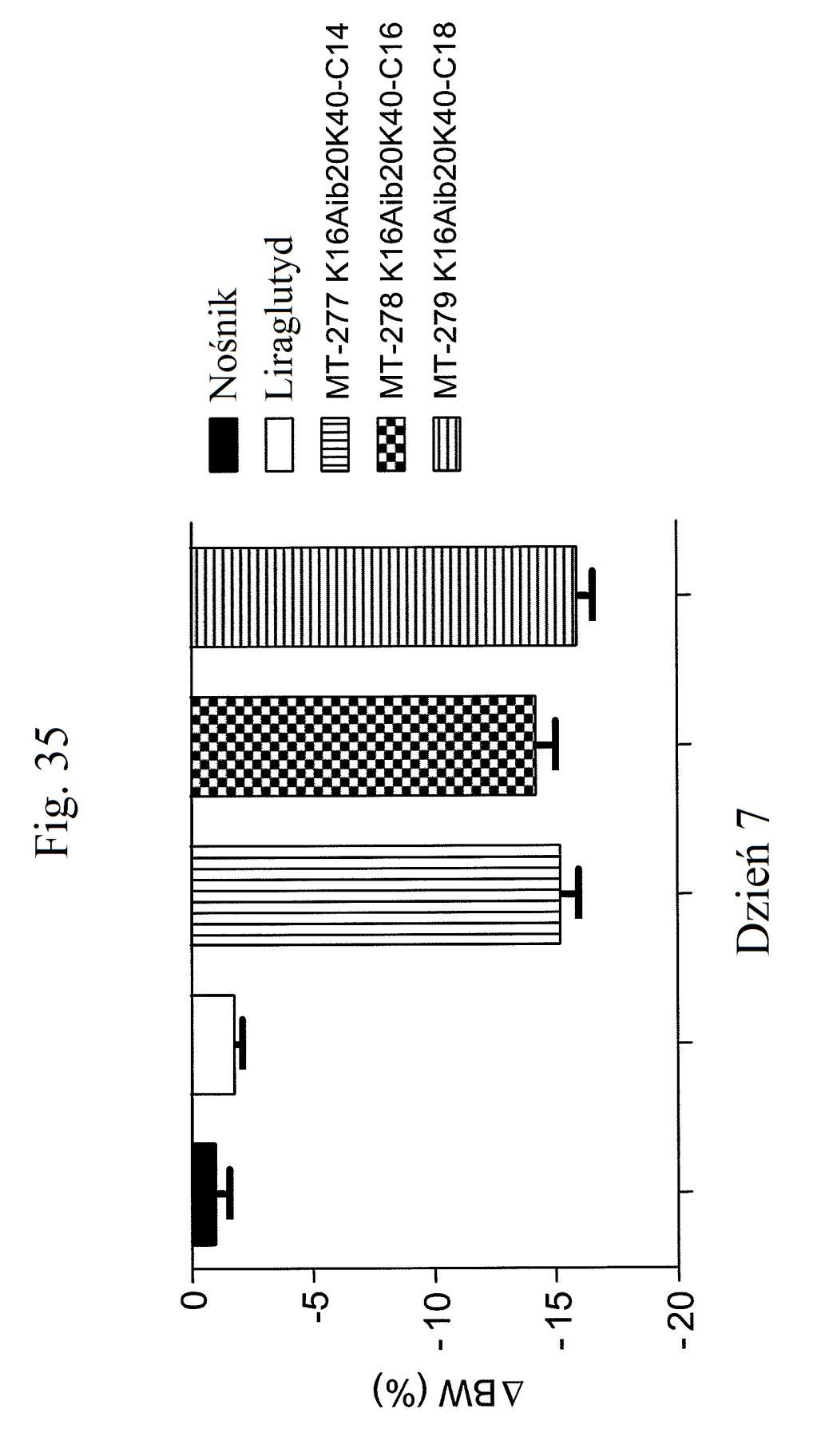
492 491
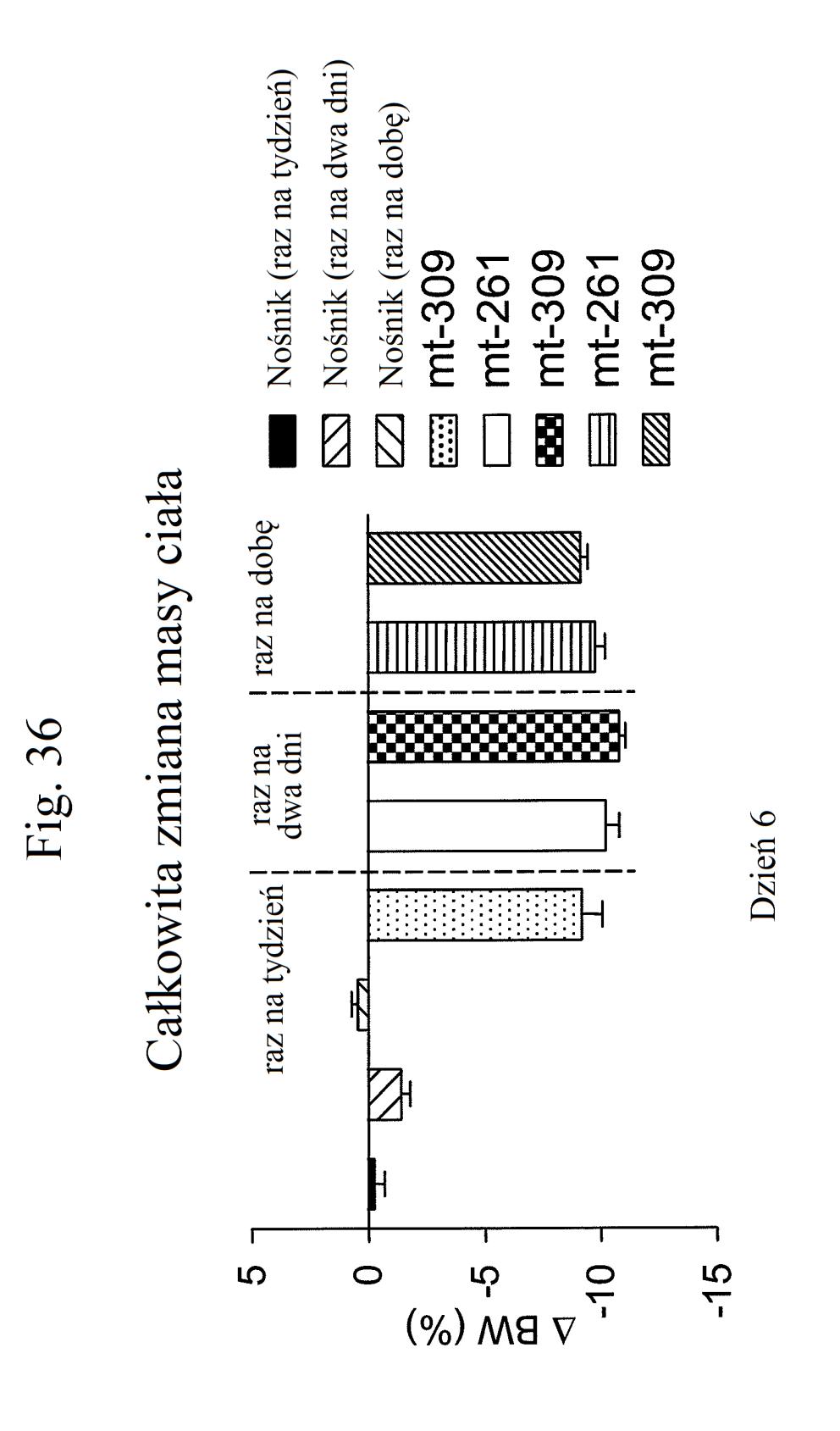
493 492
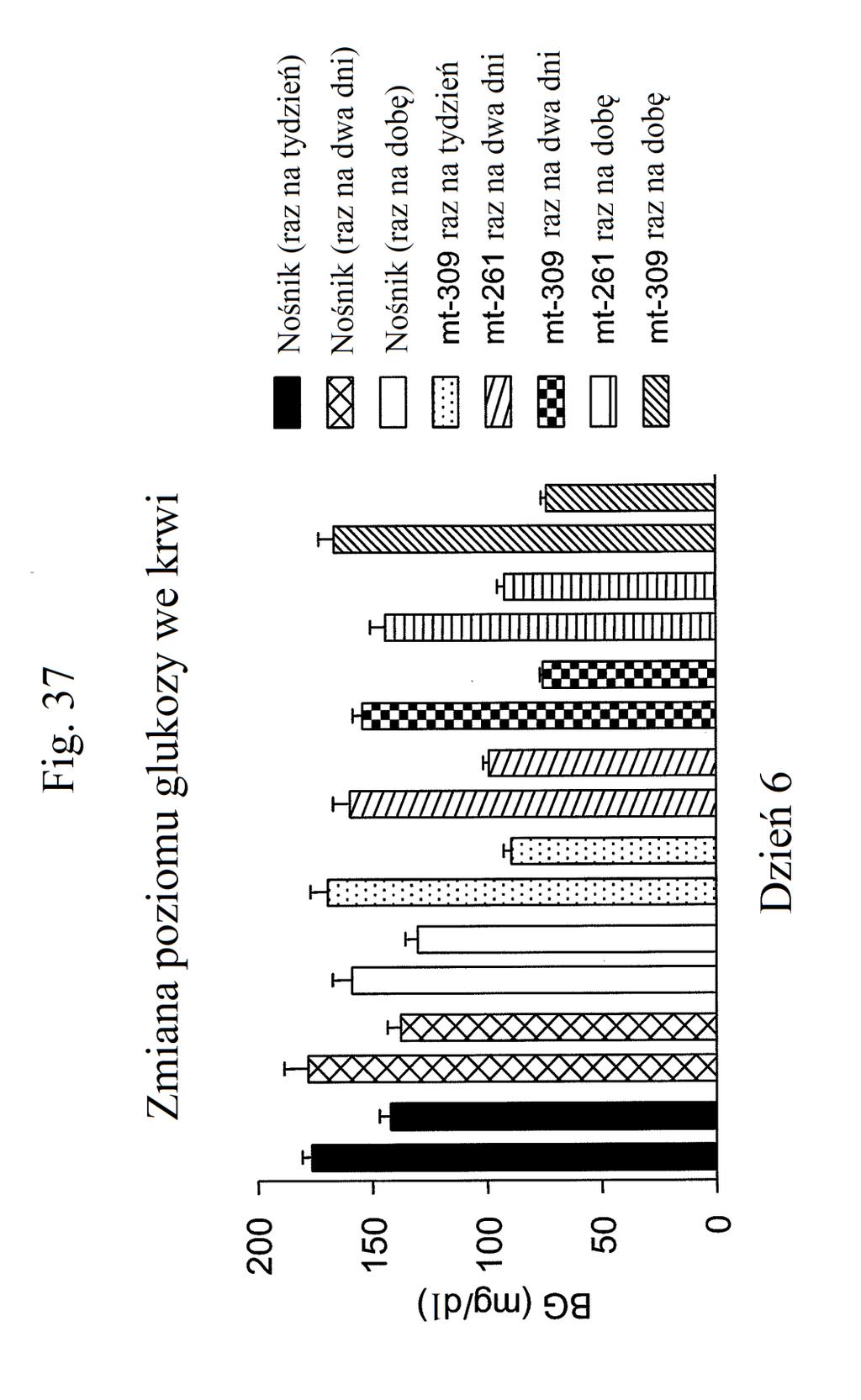
494 493
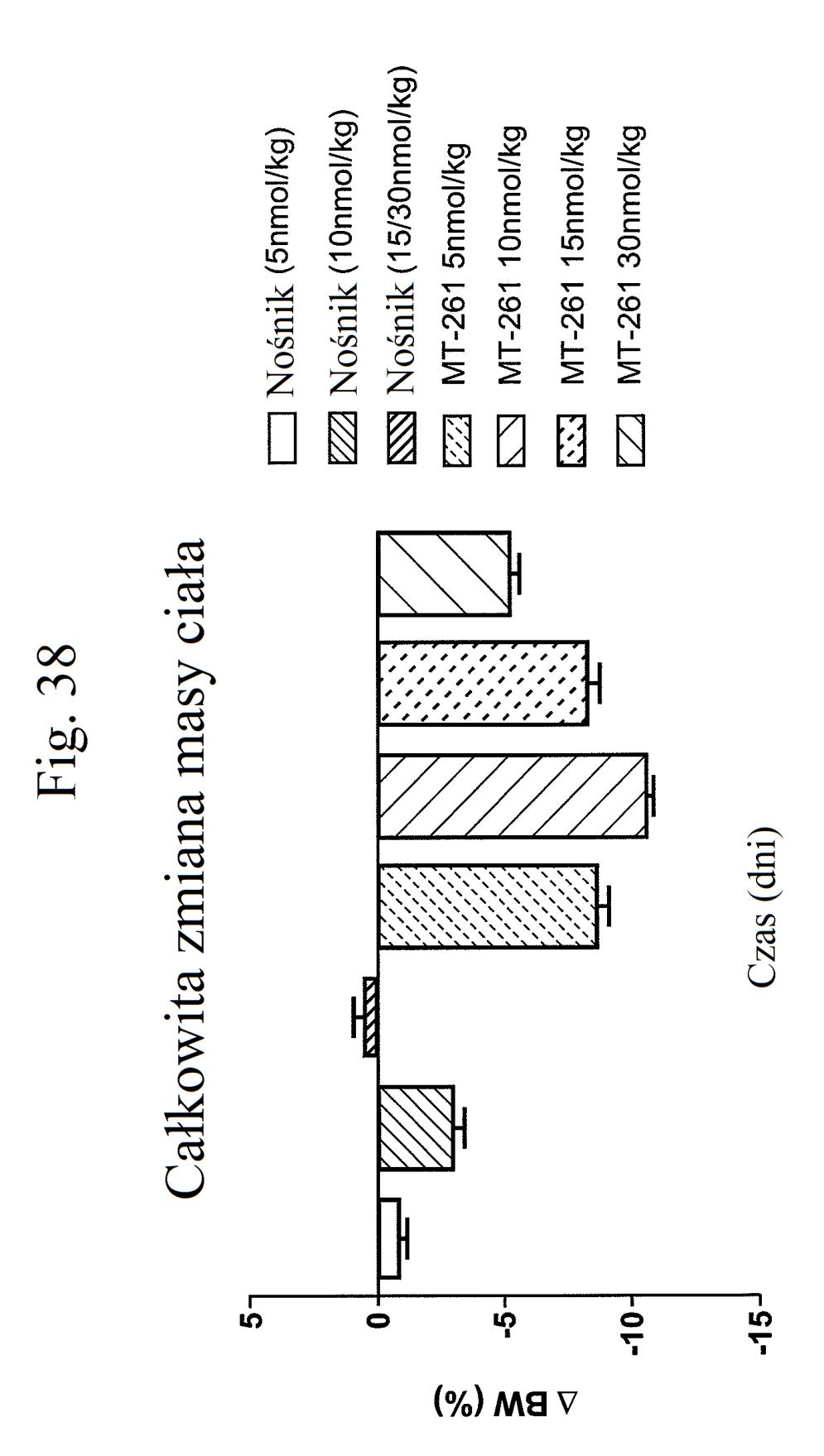
495 494
Chemiczne składniki komórek
 Chemiczne składniki komórek Pierwiastki chemiczne w komórkach: - makroelementy (pierwiastki biogenne) H, O, C, N, S, P Ca, Mg, K, Na, Cl >1% suchej masy - mikroelementy Fe, Cu, Mn, Mo, B, Zn, Co, J, F
Chemiczne składniki komórek Pierwiastki chemiczne w komórkach: - makroelementy (pierwiastki biogenne) H, O, C, N, S, P Ca, Mg, K, Na, Cl >1% suchej masy - mikroelementy Fe, Cu, Mn, Mo, B, Zn, Co, J, F
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1968711 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 05.01.2007 07712641.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1968711 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 05.01.2007 07712641.5
Przegląd budowy i funkcji białek
 Przegląd budowy i funkcji białek Co piszą o białkach? Wyraz wprowadzony przez Jönsa J. Berzeliusa w 1883 r. w celu podkreślenia znaczenia tej grupy związków. Termin pochodzi od greckiego słowa proteios,
Przegląd budowy i funkcji białek Co piszą o białkach? Wyraz wprowadzony przez Jönsa J. Berzeliusa w 1883 r. w celu podkreślenia znaczenia tej grupy związków. Termin pochodzi od greckiego słowa proteios,
21. Wstęp do chemii a-aminokwasów
 21. Wstęp do chemii a-aminokwasów Chemia rganiczna, dr hab. inż. Mariola Koszytkowska-Stawińska, WChem PW; 2016/2017 1 21.1. Budowa ogólna a-aminokwasów i klasyfikacja peptydów H 2 N H kwas 2-aminooctowy
21. Wstęp do chemii a-aminokwasów Chemia rganiczna, dr hab. inż. Mariola Koszytkowska-Stawińska, WChem PW; 2016/2017 1 21.1. Budowa ogólna a-aminokwasów i klasyfikacja peptydów H 2 N H kwas 2-aminooctowy
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680075 (13) T3 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2004
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1680075 (13) T3 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2004
Budowa aminokwasów i białek
 Biofizyka Ćwiczenia 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas Budowa aminokwasów i białek E.Banachowicz 1 Ogólna budowa aminokwasów α w neutralnym p α N 2 COO N
Biofizyka Ćwiczenia 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas Budowa aminokwasów i białek E.Banachowicz 1 Ogólna budowa aminokwasów α w neutralnym p α N 2 COO N
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1747298 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.7 (51) Int. Cl. C22C14/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1747298 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 02.05.2005 05747547.7 (51) Int. Cl. C22C14/00 (2006.01)
46 i 47. Wstęp do chemii -aminokwasów
 46 i 47. Wstęp do chemii -aminokwasów Chemia rganiczna, dr hab. inż. Mariola Koszytkowska-Stawińska, WChem PW; 2017/2018 1 21.1. Budowa ogólna -aminokwasów i klasyfikacja peptydów H 2 H H 2 R H R R 1 H
46 i 47. Wstęp do chemii -aminokwasów Chemia rganiczna, dr hab. inż. Mariola Koszytkowska-Stawińska, WChem PW; 2017/2018 1 21.1. Budowa ogólna -aminokwasów i klasyfikacja peptydów H 2 H H 2 R H R R 1 H
Informacje. W sprawach organizacyjnych Slajdy z wykładów
 Biochemia Informacje W sprawach organizacyjnych malgorzata.dutkiewicz@wum.edu.pl Slajdy z wykładów www.takao.pl W sprawach merytorycznych Takao Ishikawa (takao@biol.uw.edu.pl) Kiedy? Co? Kto? 24 lutego
Biochemia Informacje W sprawach organizacyjnych malgorzata.dutkiewicz@wum.edu.pl Slajdy z wykładów www.takao.pl W sprawach merytorycznych Takao Ishikawa (takao@biol.uw.edu.pl) Kiedy? Co? Kto? 24 lutego
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886669 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 02.08.2007 07113670.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886669 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 02.08.2007 07113670.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 213136 (96) Data i numer zgłoszenia patentu europejskiego: 14.03.2008 08723469.6 (13) (1) T3 Int.Cl. F24D 19/ (2006.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 213136 (96) Data i numer zgłoszenia patentu europejskiego: 14.03.2008 08723469.6 (13) (1) T3 Int.Cl. F24D 19/ (2006.01) Urząd
etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
 Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1799953 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.08.2005 05770398.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1799953 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.08.2005 05770398.5
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1787644 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 07.11.2006 06123574.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1787644 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 07.11.2006 06123574.3
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1773451 (96) Data i numer zgłoszenia patentu europejskiego: 08.06.2005 05761294.7 (13) (51) T3 Int.Cl. A61K 31/4745 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 8294 (96) Data i numer zgłoszenia patentu europejskiego: 2.01.08 08001421.0 (13) (1) T3 Int.Cl. B62D /04 (06.01) Urząd Patentowy
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 8294 (96) Data i numer zgłoszenia patentu europejskiego: 2.01.08 08001421.0 (13) (1) T3 Int.Cl. B62D /04 (06.01) Urząd Patentowy
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2300459. (96) Data i numer zgłoszenia patentu europejskiego: 22.06.2009 09772333.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2300459. (96) Data i numer zgłoszenia patentu europejskiego: 22.06.2009 09772333.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2049 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.06.09 09772333.2 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2049 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.06.09 09772333.2 (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 223771 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.08 0886773.1 (13) (1) T3 Int.Cl. A47L 1/42 (06.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 223771 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.08 0886773.1 (13) (1) T3 Int.Cl. A47L 1/42 (06.01) Urząd
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2326237 (96) Data i numer zgłoszenia patentu europejskiego: 07.07.2009 09780285.4 (13) (51) T3 Int.Cl. A47L 15/50 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2326237 (96) Data i numer zgłoszenia patentu europejskiego: 07.07.2009 09780285.4 (13) (51) T3 Int.Cl. A47L 15/50 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2210706 (96) Data i numer zgłoszenia patentu europejskiego: 21.01.2010 10000580.0 (13) (51) T3 Int.Cl. B24B 21/20 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2210706 (96) Data i numer zgłoszenia patentu europejskiego: 21.01.2010 10000580.0 (13) (51) T3 Int.Cl. B24B 21/20 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2003466 (96) Data i numer zgłoszenia patentu europejskiego: 12.06.2008 08460024.6 (13) (51) T3 Int.Cl. G01S 5/02 (2010.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2003466 (96) Data i numer zgłoszenia patentu europejskiego: 12.06.2008 08460024.6 (13) (51) T3 Int.Cl. G01S 5/02 (2010.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1505553. (96) Data i numer zgłoszenia patentu europejskiego: 05.08.2004 04018511.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1505553. (96) Data i numer zgłoszenia patentu europejskiego: 05.08.2004 04018511.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 3 (96) Data i numer zgłoszenia patentu europejskiego: 0.08.04 0401811.8 (13) (1) T3 Int.Cl. G08C 17/00 (06.01) Urząd Patentowy
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 3 (96) Data i numer zgłoszenia patentu europejskiego: 0.08.04 0401811.8 (13) (1) T3 Int.Cl. G08C 17/00 (06.01) Urząd Patentowy
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2122 (96) Data i numer zgłoszenia patentu europejskiego: 2..07 07866441.4 (13) (1) T3 Int.Cl. D21H 19/06 (06.01) Urząd Patentowy
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2122 (96) Data i numer zgłoszenia patentu europejskiego: 2..07 07866441.4 (13) (1) T3 Int.Cl. D21H 19/06 (06.01) Urząd Patentowy
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2259949 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2009 09727379.1 (13) (51) T3 Int.Cl. B60L 11/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2259949 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2009 09727379.1 (13) (51) T3 Int.Cl. B60L 11/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 221611 (96) Data i numer zgłoszenia patentu europejskiego: 19.01. 000481.1 (13) (1) T3 Int.Cl. B28C /42 (06.01) B60P 3/16
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 221611 (96) Data i numer zgłoszenia patentu europejskiego: 19.01. 000481.1 (13) (1) T3 Int.Cl. B28C /42 (06.01) B60P 3/16
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2179743 (96) Data i numer zgłoszenia patentu europejskiego: 13.07.2009 09460028.5 (13) (51) T3 Int.Cl. A61K 38/18 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2179743 (96) Data i numer zgłoszenia patentu europejskiego: 13.07.2009 09460028.5 (13) (51) T3 Int.Cl. A61K 38/18 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2190940 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.09.2008 08802024.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2190940 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.09.2008 08802024.3
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2555663 (96) Data i numer zgłoszenia patentu europejskiego: 06.04.2011 11730434.5 (13) (51) T3 Int.Cl. A47L 15/42 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2555663 (96) Data i numer zgłoszenia patentu europejskiego: 06.04.2011 11730434.5 (13) (51) T3 Int.Cl. A47L 15/42 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2084998 (96) Data i numer zgłoszenia patentu europejskiego: 29.01.2009 09151619.5 (13) (51) T3 Int.Cl. A47F 3/14 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2084998 (96) Data i numer zgłoszenia patentu europejskiego: 29.01.2009 09151619.5 (13) (51) T3 Int.Cl. A47F 3/14 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 161679 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 24.06.0 064.7 (1) Int. Cl. B60R21/01 (06.01) (97) O udzieleniu
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 161679 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 24.06.0 064.7 (1) Int. Cl. B60R21/01 (06.01) (97) O udzieleniu
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2047071 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.07.2007 07786251.4
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2047071 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21.07.2007 07786251.4
Leczenie cukrzycy typu 2- nowe możliwości
 Leczenie cukrzycy typu 2- nowe możliwości Dr n. med. Iwona Jakubowska Oddział Diabetologii, Endokrynologii i Chorób Wewnętrznych SP ZOZ Woj,. Szpital Zespolony Im. J. Śniadeckiego w Białymstoku DEFINICJA
Leczenie cukrzycy typu 2- nowe możliwości Dr n. med. Iwona Jakubowska Oddział Diabetologii, Endokrynologii i Chorób Wewnętrznych SP ZOZ Woj,. Szpital Zespolony Im. J. Śniadeckiego w Białymstoku DEFINICJA
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1591364 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.04.2005 05103299.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1591364 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.04.2005 05103299.3
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1810954 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.2006 06025226.9 (13) (51) T3 Int.Cl. C03B 9/41 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1810954 (96) Data i numer zgłoszenia patentu europejskiego: 06.12.2006 06025226.9 (13) (51) T3 Int.Cl. C03B 9/41 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2127498 (96) Data i numer zgłoszenia patentu europejskiego: 14.02.2008 08716843.1 (13) (51) T3 Int.Cl. H05B 41/288 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2127498 (96) Data i numer zgłoszenia patentu europejskiego: 14.02.2008 08716843.1 (13) (51) T3 Int.Cl. H05B 41/288 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2370461 (96) Data i numer zgłoszenia patentu europejskiego: 15.12.2008 08875672.1 (13) (51) T3 Int.Cl. C07K 14/605 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2370461 (96) Data i numer zgłoszenia patentu europejskiego: 15.12.2008 08875672.1 (13) (51) T3 Int.Cl. C07K 14/605 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") PL/EP 1699990 T3 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1699990 (96) Data i numer zgłoszenia patentu europejskiego: 09.11.2004 04800186.1 (13) (51) T3 Int.Cl. E04G
PL/EP 1699990 T3 RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1699990 (96) Data i numer zgłoszenia patentu europejskiego: 09.11.2004 04800186.1 (13) (51) T3 Int.Cl. E04G
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 28647 (96) Data i numer zgłoszenia patentu europejskiego: 30.03.09 091662.2 (13) (1) T3 Int.Cl. C07D 333/28 (06.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 28647 (96) Data i numer zgłoszenia patentu europejskiego: 30.03.09 091662.2 (13) (1) T3 Int.Cl. C07D 333/28 (06.01) Urząd
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 172874 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 04.0.2006 0611312. (1) Int. Cl. B23B31/28 (2006.01) (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 172874 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 04.0.2006 0611312. (1) Int. Cl. B23B31/28 (2006.01) (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1730054 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 22.03.2005 05731932.9 (51) Int. Cl. B65G17/06 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1730054 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 22.03.2005 05731932.9 (51) Int. Cl. B65G17/06 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1755549 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.04.2005 05780098.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1755549 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 22.04.2005 05780098.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1648484 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 01.06.04 047366.4 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1648484 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 01.06.04 047366.4 (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2321656 (96) Data i numer zgłoszenia patentu europejskiego:.08.09 09807498.2 (13) (51) T3 Int.Cl. G01R /18 (06.01) G01R 19/
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2321656 (96) Data i numer zgłoszenia patentu europejskiego:.08.09 09807498.2 (13) (51) T3 Int.Cl. G01R /18 (06.01) G01R 19/
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 182634 (96) Data i numer zgłoszenia patentu europejskiego: 19.04.07 070963.1 (13) T3 (1) Int. Cl. F16H/17 F16H7/04 (06.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 182634 (96) Data i numer zgłoszenia patentu europejskiego: 19.04.07 070963.1 (13) T3 (1) Int. Cl. F16H/17 F16H7/04 (06.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1624265 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2005 05106119.0 (13) T3 (51) Int. Cl. F25D23/06 F25D25/02
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1624265 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2005 05106119.0 (13) T3 (51) Int. Cl. F25D23/06 F25D25/02
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1999308 (96) Data i numer zgłoszenia patentu europejskiego: 28.03.2007 07727422.3 (13) (51) T3 Int.Cl. D06F 35/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1999308 (96) Data i numer zgłoszenia patentu europejskiego: 28.03.2007 07727422.3 (13) (51) T3 Int.Cl. D06F 35/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 240040 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 09.07. 007077.0 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 240040 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 09.07. 007077.0 (97)
(96) Data i numer zgłoszenia patentu europejskiego:
 Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1690978 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2005 05101042.9 (13) T3 (51) Int. Cl. D06F81/08 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1690978 (96) Data i numer zgłoszenia patentu europejskiego: 11.02.2005 05101042.9 (13) T3 (51) Int. Cl. D06F81/08 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2353894 (96) Data i numer zgłoszenia patentu europejskiego: 19.02.2010 10001703.7 (13) (51) T3 Int.Cl. B60D 5/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2353894 (96) Data i numer zgłoszenia patentu europejskiego: 19.02.2010 10001703.7 (13) (51) T3 Int.Cl. B60D 5/00 (2006.01)
Bioinformatyka. z sylabusu... (wykład monograficzny) wykład 1. E. Banachowicz. Wykład monograficzny Bioinformatyka.
 wykład 1. E. Banachowicz. Wykład monograficzny Bioinformatyka.") Bioinformatyka (wykład monograficzny) wykład 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas z sylabusu... Wykład 1, 2006 1 Co to jest Bioinformatyka? Zastosowanie technologii
Bioinformatyka (wykład monograficzny) wykład 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas z sylabusu... Wykład 1, 2006 1 Co to jest Bioinformatyka? Zastosowanie technologii
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 16234 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 18..0 0022716.4 (1) Int. Cl. B6D71/00 (06.01) (97) O udzieleniu
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 16234 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 18..0 0022716.4 (1) Int. Cl. B6D71/00 (06.01) (97) O udzieleniu
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1561894 (96) Data i numer zgłoszenia patentu europejskiego: 25.01.2005 05001385.3 (13) (51) T3 Int.Cl. E06B 3/66 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1561894 (96) Data i numer zgłoszenia patentu europejskiego: 25.01.2005 05001385.3 (13) (51) T3 Int.Cl. E06B 3/66 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1660738 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 28.04.2005 05737864.8 (51) Int. Cl. E04G1/32 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1660738 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 28.04.2005 05737864.8 (51) Int. Cl. E04G1/32 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1816307 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:.07.06 060114.3 (1) Int. Cl. E06B9/68 (06.01) (97) O udzieleniu
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1816307 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:.07.06 060114.3 (1) Int. Cl. E06B9/68 (06.01) (97) O udzieleniu
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1671552 (96) Data i numer zgłoszenia patentu europejskiego: 02.12.2005 05026319.3 (13) T3 (51) Int. Cl. A23L1/305 A23J3/16
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1671552 (96) Data i numer zgłoszenia patentu europejskiego: 02.12.2005 05026319.3 (13) T3 (51) Int. Cl. A23L1/305 A23J3/16
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1701111 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 11.03.2005 05090064.6 (51) Int. Cl. F24H9/20 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1701111 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 11.03.2005 05090064.6 (51) Int. Cl. F24H9/20 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1605897 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 19.03.2004 04722142.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1605897 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 19.03.2004 04722142.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1732433 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.01.2005 05702820.1
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1732433 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 27.01.2005 05702820.1
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1658064 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.6 (51) Int. Cl. A61K31/37 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1658064 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 06.07.2004 04740699.6 (51) Int. Cl. A61K31/37 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2328822 (96) Data i numer zgłoszenia patentu europejskiego: 02.09.2009 09782487.4 (13) (51) T3 Int.Cl. B65G 15/38 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2328822 (96) Data i numer zgłoszenia patentu europejskiego: 02.09.2009 09782487.4 (13) (51) T3 Int.Cl. B65G 15/38 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1614553 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 01.07.2005 05014326.2 (51) Int. Cl. B60C27/06 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1614553 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 01.07.2005 05014326.2 (51) Int. Cl. B60C27/06 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2528702 (96) Data i numer zgłoszenia patentu europejskiego: 03.12.2010 10796315.9 (13) (51) T3 Int.Cl. B21D 53/36 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2528702 (96) Data i numer zgłoszenia patentu europejskiego: 03.12.2010 10796315.9 (13) (51) T3 Int.Cl. B21D 53/36 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886585 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.07.2006 06291197.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1886585 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.07.2006 06291197.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1477128 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 14.05.2004 04076445.8 (51) Int. Cl. A61D1/02 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1477128 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 14.05.2004 04076445.8 (51) Int. Cl. A61D1/02 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1947302. (96) Data i numer zgłoszenia patentu europejskiego: 03.12.2007 07122193.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1947302. (96) Data i numer zgłoszenia patentu europejskiego: 03.12.2007 07122193.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1947302 (96) Data i numer zgłoszenia patentu europejskiego: 03.12.2007 07122193.1 (13) (51) T3 Int.Cl. F01M 11/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1947302 (96) Data i numer zgłoszenia patentu europejskiego: 03.12.2007 07122193.1 (13) (51) T3 Int.Cl. F01M 11/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1740398 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 1.03.200 071703.9 (1) Int. Cl. B60C1/06 (2006.01) (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1740398 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 1.03.200 071703.9 (1) Int. Cl. B60C1/06 (2006.01) (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609. (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609. (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609 (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.2 (13) (51) T3 Int.Cl. A61K 38/17 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1879609 (96) Data i numer zgłoszenia patentu europejskiego: 04.05.2006 06742792.2 (13) (51) T3 Int.Cl. A61K 38/17 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2480370 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 08.09.2010 10773557.3
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2480370 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 08.09.2010 10773557.3
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2776315 (96) Data i numer zgłoszenia patentu europejskiego: 19.07.2013 13753588.6 (13) (51) T4 Int.Cl. B64C 29/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2776315 (96) Data i numer zgłoszenia patentu europejskiego: 19.07.2013 13753588.6 (13) (51) T4 Int.Cl. B64C 29/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1854925 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2005 05826699.0 (13) (51) T3 Int.Cl. E03D 1/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1854925 (96) Data i numer zgłoszenia patentu europejskiego: 16.12.2005 05826699.0 (13) (51) T3 Int.Cl. E03D 1/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2828428 (96) Data i numer zgłoszenia patentu europejskiego: 12.03.13 13731877.0 (13) (1) T3 Int.Cl. D0B 19/12 (06.01) D0B
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2828428 (96) Data i numer zgłoszenia patentu europejskiego: 12.03.13 13731877.0 (13) (1) T3 Int.Cl. D0B 19/12 (06.01) D0B
Aminokwasy, peptydy i białka. Związki wielofunkcyjne
 Aminokwasy, peptydy i białka Związki wielofunkcyjne Aminokwasy, peptydy i białka Aminokwasy, peptydy i białka: - wiadomości ogólne Aminokwasy: - ogólna charakterystyka - budowa i nazewnictwo - właściwości
Aminokwasy, peptydy i białka Związki wielofunkcyjne Aminokwasy, peptydy i białka Aminokwasy, peptydy i białka: - wiadomości ogólne Aminokwasy: - ogólna charakterystyka - budowa i nazewnictwo - właściwości
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1659297 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 12.10.2005 05354036.5
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1659297 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 12.10.2005 05354036.5
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445326 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.10.2011 11186353.6
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445326 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.10.2011 11186353.6
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1890471 (96) Data i numer zgłoszenia patentu europejskiego: 19.10.2006 06791271.7 (13) (51) T3 Int.Cl. H04M 3/42 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1890471 (96) Data i numer zgłoszenia patentu europejskiego: 19.10.2006 06791271.7 (13) (51) T3 Int.Cl. H04M 3/42 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445186 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2011 11184611.9
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2445186 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.10.2011 11184611.9
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1668001 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.09.2004 04784968.2
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1668001 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 24.09.2004 04784968.2
Profil metaboliczny róŝnych organów ciała
 Profil metaboliczny róŝnych organów ciała Uwaga: tkanka tłuszczowa (adipose tissue) NIE wykorzystuje glicerolu do biosyntezy triacylogliceroli Endo-, para-, i autokrynna droga przekazu informacji biologicznej.
Profil metaboliczny róŝnych organów ciała Uwaga: tkanka tłuszczowa (adipose tissue) NIE wykorzystuje glicerolu do biosyntezy triacylogliceroli Endo-, para-, i autokrynna droga przekazu informacji biologicznej.
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1890558 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.05.2006 06755505.2
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1890558 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 18.05.2006 06755505.2
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1711158 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 16.11.2004 04806793.8
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1711158 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 16.11.2004 04806793.8
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2052830. (96) Data i numer zgłoszenia patentu europejskiego: 21.10.2008 08018365.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2052830. (96) Data i numer zgłoszenia patentu europejskiego: 21.10.2008 08018365.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 202830 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21..2008 0801836.0 (97)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 202830 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 21..2008 0801836.0 (97)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2638340 (96) Data i numer zgłoszenia patentu europejskiego: 10.11.2011 11781794.0 (13) (51) T3 Int.Cl. F25D 23/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2638340 (96) Data i numer zgłoszenia patentu europejskiego: 10.11.2011 11781794.0 (13) (51) T3 Int.Cl. F25D 23/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1449961 (96) Data i numer zgłoszenia patentu europejskiego: 14.04.2004 04405227.2 (13) T3 (51) Int. Cl. E01B9/14 F16B13/00
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1449961 (96) Data i numer zgłoszenia patentu europejskiego: 14.04.2004 04405227.2 (13) T3 (51) Int. Cl. E01B9/14 F16B13/00
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2086467 (96) Data i numer zgłoszenia patentu europejskiego: 26.11.2007 07824706.1 (13) (51) T3 Int.Cl. A61F 2/16 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2086467 (96) Data i numer zgłoszenia patentu europejskiego: 26.11.2007 07824706.1 (13) (51) T3 Int.Cl. A61F 2/16 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2074843. (96) Data i numer zgłoszenia patentu europejskiego: 27.09.2007 07818485.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2074843. (96) Data i numer zgłoszenia patentu europejskiego: 27.09.2007 07818485.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 74843 (96) Data i numer zgłoszenia patentu europejskiego: 27.09.07 0781848.0 (13) (1) T3 Int.Cl. H04W 4/12 (09.01) Urząd
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 74843 (96) Data i numer zgłoszenia patentu europejskiego: 27.09.07 0781848.0 (13) (1) T3 Int.Cl. H04W 4/12 (09.01) Urząd
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2057877 (96) Data i numer zgłoszenia patentu europejskiego: 03.11.2008 08019246.1 (13) (51) T3 Int.Cl. A01C 23/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2057877 (96) Data i numer zgłoszenia patentu europejskiego: 03.11.2008 08019246.1 (13) (51) T3 Int.Cl. A01C 23/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2143572 (96) Data i numer zgłoszenia patentu europejskiego: 18.05.2009 09160454.6 (13) (51) T3 Int.Cl. B60C 11/13 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2143572 (96) Data i numer zgłoszenia patentu europejskiego: 18.05.2009 09160454.6 (13) (51) T3 Int.Cl. B60C 11/13 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1912546. (96) Data i numer zgłoszenia patentu europejskiego: 11.08.2006 06761852.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1912546. (96) Data i numer zgłoszenia patentu europejskiego: 11.08.2006 06761852.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1912546 (96) Data i numer zgłoszenia patentu europejskiego: 11.08.2006 06761852.0 (13) (51) T3 Int.Cl. A47J 41/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1912546 (96) Data i numer zgłoszenia patentu europejskiego: 11.08.2006 06761852.0 (13) (51) T3 Int.Cl. A47J 41/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1663252 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 10.09.2004 04786930.0
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1663252 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 10.09.2004 04786930.0
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2224595 (96) Data i numer zgłoszenia patentu europejskiego: 10.02.2010 10001353.1 (13) (51) T3 Int.Cl. H03K 17/96 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2224595 (96) Data i numer zgłoszenia patentu europejskiego: 10.02.2010 10001353.1 (13) (51) T3 Int.Cl. H03K 17/96 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2383703 (96) Data i numer zgłoszenia patentu europejskiego: 29.04. 40068.1 (13) (1) T3 Int.Cl. G07B 1/06 (11.01) G08G 1/017
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2383703 (96) Data i numer zgłoszenia patentu europejskiego: 29.04. 40068.1 (13) (1) T3 Int.Cl. G07B 1/06 (11.01) G08G 1/017
Recenzja rozprawy doktorskiej mgr Karoliny Grabowskiej zatytułowanej Cykliczne peptydy o aktywności antyangiogennej
 Prof. dr hab. Zbigniew Szewczuk Wrocław, 2018-04-17 Wydział Chemii Uniwersytetu Wrocławskiego ul. F. Joliot-Curie 14, 50-383 Wrocław tel.: +48-71-3757212 e-mail: zbigniew.szewczuk@chem.uni.wroc.pl Recenzja
Prof. dr hab. Zbigniew Szewczuk Wrocław, 2018-04-17 Wydział Chemii Uniwersytetu Wrocławskiego ul. F. Joliot-Curie 14, 50-383 Wrocław tel.: +48-71-3757212 e-mail: zbigniew.szewczuk@chem.uni.wroc.pl Recenzja
Struktura i funkcja białek (I mgr)
") Struktura i funkcja białek (I mgr) Dr Filip Jeleń fj@protein.pl http://www.protein.pl/ Jeremy M. Berg, John L. Tymoczko, Lubert Stryer Biochemia Carl Branden, John Tooze Introduction to Protein Structure
Struktura i funkcja białek (I mgr) Dr Filip Jeleń fj@protein.pl http://www.protein.pl/ Jeremy M. Berg, John L. Tymoczko, Lubert Stryer Biochemia Carl Branden, John Tooze Introduction to Protein Structure
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1712702 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 28.03.2006 06006359.1 (51) Int. Cl. E04F15/02 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1712702 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 28.03.2006 06006359.1 (51) Int. Cl. E04F15/02 (2006.01)
Struktura biomakromolekuł chemia biologiczna III rok
 truktura biomakromolekuł chemia biologiczna III rok jak są zbudowane białka? dlaczego białka są tak zbudowane? co z tego wynika? 508 13 604 liczba struktur dostępnych w Protein Data Bank wynosi aktualnie
truktura biomakromolekuł chemia biologiczna III rok jak są zbudowane białka? dlaczego białka są tak zbudowane? co z tego wynika? 508 13 604 liczba struktur dostępnych w Protein Data Bank wynosi aktualnie
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1734922 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.03.2005 05728244.4
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1734922 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 11.03.2005 05728244.4
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) (13) T3 (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1661542 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 12.08.2004 04762070.3 (51) Int. Cl. A61G7/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 1661542 (13) T3 (96) Data i numer zgłoszenia patentu europejskiego: 12.08.2004 04762070.3 (51) Int. Cl. A61G7/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2744371 (96) Data i numer zgłoszenia patentu europejskiego: 07.09.2012 12777842.1 (13) (51) T3 Int.Cl. A47B 88/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2744371 (96) Data i numer zgłoszenia patentu europejskiego: 07.09.2012 12777842.1 (13) (51) T3 Int.Cl. A47B 88/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2468142. (96) Data i numer zgłoszenia patentu europejskiego: 21.12.2011 11194996.
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2468142. (96) Data i numer zgłoszenia patentu europejskiego: 21.12.2011 11194996.") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2468142 (96) Data i numer zgłoszenia patentu europejskiego: 21.12.2011 11194996.2 (13) (51) T3 Int.Cl. A47C 23/00 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2468142 (96) Data i numer zgłoszenia patentu europejskiego: 21.12.2011 11194996.2 (13) (51) T3 Int.Cl. A47C 23/00 (2006.01)
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2949485 (96) Data i numer zgłoszenia patentu europejskiego: 06.10.2014 14187774.6 (13) (51) T3 Int.Cl. B60C 23/04 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2949485 (96) Data i numer zgłoszenia patentu europejskiego: 06.10.2014 14187774.6 (13) (51) T3 Int.Cl. B60C 23/04 (2006.01)