O elektryczności neuronalnej i kanałach jonowych nieco więcej
|
|
|
- Krzysztof Sikora
- 8 lat temu
- Przeglądów:
Transkrypt
1 O elektryczności neuronalnej i kanałach jonowych nieco więcej
2 4.13 Molecular structure of the Na + /K + pump. (Part 1)

3 4.11 Ionic movements due to the Na + /K + pump. (Part 1) Np. przez dinitrofenol blokujący syntezę ATP Doświadczenia lat 50-tych z izotopem Na i blokowaniem ATP-azy ouabainą (g-strofantyna) Strophanthus gratus i Acokanthera ouabaio

4 4.12 Electrogenic transport of ions by the Na + /K + pump can influence membrane potential. Po silnym pobudzeniu neuronu/aksonu następuje stymulacja ATP-azy która w efekcie prowadzi do hyperpolaryzacji błony. Ouabaina zapobiega hyperpolaryzacji w takim eksperymencie.
5 Potencjał spoczynkowy różni się od potencjału równowagi dla potasu (jak również od pozostałych jonów!) E K = -102mV E Cl = -76mV E Na = 56mV E Ca = 125mV Różne populacje komórek w CSN mają różne wartości potencjału spoczynkowego. Mogą wartości zależeć od pory dnia, np. neurony wzgórza są hyperspolaryzowane w nocy zmniejszając w ten sposób impulsację dokorową.
6 Prąd jonowy Ruch określonych jonów poprzez błonę komórkową jest zależny od: różnicy potencjału spoczynkowego E m i potencjału równowagi dla określonego jonu np. dla potasu E k Przewodnictwa błony dla określonego jonu (odwrotność oporu elektrycznego zgodnie z prawem Ohma tj. i = V/R). Jeśli E m = E k prądu nie ma. Dla potasu: I k = g k (E m E k ) gdzie I k oznacza prąd jonowy potasu; g k oznacza przewodnictwo dla potasu (g = 1 Siemen gdy 1 volt powoduje przepływ 1 ampera)

7 Box A Voltage Clamp Method Kenneth Steward Cole Techniki opracowanej przez Kennetha S. Cole a tej używali w latach 50-tych Hodgkin i Huxley (nobel 1963)

8 3.5 Pharmacological separation of Na + and K + currents into components. Chlorek czteroetyloamoniowy doświadczenia z lat 60-tych

9 Wyniki badań Hodgkina i Huxley a nad przewodnictwem jonowym w zależności od napięcia błonowego w eksp. Voltage-clamp
 ze zmianą stężenia pozakomórkowego K Potencjał równowagi zachowuje się")
10 Potencjał spoczynkowy a stężenie jonów potasu Uwaga: stężenie wewnątrzkomórkowe potasu u kałamarnicy ok. 400 mm Hodkin i Katz w 1949 r eksperymenty (na aksonie kałamarnicy) ze zmianą stężenia pozakomórkowego K Potencjał równowagi zachowuje się prawie zgodnie z równaniem Nernsta
11 Dowód, że potencjał spoczynkowy określa głównie gradient stężenia potasu Czarna linia predykcja potencjału zgodnie z równaniem Nernsta Czerwona linia rzeczywiste dane pomiarowe potencjału Wykresy najbardziej się różnią dla niższych stężeń potasu (wpływ innych jonów)
12 Błona komórkowa jest znacznie bardziej przepuszczalna dla K niż dla innych jonów Wniosek: potas najbardziej wpływa na zachowanie potencjału błonowego

13 Rola sodu w gerneracji potencjału czynnościowego Wniosek Hodgkina i Katza : w czasie Pcz następuje gwałtowny wzrost przepuszczalności dla sodu
14 O wartości maks. potencjału czynnościowego decyduje stężenie pozakomórkowe sodu (limituje potencjał równowagi dla sodu) O wartości potencjału spoczynkowego decyduje stężenie potasu (wniosek: w potencjale spoczynkowym dominuje przewodnictwo potasowe)

15 Box B Action Potential Form and Nomenclature Pcz w różnych neuronach ma różny kształt ( waveform ) ale wnioski z eksperymentów na kałamarnicy zasadniczo obowiązują wszędzie A- squid, B akson motoneuronu żaby, C ciało motoneuronu żaby, D- neuron oliwki guinea pig, E- kom.purkinjego guinea pig Różne kształty Pcz są związane z dodatkowymi prądami (kanałami) jonowymi (Ca-, Kir)
16 Kanały jonowe
17 Techniki patch-clamp otworzyły drogę do poznania bardzo wielu kanałów jonowych o różnych własnościach Typy kanałów jonowych: 1) napięciowo-zależne 8) kanały przeciekowe (leakage channels) 2) aktywowane ligandem 3) aktywowane fizyczną zmianą kształtu (rozciąganiem) 4) zależne od temperatury (z rodziny TRP) 5) zależne od kwasowości (ph acid sensing ion channels (ASICs) 6) kanały zależne od cyklicznych nukleotydów (cgmp,camp) 7) aktywowane światłem (channelrhodopsin 1-2 algi)
 umożliwiła badanie przepływów jonowych (prądów) dla indywidualnych kanałów.")

18 Technika patch-clamp (Erwin Neher, Bert Sakmann 1976 Max Planck Inst. GoettingenNobel 1991) umożliwiła badanie przepływów jonowych (prądów) dla indywidualnych kanałów. Ostatecznie udowodniła istnienie kanałów jonowoselektywnych a jednocześnie potwierdziła wcześniejsze postulaty odnośnie istnienia takich kanałów proponowane przez Hodgkina i Huxley a Box A The Patch Clamp Method (Part 1)

19 Box A The Patch Clamp Method (Part 2)

20 4.4 Types of voltage-gated ion channels.
 dla kanałów jonowych Ponadto splicing oraz edycja RNA a także zmiany posttranslacyjne (fosforylacja) gwałtownie zwiększają")
21 Techniki patch-clamp: droga do poznania bardzo wielu kanałów jonowych o różnych własnościach W badaniach wykorzystuje się techniki genetyki molekularnej szczególnie na modelu oocytów żaby z rodzaju Xenopus Obecnie znamy ponad 100 genów (najwięcej dla K) dla kanałów jonowych Ponadto splicing oraz edycja RNA a także zmiany posttranslacyjne (fosforylacja) gwałtownie zwiększają różnorodność.
22 Równanie prądu jonowego obowiązuje dla pojedynczego kanału (np. potasowego) i s = g s (E m E k ) Przeciętny kanał jonowy posiada przewodność średnią ok. 20 ps (pikosiemensów czyli S) Dla gradientu napięcia 50mV i przewodności 20pS, prąd przechodzący przez pojedynczy kanał wynosi A (ampera) = ok. 6 mln jednowartościowych jonów/sek. (1A = ok. 6 x ładunków elektrycznych / sek)


23 4.1 Measurements of ionic currents flowing through single Na + channels. Prądy dośrodkowe Makroskopowy prąd sodowy jest sumą mikroskopowych prądów pojedynczych (napięciowozależnych) kanałów
 również są zsumowanymi prądami kanałów potasowych Zarówno kanał potasowy jak i")
24 4.2 Measurements of ionic currents flowing through single K + channels. Makroskopowe prądy potasowe dozewnątrz (outward) również są zsumowanymi prądami kanałów potasowych Zarówno kanał potasowy jak i sodowy muszą posiadać voltage sensor strukturę wyczuwającą napięcie
 Tetramer zbudowany z 4 podjednostek")
25 4.8 Structure of a simple bacterial K + channel determined by crystallography. (Part 1) Tetramer zbudowany z 4 podjednostek
 Ujemne ładunki helisy odwadniają jony K Przechodzą tylko nieuwodnione")
26 4.8 Structure of a simple bacterial K + channel determined by crystallography. (Part 2) Ujemne ładunki helisy odwadniają jony K Przechodzą tylko nieuwodnione jony K+
 Eksperymenty z patch-clamp wykazały")
27 UWAGA! Wykryto również napięciowozależne kanały Na które nie są inaktywowane depolaryzacją i prowadzące do długotrwających Pcz (blokowane przez lidokainę, benzokainę) Eksperymenty z patch-clamp wykazały podobieństwa i różnice pomiędzy różnymi kanałami jonowymi Podobieństwa kanałów K i Na: jonoselektywność, zależność prawdopodobieństwa otwarcia od napięcia, zamykanie kanałów Na i K przez hyperpolaryzację różnice kanałów K i Na: w kinetyce otwarcia (szybkość, czas otwarcia), depolaryzacja w kanale Na prowadzi oprócz otwarcia także do jego inaktywacji ale nie w przypadku kanału potasowego
28 Kanały potasowe są najbardziej zróżnicowane (różnią się np. czasem inaktywacji od milisekund do minut i innymi cechami) Kanały potasowe napięciowozależne Kanały potasowe wapniowo-zależne 1. Maxi K ps, blokowane przez charybdotoksynę (ze skorpiona) 2. Kanały o pośrednim przewodnictwie (18-80 ps) aktywowane obrzękiem komórek 3. małe kanały potasowe ps, blokowane przez toksynę pszczół apaminę Dośrodkowe prostowniki (inward rectifiers) M.in. wrażliwe na ATP blokowane przez glibenclamid i tolbutamid Kanały potasowe z podwójną domeną wewnętrznego otworu (2-pore domain=tandem pore domain, tzw. leak channels) (dimery!) M.in. kanały zależne od ph


29 4.6 Topology of principal subunits of voltage-gated, K +, 2-pore (tandem) zbudowany z 2 podj. Pozostałe z 4 podj.
30 Dwie zasadnicze podgrupy kanałów potasowych: 1. prostujące kanały odkomórkowe Rola w hyperpolaryzacji następczej występującej w przebiegu potencjału czynnościowego 2. dokomórkowe kanały prostujące (K-ir) K-ir grają istotną rolę w kontroli i regulacji potencjału spoczynkowego oraz wartości potencjału progowego.




31 Różne własności typów kanałów K +. Eksperymenty na oocytach X.laevis z ekspresją różnych typów kanałów K i pomiarami technika voltage-clamp Kir = Kanały K częściowo napięciowozależne
32 Potasowe dowewnątrz prostujące kanały jonowe (K-ir) Cechy: - osłabione przewodnictwo w warunkach depolaryzacji - podwyższone w warunkach hyperpolaryzacji - zdolność do wytwarzania większego dokomórkowego napływu jonów niż wypływu. - Blokowanie kierunku od (wypływu jonów potasowych) w warunkach depolaryzacji jest skutkiem działania wewnątrzkomórkowego magnezu (Mg2+) oraz polyamin (spermina, putrescyna, spermidyna).
33 dośrodkowe prostujące kanały potasowe pozwalają na dłuższe odpowiedzi depolaryzacyjne np. w sercu (gdzie potencjał czynnościowy trwa msec), grają rolę w tzw fertilization potential w komórkach jajowych (który trwa minuty). Zapobiegają utracie K + w czasie przedłużonej depolaryzacji i mogą pozwalać na re-entry K + T-tubul w mięśniach.
:. Kir4.")
34 Ekspresja Kir w ślimaku Immunohistochemiczna lokalizacja Kir4.1 w ślimaku szczura. (p-ciało anti-kir4.1):. Kir4.1 w zwoju spiralnym ślimaka, w stria vascularis, w komórkach Deiters a i komórkach kolumnowych (pillar cells) organu Cortiego (mała strzałka).
 Kanał Na zbudowany z 1 podj α (gen SCN1A) i 1-2 podj β (regulacyjne) - gen")
35 4.6 Topology of principal subunits of voltage-gated Na channels. (Part 1) Kanał Na zbudowany z 1 podj α (gen SCN1A) i 1-2 podj β (regulacyjne) - gen SCN1B Jest 9 typów podj α
36 Przykłady ( channelopathies ) Nie ma wytłumaczenia dlaczego określona mutacja kanału jonowego powoduje dane objawy Różne typy mutacji danym kanale jonowym różnią się także fenotypowo (objawami). Zwykle mutacje missense łagodniejsze od trunkacji. Istotne także miejsce mutacji missense. 1. Zaburzenia napięciowo-zależnych kanałów wapniowych (Ca): 1. Familial hemiplegic migraine (FHM) = (rodzinna migrena z hemiplegią) 2. Episodic ataxia type 2 (EA2) 3. X-linked congenital stationary night blindness (CSNB) 2. Defekt genu kanału sodowego (Na): SCN1A (dla α-podj) 1. Generalized epilepsy with febrile seizures plus (GEFS+) 2. Zesp. Dravet = severe myoclonic epilepsy of infancy (SMEI) w 95% mutacje SCN1A to nowe mutacje! Początek w 1-r.ż, spowolnienie rozwoju 3. Defekt genu kanału sodowego (Na): SCN1B (dla β-podj) 1. Generalized epilepsy with febrile seizures plus (GEFS+) 4. Defekt kanałów potasowych (K) 1. Benign familial neonatal convulsions (BFNC) objawy drgawek zanikają po okresie noworodkowym
37 Przykłady chorób spowodowanych nieprawidłowością działania kanałów 1. Mukowiscydoza: zmutowany kanał chlorkowy (epithelium chloride channel Cystic Fibrosis Transmembrane Conductance Regulator CFTR) 2. Zesp. Bartter a : Alkaloza, hipokaliemia, hyperaldosteronizm, hyperreninemia, bez nadciśnienia (brak odpowiedzi na aldosteron), zab. wzrastania, słabość mięśniowa, zaparcia, wielomocz.: mutacja napięciowozależnego kanału chlorkowego. 3. Myotonia congenita: Inna mutacja tego samego kanału (napięciowozależnego kanału chlorkowego). 4. Cholera: biegunka działanie toksyny bakteryjnej pobudzającej camp w nabłonku jelita co pobudza kanał CFTR i wywołuje sekrecję chloru do światła jelita. (w mukowiscydozie toksyna nie działa!). 5. Rodzinna hypoglikemia z hyperinsulinizmem; mutacja ATPzależnego kanału potasowego w komórkach beta trzustki powoduje, że kanał stymuluje wydzielanie insuliny 6. Zesp. Liddle a: wrodzone nadciśnienie: nadaktywność kanału sodowego w nabłonkach. 7. Zesp. Lamberta-Eatona (miasteniczny): przeciwciała przeciw kanałom Ca

38 Aktywowane ligandem kanały jonowe nie są zazwyczaj tak wysokoselektywne dla określonych jonów jak kanały napięciowo-zależne Oprócz błony cytoplazmatycznej obecne są w błonach organelli wewnątrzkomórkowych Kanały K aktywowane wapniem, kanały aktywowane cyklicznymi nukleotydami oraz ASICs mają wewnątrzkomórkowe domeny detekcji ligandu (Kanały aktywowane cykl. nukleotydami graja rolę m.in. w węchu i recepcji światła)!
39 Technika patch-clamp w badaniu kanałów jonowych aktywowanych ligandem A) Eksperyment typu outside-out pipeta zawiera roztwór o składzie podobnym do cytoplazmy. Na zewnątrz błony z kanałem roztwór jest podobny do zewnątrzkomórkowego. Mierzone jest natężenie prądu. Natomiast woltaż jest stabilizowany na dowolnie wybranej wartości
40 Technika patch-clamp B) Przepływ prądu płynącego przez pojedynczy kanał jonowy po podaniu od zewnątrz acetylocholiny (Ach) w sposób ciągły. Prąd PŁYNIE W POSTACI IMPULSÓW.( wszystko-albo-nic ). Zwiększenie stężenia Ach nie powoduje zmiany natężenia prądu lecz wzrost PRAWDOPODOBIEŃSTWA otwarcia kanału! Czas otwarcia jest różny ale amplituda (natężenie prądu) zawsze ta sama. Efekt postsynaptyczny jest wynikiem sumowania potencjałów z wielu kanałów jonowych.
41 Technika patch-clamp z kanałem receptorowym: podawanie N-T przy zmienianych wartościach potencjału stabilizowanego napięcia
42 Technika patch-clamp kanałem receptorowym: podawanie N-T przy zmienianych wartościach potencjału stabilizowanego napięcia oraz zmienianych stężeń jonów: wnioski: 1) Po związaniu z NT częstotliwość i średni czas otwarcia kanału są niezależne od napięcia 2) Kierunek i amplituda prądu zależy od napięcia. 3) Kierunek prądu dąży do osiągnięcia równowagi zgodnie z równaniem Goldmana-Hodgkina-Katza. 4) Testowany kanał jest tak samo przepuszczalny dla K+ i Na+ (ale różne kanały odpowiedzialne za wytwarzanie EPSP są różnie przepuszczalne dla K+ i dla Na+.) I sc = sc (V m E r ) Single channel reversal (null) potential receptora, w tym przyp.= 0 (Są one różne dla różnych receptorów) Prąd płynie na zewnątrz
43 Kablowe właściwości neuronów i ich wypustek

44 3.10 Passive current flow in an axon. (Part 1)
45 Elektrotonus, potencjały elektrotoniczne Potencjały podprogowe: rozprzestrzenianie biernie (elektrotoniczne) osłabienie: a) z odległością od miejsca ich powstania b) z upływem czasu. Własności elektrotoniczne neuronów nazywane są też właściwościami kablowymi
46 Kablowe właściwości neuronu Równanie kablowe: opis relacji (zmian) napięcia wzdłuż (modelowej) wypustki neuronu do odległości i czasu jego rozwiązanie: V x = V 0 e x/ V t = V 0 e t/ = tzw. stałą długości (length constant, space constant, characteristic length) = odległość od punktu 0, w której napięcie zmniejszy się do 37% wartości w punkcie 0.
47 Box C Passive Membrane Properties (Part 1) V x = V 0 e x/
 jest większa w aksonie zmielinizowanym i dlatego lokalne prądy operują na dalszym")
48 Electrotoniczny zanik napięcia wzdłuż niezmielinizowanego (schemat górny) i zmielinizowanego (schemat dolny). Stała długości (Length constant - λ) jest większa w aksonie zmielinizowanym i dlatego lokalne prądy operują na dalszym dystansie wzdłuż aksonu.
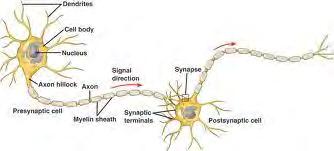
49
O elektryczności neuronalnej i kanałach jonowych nieco więcej
 O elektryczności neuronalnej i kanałach jonowych nieco więcej 4.13 Molecular structure of the Na + /K + pump. (Part 1) 4.11 Ionic movements due to the Na + /K + pump. (Part 1) Np. przez dinitrofenol blokujący
O elektryczności neuronalnej i kanałach jonowych nieco więcej 4.13 Molecular structure of the Na + /K + pump. (Part 1) 4.11 Ionic movements due to the Na + /K + pump. (Part 1) Np. przez dinitrofenol blokujący
O elektryczności neuronalnej i kanałach jonowych nieco więcej
 O elektryczności neuronalnej i kanałach jonowych nieco więcej 4.13 Molecular structure of the Na + /K + pump. (Part 1) 4.11 Ionic movements due to the Na + /K + pump. (Part 1) Np. przez dinitrofenol blokujący
O elektryczności neuronalnej i kanałach jonowych nieco więcej 4.13 Molecular structure of the Na + /K + pump. (Part 1) 4.11 Ionic movements due to the Na + /K + pump. (Part 1) Np. przez dinitrofenol blokujący
Potencjał spoczynkowy i czynnościowy
 Potencjał spoczynkowy i czynnościowy Marcin Koculak Biologiczne mechanizmy zachowania https://backyardbrains.com/ Powtórka budowy komórki 2 Istota prądu Prąd jest uporządkowanym ruchem cząstek posiadających
Potencjał spoczynkowy i czynnościowy Marcin Koculak Biologiczne mechanizmy zachowania https://backyardbrains.com/ Powtórka budowy komórki 2 Istota prądu Prąd jest uporządkowanym ruchem cząstek posiadających
Budowa i zróżnicowanie neuronów - elektrofizjologia neuronu
 Budowa i zróżnicowanie neuronów - elektrofizjologia neuronu Neuron jest podstawową jednostką przetwarzania informacji w mózgu. Sygnał biegnie w nim w kierunku od dendrytów, poprzez akson, do synaps. Neuron
Budowa i zróżnicowanie neuronów - elektrofizjologia neuronu Neuron jest podstawową jednostką przetwarzania informacji w mózgu. Sygnał biegnie w nim w kierunku od dendrytów, poprzez akson, do synaps. Neuron
Elektrofizjologia neuronu
 Spis treści Co to jest neuron? 2008-11-13 Spis treści Co to jest neuron? Wstęp Rola jonów w działaniu neronu Potencjał membranowy Stan równowagi Bramki jonowe Dynamika bramek jonowych Model Hodgkina-Huxley
Spis treści Co to jest neuron? 2008-11-13 Spis treści Co to jest neuron? Wstęp Rola jonów w działaniu neronu Potencjał membranowy Stan równowagi Bramki jonowe Dynamika bramek jonowych Model Hodgkina-Huxley
Z47 BADANIA WŁAŚCIWOŚCI ELEKTROFIZJOLOGICZNYCH BŁON KOMÓRKOWYCH
 Z47 BADANIA WŁAŚCIWOŚCI ELEKTROFIZJOLOGICZNYCH BŁON KOMÓRKOWYCH I. Cel ćwiczenia Celem ćwiczenia jest zapoznanie się z podstawową wiedzą na temat pomiarów elektrofizjologicznych żywych komórek metodą Patch
Z47 BADANIA WŁAŚCIWOŚCI ELEKTROFIZJOLOGICZNYCH BŁON KOMÓRKOWYCH I. Cel ćwiczenia Celem ćwiczenia jest zapoznanie się z podstawową wiedzą na temat pomiarów elektrofizjologicznych żywych komórek metodą Patch
Błona komórkowa grubość od 50 do 100 A. Istnieje pewna różnica potencjałów, po obu stronach błony, czyli na błonie panuje pewne
 Błona komórkowa grubość od 50 do 100 A Istnieje pewna różnica potencjałów, po obu stronach błony, czyli na błonie panuje pewne napięcie elektryczne, zwane napięciem na błonie. Różnica potencjałów to ok.
Błona komórkowa grubość od 50 do 100 A Istnieje pewna różnica potencjałów, po obu stronach błony, czyli na błonie panuje pewne napięcie elektryczne, zwane napięciem na błonie. Różnica potencjałów to ok.
Droga impulsu nerwowego w organizmie człowieka
 Droga impulsu nerwowego w organizmie człowieka Impuls nerwowy Impuls nerwowy jest zjawiskiem elektrycznym zachodzącym na powierzchni komórki nerwowej i pełni podstawową rolę w przekazywaniu informacji
Droga impulsu nerwowego w organizmie człowieka Impuls nerwowy Impuls nerwowy jest zjawiskiem elektrycznym zachodzącym na powierzchni komórki nerwowej i pełni podstawową rolę w przekazywaniu informacji
Praktyczne aspekty modelowania układu nerwowego
 Praktyczne aspekty modelowania układu nerwowego Ćwiczenia 2 Model Hodgkina-Huxleya dr Daniel Wójcik na podstawie The Book of GENESIS Wprowadzenie do interfejsu graficznego GENESIS Przećwiczymy obsługę
Praktyczne aspekty modelowania układu nerwowego Ćwiczenia 2 Model Hodgkina-Huxleya dr Daniel Wójcik na podstawie The Book of GENESIS Wprowadzenie do interfejsu graficznego GENESIS Przećwiczymy obsługę
Właściwości błony komórkowej
 Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność Transport przez błony Współczynnik przepuszczalności [cm/s] RóŜnice składu jonowego między wnętrzem komórki ssaka a otoczeniem
Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność Transport przez błony Współczynnik przepuszczalności [cm/s] RóŜnice składu jonowego między wnętrzem komórki ssaka a otoczeniem
Transportowane cząsteczki CO O, 2, NO, H O, etanol, mocznik... Zgodnie z gradientem: stężenia elektrochemicznym gradient stężeń
 Transportowane cząsteczki Transport przez błony Transport bierny szybkość transportu gradien t stężeń kanał nośnik Transport z udziałem nośnika: dyfuzja prosta dyfuzja prosta CO 2, O 2, NO,, H 2 O, etanol,
Transportowane cząsteczki Transport przez błony Transport bierny szybkość transportu gradien t stężeń kanał nośnik Transport z udziałem nośnika: dyfuzja prosta dyfuzja prosta CO 2, O 2, NO,, H 2 O, etanol,
Błona komórkowa grubość od 50 do 100 A. Istnieje pewna różnica potencjałów, po obu stronach błony, czyli na błonie panuje pewne
 Błona komórkowa grubość od 50 do 100 A Istnieje pewna różnica potencjałów, po obu stronach błony, czyli na błonie panuje pewne napięcie elektryczne, zwane napięciem na błonie. Różnica potencjałów to ok.
Błona komórkowa grubość od 50 do 100 A Istnieje pewna różnica potencjałów, po obu stronach błony, czyli na błonie panuje pewne napięcie elektryczne, zwane napięciem na błonie. Różnica potencjałów to ok.
Wstęp do sieci neuronowych, wykład 15, Neuron Hodgkina-Huxleya
 Wstęp do sieci neuronowych, wykład 15, Neuron Hodgkina-Huxleya Maja Czoków, Jarosław Piersa, Andrzej Rutkowski Wydział Matematyki i Informatyki, Uniwersytet Mikołaja Kopernika 2019-01-21 Projekt pn. Wzmocnienie
Wstęp do sieci neuronowych, wykład 15, Neuron Hodgkina-Huxleya Maja Czoków, Jarosław Piersa, Andrzej Rutkowski Wydział Matematyki i Informatyki, Uniwersytet Mikołaja Kopernika 2019-01-21 Projekt pn. Wzmocnienie
c stężenie molowe, V średnia prędkość molekuł
 Elektrodyfuzja, prąd jonowy i biopotencjały elektryczne.. Zjawiska elektryczne towarzyszące dyfuzji jonów oraz różnice ich stężeń powodują, że potencjały elektryczne roztworów po obu stronach błony są
Elektrodyfuzja, prąd jonowy i biopotencjały elektryczne.. Zjawiska elektryczne towarzyszące dyfuzji jonów oraz różnice ich stężeń powodują, że potencjały elektryczne roztworów po obu stronach błony są
Transport przez błony
 Transport przez błony Transport bierny Nie wymaga nakładu energii Transport aktywny Wymaga nakładu energii Dyfuzja prosta Dyfuzja ułatwiona Przenośniki Kanały jonowe Transport przez pory w błonie jądrowej
Transport przez błony Transport bierny Nie wymaga nakładu energii Transport aktywny Wymaga nakładu energii Dyfuzja prosta Dyfuzja ułatwiona Przenośniki Kanały jonowe Transport przez pory w błonie jądrowej
Biologiczne mechanizmy zachowania
 Biologiczne mechanizmy zachowania Przekaźnictwo chemiczne w mózgu mgr Monika Mazurek IPs UJ Odkrycie synaps Ramon y Cajal (koniec XIX wieku) neurony nie łączą się między sobą, między nimi jest drobna szczelina.
Biologiczne mechanizmy zachowania Przekaźnictwo chemiczne w mózgu mgr Monika Mazurek IPs UJ Odkrycie synaps Ramon y Cajal (koniec XIX wieku) neurony nie łączą się między sobą, między nimi jest drobna szczelina.
Kanały jonowe i pompy błonowe
 Kanały jonowe i pompy błonowe Jak badad przepływ jonów? Patch-clamp -zassanie powoduje ścisłe połączenie błony komórkowej z kapilarą (opornośd miedzy wnętrzem pipety a otaczającym roztworem = 10^9 omów)
Kanały jonowe i pompy błonowe Jak badad przepływ jonów? Patch-clamp -zassanie powoduje ścisłe połączenie błony komórkowej z kapilarą (opornośd miedzy wnętrzem pipety a otaczającym roztworem = 10^9 omów)
Tkanka nerwowa. Komórki: komórki nerwowe (neurony) sygnalizacja komórki neurogleju (glejowe) ochrona, wspomaganie
 sygnalizacja komórki neurogleju (glejowe) ochrona, wspomaganie") Komórki: komórki nerwowe (neurony) sygnalizacja komórki neurogleju (glejowe) ochrona, wspomaganie Tkanka nerwowa Substancja międzykomórkowa: prawie nieobecna (blaszki podstawne) pobudliwość przewodnictwo
Komórki: komórki nerwowe (neurony) sygnalizacja komórki neurogleju (glejowe) ochrona, wspomaganie Tkanka nerwowa Substancja międzykomórkowa: prawie nieobecna (blaszki podstawne) pobudliwość przewodnictwo
Właściwości błony komórkowej
 Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność szybka dyfuzja: O 2, CO 2, N 2, benzen Dwuwarstwa lipidowa - przepuszczalność Współczynnik przepuszczalności [cm/s] 1 Transport
Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność szybka dyfuzja: O 2, CO 2, N 2, benzen Dwuwarstwa lipidowa - przepuszczalność Współczynnik przepuszczalności [cm/s] 1 Transport
Właściwości błony komórkowej
 Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność Transport przez błony Cząsteczki < 150Da Błony - selektywnie przepuszczalne RóŜnice składu jonowego między wnętrzem komórki ssaka
Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność Transport przez błony Cząsteczki < 150Da Błony - selektywnie przepuszczalne RóŜnice składu jonowego między wnętrzem komórki ssaka
Krwiobieg duży. Krwiobieg mały
 Mięsień sercowy Budowa serca Krązenie krwi Krwiobieg duży Krew (bogata w tlen) wypływa z lewej komory serca przez zastawkę aortalną do głównej tętnicy ciała, aorty, rozgałęzia się na mniejsze tętnice,
Mięsień sercowy Budowa serca Krązenie krwi Krwiobieg duży Krew (bogata w tlen) wypływa z lewej komory serca przez zastawkę aortalną do głównej tętnicy ciała, aorty, rozgałęzia się na mniejsze tętnice,
Wprowadzenie do pulsujących sieci neuronowych
 Zakład Algebry i Kombinatoryki Wydział Matematyki i Nauk Informacyjnych 31 maja 2017 Wstęp Plan prezentacji Biologiczna inspiracja modeli neuronów. Modelowe neuronów naturalnych. Neurony trzeciej generacji
Zakład Algebry i Kombinatoryki Wydział Matematyki i Nauk Informacyjnych 31 maja 2017 Wstęp Plan prezentacji Biologiczna inspiracja modeli neuronów. Modelowe neuronów naturalnych. Neurony trzeciej generacji
Czynności komórek nerwowych. Adriana Schetz IF US
 Czynności komórek nerwowych Adriana Schetz IF US Plan wykładu 1. Komunikacja mędzykomórkowa 2. Neurony i komórki glejowe jedność architektoniczna 3. Czynności komórek nerwowych Komunikacja międzykomórkowa
Czynności komórek nerwowych Adriana Schetz IF US Plan wykładu 1. Komunikacja mędzykomórkowa 2. Neurony i komórki glejowe jedność architektoniczna 3. Czynności komórek nerwowych Komunikacja międzykomórkowa
Właściwości błony komórkowej
 Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność Glikokaliks glikokaliks cytoplazma jądro błona komórkowa Mikrografia elektronowa powierzchni limfocytu ludzkiego (wybarwienie
Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność Glikokaliks glikokaliks cytoplazma jądro błona komórkowa Mikrografia elektronowa powierzchni limfocytu ludzkiego (wybarwienie
Laboratorium Fizjologii
 Laboratorium Fizjologii Wstęp do neurofizjologii - potencjał czynnościowy i połączenia synaptyczne 1 Wstęp Potencjał równowagi Jeśli po dwu stronach przegrody przepuszczalnej dla jonów wytworzyć różnicę
Laboratorium Fizjologii Wstęp do neurofizjologii - potencjał czynnościowy i połączenia synaptyczne 1 Wstęp Potencjał równowagi Jeśli po dwu stronach przegrody przepuszczalnej dla jonów wytworzyć różnicę
Fizjologia człowieka
 Akademia Wychowania Fizycznego i Sportu w Gdańsku Katedra: Promocji Zdrowia Zakład: Biomedycznych Podstaw Zdrowia Fizjologia człowieka Osoby prowadzące przedmiot: Prof. nadzw. dr hab. Zbigniew Jastrzębski
Akademia Wychowania Fizycznego i Sportu w Gdańsku Katedra: Promocji Zdrowia Zakład: Biomedycznych Podstaw Zdrowia Fizjologia człowieka Osoby prowadzące przedmiot: Prof. nadzw. dr hab. Zbigniew Jastrzębski
Tkanka nerwowa. neurony (pobudliwe) odbieranie i przekazywanie sygnałów komórki glejowe (wspomagające)
 odbieranie i przekazywanie sygnałów komórki glejowe (wspomagające)") Tkanka nerwowa neurony (pobudliwe) odbieranie i przekazywanie sygnałów komórki glejowe (wspomagające) Sygnalizacja w komórkach nerwowych 100 tys. wejść informacyjnych przyjmowanie sygnału przewodzenie
Tkanka nerwowa neurony (pobudliwe) odbieranie i przekazywanie sygnałów komórki glejowe (wspomagające) Sygnalizacja w komórkach nerwowych 100 tys. wejść informacyjnych przyjmowanie sygnału przewodzenie
Model błony neuronowej
 Model błony neuronowej 1 Modelowanie pewnych aspektów czynności mózgu Neuron McCullocha i Pits a. Pierwsze próby matematycznego opisu czynności neuronów i próby zrozumienia w oparciu o te modele czynności
Model błony neuronowej 1 Modelowanie pewnych aspektów czynności mózgu Neuron McCullocha i Pits a. Pierwsze próby matematycznego opisu czynności neuronów i próby zrozumienia w oparciu o te modele czynności
Modelowanie pewnych aspektów czynności mózgu
 Tutorial: Modelowanie czynności neuronów i pewnych aspektów czynności mózgu 1 Modelowanie pewnych aspektów czynności mózgu Neuron McCullocha i Pits a. Pierwsze próby matematycznego opisu czynności neuronów
Tutorial: Modelowanie czynności neuronów i pewnych aspektów czynności mózgu 1 Modelowanie pewnych aspektów czynności mózgu Neuron McCullocha i Pits a. Pierwsze próby matematycznego opisu czynności neuronów
Transport przez błonę komórkową cd. Modelowanie
 Transport przez błonę komórkową cd. Modelowanie Kanały jonowe posiadają zdolność do kontrolowanego przepuszczania jonów. występowanie w nich tzw. pora wodna hydrofilowa przestrzeń wewnątrz białka, przez
Transport przez błonę komórkową cd. Modelowanie Kanały jonowe posiadają zdolność do kontrolowanego przepuszczania jonów. występowanie w nich tzw. pora wodna hydrofilowa przestrzeń wewnątrz białka, przez
n liczba moli elektronów E siła elektromotoryczna ogniwa F = en A stała Faradaya C/mol
 Zmiana entalpii swobodnej G podczas reakcji w której zachodzi przepływ elektronów jest pracą nieobjętościową i może być wyrażona jako iloczyn napięcie i ładunku. na przykład procesy oksydo-redukcyjne zachodzące
Zmiana entalpii swobodnej G podczas reakcji w której zachodzi przepływ elektronów jest pracą nieobjętościową i może być wyrażona jako iloczyn napięcie i ładunku. na przykład procesy oksydo-redukcyjne zachodzące
biologiczne mechanizmy zachowania seminarium + laboratorium M.Eng. Michal Adam Michalowski
 biologiczne mechanizmy zachowania seminarium + laboratorium M.Eng. Michal Adam Michalowski michal.michalowski@uwr.edu.pl michaladamichalowski@gmail.com michal.michalowski@uwr.edu.pl https://mmichalowskiuwr.wordpress.com/
biologiczne mechanizmy zachowania seminarium + laboratorium M.Eng. Michal Adam Michalowski michal.michalowski@uwr.edu.pl michaladamichalowski@gmail.com michal.michalowski@uwr.edu.pl https://mmichalowskiuwr.wordpress.com/
Biologiczne mechanizmy zachowania - fizjologia. zajecia 1 :
 Biologiczne mechanizmy zachowania - fizjologia zajecia 1 : 8.10.15 Kontakt: michaladammichalowski@gmail.com https://mmichalowskiuwr.wordpress.com/ II gr 08:00 10:0 III gr 10:15 11:45 IV gr 12:00 13:30
Biologiczne mechanizmy zachowania - fizjologia zajecia 1 : 8.10.15 Kontakt: michaladammichalowski@gmail.com https://mmichalowskiuwr.wordpress.com/ II gr 08:00 10:0 III gr 10:15 11:45 IV gr 12:00 13:30
ROLA WAPNIA W FIZJOLOGII KOMÓRKI
 ROLA WAPNIA W FIZJOLOGII KOMÓRKI Michał M. Dyzma PLAN REFERATU Historia badań nad wapniem Domeny białek wiążące wapń Homeostaza wapniowa w komórce Komórkowe rezerwuary wapnia Białka buforujące Pompy wapniowe
ROLA WAPNIA W FIZJOLOGII KOMÓRKI Michał M. Dyzma PLAN REFERATU Historia badań nad wapniem Domeny białek wiążące wapń Homeostaza wapniowa w komórce Komórkowe rezerwuary wapnia Białka buforujące Pompy wapniowe
Fizjologia czlowieka seminarium + laboratorium. M.Eng. Michal Adam Michalowski
 Fizjologia czlowieka seminarium + laboratorium M.Eng. Michal Adam Michalowski michal.michalowski@uwr.edu.pl michaladamichalowski@gmail.com michal.michalowski@uwr.edu.pl https://mmichalowskiuwr.wordpress.com/
Fizjologia czlowieka seminarium + laboratorium M.Eng. Michal Adam Michalowski michal.michalowski@uwr.edu.pl michaladamichalowski@gmail.com michal.michalowski@uwr.edu.pl https://mmichalowskiuwr.wordpress.com/
SPIS TREŚCI CZĘŚĆ LITERATUROWA OBJAŚNIENIE STOSOWANYCH W PRACY SKRÓTÓW... 6 OBJAŚNIENIE STOSOWANYCH W PRACY OZNACZEŃ... 8.
 SPIS TREŚCI OBJAŚNIENIE STOSOWANYCH W PRACY SKRÓTÓW... 6 OBJAŚNIENIE STOSOWANYCH W PRACY OZNACZEŃ... 8 Rozdział 1 CEL PRACY 1.1. Przedmiot prowadzonych badań... l1 1.2. Cel pracy... 15 CZĘŚĆ LITERATUROWA
SPIS TREŚCI OBJAŚNIENIE STOSOWANYCH W PRACY SKRÓTÓW... 6 OBJAŚNIENIE STOSOWANYCH W PRACY OZNACZEŃ... 8 Rozdział 1 CEL PRACY 1.1. Przedmiot prowadzonych badań... l1 1.2. Cel pracy... 15 CZĘŚĆ LITERATUROWA
Opracowanie modelu matryc mikroelektrodowych oraz układu scalonego do elektrycznej stymulacji żywych sieci neuronowych
 Akademia Górniczo-Hutnicza im. Stanisława Staszica w Krakowie Wydział Fizyki i Informatyki Stosowanej Zakład Elektroniki Jądrowej Rozprawa doktorska Opracowanie modelu matryc mikroelektrodowych oraz układu
Akademia Górniczo-Hutnicza im. Stanisława Staszica w Krakowie Wydział Fizyki i Informatyki Stosowanej Zakład Elektroniki Jądrowej Rozprawa doktorska Opracowanie modelu matryc mikroelektrodowych oraz układu
Fizjologiczne podstawy badań elektrofizjologicznych obwodowego układu nerwowego
 neuroelektrofizjologia Fizjologiczne podstawy badań elektrofizjologicznych obwodowego układu nerwowego Rafał Rola I Klinika Neurologiczna, Instytut Psychiatrii i Neurologii, Warszawa Adres do korespondencji:
neuroelektrofizjologia Fizjologiczne podstawy badań elektrofizjologicznych obwodowego układu nerwowego Rafał Rola I Klinika Neurologiczna, Instytut Psychiatrii i Neurologii, Warszawa Adres do korespondencji:
LABORATORIUM BIOMECHANIKI
 LABORATORIUM BIOMECHANIKI ĆWICZENIE NR 1 BADANIE PARAMETRÓW SYGNAŁU ELEKTROMIOGRAFICZNEGO ORAZ WYZNACZANIE CZASU REFRAKCJI UKŁADU NERWOWO - MIĘŚNIOWEGO 1. CEL ĆWICZENIA Celem ćwiczenia jest zapoznanie
LABORATORIUM BIOMECHANIKI ĆWICZENIE NR 1 BADANIE PARAMETRÓW SYGNAŁU ELEKTROMIOGRAFICZNEGO ORAZ WYZNACZANIE CZASU REFRAKCJI UKŁADU NERWOWO - MIĘŚNIOWEGO 1. CEL ĆWICZENIA Celem ćwiczenia jest zapoznanie
INHIBICJA KANAŁÓW JONOWYCH POSZUKIWANIE NOWYCH LEKÓW.
 INHIBICJA KANAŁÓW JONOWYCH POSZUKIWANIE NOWYCH LEKÓW. Dariusz Matosiuk Katedra i Zakład Syntezy i Technologii Chemicznej Środków Leczniczych, Wydział Farmaceutyczny z Oddziałem Analityki Medycznej Uniwersytet
INHIBICJA KANAŁÓW JONOWYCH POSZUKIWANIE NOWYCH LEKÓW. Dariusz Matosiuk Katedra i Zakład Syntezy i Technologii Chemicznej Środków Leczniczych, Wydział Farmaceutyczny z Oddziałem Analityki Medycznej Uniwersytet
Biologiczne mechanizmy zachowania - fizjologia. zajecia 8 :
 Biologiczne mechanizmy zachowania - fizjologia zajecia 8 : 19.11.15 Kontakt: michaladammichalowski@gmail.com https://mmichalowskiuwr.wordpress.com/ I gr 08:30 10:00 II gr 10:15 11:45 III gr 12:00 13:30
Biologiczne mechanizmy zachowania - fizjologia zajecia 8 : 19.11.15 Kontakt: michaladammichalowski@gmail.com https://mmichalowskiuwr.wordpress.com/ I gr 08:30 10:00 II gr 10:15 11:45 III gr 12:00 13:30
Dr inż. Marta Kamińska
 Nowe techniki i technologie dla medycyny Dr inż. Marta Kamińska Układ nerwowy Układ nerwowy zapewnia łączność organizmu ze światem zewnętrznym, zezpala układy w jedną całość, zprawując jednocześnie nad
Nowe techniki i technologie dla medycyny Dr inż. Marta Kamińska Układ nerwowy Układ nerwowy zapewnia łączność organizmu ze światem zewnętrznym, zezpala układy w jedną całość, zprawując jednocześnie nad
Procesy stochastyczne w kardiologii od elektrofizjologii do zmienności rytmu serca cz. 1. Monika Petelczyc Wydział Fizyki Politechnika Warszawska
 Procesy stochastyczne w kardiologii od elektrofizjologii do zmienności rytmu serca cz. 1 Monika Petelczyc Wydział Fizyki Politechnika Warszawska Od poziomu makro do komórki http://www.myofilament.org/photosandmovies/movies.htm
Procesy stochastyczne w kardiologii od elektrofizjologii do zmienności rytmu serca cz. 1 Monika Petelczyc Wydział Fizyki Politechnika Warszawska Od poziomu makro do komórki http://www.myofilament.org/photosandmovies/movies.htm
Co to są wzorce rytmów?
 Sieci neuropodobne XII, Centralne generatory wzorców 1 Co to są wzorce rytmów? Centralne generatory rytmów są układami neuronowymi powodujących cykliczną aktywację odpowiednich mięśni, mogą działać w pewnym
Sieci neuropodobne XII, Centralne generatory wzorców 1 Co to są wzorce rytmów? Centralne generatory rytmów są układami neuronowymi powodujących cykliczną aktywację odpowiednich mięśni, mogą działać w pewnym
Receptory neurotransmiterów, klasyfikacja, drogi wewnątrzkomórkowej transdukcji sygnału
 Receptory neurotransmiterów, klasyfikacja, drogi wewnątrzkomórkowej transdukcji sygnału Zasadnicze zjawiska i procesy elektryczne w neurobiologii i metody ich badań zjawisk elektrycznych w neurobiologii
Receptory neurotransmiterów, klasyfikacja, drogi wewnątrzkomórkowej transdukcji sygnału Zasadnicze zjawiska i procesy elektryczne w neurobiologii i metody ich badań zjawisk elektrycznych w neurobiologii
Diagnostyka i protetyka słuchu i wzroku. Układ nerwowy człowieka. Przygotowała: prof. Bożena Kostek
 Diagnostyka i protetyka słuchu i wzroku Układ nerwowy człowieka Przygotowała: prof. Bożena Kostek receptory ośrodkowy układ nerwowy efektory układ autonomiczny ... ośrodkowy układ nerwowy receptory...
Diagnostyka i protetyka słuchu i wzroku Układ nerwowy człowieka Przygotowała: prof. Bożena Kostek receptory ośrodkowy układ nerwowy efektory układ autonomiczny ... ośrodkowy układ nerwowy receptory...
Model Poissona-Nernsta-Plancka w predykcji struktury kanałów białkowych
 Model Poissona-Nernsta-Plancka w predykcji struktury kanałów białkowych mgr inż. Witold Dyrka Opiekun: dr hab. inż. Małgorzata Kotulska Instytut Inżynierii Biomedycznej i Pomiarowej Plan wystąpienia Nanopory
Model Poissona-Nernsta-Plancka w predykcji struktury kanałów białkowych mgr inż. Witold Dyrka Opiekun: dr hab. inż. Małgorzata Kotulska Instytut Inżynierii Biomedycznej i Pomiarowej Plan wystąpienia Nanopory
Podstawy Elektrotechniki i Elektroniki. Opracował: Mgr inż. Marek Staude
 Podstawy Elektrotechniki i Elektroniki Opracował: Mgr inż. Marek Staude Część 1 Podstawowe prawa obwodów elektrycznych Prąd elektryczny definicja fizyczna Prąd elektryczny powstaje jako uporządkowany ruch
Podstawy Elektrotechniki i Elektroniki Opracował: Mgr inż. Marek Staude Część 1 Podstawowe prawa obwodów elektrycznych Prąd elektryczny definicja fizyczna Prąd elektryczny powstaje jako uporządkowany ruch
Wykład I. Komórka. 1. Bioczasteczki : węglowodany, białka, tłuszcze nukleotydy
 Wykład I. Komórka 1. Bioczasteczki : węglowodany, białka, tłuszcze nukleotydy 2. Funkcje białek błonowych: 1. Transport: a. bierny b. czynny, z wykorzystaniem energii 2. Aktywność enzymatyczna 3. Receptory
Wykład I. Komórka 1. Bioczasteczki : węglowodany, białka, tłuszcze nukleotydy 2. Funkcje białek błonowych: 1. Transport: a. bierny b. czynny, z wykorzystaniem energii 2. Aktywność enzymatyczna 3. Receptory
Dywergencja/konwergencja połączeń między neuronami
 OD NEURONU DO SIECI: MODELOWANIE UKŁADU NERWOWEGO Własności sieci, plastyczność synaps Stefan KASICKI SWPS, SPIK wiosna 2007 s.kasicki@nencki.gov.pl Dywergencja/konwergencja połączeń między neuronami 1
OD NEURONU DO SIECI: MODELOWANIE UKŁADU NERWOWEGO Własności sieci, plastyczność synaps Stefan KASICKI SWPS, SPIK wiosna 2007 s.kasicki@nencki.gov.pl Dywergencja/konwergencja połączeń między neuronami 1
Modelowanie wektora magnetycznego serca na podstawie jonowych prądów komórkowych
 Modelowanie wektora magnetycznego serca na podstawie jonowych prądów komórkowych Wstęp Podstawy modelu komórkowego Proces pobudzenia serca Wektor magnetyczny serca MoŜliwości diagnostyczne Wstęp Przepływający
Modelowanie wektora magnetycznego serca na podstawie jonowych prądów komórkowych Wstęp Podstawy modelu komórkowego Proces pobudzenia serca Wektor magnetyczny serca MoŜliwości diagnostyczne Wstęp Przepływający
Tom 54 2005 Numer 4 (269) Strony 373 379
 Strony 373 379") Tom 54 2005 Numer 4 (269) Strony 373 379 Piotr Koprowski, Wojciech Grajkowski, Andrzej Kubalski Zakład Biologii Komórki Instytut Biologii Doświadczalnej PAN Pasteura 3, 02-093 Warszawa email:a.kubalski@nencki.gov.pl
Tom 54 2005 Numer 4 (269) Strony 373 379 Piotr Koprowski, Wojciech Grajkowski, Andrzej Kubalski Zakład Biologii Komórki Instytut Biologii Doświadczalnej PAN Pasteura 3, 02-093 Warszawa email:a.kubalski@nencki.gov.pl
MECHANIZMY RUCHÓW KOMÓRKOWYCH - DZIAŁANIE ANESTETYKÓW NA KOMÓRKI
 MECHANIZMY RUCHÓW KOMÓRKOWYCH - DZIAŁANIE ANESTETYKÓW NA KOMÓRKI Zakres materiału, który naleŝy przygotować do ćwiczeń: 1) Budowa błony komórkowej 2) Mechanizm działania anestetyków 3) Aktywność ruchowa
MECHANIZMY RUCHÓW KOMÓRKOWYCH - DZIAŁANIE ANESTETYKÓW NA KOMÓRKI Zakres materiału, który naleŝy przygotować do ćwiczeń: 1) Budowa błony komórkowej 2) Mechanizm działania anestetyków 3) Aktywność ruchowa
biologiczne mechanizmy zachowania seminarium + laboratorium M.Eng. Michal Adam Michalowski
 biologiczne mechanizmy zachowania seminarium + laboratorium M.Eng. Michal Adam Michalowski michal.michalowski@uwr.edu.pl michaladamichalowski@gmail.com michal.michalowski@uwr.edu.pl https://mmichalowskiuwr.wordpress.com/
biologiczne mechanizmy zachowania seminarium + laboratorium M.Eng. Michal Adam Michalowski michal.michalowski@uwr.edu.pl michaladamichalowski@gmail.com michal.michalowski@uwr.edu.pl https://mmichalowskiuwr.wordpress.com/
SKRYPT DO ĆWICZEŃ Z BIOFIZYKI. dla studentów biologii i biotechnologii UMCS w Lublinie. Wydawnictwo Uniwersytetu Marii Curie-Skłodowskiej
 SKRYPT DO ĆWICZEŃ Z BIOFIZYKI dla studentów biologii i biotechnologii UMCS w Lublinie Wydawnictwo Uniwersytetu Marii Curie-Skłodowskiej SKRYPT DO ĆWICZEŃ Z BIOFIZYKI dla studentów biologii i biotechnologii
SKRYPT DO ĆWICZEŃ Z BIOFIZYKI dla studentów biologii i biotechnologii UMCS w Lublinie Wydawnictwo Uniwersytetu Marii Curie-Skłodowskiej SKRYPT DO ĆWICZEŃ Z BIOFIZYKI dla studentów biologii i biotechnologii
Kanałopatie neuronalne. Neuronal channelopathies. *Rafał Rola, Danuta Ryglewicz. Postępy Nauk Medycznych, t. XXV, nr 1, 2012
 Postępy Nauk Medycznych, t. XXV, nr 1, 2012 Borgis *Rafał Rola, Danuta Ryglewicz Kanałopatie neuronalne Neuronal channelopathies I Klinika Neurologii, Instytut Psychiatrii i Neurologii w Warszawie Kierownik
Postępy Nauk Medycznych, t. XXV, nr 1, 2012 Borgis *Rafał Rola, Danuta Ryglewicz Kanałopatie neuronalne Neuronal channelopathies I Klinika Neurologii, Instytut Psychiatrii i Neurologii w Warszawie Kierownik
Neurobiologia wybrane zagadnienia neuroscience
 Neurobiologia wybrane zagadnienia neuroscience Dariusz Adamek Zakład Neuropatologii CM UJ Materiały do wykładów z neurobiologii dla studentów Wydziału Lekarskiego i Stomatologii CM UJ Materiały stanowią
Neurobiologia wybrane zagadnienia neuroscience Dariusz Adamek Zakład Neuropatologii CM UJ Materiały do wykładów z neurobiologii dla studentów Wydziału Lekarskiego i Stomatologii CM UJ Materiały stanowią
Biofizyka
 Biofizyka 12 21. 01. 2008 Zachowanie się układów różnym stopniu złożoności w zewnętrznym polu elektrycznym Właściwości elektryczne substancji Elektryczne właściwości substancji uwarunkowane są dwiema podstawowymi
Biofizyka 12 21. 01. 2008 Zachowanie się układów różnym stopniu złożoności w zewnętrznym polu elektrycznym Właściwości elektryczne substancji Elektryczne właściwości substancji uwarunkowane są dwiema podstawowymi
Neurologia dla studentów wydziału pielęgniarstwa. Bożena Adamkiewicz Andrzej Głąbiński Andrzej Klimek
 Neurologia dla studentów wydziału pielęgniarstwa Bożena Adamkiewicz Andrzej Głąbiński Andrzej Klimek Spis treści Wstęp... 7 Część I. Wiadomości ogólne... 9 1. Podstawy struktury i funkcji układu nerwowego...
Neurologia dla studentów wydziału pielęgniarstwa Bożena Adamkiewicz Andrzej Głąbiński Andrzej Klimek Spis treści Wstęp... 7 Część I. Wiadomości ogólne... 9 1. Podstawy struktury i funkcji układu nerwowego...
Rozdział 4. nierównomierne rozmieszczenie jonów?
 PRZEWODNICTWO NERWOWE I TRANSMISJA SYNAPTYCZNA Rozdział 4 Potencjał błonowy różnica w ładunku elektrycznym (potencjałów) pomiędzy wnętrzem a zewnętrzem komórki Jak go zarejestrować? używając mikroelektrod.
PRZEWODNICTWO NERWOWE I TRANSMISJA SYNAPTYCZNA Rozdział 4 Potencjał błonowy różnica w ładunku elektrycznym (potencjałów) pomiędzy wnętrzem a zewnętrzem komórki Jak go zarejestrować? używając mikroelektrod.
SIECI NEURONOWE Liniowe i nieliniowe sieci neuronowe
 SIECI NEURONOWE Liniowe i nieliniowe sieci neuronowe JOANNA GRABSKA-CHRZĄSTOWSKA Wykłady w dużej mierze przygotowane w oparciu o materiały i pomysły PROF. RYSZARDA TADEUSIEWICZA BUDOWA RZECZYWISTEGO NEURONU
SIECI NEURONOWE Liniowe i nieliniowe sieci neuronowe JOANNA GRABSKA-CHRZĄSTOWSKA Wykłady w dużej mierze przygotowane w oparciu o materiały i pomysły PROF. RYSZARDA TADEUSIEWICZA BUDOWA RZECZYWISTEGO NEURONU
Drgania wymuszone - wahadło Pohla
 Zagadnienia powiązane Częstość kołowa, częstotliwość charakterystyczna, częstotliwość rezonansowa, wahadło skrętne, drgania skrętne, moment siły, moment powrotny, drgania tłumione/nietłumione, drgania
Zagadnienia powiązane Częstość kołowa, częstotliwość charakterystyczna, częstotliwość rezonansowa, wahadło skrętne, drgania skrętne, moment siły, moment powrotny, drgania tłumione/nietłumione, drgania
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2122364 (96) Data i numer zgłoszenia patentu europejskiego: 1.02.08 08707734.3 (13) (1) T3 Int.Cl. G01N 33/68 (06.01) G01N
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2122364 (96) Data i numer zgłoszenia patentu europejskiego: 1.02.08 08707734.3 (13) (1) T3 Int.Cl. G01N 33/68 (06.01) G01N
Cel modelowania neuronów realistycznych biologicznie:
 Sieci neuropodobne XI, modelowanie neuronów biologicznie realistycznych 1 Cel modelowania neuronów realistycznych biologicznie: testowanie hipotez biologicznych i fizjologicznych eksperymenty na modelach
Sieci neuropodobne XI, modelowanie neuronów biologicznie realistycznych 1 Cel modelowania neuronów realistycznych biologicznie: testowanie hipotez biologicznych i fizjologicznych eksperymenty na modelach
Co się stanie, gdy połączymy szeregowo dwie żarówki?
 Różne elementy układu elektrycznego można łączyć szeregowo. Z wartości poszczególnych oporów, można wyznaczyć oporność całkowitą oraz całkowite natężenie prądu. Zadania 1. Połącz szeregowo dwie identyczne
Różne elementy układu elektrycznego można łączyć szeregowo. Z wartości poszczególnych oporów, można wyznaczyć oporność całkowitą oraz całkowite natężenie prądu. Zadania 1. Połącz szeregowo dwie identyczne
Tkanka mięśniowa pobudliwość kurczliwość Miofilamenty nie kurczą się, lecz przesuwają względem siebie ( główki miozyny kroczą po aktynie)
") Tkanka mięśniowa Aparat kuczliwy: miofilamenty cienkie (aktyna i białka pomocnicze) miofilamenty grube (miozyna 2) pobudliwość kurczliwość Miofilamenty nie kurczą się, lecz przesuwają względem siebie (
Tkanka mięśniowa Aparat kuczliwy: miofilamenty cienkie (aktyna i białka pomocnicze) miofilamenty grube (miozyna 2) pobudliwość kurczliwość Miofilamenty nie kurczą się, lecz przesuwają względem siebie (
biologia w gimnazjum OBWODOWY UKŁAD NERWOWY
 biologia w gimnazjum 2 OBWODOWY UKŁAD NERWOWY BUDOWA KOMÓRKI NERWOWEJ KIERUNEK PRZEWODZENIA IMPULSU NEROWEGO DENDRYT ZAKOŃCZENIA AKSONU CIAŁO KOMÓRKI JĄDRO KOMÓRKOWE AKSON OSŁONKA MIELINOWA Komórka nerwowa
biologia w gimnazjum 2 OBWODOWY UKŁAD NERWOWY BUDOWA KOMÓRKI NERWOWEJ KIERUNEK PRZEWODZENIA IMPULSU NEROWEGO DENDRYT ZAKOŃCZENIA AKSONU CIAŁO KOMÓRKI JĄDRO KOMÓRKOWE AKSON OSŁONKA MIELINOWA Komórka nerwowa
Ćwiczenie 1: Wyznaczanie warunków odporności, korozji i pasywności metali
 Ćwiczenie 1: Wyznaczanie warunków odporności, korozji i pasywności metali Wymagane wiadomości Podstawy korozji elektrochemicznej, wykresy E-pH. Wprowadzenie Główną przyczyną zniszczeń materiałów metalicznych
Ćwiczenie 1: Wyznaczanie warunków odporności, korozji i pasywności metali Wymagane wiadomości Podstawy korozji elektrochemicznej, wykresy E-pH. Wprowadzenie Główną przyczyną zniszczeń materiałów metalicznych
Genetyka padaczek. Joanna Jędrzejczak Klinika Neurologii i Epileptologii CMKP Warszawa
 Genetyka padaczek Joanna Jędrzejczak Klinika Neurologii i Epileptologii CMKP Warszawa Genetyka padaczek Cel Mapa genowa w padaczce Przegląd najnowszych badań związanych z mutacjami genów odpowiedzialnych
Genetyka padaczek Joanna Jędrzejczak Klinika Neurologii i Epileptologii CMKP Warszawa Genetyka padaczek Cel Mapa genowa w padaczce Przegląd najnowszych badań związanych z mutacjami genów odpowiedzialnych
Biologiczne mechanizmy zachowania - fizjologia. zajecia 6 :
 Biologiczne mechanizmy zachowania - fizjologia zajecia 6 : 12.11.15 Kontakt: michaladammichalowski@gmail.com https://mmichalowskiuwr.wordpress.com/ I gr 08:30 10:00 (s. Cybulskiego; 08.10. 19.11.) II gr
Biologiczne mechanizmy zachowania - fizjologia zajecia 6 : 12.11.15 Kontakt: michaladammichalowski@gmail.com https://mmichalowskiuwr.wordpress.com/ I gr 08:30 10:00 (s. Cybulskiego; 08.10. 19.11.) II gr
Przyrządy i Układy Półprzewodnikowe
 VI. Prostownik jedno i dwupołówkowy Cel ćwiczenia: Poznanie zasady działania układu prostownika jedno i dwupołówkowego. A) Wstęp teoretyczny Prostownik jest układem elektrycznym stosowanym do zamiany prądu
VI. Prostownik jedno i dwupołówkowy Cel ćwiczenia: Poznanie zasady działania układu prostownika jedno i dwupołówkowego. A) Wstęp teoretyczny Prostownik jest układem elektrycznym stosowanym do zamiany prądu
Procesy stochastyczne w kardiologii od elektrofizjologii do zmienności rytmu serca cz. 2. Monika Petelczyc Wydział Fizyki Politechnika Warszawska
 Procesy stochastyczne w kardiologii od elektrofizjologii do zmienności rytmu serca cz. 2 Monika Petelczyc Wydział Fizyki Politechnika Warszawska Model elektryczny błony Hipoteza błonowa Bernsteina (192):
Procesy stochastyczne w kardiologii od elektrofizjologii do zmienności rytmu serca cz. 2 Monika Petelczyc Wydział Fizyki Politechnika Warszawska Model elektryczny błony Hipoteza błonowa Bernsteina (192):
Czy prąd przepływający przez ciecz zmienia jej własności chemiczne?
 Czy prąd przepływający przez ciecz zmienia Zadanie Zmierzenie charakterystyki prądowo-napięciowej elektrolitu zawierającego roztwór siarczanu miedzi dla elektrod miedzianych. Obserwacja widocznych zmian
Czy prąd przepływający przez ciecz zmienia Zadanie Zmierzenie charakterystyki prądowo-napięciowej elektrolitu zawierającego roztwór siarczanu miedzi dla elektrod miedzianych. Obserwacja widocznych zmian
Praktyczne aspekty modelowania układu nerwowego
 Praktyczne aspekty modelowania układu nerwowego Ćwiczenia 1 Modelowanie układu nerwowego w GENESIS dr Daniel Wójcik d.wojcik [malpa] nencki.gov.pl na podstawie The Book of GENESIS Modelowanie 1)Tworzymy
Praktyczne aspekty modelowania układu nerwowego Ćwiczenia 1 Modelowanie układu nerwowego w GENESIS dr Daniel Wójcik d.wojcik [malpa] nencki.gov.pl na podstawie The Book of GENESIS Modelowanie 1)Tworzymy
Natężenie prądu elektrycznego
 Natężenie prądu elektrycznego Wymuszenie w przewodniku różnicy potencjałów powoduje przepływ ładunków elektrycznych. Powszechnie przyjmuje się, że przepływający prąd ma taki sam kierunek jak przepływ ładunków
Natężenie prądu elektrycznego Wymuszenie w przewodniku różnicy potencjałów powoduje przepływ ładunków elektrycznych. Powszechnie przyjmuje się, że przepływający prąd ma taki sam kierunek jak przepływ ładunków
Historia elektrochemii
 Historia elektrochemii Luigi Galvani (1791): elektryczność zwierzęca Od żab do ogniw Alessandro Volta (około 1800r): weryfikacja doświadczeń Galvaniego Umieszczenie dwóch różnych metali w ciele żaby może
Historia elektrochemii Luigi Galvani (1791): elektryczność zwierzęca Od żab do ogniw Alessandro Volta (około 1800r): weryfikacja doświadczeń Galvaniego Umieszczenie dwóch różnych metali w ciele żaby może
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU HALO, NEURON. ZGŁOŚ SIĘ.
 SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU HALO, NEURON. ZGŁOŚ SIĘ. SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy.
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU HALO, NEURON. ZGŁOŚ SIĘ. SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy.
Prąd elektryczny - przepływ ładunku
 Prąd elektryczny - przepływ ładunku I Q t Natężenie prądu jest to ilość ładunku Q przepływającego przez dowolny przekrój przewodnika w ciągu jednostki czasu t. Dla prądu stałego natężenie prądu I jest
Prąd elektryczny - przepływ ładunku I Q t Natężenie prądu jest to ilość ładunku Q przepływającego przez dowolny przekrój przewodnika w ciągu jednostki czasu t. Dla prądu stałego natężenie prądu I jest
LECZENIE PRZEWLEKŁYCH ZAKAŻEŃ PŁUC U PACJENTÓW
 Nazwa programu: Terapeutyczne Programy Zdrowotne 2012 Załącznik nr 30 do Zarządzenia Nr 59/2011/DGL Prezesa NFZ z dnia 10 października 2011 roku LECZENIE PRZEWLEKŁYCH ZAKAŻEŃ PŁUC U PACJENTÓW Z MUKOWISCYDOZĄ
Nazwa programu: Terapeutyczne Programy Zdrowotne 2012 Załącznik nr 30 do Zarządzenia Nr 59/2011/DGL Prezesa NFZ z dnia 10 października 2011 roku LECZENIE PRZEWLEKŁYCH ZAKAŻEŃ PŁUC U PACJENTÓW Z MUKOWISCYDOZĄ
Tkanka nerwowa. pobudliwość przewodnictwo
 Komórki: komórki nerwowe (neurony) - sygnalizacja, neurosekrecja komórki neurogleju (glejowe) - ochrona, wspomaganie Tkanka nerwowa Substancja międzykomórkowa: prawie nieobecna (blaszki podstawne) pobudliwość
Komórki: komórki nerwowe (neurony) - sygnalizacja, neurosekrecja komórki neurogleju (glejowe) - ochrona, wspomaganie Tkanka nerwowa Substancja międzykomórkowa: prawie nieobecna (blaszki podstawne) pobudliwość
BIOLOGICZNE MECHANIZMY ZACHOWANIA II JĄDRA PODSTAWY KRESOMÓZGOWIA I KONTROLA RUCHOWA
 BIOLOGICZNE MECHANIZMY ZACHOWANIA II JĄDRA PODSTAWY KRESOMÓZGOWIA I KONTROLA RUCHOWA MECHANIZMY KONTROLI RUCHOWEJ SYSTEMY ZSTĘPUJĄCE Korowe ośrodki motoryczne Kora motoryczna (planowanie, inicjacja i kierowanie
BIOLOGICZNE MECHANIZMY ZACHOWANIA II JĄDRA PODSTAWY KRESOMÓZGOWIA I KONTROLA RUCHOWA MECHANIZMY KONTROLI RUCHOWEJ SYSTEMY ZSTĘPUJĄCE Korowe ośrodki motoryczne Kora motoryczna (planowanie, inicjacja i kierowanie
ZAJĘCIA 1. uczenie się i pamięć mechanizmy komórkowe. dr Marek Binder Zakład Psychofizjologii
 ZAJĘCIA 1 uczenie się i pamięć mechanizmy komórkowe dr Marek Binder Zakład Psychofizjologii problem engramu dwa aspekty poziom systemowy które części mózgu odpowiadają za pamięć gdzie tworzy się engram?
ZAJĘCIA 1 uczenie się i pamięć mechanizmy komórkowe dr Marek Binder Zakład Psychofizjologii problem engramu dwa aspekty poziom systemowy które części mózgu odpowiadają za pamięć gdzie tworzy się engram?
Składniki cytoszkieletu. Szkielet komórki
 Składniki cytoszkieletu. Szkielet komórki aktynowe pośrednie aktynowe pośrednie 1 Elementy cytoszkieletu aktynowe pośrednie aktynowe filamenty aktynowe inaczej mikrofilamenty filamenty utworzone z aktyny
Składniki cytoszkieletu. Szkielet komórki aktynowe pośrednie aktynowe pośrednie 1 Elementy cytoszkieletu aktynowe pośrednie aktynowe filamenty aktynowe inaczej mikrofilamenty filamenty utworzone z aktyny
Wzmacniacze operacyjne
 Wzmacniacze operacyjne Cel ćwiczenia Celem ćwiczenia jest badanie podstawowych układów pracy wzmacniaczy operacyjnych. Wymagania Wstęp 1. Zasada działania wzmacniacza operacyjnego. 2. Ujemne sprzężenie
Wzmacniacze operacyjne Cel ćwiczenia Celem ćwiczenia jest badanie podstawowych układów pracy wzmacniaczy operacyjnych. Wymagania Wstęp 1. Zasada działania wzmacniacza operacyjnego. 2. Ujemne sprzężenie
Źródła zasilania i parametry przebiegu zmiennego
 POLIECHNIKA ŚLĄSKA WYDZIAŁ INŻYNIERII ŚRODOWISKA I ENERGEYKI INSYU MASZYN I URZĄDZEŃ ENERGEYCZNYCH LABORAORIUM ELEKRYCZNE Źródła zasilania i parametry przebiegu zmiennego (E 1) Opracował: Dr inż. Włodzimierz
POLIECHNIKA ŚLĄSKA WYDZIAŁ INŻYNIERII ŚRODOWISKA I ENERGEYKI INSYU MASZYN I URZĄDZEŃ ENERGEYCZNYCH LABORAORIUM ELEKRYCZNE Źródła zasilania i parametry przebiegu zmiennego (E 1) Opracował: Dr inż. Włodzimierz
Autonomiczny układ nerwowy - AUN
 Autonomiczny układ nerwowy - AUN AUN - różnice anatomiczne część współczulna część przywspółczulna włókna nerwowe tworzą odrębne nerwy (nerw trzewny większy) wchodzą w skład nerwów czaszkowych lub rdzeniowych
Autonomiczny układ nerwowy - AUN AUN - różnice anatomiczne część współczulna część przywspółczulna włókna nerwowe tworzą odrębne nerwy (nerw trzewny większy) wchodzą w skład nerwów czaszkowych lub rdzeniowych
Tkanka nerwowa. pobudliwość przewodnictwo
 Komórki: komórki nerwowe (neurony) - sygnalizacja, neurosekrecja komórki neurogleju (glejowe) - ochrona, wspomaganie Tkanka nerwowa Substancja międzykomórkowa: prawie nieobecna (blaszki podstawne) pobudliwość
Komórki: komórki nerwowe (neurony) - sygnalizacja, neurosekrecja komórki neurogleju (glejowe) - ochrona, wspomaganie Tkanka nerwowa Substancja międzykomórkowa: prawie nieobecna (blaszki podstawne) pobudliwość
Kompartmenty wodne ustroju
 Kompartmenty wodne ustroju Tomasz Irzyniec Oddział Nefrologii, Szpital MSWiA Katowice Zawartość wody w ustroju jest funkcją wieku, masy ciała i zawartości tłuszczu u dzieci zawartość wody wynosi około
Kompartmenty wodne ustroju Tomasz Irzyniec Oddział Nefrologii, Szpital MSWiA Katowice Zawartość wody w ustroju jest funkcją wieku, masy ciała i zawartości tłuszczu u dzieci zawartość wody wynosi około
Różne dziwne przewodniki
 Różne dziwne przewodniki czyli trzy po trzy o mechanizmach przewodzenia prądu elektrycznego Przewodniki elektronowe Metale Metale (zwane również przewodnikami) charakteryzują się tym, że elektrony ich
Różne dziwne przewodniki czyli trzy po trzy o mechanizmach przewodzenia prądu elektrycznego Przewodniki elektronowe Metale Metale (zwane również przewodnikami) charakteryzują się tym, że elektrony ich
Złącza p-n, zastosowania. Własności złącza p-n Dioda LED Fotodioda Dioda laserowa Tranzystor MOSFET
 Złącza p-n, zastosowania Własności złącza p-n Dioda LED Fotodioda Dioda laserowa Tranzystor MOSFET Złącze p-n, polaryzacja złącza, prąd dyfuzyjny (rekombinacyjny) Elektrony z obszaru n na złączu dyfundują
Złącza p-n, zastosowania Własności złącza p-n Dioda LED Fotodioda Dioda laserowa Tranzystor MOSFET Złącze p-n, polaryzacja złącza, prąd dyfuzyjny (rekombinacyjny) Elektrony z obszaru n na złączu dyfundują
ANALOGOWY MODEL TRANSMISJI SYNAPTYCZNEJ
 Ćwiczenie nr 17 ANALOGOWY MODEL TRANSMISJI SYNAPTYCZNEJ Aparatura Komputer wraz z neurosymulatorem, Cobra3. Przebieg ćwiczenia W ramach ćwiczenia przeprowadzone zostaną następujące badania: A. Wyznaczanie
Ćwiczenie nr 17 ANALOGOWY MODEL TRANSMISJI SYNAPTYCZNEJ Aparatura Komputer wraz z neurosymulatorem, Cobra3. Przebieg ćwiczenia W ramach ćwiczenia przeprowadzone zostaną następujące badania: A. Wyznaczanie
biologiczne mechanizmy zachowania seminarium + laboratorium M.Eng. Michal Adam Michalowski
 biologiczne mechanizmy zachowania seminarium + laboratorium M.Eng. Michal Adam Michalowski michal.michalowski@uwr.edu.pl michaladamichalowski@gmail.com michal.michalowski@uwr.edu.pl https://mmichalowskiuwr.wordpress.com/
biologiczne mechanizmy zachowania seminarium + laboratorium M.Eng. Michal Adam Michalowski michal.michalowski@uwr.edu.pl michaladamichalowski@gmail.com michal.michalowski@uwr.edu.pl https://mmichalowskiuwr.wordpress.com/
Nauka o Materiałach. Wykład XI. Właściwości cieplne. Jerzy Lis
 Nauka o Materiałach Wykład XI Właściwości cieplne Jerzy Lis Nauka o Materiałach Treść wykładu: 1. Stabilność termiczna materiałów 2. Pełzanie wysokotemperaturowe 3. Przewodnictwo cieplne 4. Rozszerzalność
Nauka o Materiałach Wykład XI Właściwości cieplne Jerzy Lis Nauka o Materiałach Treść wykładu: 1. Stabilność termiczna materiałów 2. Pełzanie wysokotemperaturowe 3. Przewodnictwo cieplne 4. Rozszerzalność
Tkanka nerwowa. pobudliwość przewodnictwo
 Komórki: komórki nerwowe (neurony) - sygnalizacja, neurosekrecja komórki neurogleju (glejowe) - ochrona, wspomaganie Tkanka nerwowa Substancja międzykomórkowa: prawie nieobecna (blaszki podstawne) pobudliwość
Komórki: komórki nerwowe (neurony) - sygnalizacja, neurosekrecja komórki neurogleju (glejowe) - ochrona, wspomaganie Tkanka nerwowa Substancja międzykomórkowa: prawie nieobecna (blaszki podstawne) pobudliwość
Jednostka chorobowa. 235200 HFE HFE 235200 Wykrycie mutacji w genie HFE odpowiedzialnych za heterochromatozę. Analiza mutacji w kodonach: C282Y, H63D.
 Jednostka chorobowa Jednostka Oznaczenie Chorobowa testu OMIM TM Badany Gen Literatura Gen OMIM TM Opis/cel badania Zakres analizy Materiał biologiczny Czas analizy [dni roboczych] Cena [PLN] HEMOCHROMATOZA
Jednostka chorobowa Jednostka Oznaczenie Chorobowa testu OMIM TM Badany Gen Literatura Gen OMIM TM Opis/cel badania Zakres analizy Materiał biologiczny Czas analizy [dni roboczych] Cena [PLN] HEMOCHROMATOZA
ZJAWISKA TERMOELEKTRYCZNE
 Wstęp W ZJAWISKA ERMOELEKRYCZNE W.1. Wstęp Do zjawisk termoelektrycznych zaliczamy: zjawisko Seebecka - efekt powstawania różnicy potencjałów elektrycznych na styku metali lub półprzewodników, zjawisko
Wstęp W ZJAWISKA ERMOELEKRYCZNE W.1. Wstęp Do zjawisk termoelektrycznych zaliczamy: zjawisko Seebecka - efekt powstawania różnicy potencjałów elektrycznych na styku metali lub półprzewodników, zjawisko
DZIAŁ I. Zalecane źródła informacji Fizjologia człowieka. Podręcznik dla studentów medycyny. Red. Stanisław J. Konturek, Elservier Urban&Partner 2007
 DZIAŁ I. PODSTAWY REGULACJI I KONTROLI CZYNNOŚCI ORGANIZMU. TKANKI POBUDLIWE. Ćw. 1. Fizjologia jako nauka o homeostazie. (1-2 X 2012) 1. Wprowadzenie do przedmiotu. 2. Fizjologia i jej znaczenie w naukach
DZIAŁ I. PODSTAWY REGULACJI I KONTROLI CZYNNOŚCI ORGANIZMU. TKANKI POBUDLIWE. Ćw. 1. Fizjologia jako nauka o homeostazie. (1-2 X 2012) 1. Wprowadzenie do przedmiotu. 2. Fizjologia i jej znaczenie w naukach
FIZJOLOGIA ZWIERZĄT prof. dr hab. Krystyna Skwarło-Sońta rok akad. 2012/2013
 FIZJOLOGIA ZWIERZĄT prof. dr hab. Krystyna Skwarło-Sońta rok akad. 2012/2013 CZYNNIKI ŚRODOWISKOWE Zmieniająca się w ciągu roku długość dnia i nocy (fotoperiod), wyznacza sezonowość zmian pozostałych czynników:
FIZJOLOGIA ZWIERZĄT prof. dr hab. Krystyna Skwarło-Sońta rok akad. 2012/2013 CZYNNIKI ŚRODOWISKOWE Zmieniająca się w ciągu roku długość dnia i nocy (fotoperiod), wyznacza sezonowość zmian pozostałych czynników:
Wykład 3 Zjawiska transportu Dyfuzja w gazie, przewodnictwo cieplne, lepkość gazu, przewodnictwo elektryczne
 Wykład 3 Zjawiska transportu Dyfuzja w gazie, przewodnictwo cieplne, lepkość gazu, przewodnictwo elektryczne W3. Zjawiska transportu Zjawiska transportu zachodzą gdy układ dąży do stanu równowagi. W zjawiskach
Wykład 3 Zjawiska transportu Dyfuzja w gazie, przewodnictwo cieplne, lepkość gazu, przewodnictwo elektryczne W3. Zjawiska transportu Zjawiska transportu zachodzą gdy układ dąży do stanu równowagi. W zjawiskach