Natknąłem się na nieco archiwalny wątek na forum dyskusyjnym, jakie funkcjonuje na portalu racjonalista. Cytowany przez kogoś autor usiłuje w nim podw
|
|
|
- Justyna Kowalik
- 8 lat temu
- Przeglądów:
Transkrypt
1 Cz:1
2 Natknąłem się na nieco archiwalny wątek na forum dyskusyjnym, jakie funkcjonuje na portalu racjonalista. Cytowany przez kogoś autor usiłuje w nim podważyć nieredukowalną złożoność symbolu teorii inteligentnego projektu w biologii -wici bakteryjnej. Postanowiłem w oparciu o dyskusję z treścią tego tekstu wykazać błędy i niedociągnięcia modelu kooptacji. Krytyk Powołuje się na ogólnikowy model Nicholasa J. Matzke, w którym próbuje opisać ewolucję wici bakteryjnej w wyniku połączenia się i dalszej współpracy odrębnych podzespołów, które wcześniej pełniły inne funkcje (kooptacja) Ten filmik i schemat ilustrują, jak system wydzielniczy typu III miał wejść w skład budowy wici bakteryjnej: [1] v=sdwtwnpyr9w&list=plpqnr5ubp_6et_8paumjvryvw7dd sga8e%20
3
4 Twórca hipotezy kooptacji twierdzi, że mechanizm lokomocyjny bakterii e. cola powstał w wyniku połączenia się aparatu wydzielniczego typu III oraz syntazy ATP z resztą wici bakteryjnej oraz, że został radykalnie zmodyfikowany w celu pełnienia nowej funkcji, które będą omówione w dalszej części niniejszego tekstu. Założeniem pomysłu jest, że aparat wydzielniczy typu III zaczął w silniku bakteryjnym pełnić funkcję urządzenia transportującego białka, które budują wić bakteryjną [flageliny]: W poście tym będę cytował fragmenty wpisu z forum racjonalisty i komentował. Wzbogacę moje komentarze odpowiednimi ilustracjami, żeby każdy zainteresowany mógł sobie je obejrzeć i jeszcze lepiej zrozumieć zawartą w niniejszej polemice argumentację. Na wstępie zamieszczę kilka bardziej szczegółowych informacji technicznych na temat budowy silnika bakteryjnego. Należy je tutaj zamieścić, ponieważ to pozwoli każdemu czytelnikowi uświadomić sobie, jakiej jakości powinien być rzeczywisty model opisujący w sposób szczegółowy ewolucję wici bakteryjnej. Powinno się w nim krok po kroku wyjaśnić, jak niedopasowane
5 początkowo elementy zostały tak wyszlifowane przez ewolucję, że zaczęły być komplementarne w stosunku do siebie, jak puzle. Takie wyjaśnienie jest konieczne, ponieważ na każdym takim kroku, który nie dałby korzyści ewolucja by się zatrzymała i silnik bakteryjny by nie powstał. Więc nie chodzi tutaj tylko o czepianie się niedopracowanych szczegółów. Nie będę w tym tekście opisywał wszystkich trudności związanych z modelem kooptacji, ponieważ musiałbym w tym celu napisać grubą książkę. Skoncentruję się więc na najbardziej istotnych kwestiach. Białka niedopasowane: Białka dopasowane:
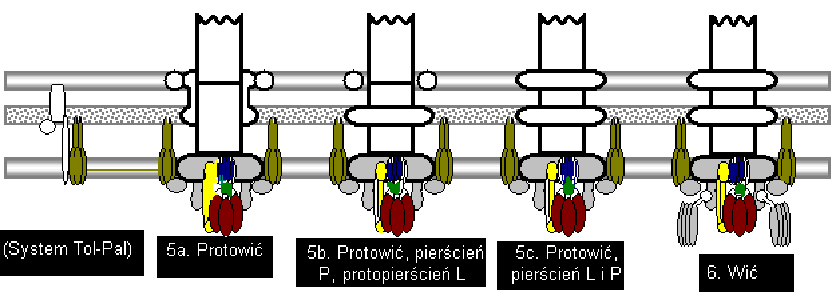
6 W rzęsce [wici] bakterii gram-ujemnej, na którą składa się około 25 białek w bardzo zróżnicowanej liczbie kopii, można wyróżnić trzy części: Ciało podstawowe, to obrotowy rotor o bardzo złożonej strukturze, który składa się z około 20 białek. Kotwiczy ono rzęskę w osłonach komórkowych i nadaje jej ruch obrotowy. Ciało podstawowe jet najbardziej złożoną częścią rzęski. Zbudowane jest z czterech pierścieni, przez który przechodzi centralny, pusty w środku rdzeń [kanał służący do transportu białek]
7 [2] Nazwa poszczególnych pierścieni wiąże się z ich rozmieszczeniem w osłonach. Najwcześniej powstająca część rzęski, pierścień MS, jest zbudowany z jednego białka [FliF]. Początkowo sądzono, że M i S to dwa pierścienie, lecz okazało się, że są tworzone przez różne domeny tego samego białka. Z zewnętrznej powierzchni pierścienia MS, na jego peryferycznej części, związane jest białko FliG. Pierścień ten znajduje się w błonie cytoplazm;atycznej i nad nią (ang. membrane /supramembrane). Pierścień P, zbudowany z białka Flil, jest zakotwiczony w warstwie mureiny (peptydoglikanu), a pierścień L (białko FliH) jest na poziomie lipopolisacharydowej warstwy błony zewnętrznej (LPS).
8 [3] Poniżej pierścienia MS znajduje się duża, przypominająca bęben struktura, zwana pierścieniem C
9 (cytoplazmatyczny) (białka FliM, FliN), w obrębie której występują białka FlhA i FlhB oraz Fli H, I, O, P, Q i R. Białka te stanowią kompleks aparatu transportowego, uczestniczącego w procesie transportu białek tworzących między innymi strukturę haka i włókna rzęski, będącego analogiem funkcjonalnym i strukturalnym kompleksu białkowego sekrecji typu III w komórkach wielu bakterii patogennych; Białka pierścienia C (FliM, FliN) oraz białko FliG wspólnie są określane, jako rotor lub kompleks przełącznikowy (ang. swith complex), gdyż decydują one o kierunku obrotu rzęski. Z białkiem FliM wiąże się białko CheY~P w procesie chemotaksji. Na poziomie pierścienia MS są związane białka statora, MotA i MotB. Drugie białko ma domenę wiązania z mureiną. W każdym obrotowym motorze znajduje się układ do 8 kompleksów każdy o składzie MotA4MotB2. Liczba kopii poszczególnych białek decydujących o obrocie rzęski wynosi: MotA do 32; MotB do 16; FliG -34; FliM34; FliN ponad 100. Na podstawie: 'Biologia molekularna bakterii', str. 107, 108m i 550, Warszawa 2006
10 Przebieg montażu silnika bakteryjnego: [4]
11
12
13 DYSKUSJA Jakiś czas temu miałem okazję rozmawiać z panem Kotasiem o ile dobrze pamiętam na temat wici bakteryjnej. Generalnie rzecz biorąc zrozumiałem to tak: Okazuje się jednak, że wić składa się z elementów, które wskazują na swoją odrębność. Kolejno "Podstawka", tworząca kanał protonowy u innych bakterii i jest transporterem jonów magnezu. Ponadto wskazano, że cały ten układ jest zaskakująco podobny do systemu sekrecyjnego III u bakterii. Jest to jeden z systemów przenoszących białka na zewnątrz komórki i w ten sposób bakteria może wydzielać białka do środowiska [..]. Aparat wydzielniczy typu III Służy bakteriom do infekowania komórek eukariotycznych zjadliwymi białkami. Jego funkcja przypomina tą, jaką pełni strzykawka. Aparat wydzielniczy typu III jest ponadto urządzeniem nieredukowalnie złożonym. Na tych filmach można zobaczyć, jak służy bakterii salmonelli i dodatkowo przyjrzeć się jego budowie:
14 Należy dodać, że według modelu kooptacji nie taki sam układ wydzielniczy typu III wszedł w skład elementów budujących wić bakteryjną, tylko do niego podobny. Nie występujący współcześnie w przyrodzie. Syntaza ATP - silnik molekularny służący komórkom do produkcji energii - którego zmodyfikowana przez ewolucję wersja rzekomo wchodzi w skład budowy aparatu odpowiedzialnego za transport białek do wici też jest kompleksem nieredukowalnie złożonym:
15 Na poniższych ilustracjach pokazano białka opiekuńcze, które w cytoplazmie łączą się z flagelinami budującymi wić bakteryjną i eksportują je do bramy aparatu transportującego. Zwolennicy hipotezy kooptacji twierdzą, że podobieństwo w budowie tych wyspecjalizowanych molekuł dowodzi ich pochodzenia od syntazy ATP. Nie wyjaśniają jednak, jak mogła przebiegać ich stopniowa ewolucja, dostosowując je do funkcji, jakie pełnią obecnie: [6]
16 Neodarwiniści w podobny sposób tłumaczą ewolucję syntazy ATP, jak się próbuje wyjaśnić genezę wici bakteryjnej. W proponowanym modelu zakłada się połączenie dwóch osobnych modułów, które wcześniej pełniły inne funkcje. Pomysł ten nazwano: modularna hipoteza w ewolucji syntazy ATP. W druga część niniejszego artykułu zawiera krytykę tej koncepcji. Ilustrację wykonał Jerzy Dzik [5]
17 Autor dyskutowanego tekstu ciągnie dalej: [...] Rzekoma nieredukowalność tego systemu [wici bakteryjnej] upadła kiedy okazało się, że jest to system homologiczny funkcjonalnie do systemu sekrecji typu III. Napęd protonowy,który napędza ruch wici jest wymagany do transportu białek w tym systemie. Efekt ruchu wici jest zatem uzyskany przy okazji, co wyjaśnia pochodzenie ewolucyjne wici i systemu sekrecyjnego typu III. Wić powstaje w wypadku transportu flagelin przez ten system. [...] Za transport białek do wici odpowiedzialna jest początkowo ATP -aza, która stanowi element aparatu dostarczającego flageliny. Dopiero następnie pomaga w tym pęd wirującego rotora. Aparat wydzielniczy typu III się nie kręci, więc nie można twierdzić, że mógł on być pod tym względem prekursorem ewolucyjnym silnika bakteryjnego. Możliwość nabycia przez tą strukturę możliwości obracania się wymagałoby przebycia wielu trudności, które są nie do ominięcia na drodze stopniowej ewolucji. Do tego, aby silnik bakteryjny mógł się obracać, czy zmieniać kierunek wirowania, potrzeba wielu dodatkowych elementów i daleko idących modyfikacji rzekomo homologicznych do wiciowych elementów występujących w aparacie wydzielniczym typu III. Na kolejnych ilustracjach pokazano w jaki sposób działa system transportu białek w wici bakteryjnej. Dostosowanie systemu wydzielniczego typu III, który pierwotnie pełnił rolę strzykawki infekującej komórki do tego typu funkcji również wymagałoby radykalnych modyfikacji, z którymi stopniowa ewolucja by sobie nie poradziła. W procesie budowy systemu odpowiedzialnego za ruch bakterii bierze udział kilka
18 różnych białek, budujących hak czy samo włókno (śrubę). Nie koniec na tym. Żeby umożliwić komórce ich dostarczenie do bramy prowadzącej do kanały wiodącego do końca wici, musiałyby powstać odpowiednie białka transportujące oraz system kontroli tego procesu. Hipoteza kooptacji nie tłumaczy krok po kroku, jak mogłoby się to stać. Jest ona pełna uogólnień i skoków myślowych. Opis tego, co przedstawiają poszczególne ilustracje jest pod nimi. [6]
19
20
21
22 Nie od rzeczy będzie wspomnieć o nadzwyczajnych właściwościach białka flageliny, z którego zbudowana jest wić. Cząsteczki tego białka (oznaczonego jako FliC) o masie cząsteczkowej mają zdolność do łączenia się razem w regularne struktury podobne do nici i spirali o różnym promieniu krzywizny. Jest to proces przypominający krystalizację. Bakteria buduje z flageliny zawsze ściśle określoną formę sztywnej, skręconej w kształt korkociągu rureczki. Podczas montażu wici powstaje pytanie w jaki sposób tysiące cząsteczek flageliny dostarczane są do nowo
23 powstającej i wydłużającej się wici (a musi być to mechanizm niezwykle wydajny, ponieważ jeden skręt spirali wici składa się z około 5000 cząsteczek tego białka). Otóż flagellina transportowana jest na koniec aktualnie montowanego odcinka poprzez specjalny kanał wewnątrz budowanej wici w tempie około 50 cząsteczek flagelliny na sekundę. Proces budowy wici wieńczy umieszczenie na jej końcu specjalnej nasadki. Utworzona ostatecznie wić jest strukturą prawo- lub lewoskrętną, przy czym zmiana skrętności wici zachodzi przy każdej zmianie kierunku obrotów silnika obracającego wić: Przy obrotach silniczka zgodnego z ruchem wskazówek zegara wić wykazuje prawoskrętność i odwrotnie, przy obrotach w przeciwnym kierunku wykazuje lewoskrętność. Wić nadająca bakterii ruch postępowy w płynnym środowisku jest zatem odpowiednikiem śruby okrętowej lub śmigła. Wszystkie te właściwości wici narzucają szczególne wymagania wobec materiału, z którego jest ona zbudowana. [7]

![[statora] przez które przepływają protony](/docs-images/51/27966335/images/24-0.png "wprawiające silnik bakteryjny w ruch")
24 Tutaj natomiast schematy obrazujące prawdopodobne sposoby pracy elementów [statora] przez które przepływają protony wprawiające silnik bakteryjny w ruch obrotowy: [8]
25
26 Mechanizm generowania obrotów rotora jest słabo poznany. Wiadomo jednak, że odpowiadają za to złożone interakcje pomiędzy nim a statorem. Istnieją trzy modele tłumaczące ten machanizm: model turbiny wodnej, model kołowrotka i model elektrostatycznej turbiny protonowej. Według modelu turbiny wodnej, protony lub jony sodu płynące na powierzchni białek statora MotA i MotB wywołują kierunkowy ruch cząsteczek wody, który zmienia sferyczny kształt tych białek, dzięki czemu wywierają one nacisk na rotor w ten sposób go napędzając. Model kołowrotka zakłada, że protony/jony sodu wpływają do specjalnego tunelu w białkach kompleksu statora MotA i MotB i kierowane są do specyficznych komponentów rotora. Wymusza to ruch rotora, który dalej przekazuje te protony/jony sodu do następnego tunelu w statorze, skąd płyną one już do cytoplazmy (zob. rys. 3).
27 Model kołowrotka. Zakłada on, że strumień protonów lub jonów sodu płynie przez specjalny tunel w białku statora, nastpnie przepływa przez część rotora napędzając go w ten sposób i sąsiednim tunelem w statorze spływa do wnętrza komórki. Model elektrostatycznej turbiny protonowej przewiduje, że protony/jony sodu przepływające przez tunel w białkach statora MotA i MotB oddziałują z ładunkami elektrostatycznymi precyzyjnie rozmieszczonymi na tworzących pierścień rotora cząsteczkach białka FliG, stwarzając w ten sposób dynamiczne pole elektrostatyczne, które napędza rotor (zob. rys. 4). Wydaje się, że na dzień dzisiejszy ten właśnie model jest wśród badaczy najbardziej popularny. Przypuszczalny model generowania obrotów rotora w
28 wyniku elektrostatycznego oddziaływania pomiędzy strumieniem protonów lub jonów sodu przepływających przez tunel w białku statora MotA i MotB a ładunkami na powierzchni białkia rotora FliG. Niezależnie od tego, który model jest poprawny, obliczono, że jeden obrót silnika wymaga przepływu około tysiąca protonów. Obroty rotora napędzają ośkę, która z kolei przenosi obroty przez błonę komórkową i poprzez kątnik (uniwersalne łącze) nadaje ruch wici. [7] Autor krytykujący koncepcję nieredukowalnej złożoności wici bakteryjnej nie wiedział, lub nie chciał przyjąć do wiadomości [reguła wśród zwolenników samodziejstwa], że Michael Behe odpowiedział na zarzuty, których użył. Zacytuję, co napisał na ten temat, ale zanim zapoznacie się z tym materiałem proszę przeczytać prawdziwą, niezmanipulowaną definicję nieredukowalnej złożoności. Wówczas zrozumienie wywodu Behe'ego nie nastręczy nikomu trudności;:
29 Michael Behe napisał [9]: ( )W końcu, zamiast pokazać, w jaki sposób ich teoria radzi sobie z tym problemem, darwiniści starają się obejść problem nieredukowalnej złożoności przy pomocy gierek słownych. Podczas debaty, sponsorowanej przez American Museum of Natural History, która odbyła się w kwietniu 2002 roku między zwolennikami i przeciwnikami teorii inteligentnego projektu, Kenneth Miller rzeczywiście stwierdził (.)że pułapka na myszy nie jest nieredukowalnie złożona, gdyż jej podzbiory, a nawet każda osobna część, wciąż mogą funkcjonować niezależnie od tego układu. Miller zauważył, że drążek przytrzymujący z pułapki na myszy może służyć jako wykałaczka, a więc nadal pełni funkcję, nie będąc częścią pułapki na myszy. Wszystkich części pułapki można użyć jako przycisku do papieru ciągnął dalej więc każda z nich pełni jakieś funkcje. A skoro każdy przedmiot, który posiada masę, może posłużyć jako przycisk do papieru, to każda część czegokolwiek pełni swoją własną funkcję. Czary mary, nie istnieje nic takiego jak nieredukowalna zło- żoność.w taki oto prosty sposób wyjaśniono poważny problem dla
30 gradualizmu, który każde dziecko może dostrzec w systemach, takich jak pułapka na myszy.oczywiście, powyższe proste wyjaśnienie opiera się na ewidentnie błędnym przekonaniu, wyraźnej dwuznaczności. Miller używa słowa funkcja w dwóch różnych sensach. Przypomnijmy sobie, że definicja nieredukowalnej złożoności mówi, iż usunięcie jakiejś części powoduje, że system przestaje sprawnie funkcjonować. Nie wspominając o tym w swym wystąpieniu, Miller przenosi nacisk z osobnej funkcji samego nienaruszonego systemu na kwestię, czy możemy znaleźć inne zastosowanie (czy funkcję ) dla niektórych jego części. Jeśli jednak usunie się jakąś część z przedstawionej przeze mnie pułapki, to nie złapie ona już myszy. System faktycznie przestaje sprawnie funkcjonować, a więc jest nieredukowalnie złożony właśnie tak jak napisałem. Co więcej, funkcje tak łatwo przypisywane przez Millera częściom pułapki przycisk do papieru, wykałaczka, łańcuszek na klucze i tak dalej mają niewiele, albo nic wspólnego z funkcją całego układu łapaniem myszy, a więc nie daje nam to żadnej wskazówki dla wyjaśnienia, w jaki sposób funkcja systemu mogła powstać stopniowo. Miller nie wyjaśnił właściwie niczego. Pozostawiając problem pułapki na myszy za sobą, Miller przeszedł następnie do omówienia wici bakteryjnej i ponownie odwołał się do tego samego błędnego przekonania. Jeżeli nie pozostało nic innego, należy podziwiać tę zapierającą dech zuchwałość próby słownego obrócenia kolejnego poważnego problemu darwinizmu na jego korzyść. W ostatnich latach wykazano, że wić bakteryjna jest znacznie bardziej skomplikowanym systemem niż dotąd sądzono. Działa ona nie tylko jako urządzenie o napędzie
31 obrotowym, ale w jej skład wchodzi także wyszukany mechanizm transportujący białka z wewnątrz na zewnątrz komórki, tworzące wierzchni fragment wici. Miller bez zmrużenia oczu zapewnia, że wić nie jest nieredukowalnie złożona, gdyż pewnychbiałek wici może brakować, a pozostała reszta być może niezależnie może nadal transportować białka. (Białka podobne ale nie identyczne do białek znajdowanych w wici występują w systemie wydzielinowym typu III u niektórych bakterii). Miller ponownie popadł w dwuznaczność, przenosząc nacisk z funkcji układu, który działa jak maszyna o napędzie obrotowym, na zdolność podzbioru tego systemu do transportowania białek przez membranę. Jednak, jak argumentowałem, usunięcie części wici całkowicie odbiera temu układowi zdolność do funkcjonowania jak maszyna o napędzie obrotowym. Dlatego, niezgodnie z twierdzeniami Millera, wić rzeczywiście jest nieredukowalnie złożona. Co więcej, funkcja transportowania białek ma bezpośrednio tyle wspólnego z funkcją napędzania obrotowego, ile wykałaczka z pułapką na myszy. Tak więc odkrycie dodatkowej funkcji transportowania białek nie mówi nam niczego o tym, jak procesy darwinowskie mogły złożyć maszynę o napędzie obrotowym.
32 Modyfikacje i dodawanie części Neodarwiniści twierdzą, że wić powstała w wyniku połączenia się systemu wydzielniczego typu III [oraz późniejszej jego modyfikacji] z resztą części obecnych w wici bakteryjnej [tworzących głównie silnik]. Nie prezentują szczegółowego modelu teoretycznego opisującego, jak to się mogło krok po kroku stać, przy zachowaniu funkcji przez ćwierć czy półsilniki [formy przejściowe] - tylko jedynie ogólnikowe pomysły mające status komiksowych bajeczek. Niemniej neodarwiniści wskazują na pewne podobieństwa pomiędzy białkami budującymi niektóre elementy wici i systemu wydzielniczego typu III, nazywając je homologiami, co ma sugerować ich wspólne ewolucyjne pochodzenie. W artykule znakomicie pokazano te podobieństwa, ale też zasadnicze różnice, o których w rozmaitych propagandówkach odwołujących się do hipotezy kooptacji raczej się nie wspomina. Podkreślam: mamy do czynienia jedynie z podobieństwami, a nie homologiami w pełnym tego słowa znaczeniu, ponieważ te białka nie są takie same, a niektóre z nich mimo podobieństw pełnią zupełnie inne funkcje. Zmodyfikowanie białek obecnych w aparacie wydzielniczym typu III i przystosowanie ich do pełnienia funkcji rotora wymagałoby długiej ewolucji i ponadto dodania innych niezbędnych części, na przykład w postaci precyzyjnie dopasowanych statorów, przez które przepływają protony napędzające rotor. Sam rotor musiałby zostać ściśle dopasowany do statorów, a
33 odpowiednie jego domeny musiałyby nabyć możliwości wiązania protonów i ich uwalniania do wnętrza komórki po obrocie rotora. Poza tym musiałyby zajść inne modyfikacje umożliwiające ruch obrotowy rotora i wału przenoszącego siłę jego pracy na hak i później na wić [śrubę napędową]. Aparat wydzielniczy typu III się nie kręci, to prosta i ostro zakończona rura [pilus], sztywno osadzona w błonie komórkowej, przez którą z wnętrza komórki pompowane są białka na zewnątrz [konkretnie do infekowanej komórki eukariotycznej]; [10]
34 Podobne do wiciowych białka w systemie wydzielniczym typu III przede wszystkim spełniają funkcję pierścieni stacjonarnie zakotwiczających to urządzenie w błonie komórkowej. Transformacje białek obecnych w aparacie wydzielniczym typu III w obecne u wici bakteryjnego wymagałyby wielu korzystnych
35 mutacji [dużej ilości poszczególnych kroków] i każda taka zmiana musiałaby dawać korzyści w postaci przewagi selekcyjnej. Takiej ewolucji nikt nie potrafi sobie wyobrazić i szczegółowo opisać w rzetelnym i naukowym modelu teoretycznym, ponieważ jest to po prostu niemożliwe. Taka postulowana transformacja musiałaby dokonać wielkich skoków ewolucyjnych i to równocześnie, na wielu poziomach organizacji strukturalnej, a to jest niemożliwe. [11]
36 [12]
37
38
39
40
41
42
43 Film dokumentalny omawiający między innymi problemy hipotezy kooptacji. Wypowiada się w nim również Michael Behe: [14]
44 [15]
45 [16] Forumowicz z racjonalisty.pl kontynuuje: [...]Ponadto wykazano zróżnicowanie budowy wici u różnych bakterii a w niektórych wykazano obecność tylko części białek tworzących wić. Wić zatem stała się dowodem na możliwość ewolucji. Zwolennicy ewolucjonizmu twierdzą, że neodarwinowskiego pochodzenia wici bakteryjnej dowodzą odkrycia różnych jej rodzajów pomiędzy którymi nie znaleziono homologii. Uznano więc, że te rózne silniki powstały niezależnie od sienie, w wyniku ewolucji zbieżnej [konwergencji]. Wyciągnięto z tego faktu
46 nieuprawniony wniosek: 'skoro wić powstała tyle razy w przyrodzie, to znaczy, że ewoluuje szybko i łatwo. Jest to argument oparty na błędzie logicznym. Neodarwiniści nie zaprezentowali, jak dotąd żadnego przekonującego modelu, który szczegółowo wyjaśniałby ewolucję wici bakteryjnej - jak można było się przekonać mamy do czynienia jedynie z takimi sobie bajeczkami. A więc ich błąd polega na próbach wspierania jednej hipotezy inną, która sama domaga się uzasadnienia; Znane są trzy typy wici, które mają całkowicie odmienną ultrastrukturę i nie wykazują homologii wici bakterii, archeonów i eukariontów. Budowa wici tych ostatnich stała się jedną z podstaw współczesnych podziałów systematycznych wyróżniających sześć supergrup jądrowców. Tutaj znalazłem pewną wypowiedź na ten temat: [..] "Po pierwsze, z funkcjonalną wicią połączony jest skomplikowany system kontrolny, który mówi jej, kiedy ma rotować i kiedy przestać, a czasem, kiedy się odwrócić i obracać się w przeciwnym kierunku. Umożliwia to bakterii poruszanie się w stronę odpowiedniego sygnału, jak i w stronę przeciwną, zamiast w kierunku przypadkowym, co ułatwiałoby jej popłynięcie w niewłaściwym kierunku. Problem wyjaśnienia pochodzenia wici nie ogranicza się zatem do niej samej, lecz obejmuje także sprzęgnięty z nią system kontrolny[...].
47 Zgadza się. Chodzi o mechanizm chemotaksji, który jest systemem nieredukowalnie złożonym. Można o nim poczytać tutaj:
48 I dalej: [...] Po drugie, subtelniejszy problem stanowi to, w jaki sposób poszczególne części wici złożyły się w jedną całość. Analogia do silnika zaburtowego jest nieadekwatna pod jednym względem: generalnie silnik zaburtowy montuje człowiek - inteligentny czynnik określający, które części mają się ze sobą łączyć. Jednakże informacja potrzebna do złożenia wici bakteryjnej (czy, w rzeczywistości, wszystkich pozostałych biologicznych mechanizmów molekularnych) zawiera się w białkach tworzących samą tę strukturę. Niedawno opublikowana praca pokazuje, że proces powstawania wici jest nadzwyczaj wyrafinowany i złożony. Gdy białka nie zawierają tej informacji, wić w ogóle nie powstaje. Dlatego, nawet jeśli mielibyśmy hipotetyczną komórkę, w której byłyby białka homologiczne do wszystkich części wici (być może wykonujące inne zadania niż napędzanie), lecz brakowałoby informacji mówiącej o tym, jak mają się one złożyć w wić, to i tak nie otrzymalibyśmy odpowiedniej struktury. Problem nieredukowalności nie zniknie. (Nieredukowalna złożoność: problem dla ewolucjonizmu darwinowskiego). Materiał do ściągmnięcia tutaj: action=tekst&id=70 To też jest prawda. Biogeneza [montaż w komórce] każdej złożonej maszyny komórkowej wymaga precyzyjnej sekwencji zdarzeń przebiegającej w określonej kolejności. Jakiekolwiek zakłócenia uniemożliwią prawidłowy przebieg tego procesu. Mechanizmy te nieźle poznano w przypadku innego silnika
49 molekularnego; syntazy ATP: [...] Modele składania syntazy ATP Kontrola procesu składania syntazy ATP jest niezbędna, by zapobiec tworzeniu się niekontrolowanego kanału przepływu protonów, którego obecność prowadziłaby do destabilizacji potencjału na wewnętrznej błonie mitochondrialnej i do śmierci komórki. Jest to proces niezwykle złożony, wymagający prawidłowego rozpoznania i integracji podjednostek, jak również aktywności wyżej opisanych białek wspierających etapy ekspresji podjednostek oraz koordynacji tych etapów ze składaniem kompleksu. Defekty funkcji tego enzymu spowodowane są więc nie tylko mutacjami w genach strukturalnych, ale także w genach kodujących czynniki wspomagające, działających na etapach transkrypcji, translacji, modyfikacji potranslacyjnych lub podczas składania enzymu.
50 Obecnie istnieją dwa modele proponujące kolejność składania podjednostek syntazy ATP w aktywny enzym. Uważa się, że syntetyzowane w macierzy białka Atp9 oligomeryzują tworząc pierścień, do którego dołączana jest domena F1 (Ryc. 2A). Do tego subkompleksu dołączana jest podjednostka Atp8, a potem ramię zewnętrzne enzymu. Na końcu składana jest podjednostka Atp6, co zabezpiecza przed tworzeniem się niekontrolowanego przez F1 kanału protonów i w konsekwencji niestabilności potencjału przez wewnętrzną błonę mitochondrialną. [..] [17] W przypadku syntazy ATP, czy wici bakteryjnej kontrola przebiegu biogenezy w komórce zaczyna się na poziomie regulacji ekspresji genów. W właściwym czasie i kolejności ekspresji ulegają odpowiednie geny, kodujące białka budujące te silniki, lub inne wspomagające ich montaż [białka pomocnicze], bez których cały proces utknąłby w martwym punkcie. Mamy więc do czynienia z jeszcze większą precyzją i nieredukowalną złożonością niż to się dawniej mogło wydawać.
51 Podsumowanie: - Według modelu kooptacji wić bakteryjna powstała poprzez połączenie się trzech elementów; a) Syntazy ATP, b) Aparatu wydzielniczego typu III, c) Elementów tworzących silnik obrotowym Wszystkie te trzy podzespoły z osobna są nieredukowalnie złożone, a wszystkie razem wzięte stanowią nadrzędny system nieredukowalnie złożony. Dopiero wówczas, kiedy występują w komplecie wić bakteryjna może spełniać swoje funkcje. A więc nie mogła ewoluować stopniowo, poprzez dodawanie kolejnych elementów. - System transportu białek [flagelin] do wici bakteryjnej stanowi mechanizm działający na zupełnie odmiennych zasadach niż kompleks odpowiedzialny za wprawianie wici bakteryjnej w ruch obrotowy. Tym samym podobieństwa strukturalne pomiędzy aparatem transportującym flageliny i wydzielniczym typu III nie wyjaśniają ewolucji silnika, wprawiającego w ruch wić bakteryjną. Poza tym podobieństwa nie muszą świadczyć o pokrewieństwach ewolucyjnych - równie dobrze można je interpretować, jako efekt projektu. - Model kooptacji w ewolucji wici bakteryjnej jest ogólnikowy i nie wyjaśnia krok po kroku, jak wyewoluowała wić bakteryjna. Jak widać na filmie omawiającym hipotezę kooptacji autorzy tego pomysłu nie wnikają w szczegóły i pomijają wiele trudnych etapów, których nie potrafią sobie wyobrazić. Takiej
52 powierzchownej opowieści nie można traktować, jako szczegółowego, naukowego modelu teoretycznego, tylko co najwyżej ciekawostkę. - Biogeneza wszystkich maszyn molekularnych, czy to rybosomów czy silników bakteryjnych, jest bardzo skomplikowana i precyzyjna. Jakiekolwiek zakłócenia powodują utratę funkcji. Świadczy to o istnieniu nieredukowalniej złożoności na wielu poziomach organizacji molekularnej, związanych z montażem i funkcjonowaniem tych urządzeń biologicznych. Poza tym mechanizm regulacji pracy silnika bakteryjnego [chemotaksja] również jest systemem nieredukowalnie złożonym. Warto zaznaczyć, że przebieg montażu wici bakteryjnej powinien w jakimś zakresie odzwierciedlać jej ewolucję, to znaczy kolejność poszczególnych etapów, na których niezbędne elementy stopniowo były dodawane do całego układu. Nie mogło być tak, żeby wszystko przebiegało według modelu kooptacji. Ten pomysł jest nieadekwatny do rzeczywistości. Jeżeli uczeni znokautują [spowodują, że przestanie ulegać ekspresji], jakiś istotny gen, odpowiedzialny za powstawanie białka wchodzącego w skład podstawowej budowy [nieredukowalnego rdzenia] wici bakteryjnej, to cały system się załamuje i silnik się nie wykształca. Jakże więc podczas rzekomej ewolucji tego urządzenia ewolucja mogła dodawać do całego systemu poszczególne elementy, przy czym niekompletna wić zachowywała funkcje? Jak mógł działać niekompletny silnik podczas różnych etapów ewolucji dając komórce przewagę selekcyjną?:
53
54 Dodatek: Argumentacja, z którą dyskutowałem w niniejszym poście nie stanowi nowości. Nie wiem z jakich źródeł korzystał autor wiadomo jedynie, że nie do końca rozumiał istotę argumentacji autorów, z których twórczości korzystał. W internecie można znaleźć tego typu wywody w niejednym miejscu. Np. w wikipedii. Za każdym razem autorzy różnych artykułów krytykujących koncepcję nieredukowalnej złożoności wici bakteryjnej powołują się jedynie na hipotezę kooptacji, zapewniają, że zadowalająco wyjaśnia ona genezę tego silnika molekularnego, jednak za każdym razem pomijają rzetelną jej analizę. Przykład: Podobnie wić bakteryjna, którą Behe wskazywał jako ewidentny przykład systemu nieredukowalnie złożonego, szybko okazała się składać z modułów mających samodzielne znaczenie funkcjonalne (np. "silnik" wici okazał się tożsamy z pompą używaną przez bakterie chorobotwórcze do wstrzykiwania toksyn), poszczególne białka samej wici (flageliny) są produktami zduplikowanych genów, a wiele innych elementów kompleksu wici ma odpowiedniki pełniące inne funkcje. Ponadto wykazano, że zgodnie z przewidywaniami neodarwinizmu występuje duża zmienność wici pomiędzy różnymi gatunkami bakterii (a zatem można zmieniać poszczególne elementy) oraz przykłady "narządów szczątkowych" wśród tych organelli. Okazało się też, że archebakterie mają wici podobne funkcjonalnie, jednak wyewoluowały niezależnie. Zazwyczaj złożone struktury powstają dzięki kooptacji
55 (przysposobieniu) istniejących, już złożonych, struktur (skrzydło powstające u owadów z narządów wymiany gazowej, u kręgowców z kończyn przednich), powielaniu modułów (przykładem jest duplikacja genu; szczególnym przypadkiem są duplikacje genów Hox, mogące powodować zwielokrotnienie danej struktury anatomicznej u zarodka) i innym procesów rozpoznanych przez biologię ewolucyjną. [18]
56 Źródła [1] 3.html (The bacterial flagellar motor: brilliant evolution or intelligent design? Tutaj można zapoznać się z bardziej szczegółową krytyką hipotezy kooptacji: [2] %2814% [3] cardsetid= [4] [5] sa=t&rct=j&q=&esrc=s&source=web&cd=1&cad=rja&uact=8&v ed=0ahukewiu5ljf9p7jahxjvhikhqxgah0qfggbmaa&url =http%3a%2f%2fwww.paleo.pan.pl%2fpeople%2fdzik %2FLectures %2FEwolucja_08.pdf&usg=AFQjCNELB0Z4oLDglY3BnVCBP N2NmFzt8w&bvm=bv ,d.bGQ
57 [6] [7] [8] [9] [10] [11] list=plpqnr5ubp_6e7ytbdsl1hnl_blk7hskoa&v=a_5fto P_mMY [14] "Odkrywanie Tajemnicy Życia" Lektor PL [15] 020 [16]
58 4/ss3.htm [17] O genetycznej regulacji ekspresji genów odpowiedzialnych za przebieg biogenezy wici bakteryjnej można poczytać tutaj: [18] %C5%BCono%C5%9B%C4%87 [19]
59
60 Dodatek: Hemotaksja Wykład na temat silnika bakteryjnego w jeżyku polskim:
61 Cz:2 Krytyka modularnej hipotezy ewolucji syntazy ATP Film: Syntaza ATP (polski lektor)
62 Hipoteza ewolucji modularnej syntazy ATP Uważa się, że ewolucja syntazy ATP zachodziła modułowo. Obie domeny wraz ze swoimi wartościowościami połączyły się zyskując nową funkcję. Domena F1 wykazuje znaczne podobieństwo do heksamerycznej helikazy DNA, a domena Fo jest podobna do kompleksów tworzących motor molekularny napędzający wici komórek. Heksamer α3β3 tworzący domenę F1 podobnie jak helikaza DNA składa się z obracającego się pierścienia z otworem pośrodku. U obu enzymów obroty pierścienia umożliwiają pełnienie funkcji. Helikaza DNA porusza się po helisie DNA przy jednoczesnej hydrolizie nukleotydów. Domena F1 wykorzystuje zmiany konformacyjne podczas obrotu podjednostki γ do przeprowadzenia reakcji enzymatycznej. Jony H+ przepływające przez domenę Fo w bardzo podobny sposób jak przy napędzaniu motoru molekularnego poruszającego wicią. Ich wspólną cechą jest pierścień składający się z wielu alfa-helikalnych białek, które obracają się w stosunku do innych białek zużywając jednocześnie gradient protonowy jako źródło energii. Jest to jednakże dość wątły związek, ogólna struktura motorów molekularnych jest zdecydowanie większa, zawierają 30 polipeptydów, w porównaniu do 10, 11, lub 14 znanych polipeptydów domeny Fo. Modułowa teoria pochodzenia syntazy ATP sugeruje, że dwie domeny o niezależnych funkcjach, helikaza DNA posiadająca właściwości ATPazy i motor molekularny napędzany siłą protonomotoryczną, mogły się połączyć i doprowadzić do odwrócenia właściwości ATPazy
63 powstałej z helikazy DNA. Dalszy rozwój prowadziłby do powstania kompleksy syntazy ATP znanej dzisiaj. Alternatywnie kompleks helikazy DNA i motoru molekularnego mógł wykazywać początkowo aktywność ATPazy, która przenosiła jony H+ zużywając ATP. Dalsza ewolucja kompleksy mogłaby doprowadzić do odwrócenia przeprowadzanej reakcji i powstania funkcji spełnianej przez syntazę ATP. [1] Więcej szczegółów Białka heksameryczne - odpowiedzialne za rozplatanie podwójnej helisy DNA czy DNA - RNA, za translokację kwasów nukleinowych, na przykład pakowanie ich do kapsydu bakteriofaga podczas jego biogenezy, pewne kompleksy odpowiedzialne za translokację polipeptydów na drugą stronę błony, czy podjednostka tworząca domenę F_1 służącą komórce do produkcji ATP, nie tylko są do siebie podobne strukturalnie, ale ponadto wszystkie jako rożnego rodzaju motory są zależne od hydrolizy ATP. Mimo różnych funkcji i zasadniczych różnić w budowie owe powierzchowne podobieństwa posłużyły ewolucjonistom molekularnym do wyciągnięcia życzeniowego wniosku, jakoby wszystkie te maszyny molekularne miały wspólnego przodka. Neodarwiniści zajmujący się tymi sprawami przyznają, że zrozumienie w jaki sposób te heksameryczne motory mogły pojawić się w trakcie ewolucji jest bardzo trudnym zadaniem. Syntaza ATP to kompleks złożony z dwóch domen: F_o i F_1. Pierwsza odpowiada za przepływ protonów z jednej strony błony
64 na drugą, które to po drodze łączą się z rotorem wprawiając go w ruch obrotowy. Do tego rotora przytwierdzony jest wał, który wnika do heksamerycznego agregatu tworzącego domenę F_1. Pierwszym etapem w powstaniu syntazy ATP miało być połączenie się helikazy z receptorem, który umożliwiał transport łańcucha DNA lub RNA na drugą stronę błony. Ewolucjoniści molekularni dokonali w tym momencie ogromnego skoku myślowego, ponieważ nie wyjaśnili, jak stopniowa ewolucja mogła przekształcić helikazę, maszynę molekularną, która służy komórce do rozplatania podwójnej helisy DNA lub DNA-RNA, w zupełnie inną maszynę służącą do przepychania pojedynczych łańcuchów nukleinowych przez ten kanał. Problemu tego etapu postulowanej ewolucji syntazy ATP nie będę bardziej szczegółowo tutaj rozważał, ponieważ temat ten będzie omówiony szerzej w rozdziale: problem mnogich mutacji. Kolejnym etap, jaki zakłada ten ogólnikowy i hipotetyczny scenariusz miał polegać na zmianie funkcji już wyewoluowanej translokazy. Z roli transportera łańcuchów nukleinowych na drugą stronę błony komórkowej, ewolucja miała ją przekwalifikować w translokazę transportującą polipeptydy (rozwinięte białka). W kolejnym kroku jeden z takich polipeptydów miał utknąć między białkiem heksamerycznym i kanałem przelotowym przez błonę i w przyszłości zamienić się w wyrafinowany wał, jaki napędza syntezę ATP we współczesnych komórkach. Człowiekowi, który jest niezaznajomiony z biologią molekularną taka opowieść może się wydawać poważną propozycją naukową
65 wyjaśniającą pochodzenie syntazy ATP. w dalszej części tego artykułu wykażę, że nie jest to naukowy, rzetelny i szczegółowy model teoretyczny, tylko zwykła bajeczka przypominająca okrzyk kuglarza, który wyciąga królika z kapelusza. [2]
66 Funkcje, jakie pełnią heksameryczne translokazy transportujące łańcuchy nukleinowe, lub polipeptydy mają bardzo szerokie zastosowanie w komórkach. Oto jeden z przykładów [3] Translokaza, która bierze udział w pakowaniu materiału genetycznego do kapsydy bakteriofaga podczas jego montażu w komórce
67 Która funkcja była pierwsza? Zastanówmy się skąd mógłby się wziąć ewolucyjny prekursor (wspólny przodek) wszystkich białek heksamerycznych i jaką funkcję pełnił. Załóżmy, że była nim helikaza. Helikaza wyczerpuje definicję kompleksu nieredukowalnie złożonego, ponieważ składa się z kilku ściśle dopasowanych elementów. Chociaż budują ją dwa rodzaje tych samych podzespołów białkowych, to są one ściśle do siebie dopasowane i precyzyjnie zespolone w celu pełnienia swojej roli. Ewolucja helikazy nie mogła więc następować stopniowo. Poza tym helikaza wchodzi w skład większego kompleksu enzymatycznego odpowiedzialnego za replikację DNA czy transkrypcję. To samo dotyczy syntazy
68 ATP, która jest ostatnim enzymem w łańcuchu transportu elektronów. [4]
69 W swojej polemice z Jerrym Coyne Michael Behe podsumował ten problem w następujący sposób: [5] (.)Wielu ewolucyjnych biologów bez chwili wahania sprzeciwiło się tej tezie. Książka Darwin s Black Box została szeroko zrecenzowana. W szczególności wielu znanych biologów ewolucyjnych, wszyscy zdeklarowani darwiniści, miało okazję aby ją ostro skrytykować. Być może najlepsza była dwustronicowa recenzja w Nature, najbardziej znanym czasopiśmie naukowym na świecie. Autorem był Jerry Coyne, profesor biologii ewolucyjnej na uniwersytecie w Chicago, i jak się okazało redaktor odpowiedzialny za recenzje książek w czasopiśmie Evolution, który namówił Goulda i Dawkinsa do zamieszczenia w swoim czasopiśmie recenzji swoich książek. (..) Jerry Coyne napisał: Odpowiedź na racje Behe a leży w uświadomieniu sobie, iż biochemiczne ścieżki [ ] zostały zmontowane z elementów dokooptowanych z innych ścieżek Trombina na przykład jest jedną z głównych protein odpowiedzialnych za krzepnięcie krwi, działa jednak również w procesie podziału komórek i jest powiązana z trawiennym enzymem trypsyną. Kto wie, która funkcja pojawiła się najpierw? Dobre pytanie kto wie, która funkcja pojawiła się najpierw? Nikt nie wie. Nikt także nie wie, w jaki sposób jedna funkcja może wyjaśniać drugą. Jest to jak mówienie, iż sprężynki znajdują się zarówno w zegarkach, jak i łapkach na myszy, tak więc być może jedno wyjaśnia drugie. Jednak zagadnienie, w jaki
70 sposób skomplikowane systemy biochemiczne zgromadziły się razem, tak naprawdę nie interesuje Coyne a. [6] Pod jakim wzgędem helikaza jest podobna do domeny f_1 a czym się od niej różni? Hydroliza (rozkładanie) ATP w miejscach aktywnych wywołuje u helikazy zmiany konformacyjne, na zasadzie wielopostaciowych skurczów elementów struktury tego molekularnego robota służącego do rozplatania DNA. Wydłużone domeny mają powinowactwo do DNA (lub DNA i RNA) i tam znajdują fizyczne oparcie. Podczas synchronicznych skurczów, polegających na zmianie konformacji odpowiednich domen, które można sobie wyobrazić, jako osobne motory, wywoływanych hydrolizą ATP helikaza wiruje w koło jednego łańcucha DNA i rozplata podwójną helisę. Strukturalnie domena F_1, agregat, który w syntazie ATP produkujący ATP z ADP, najbardziej przypomina helikazę Rho, biorącą udział w procesie transkrypcji. Helikaza Rho jest złożonym z sześciu podjednostek zależnym od ATP wyrafinowanym motorem molekularnym. Te sześć podjednostek działa niczym sześć cylindrów w silniku. Właśnie podjednostki te przyjmują wiele różnych, precyzyjnych konformacji, co w harmonijny sposób pozwala temu molekularnemu robotowi wiązać się z kwasem nukleinowym i znajdując w nim fizyczne oparcie, wirować w koło jednej z nici rozplatając podwójną helisę.
71 [7,8] Powyżej : struktura, wygląd oraz schemat działania helikazy, molekularnego robota, zasilanego energią pochodzącą z rozkładu ATP, który służy komórce do rozplatania DNA i RNA
72 Heksameryczna podjednostka F_1, służąca do syntezy ATP choć jet podobna strukturalnie do helikazy, to posiada zupełnie inne funkcjonalne domeny, które pozwalają jej komplementarnie dopasować się do siebie wzajemnie, oraz do wału,który łączy domenę F_1 z domeną F_o. Posiada ona również odpowiednie domeny, które pozwalają jej znaleźć fizyczne oparcie na stojanie. A więc w przeciwieństwie do helikazy domena F_1 jes tu rządzeniem stacjonarnym. Urządzenie do produkcji ATP składa się z następujących podjednostek: F1: α, β, δ, Υ, ε. Białka tworzące domenę F_1 występują w następujących ilości kopii α 3, β 3, δ 1, Υ 1, ε 1. Podjednostki α i
73 β są w pewnym sensie analogami (są do siebie bardzo podobne). Υ tworzy wał (układ rozrządu) wewnątrz pierścienia domeny F_1 utworzonego przez podjednostki α i β. U podstaw (od góry i dołu) podjednostki Υ znajdują się podjednostki δ i ε związane. z domeną F_1. Na podjednostkę F_o składają się białka c tworzące rotor w liczbie co najmniej 10 jednostek. Stator tworzy polipeptyd oznaczony a. Stojan, na którym zakotwiczona jest podjednostka F_1 oznaczamy bb. Wał łączący podjednostki F_1 i F_o wnika głęboko do podjednostki F_1 (α 3 i β 3), która jest sztywno zakotwiczona do przedłużenia statora, stojana oznaczonego symbolem b. Odpowiednio ukształtowany wał w formę układu rozrządu powoduje, że podczas jego obrotów podjednostka F_1 (α i β) synchronicznie zmienia kształty (konformacje), co umożliwia jej syntezę ATP z ADP. [9]
74 Podjednostka f_1 (α 3 i β 3), działa jak agregat. Agregat ten działa jak mechaniczna, wielofunkcyjna forma, która składa poszczególne elementy w jedną całość, w tym przypadku ADP+pi w ATP-jak pokazano na schemacie. Odwrotna praca podjednostki F_1 ( α 3 i β 3+y), rozkładanie ATP z powrotem do ADP, skutkuje tym, iż działa ona jak silnik, który napędzany energią uzyskaną z rozkładu (hydrolizy) ATP kręci wałem, a za jego pośrednictwem rotorem. więc podczas odwrotnej pracy domeny F_1 staje się ona silnikiem, natomiast domena F_o przejmuje funkcję agregatu. [10] Cząsteczka ATP
75 Czy w ogóle było możliwe aby helikaza mogła zacząć pełnić funkcję, jaką pełni domena F_1 ( α 3 i β 3)? W wyniku trywialnej zmiany w centrum aktywnym, która w przeszłości mogła się przyczynić do tego, aby odwrócić pierwotną możliwość helikazy pozwalającą hydrolizować (rozkładać) ATP do ADP do możliwości syntezy ATP z ADP? Odpowiedz na to pytanie jest stanowczo przecząca. Aby helikaza mogła stać się domeną F_1 (α i β) i aby mogła nabyć możliwości syntezy ATP, jako stacjonarny agregat, którego praca napędzana jest fizyczną siła rotacji, musiałaby przejść przez szereg skomplikowanych dostosowań. Przede wszystkim prekursor podjednostki F_1 ( α i β) musiałby wykształcić odpowiednie domeny, które poprzez fizyczny kontakt z odpowiednio ukształtowanym wałem (układem rozrządu) umożliwiałyby mu przyjmowanie odpowiednich konformacji, pozwalających na syntezę ATP z ADP. Praca domeny F_1, jako agregatu syntetyzującego ATP z ADP wcale nie jest taka prosta, jak niektórym może się wydawać. Nie polega ona tylko na przemiennym wiązaniu ADP+Pi i przy kolejnym cyklu obrotu wału uwalnianiu gotowego produktu w postaci ATP. Choć i taki cykl byłby wystarczająco złożony. Przebieg procesu syntezy ATP zademonstruję na poniższym schemacie, a następnie go omówię, wyciągając z tej wiedzy odpowiednie wnioski. [11]
76 Jak widać na uproszczonym schemacie wał wnikający do wnętrza podjednostki F_1 ( α i β) jest zbudowany asymetrycznie, co skutecznie umożliwia mu pełnienie funkcji układu rozrządu. W przypadku syntezy ATP wymagane są trzy etapy, w których domena f_1 ( α 3 i β 3) zmienia konformacje na trzy sposoby:. (1) Konfiguracja otwarta niskiego powinowactwa. Konfiguracja ta umożliwia łączenie się ADP+Pi oraz uwalnianie ATP. (2) Konfiguracja zawężająca, w której ADP+Pi są niejako dopasowywane do siebie przed ostatecznym złożeniem w kompletną cząsteczkę ATP. (3) Konfiguracja zamknięta. W tej pozycji domena katalityczna ostatecznie łączy ADP+Pi w wyniku czego powstaje gotowa do uwolnienia cząsteczka ATP.
77 Zmiany te umożliwia domenie F_1 ( α i β) precyzyjnie dopasowany odcinek wału (układ rozrządu), który wnika pomiędzy enzymy tworzące agregat do syntezy ATP ( α 3 i β 3) i harmonijnie oddziałuje z precyzyjnie ukształtowanymi domenami tych enzymów. Problem mnogich mutacji Mimo pewnych strukturalnych podobieństw pomiędzy helikazą, a domeną F_1 przekształcenie tej pierwszej w drugą wymagałoby serii mutacji, które musiałyby nastąpić jednocześnie. Prapodjednostka F_1 musiałaby nabyć zdolności do syntezy ATP oraz dopasować się do tych obszarów wału, które pozwalają na zmianę jej konformacji. Po drodze od helikazy do domeny F_1 miał się pojawić krok pośredni. Na tym etapie helikaza miała zmienić funkcję na translokazę, najpierw transportującą łańcuchy nukleinowe następnie polipeptydy. Sama możliwość zmiany kształtu podczas mechanicznego odziaływania wału wymaga specyficznej budowy domeny F_1. Innymi słowy ewolucja taka nie miała szans zajść stopniowo - za pośrednictwem doboru kumulatywnego. Żeby kwestia stała się jasna postaram się szerzej problem wyjaśnić. Wyobraź sobie, że do powstania jakieś niezbędnej korzystnej funkcji potrzebne są aż 4 mutacje: A,B,C,D. Że dopiero zestaw A,B,C,D może dać korzyść selekcyjną (dostosowanie). Jaka więc korzyść z pojedynczych mutacji: A, B, C czy D, skoro żadna z nich z osobna nie daje żadnej przewagi selekcyjnej? Każda z osobna jest neutralna, bezużyteczna? W tym przypadku, żeby cecha określana przez mutacje: A,B,C,D mogła
78 dać przewagę selekcyjną, to te 4 [kompletny zestaw] mutacje musiałyby nastąpić za jednym zamachem, a prawdopodobieństwo takiego zdarzenia jest znikome. Jeżeli każda z tych 4 mutacji z osobna nie da przewagi selekcyjnej, to taka ewolucja po prostu nie ma szans nastąpić, ponieważ wtedy nie zadziała dobór kumulatywny. Istnieje wiele cech, których funkcję określają nierozerwalne zestawy konkretnie ulokowanych w genach nukleotydów. Jeżeli mutacja wprowadzi zmiany w takich konserwatywnych sekwencjach gen przestaje spełniać swoje funkcje i organizm ginie [efekt letalny]. Wiele enzymów, czy białek strukturalnych, posiadają takie konserwatywne domeny (centra aktywne) i jakiekolwiek zaburzenia paraliżują ich funkcję. W podobny sposób zachowuje się enzym zatruty jakimś antybiotykiem. Kontakt z toksyną powoduje, że jego centrum aktywne traci swoją funkcję i przestaje pasować, jak klucz do zamka do konkretnego substratu. [12] Skoro więc do zachowania prawidłowej funkcji potrzeba kilku konkretnych nukleotydów, to jakże one mogły powstawać w wyniku mutacji, które następowały stopniowo, po kolei? Jak widzimy nie tylko nieredukowalna złożoność kompleksów
79 biochemicznych stanowi nieprzebytą przeszkodę dla neodarwinizmu, ale i problem mnogich mutacji [13] Podczas zakładanej ewolucji domeny F_1 z helikazy musiałyby zajść równocześnie wszystkie niezbędne mutacje, a nie tylko te dostosowujące ją do syntezy ATP z ADP - chodzi o centra aktywne, gdzie ADP łączy się z Pi. Musiałyby pojawić się równocześnie te mutacje, które pozwalałyby po zmianie konformacji podjednostki F_1 przyjąć centrom aktywnym odpowiednie kształty - konfiguracje: otwartą, zwężającą i zankniętą oraz wraz z tymi zmianami musiałyby pojawić się za jednym zamachem wszystkie mutacje dopasowujące wnętrze domeny F_1 do odziałującego z nią wału. To samo dotyczy samego wału, jak i reszty elementów budujących syntazę ATP. Syntaza ATP a koncepcja nieredukowalnego rdzenia Uczeni poddając odpowiednim mutacjom skrócili wał, tak iż wnikał on ledwie do podjednostki F_1, ale synteza ATP był kaleka (mało wydajna) choć zachowany był pełen obrót. Jednak wał taki można skracać tylko do pewnych granic, później już synteza ATP jest niemożliwa. Mimo skrócenia i tak musi on zachować odpowiednią konformację (kształt), ponieważ w przeciwnym razie synteza ATP nie zachodzi. Mimo skrócenia musi on pasować do górnych obszarów domen podjednostki F_1, które we współpracy z nim pozwalają mu działać na zasadzie układu rozrządu. A więc pewna forma zredukowanej domeny F_1 będzie funkcjonować,
80 niemniej z nieprawidłowo ukształtowanym wałem, lub bez wału domena ta syntetyzować ATP nie będzie. Doświadczenie to pokazuje, że w budowie syntazy ATP istnieją granice możliwości jej redukowania: nieredukowalny rdzeń (Tutaj jest praca źródłowa opisująca to doświadczenie: [14] Prawidłowy i skrócony wał
81 Naukowcy badając maszynę do syntezy ATP przeprowadzili kilka innych ciekawych doświadczeń. Otóż - przy zastosowaniu technik molekularnych - metalową kuleczkę przytwierdzono do rotora i przy pomocy magnesów wywołano sztucznie (bez współpracy ze statorem) ruch obrotowy domeny F_1, co spowodowało syntezę ATP. Z jednej strony udowodniono, że taka synteza jest możliwa bez przepływu protonów przez domenę F_o. Z drugiej strony eksperyment wykazał, że podjednostka F_1 bez energii mechanicznej, pochodzącej z podjednostki F_o nie potrafi syntetyzować ATP. Temat związany z proponowanym modelem ewolucji domeny F_1 omówiłem szeroko w pierwszej części niniejszego opracowania. Właściwie mógłbym na tym poprzestać, gdyż tego tematu dotyczy dyskusja. Postanowiłem jednak zamieścić w niniejszym opracowaniu kilka faktów odnośnie rozważań na temat ewolucji domeny F_o. W celu wyjaśniania wcześniejszych zagadnień posługiwałem się przemawiającymi do wyobraźni schematami poglądowymi. W przypadku rozważań nad możliwością ewolucji domeny F_o postąpię tak samo.
Najmniejszy silnik świata
 Marzenie nanotechnologów: silnik protonowy bakterii Escherichia coli. Fragmenty artykułu M. Ostrowskiego. Źródło: www.creationism.org.pl/artykuly/mostrowski2/ Silnik protonowy bakterii Escherichia coli
Marzenie nanotechnologów: silnik protonowy bakterii Escherichia coli. Fragmenty artykułu M. Ostrowskiego. Źródło: www.creationism.org.pl/artykuly/mostrowski2/ Silnik protonowy bakterii Escherichia coli
Można sobie wyobrazić, że ten kaleki ślimak w kolejnych pokoleniach nabiera możliwości coraz
 Czy zdarzyło Ci się kiedyś składać maszynkę do mięsa? Czy pozwolisz sobie wmówić, że mogłaby ona powstawać stopniowo poprzez dorabianie i dodawanie kolejnych elementów i na każdym etapie zachowywać swoją
Czy zdarzyło Ci się kiedyś składać maszynkę do mięsa? Czy pozwolisz sobie wmówić, że mogłaby ona powstawać stopniowo poprzez dorabianie i dodawanie kolejnych elementów i na każdym etapie zachowywać swoją
O doborach jednorazowym i kumulatywnym w ewolucji O hipotezie selekcyjnego wymiatania
 O doborach jednorazowym i kumulatywnym w ewolucji O hipotezie selekcyjnego wymiatania Neodarwiniści twierdzą, że drobne zmiany mikroewolucyjne prowadzą do większych zmian makroewolucyjnych. Jest to spekulacja
O doborach jednorazowym i kumulatywnym w ewolucji O hipotezie selekcyjnego wymiatania Neodarwiniści twierdzą, że drobne zmiany mikroewolucyjne prowadzą do większych zmian makroewolucyjnych. Jest to spekulacja
października 2013: Elementarz biologii molekularnej. Wykład nr 2 BIOINFORMATYKA rok II
 10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
Dr. habil. Anna Salek International Bio-Consulting 1 Germany
 1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Transkrypcja RNA
 SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Transkrypcja RNA SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy. 1. Karta
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Transkrypcja RNA SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy. 1. Karta
Historia informacji genetycznej. Jak ewolucja tworzy nową informację (z ma ą dygresją).
.") Historia informacji genetycznej. Jak ewolucja tworzy nową informację (z ma ą dygresją). Czym jest życie? metabolizm + informacja (replikacja) 2 Cząsteczki organiczne mog y powstać w atmosferze pierwotnej
Historia informacji genetycznej. Jak ewolucja tworzy nową informację (z ma ą dygresją). Czym jest życie? metabolizm + informacja (replikacja) 2 Cząsteczki organiczne mog y powstać w atmosferze pierwotnej
Transport przez błony
 Transport przez błony Transport bierny Nie wymaga nakładu energii Transport aktywny Wymaga nakładu energii Dyfuzja prosta Dyfuzja ułatwiona Przenośniki Kanały jonowe Transport przez pory w błonie jądrowej
Transport przez błony Transport bierny Nie wymaga nakładu energii Transport aktywny Wymaga nakładu energii Dyfuzja prosta Dyfuzja ułatwiona Przenośniki Kanały jonowe Transport przez pory w błonie jądrowej
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Teoria ewolucji. Podstawowe pojęcia. Wspólne pochodzenie.
 Teoria ewolucji Podstawowe pojęcia. Wspólne pochodzenie. Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Teoria ewolucji Podstawowe pojęcia. Wspólne pochodzenie. Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Translacja i proteom komórki
 Translacja i proteom komórki 1. Kod genetyczny 2. Budowa rybosomów 3. Inicjacja translacji 4. Elongacja translacji 5. Terminacja translacji 6. Potranslacyjne zmiany polipeptydów 7. Translacja a retikulum
Translacja i proteom komórki 1. Kod genetyczny 2. Budowa rybosomów 3. Inicjacja translacji 4. Elongacja translacji 5. Terminacja translacji 6. Potranslacyjne zmiany polipeptydów 7. Translacja a retikulum
Wykład 14 Biosynteza białek
 BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Treść tego doniesienia mocno mi podpadła, więc sprowokowalem polemikę. Do tablicy poczuł się wezwany autor badań!
 https://www.facebook.com/naukawpolsce/posts/1832898566757138? comment_id=1833531986693796&reply_comment_id=1837161782997483¬if_id=1532294745 151071¬if_t=feed_comment_reply Treść tego doniesienia
https://www.facebook.com/naukawpolsce/posts/1832898566757138? comment_id=1833531986693796&reply_comment_id=1837161782997483¬if_id=1532294745 151071¬if_t=feed_comment_reply Treść tego doniesienia
Przemiana materii i energii - Biologia.net.pl
 Ogół przemian biochemicznych, które zachodzą w komórce składają się na jej metabolizm. Wyróżnia się dwa antagonistyczne procesy metabolizmu: anabolizm i katabolizm. Szlak metaboliczny w komórce, to szereg
Ogół przemian biochemicznych, które zachodzą w komórce składają się na jej metabolizm. Wyróżnia się dwa antagonistyczne procesy metabolizmu: anabolizm i katabolizm. Szlak metaboliczny w komórce, to szereg
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU
 SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Czy priony zawsze są szkodliwe? SPIS TREŚCI: Wprowadzenie. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. Karty pracy. 1.
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Czy priony zawsze są szkodliwe? SPIS TREŚCI: Wprowadzenie. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. Karty pracy. 1.
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 5 Droga od genu do
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 5 Droga od genu do
TATA box. Enhancery. CGCG ekson intron ekson intron ekson CZĘŚĆ KODUJĄCA GENU TERMINATOR. Elementy regulatorowe
 Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Weronika Łabaj. Geometria Bolyaia-Łobaczewskiego
 Weronika Łabaj Geometria Bolyaia-Łobaczewskiego Tematem mojej pracy jest geometria hiperboliczna, od nazwisk jej twórców nazywana też geometrią Bolyaia-Łobaczewskiego. Mimo, że odkryto ją dopiero w XIX
Weronika Łabaj Geometria Bolyaia-Łobaczewskiego Tematem mojej pracy jest geometria hiperboliczna, od nazwisk jej twórców nazywana też geometrią Bolyaia-Łobaczewskiego. Mimo, że odkryto ją dopiero w XIX
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU. Czym są choroby prionowe?
 SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Czym są choroby prionowe? SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy.
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Czym są choroby prionowe? SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy.
Właściwości błony komórkowej
 Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność Transport przez błony Cząsteczki < 150Da Błony - selektywnie przepuszczalne RóŜnice składu jonowego między wnętrzem komórki ssaka
Właściwości błony komórkowej płynność asymetria selektywna przepuszczalność Transport przez błony Cząsteczki < 150Da Błony - selektywnie przepuszczalne RóŜnice składu jonowego między wnętrzem komórki ssaka
Scenariusz lekcji przyrody/biologii (2 jednostki lekcyjne)
") Joanna Wieczorek Scenariusz lekcji przyrody/biologii (2 jednostki lekcyjne) Strona 1 Temat: Budowa i funkcje kwasów nukleinowych Cel ogólny lekcji: Poznanie budowy i funkcji: DNA i RNA Cele szczegółowe:
Joanna Wieczorek Scenariusz lekcji przyrody/biologii (2 jednostki lekcyjne) Strona 1 Temat: Budowa i funkcje kwasów nukleinowych Cel ogólny lekcji: Poznanie budowy i funkcji: DNA i RNA Cele szczegółowe:
Jak powstają nowe gatunki. Katarzyna Gontek
 Jak powstają nowe gatunki Katarzyna Gontek Powstawanie gatunków (specjacja) to proces biologiczny, w wyniku którego powstają nowe gatunki organizmów. Zachodzi na skutek wytworzenia się bariery rozrodczej
Jak powstają nowe gatunki Katarzyna Gontek Powstawanie gatunków (specjacja) to proces biologiczny, w wyniku którego powstają nowe gatunki organizmów. Zachodzi na skutek wytworzenia się bariery rozrodczej
Teoria ewolucji. Podstawy wspólne pochodzenie.
 Teoria ewolucji. Podstawy wspólne pochodzenie. Ewolucja biologiczna } Znaczenie ogólne: } proces zmian informacji genetycznej (częstości i rodzaju alleli), } które to zmiany są przekazywane z pokolenia
Teoria ewolucji. Podstawy wspólne pochodzenie. Ewolucja biologiczna } Znaczenie ogólne: } proces zmian informacji genetycznej (częstości i rodzaju alleli), } które to zmiany są przekazywane z pokolenia
Ruch zwiększa recykling komórkowy Natura i wychowanie
 Wiadomości naukowe o chorobie Huntingtona. Prostym językiem. Napisane przez naukowców. Dla globalnej społeczności HD. Ruch zwiększa recykling komórkowy Ćwiczenia potęgują recykling komórkowy u myszy. Czy
Wiadomości naukowe o chorobie Huntingtona. Prostym językiem. Napisane przez naukowców. Dla globalnej społeczności HD. Ruch zwiększa recykling komórkowy Ćwiczenia potęgują recykling komórkowy u myszy. Czy
Transportowane cząsteczki CO O, 2, NO, H O, etanol, mocznik... Zgodnie z gradientem: stężenia elektrochemicznym gradient stężeń
 Transportowane cząsteczki Transport przez błony Transport bierny szybkość transportu gradien t stężeń kanał nośnik Transport z udziałem nośnika: dyfuzja prosta dyfuzja prosta CO 2, O 2, NO,, H 2 O, etanol,
Transportowane cząsteczki Transport przez błony Transport bierny szybkość transportu gradien t stężeń kanał nośnik Transport z udziałem nośnika: dyfuzja prosta dyfuzja prosta CO 2, O 2, NO,, H 2 O, etanol,
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii
 Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
POLIMERAZY DNA- PROCARYOTA
 Enzymy DNA-zależne, katalizujące syntezę DNA wykazują aktywność polimerazy zawsze w kierunku 5 3 wykazują aktywność polimerazy zawsze wobec jednoniciowej cząsteczki DNA do utworzenia kompleksu z ssdna
Enzymy DNA-zależne, katalizujące syntezę DNA wykazują aktywność polimerazy zawsze w kierunku 5 3 wykazują aktywność polimerazy zawsze wobec jednoniciowej cząsteczki DNA do utworzenia kompleksu z ssdna
Ewolucjonizm NEODARWINIZM. Dr Jacek Francikowski Uniwersyteckie Towarzystwo Naukowe Uniwersytet Śląski w Katowicach
 Ewolucjonizm NEODARWINIZM Dr Jacek Francikowski Uniwersyteckie Towarzystwo Naukowe Uniwersytet Śląski w Katowicach Główne paradygmaty biologii Wspólne początki życia Komórka jako podstawowo jednostka funkcjonalna
Ewolucjonizm NEODARWINIZM Dr Jacek Francikowski Uniwersyteckie Towarzystwo Naukowe Uniwersytet Śląski w Katowicach Główne paradygmaty biologii Wspólne początki życia Komórka jako podstawowo jednostka funkcjonalna
Teoria ewolucji. Podstawowe pojęcia. Wspólne pochodzenie.
 Teoria ewolucji Podstawowe pojęcia. Wspólne pochodzenie. Ewolucja Znaczenie ogólne: zmiany zachodzące stopniowo w czasie W biologii ewolucja biologiczna W astronomii i kosmologii ewolucja gwiazd i wszechświata
Teoria ewolucji Podstawowe pojęcia. Wspólne pochodzenie. Ewolucja Znaczenie ogólne: zmiany zachodzące stopniowo w czasie W biologii ewolucja biologiczna W astronomii i kosmologii ewolucja gwiazd i wszechświata
Dopasowanie sekwencji (sequence alignment)
") Co to jest alignment? Dopasowanie sekwencji (sequence alignment) Alignment jest sposobem dopasowania struktur pierwszorzędowych DNA, RNA lub białek do zidentyfikowanych regionów w celu określenia podobieństwa;
Co to jest alignment? Dopasowanie sekwencji (sequence alignment) Alignment jest sposobem dopasowania struktur pierwszorzędowych DNA, RNA lub białek do zidentyfikowanych regionów w celu określenia podobieństwa;
Geny i działania na nich
 Metody bioinformatyki Geny i działania na nich prof. dr hab. Jan Mulawka Trzy królestwa w biologii Prokaryota organizmy, których komórki nie zawierają jądra, np. bakterie Eukaryota - organizmy, których
Metody bioinformatyki Geny i działania na nich prof. dr hab. Jan Mulawka Trzy królestwa w biologii Prokaryota organizmy, których komórki nie zawierają jądra, np. bakterie Eukaryota - organizmy, których
Komórka eukariotyczna
 Komórka eukariotyczna http://pl.wikipedia.org/w/index.php?title=plik:hela_cells_stained_with_hoechst_33258.jpg cytoplazma + jądro komórkowe = protoplazma W cytoplazmie odbywa się: cała przemiana materii,
Komórka eukariotyczna http://pl.wikipedia.org/w/index.php?title=plik:hela_cells_stained_with_hoechst_33258.jpg cytoplazma + jądro komórkowe = protoplazma W cytoplazmie odbywa się: cała przemiana materii,
Temat: Komórka jako podstawowa jednostka strukturalna i funkcjonalna organizmu utrwalenie wiadomości.
 SCENARIUSZ LEKCJI BIOLOGII DLA KLASY I GIMNAZJUM Temat: Komórka jako podstawowa jednostka strukturalna i funkcjonalna organizmu utrwalenie wiadomości. Cele: Utrwalenie pojęć związanych z budową komórki;
SCENARIUSZ LEKCJI BIOLOGII DLA KLASY I GIMNAZJUM Temat: Komórka jako podstawowa jednostka strukturalna i funkcjonalna organizmu utrwalenie wiadomości. Cele: Utrwalenie pojęć związanych z budową komórki;
CORAZ BLIŻEJ ISTOTY ŻYCIA WERSJA A. imię i nazwisko :. klasa :.. ilość punktów :.
 CORAZ BLIŻEJ ISTOTY ŻYCIA WERSJA A imię i nazwisko :. klasa :.. ilość punktów :. Zadanie 1 Przeanalizuj schemat i wykonaj polecenia. a. Wymień cztery struktury występujące zarówno w komórce roślinnej,
CORAZ BLIŻEJ ISTOTY ŻYCIA WERSJA A imię i nazwisko :. klasa :.. ilość punktów :. Zadanie 1 Przeanalizuj schemat i wykonaj polecenia. a. Wymień cztery struktury występujące zarówno w komórce roślinnej,
Zarówno u organizmów eukariotycznych, jak i prokariotycznych proces replikacji ma charakter semikonserwatywny.
 HIPTEZY WYJAŚIAJĄCE MECHAIZM REPLIKACJI C. Model replikacji semikonserwatywnej zakłada on, że obie nici macierzystej cząsteczki DA są matrycą dla nowych, dosyntetyzowywanych nici REPLIKACJA każda z dwóch
HIPTEZY WYJAŚIAJĄCE MECHAIZM REPLIKACJI C. Model replikacji semikonserwatywnej zakłada on, że obie nici macierzystej cząsteczki DA są matrycą dla nowych, dosyntetyzowywanych nici REPLIKACJA każda z dwóch
Zacznijmy więc pracę z repozytorium. Pierwsza konieczna rzecz do rozpoczęcia pracy z repozytorium, to zalogowanie się w serwisie:
 Repozytorium służy do przechowywania plików powstających przy pracy nad projektami we w miarę usystematyzowany sposób. Sam mechanizm repozytorium jest zbliżony do działania systemu plików, czyli składa
Repozytorium służy do przechowywania plików powstających przy pracy nad projektami we w miarę usystematyzowany sposób. Sam mechanizm repozytorium jest zbliżony do działania systemu plików, czyli składa
Wymagania edukacyjne
 Rok szkolny 2018/2019 Wymagania edukacyjne Przedmiot Klasa Nauczyciel uczący Poziom biologia 1t Edyta Nowak podstawowy Ocena dopuszczająca Ocenę dopuszczającą otrzymuje uczeń, który: przyswoił treści konieczne,
Rok szkolny 2018/2019 Wymagania edukacyjne Przedmiot Klasa Nauczyciel uczący Poziom biologia 1t Edyta Nowak podstawowy Ocena dopuszczająca Ocenę dopuszczającą otrzymuje uczeń, który: przyswoił treści konieczne,
Informacje dotyczące pracy kontrolnej
 Informacje dotyczące pracy kontrolnej Słuchacze, którzy z przyczyn usprawiedliwionych nie przystąpili do pracy kontrolnej lub otrzymali z niej ocenę negatywną zobowiązani są do dnia 06 grudnia 2015 r.
Informacje dotyczące pracy kontrolnej Słuchacze, którzy z przyczyn usprawiedliwionych nie przystąpili do pracy kontrolnej lub otrzymali z niej ocenę negatywną zobowiązani są do dnia 06 grudnia 2015 r.
Algorytm genetyczny (genetic algorithm)-
-") Optymalizacja W praktyce inżynierskiej często zachodzi potrzeba znalezienia parametrów, dla których system/urządzenie będzie działać w sposób optymalny. Klasyczne podejście do optymalizacji: sformułowanie
Optymalizacja W praktyce inżynierskiej często zachodzi potrzeba znalezienia parametrów, dla których system/urządzenie będzie działać w sposób optymalny. Klasyczne podejście do optymalizacji: sformułowanie
Podstawy biologii. Informacja, struktura i metabolizm.
 Podstawy biologii Informacja, struktura i metabolizm. Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Podstawy biologii Informacja, struktura i metabolizm. Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
 Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
Od redakcji. Symbolem oznaczono zadania wykraczające poza zakres materiału omówionego w podręczniku Fizyka z plusem cz. 1.
 Od redakcji Niniejszy zbiór zadań powstał z myślą o tych wszystkich, dla których rozwiązanie zadania z fizyki nie polega wyłącznie na mechanicznym przekształceniu wzorów i podstawieniu do nich danych.
Od redakcji Niniejszy zbiór zadań powstał z myślą o tych wszystkich, dla których rozwiązanie zadania z fizyki nie polega wyłącznie na mechanicznym przekształceniu wzorów i podstawieniu do nich danych.
TEORIA KOMÓRKI (dlaczego istnieją osobniki?)
") Wstęp do biologii 2. TEORIA KOMÓRKI (dlaczego istnieją osobniki?) Jerzy Dzik Instytut Paleobiologii PAN Instytut Zoologii UW 2015 WSPÓLNE WŁAŚCIWOŚCI dzisiejszych organizmów procesy życiowe katalizowane
Wstęp do biologii 2. TEORIA KOMÓRKI (dlaczego istnieją osobniki?) Jerzy Dzik Instytut Paleobiologii PAN Instytut Zoologii UW 2015 WSPÓLNE WŁAŚCIWOŚCI dzisiejszych organizmów procesy życiowe katalizowane
Zawartość. Wstęp 1. Historia wirusologii. 2. Klasyfikacja wirusów
 Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
lider projektu: finansowanie:
 lider projektu: finansowanie: - zapoznanie się z możliwościami budowania programów w Lego Mindstorms EV3 - budowa prostego robota z jednym silnikiem i jednym czujnikiem - naładowane zestawy Lego Mindstorms
lider projektu: finansowanie: - zapoznanie się z możliwościami budowania programów w Lego Mindstorms EV3 - budowa prostego robota z jednym silnikiem i jednym czujnikiem - naładowane zestawy Lego Mindstorms
Podstawy biologii. Informacja genetyczna. Co to jest ewolucja.
 Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Materiał genetyczny Materiałem genetycznym są kwasy nukleinowe Materiałem genetycznym organizmów komórkowych jest kwas deoksyrybonukleinowy
Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Materiał genetyczny Materiałem genetycznym są kwasy nukleinowe Materiałem genetycznym organizmów komórkowych jest kwas deoksyrybonukleinowy
Zajęcia nr. 3 notatki
 Zajęcia nr. 3 notatki 22 kwietnia 2005 1 Funkcje liczbowe wprowadzenie Istnieje nieskończenie wiele funkcji w matematyce. W dodaktu nie wszystkie są liczbowe. Rozpatruje się funkcje które pobierają argumenty
Zajęcia nr. 3 notatki 22 kwietnia 2005 1 Funkcje liczbowe wprowadzenie Istnieje nieskończenie wiele funkcji w matematyce. W dodaktu nie wszystkie są liczbowe. Rozpatruje się funkcje które pobierają argumenty
Na początek przyjrzymy się więc, jak komórka rośliny produkuje ATP, korzystając z energii światła w fazie jasnej fotosyntezy.
 Fotosynteza jako forma biosyntezy Bogactwo molekuł biologicznych przedstawionych w poprzednim rozdziale to efekt ich wytwarzania w komórkach w wyniku różnorodnych powiązanych ze sobą procesów chemicznych.
Fotosynteza jako forma biosyntezy Bogactwo molekuł biologicznych przedstawionych w poprzednim rozdziale to efekt ich wytwarzania w komórkach w wyniku różnorodnych powiązanych ze sobą procesów chemicznych.
Uczeń potrafi. Dział Rozdział Temat lekcji
 Plan wynikowy z biologii- zakres podstawowy, dla klasy III LO i III i IV Technikum LO im.ks. Jerzego Popiełuszki oraz Technikum w Suchowoli Nauczyciel: Katarzyna Kotiuk Nr programu: DKOS-4015-5/02 Dział
Plan wynikowy z biologii- zakres podstawowy, dla klasy III LO i III i IV Technikum LO im.ks. Jerzego Popiełuszki oraz Technikum w Suchowoli Nauczyciel: Katarzyna Kotiuk Nr programu: DKOS-4015-5/02 Dział
Transport makrocząsteczek
 Komórka eukariotyczna cytoplazma + jądro komórkowe = protoplazma W cytoplazmie odbywa się: cała przemiana materii, dzięki której organizm uzyskuje energię biosynteza białka i innych związków Transport
Komórka eukariotyczna cytoplazma + jądro komórkowe = protoplazma W cytoplazmie odbywa się: cała przemiana materii, dzięki której organizm uzyskuje energię biosynteza białka i innych związków Transport
Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny
 Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny Zadanie 1 1 pkt. za prawidłowe podanie typów dla obydwu zwierząt oznaczonych literami A oraz B. A. ramienionogi, B. mięczaki A.
Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny Zadanie 1 1 pkt. za prawidłowe podanie typów dla obydwu zwierząt oznaczonych literami A oraz B. A. ramienionogi, B. mięczaki A.
Komputerowe wspomaganie projektowanie leków
 Komputerowe wspomaganie projektowanie leków wykład VI Prof. dr hab. Sławomir Filipek Grupa BIOmodelowania Uniwersytet Warszawski, Wydział Chemii oraz Centrum Nauk Biologiczno-Chemicznych Cent-III www.biomodellab.eu
Komputerowe wspomaganie projektowanie leków wykład VI Prof. dr hab. Sławomir Filipek Grupa BIOmodelowania Uniwersytet Warszawski, Wydział Chemii oraz Centrum Nauk Biologiczno-Chemicznych Cent-III www.biomodellab.eu
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych
 Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Interfaza to niemal 90% cyklu komórkowego. Dzieli się na 3 fazy: G1, S i G2.
 W wyniku podziału komórki powstaje komórka potomna, która ma o połowę mniej DNA od komórki macierzystej i jest o połowę mniejsza. Aby komórka potomna była zdolna do kolejnego podziału musi osiągnąć rozmiary
W wyniku podziału komórki powstaje komórka potomna, która ma o połowę mniej DNA od komórki macierzystej i jest o połowę mniejsza. Aby komórka potomna była zdolna do kolejnego podziału musi osiągnąć rozmiary
(węglowodanów i tłuszczów) Podstawowym produktem (nośnikiem energii) - ATP
 Podstawowym produktem (nośnikiem energii) - ATP") śycie - wymaga nakładu energii źródłem - promienie świetlne - wykorzystywane do fotosyntezy - magazynowanie energii w wiązaniach chemicznych Wszystkie organizmy (a zwierzęce wyłącznie) pozyskują energię
śycie - wymaga nakładu energii źródłem - promienie świetlne - wykorzystywane do fotosyntezy - magazynowanie energii w wiązaniach chemicznych Wszystkie organizmy (a zwierzęce wyłącznie) pozyskują energię
Znaleźć wzór ogólny i zbadać istnienie granicy ciągu określonego rekurencyjnie:
 Ciągi rekurencyjne Zadanie 1 Znaleźć wzór ogólny i zbadać istnienie granicy ciągu określonego rekurencyjnie: w dwóch przypadkach: dla i, oraz dla i. Wskazówka Należy poszukiwać rozwiązania w postaci, gdzie
Ciągi rekurencyjne Zadanie 1 Znaleźć wzór ogólny i zbadać istnienie granicy ciągu określonego rekurencyjnie: w dwóch przypadkach: dla i, oraz dla i. Wskazówka Należy poszukiwać rozwiązania w postaci, gdzie
Metoda 5-WHY. Metoda 5-WHY. Wydanie 1. Zbigniew Huber. Maj 2006. Artykuł dostępny na stronie autora: http://www.huber.pl.
 Metoda 5-WHY Wydanie 1 Zbigniew Huber Maj 2006 Artykuł dostępny na stronie autora: http://www.huber.pl Copyright by Zbigniew Huber Strona 1 z 6 Wstęp Rozwiązanie jakiegoś problemu i wprowadzenie skutecznego
Metoda 5-WHY Wydanie 1 Zbigniew Huber Maj 2006 Artykuł dostępny na stronie autora: http://www.huber.pl Copyright by Zbigniew Huber Strona 1 z 6 Wstęp Rozwiązanie jakiegoś problemu i wprowadzenie skutecznego
Informacje. Kontakt: Paweł Golik, Ewa Bartnik. Instytut Genetyki i Biotechnologii, Pawińskiego 5A.
 Podstawy genetyki Informacje Kontakt: Paweł Golik, Ewa Bartnik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl; ebartnik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Podstawy genetyki Informacje Kontakt: Paweł Golik, Ewa Bartnik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl; ebartnik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Sposoby przedstawiania algorytmów
 Temat 1. Sposoby przedstawiania algorytmów Realizacja podstawy programowej 5. 1) wyjaśnia pojęcie algorytmu, podaje odpowiednie przykłady algorytmów rozwiązywania różnych problemów; 2) formułuje ścisły
Temat 1. Sposoby przedstawiania algorytmów Realizacja podstawy programowej 5. 1) wyjaśnia pojęcie algorytmu, podaje odpowiednie przykłady algorytmów rozwiązywania różnych problemów; 2) formułuje ścisły
Podstawy biologii. Informacja genetyczna. Co to jest ewolucja.
 Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Zarys biologii molekularnej genu Podstawowe procesy genetyczne Replikacja powielanie informacji Ekspresja wyrażanie (realizowanie funkcji)
Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Zarys biologii molekularnej genu Podstawowe procesy genetyczne Replikacja powielanie informacji Ekspresja wyrażanie (realizowanie funkcji)
Programowanie i techniki algorytmiczne
 Temat 2. Programowanie i techniki algorytmiczne Realizacja podstawy programowej 1) wyjaśnia pojęcie algorytmu, podaje odpowiednie przykłady algorytmów rozwiązywania różnych 2) formułuje ścisły opis prostej
Temat 2. Programowanie i techniki algorytmiczne Realizacja podstawy programowej 1) wyjaśnia pojęcie algorytmu, podaje odpowiednie przykłady algorytmów rozwiązywania różnych 2) formułuje ścisły opis prostej
6. Z pięciowęglowego cukru prostego, zasady azotowej i reszty kwasu fosforowego, jest zbudowany A. nukleotyd. B. aminokwas. C. enzym. D. wielocukier.
 ID Testu: F5679R8 Imię i nazwisko ucznia Klasa Data 1. Na indywidualne cechy danego osobnika ma (maja) wpływ A. wyłacznie czynniki środowiskowe. B. czynniki środowiskowe i materiał genetyczny. C. wyłacznie
ID Testu: F5679R8 Imię i nazwisko ucznia Klasa Data 1. Na indywidualne cechy danego osobnika ma (maja) wpływ A. wyłacznie czynniki środowiskowe. B. czynniki środowiskowe i materiał genetyczny. C. wyłacznie
Ćwiczenie 14. Maria Bełtowska-Brzezinska KINETYKA REAKCJI ENZYMATYCZNYCH
 Ćwiczenie 14 aria Bełtowska-Brzezinska KINETYKA REAKCJI ENZYATYCZNYCH Zagadnienia: Podstawowe pojęcia kinetyki chemicznej (szybkość reakcji, reakcje elementarne, rząd reakcji). Równania kinetyczne prostych
Ćwiczenie 14 aria Bełtowska-Brzezinska KINETYKA REAKCJI ENZYATYCZNYCH Zagadnienia: Podstawowe pojęcia kinetyki chemicznej (szybkość reakcji, reakcje elementarne, rząd reakcji). Równania kinetyczne prostych
Recenzja pracy doktorskiej mgr Tomasza Świsłockiego pt. Wpływ oddziaływań dipolowych na własności spinorowego kondensatu rubidowego
 Prof. dr hab. Jan Mostowski Instytut Fizyki PAN Warszawa Warszawa, 15 listopada 2010 r. Recenzja pracy doktorskiej mgr Tomasza Świsłockiego pt. Wpływ oddziaływań dipolowych na własności spinorowego kondensatu
Prof. dr hab. Jan Mostowski Instytut Fizyki PAN Warszawa Warszawa, 15 listopada 2010 r. Recenzja pracy doktorskiej mgr Tomasza Świsłockiego pt. Wpływ oddziaływań dipolowych na własności spinorowego kondensatu
Bioinformatyka Laboratorium, 30h. Michał Bereta
 Bioinformatyka Laboratorium, 30h Michał Bereta mbereta@pk.edu.pl www.michalbereta.pl 1 Często dopasować chcemy nie dwie sekwencje ale kilkanaście lub więcej 2 Istnieją dokładne algorytmy, lecz są one niewydajne
Bioinformatyka Laboratorium, 30h Michał Bereta mbereta@pk.edu.pl www.michalbereta.pl 1 Często dopasować chcemy nie dwie sekwencje ale kilkanaście lub więcej 2 Istnieją dokładne algorytmy, lecz są one niewydajne
TRANSKRYPCJA - I etap ekspresji genów
 Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
Książka kucharska, przepis i ciasto z wiśniami
 Wiadomości naukowe o chorobie Huntingtona. Prostym językiem. Napisane przez naukowców. Dla globalnej społeczności HD. Ryzykowne splatanie: nowe spojrzenie na szkodliwe białko choroby Huntingtona Badacze
Wiadomości naukowe o chorobie Huntingtona. Prostym językiem. Napisane przez naukowców. Dla globalnej społeczności HD. Ryzykowne splatanie: nowe spojrzenie na szkodliwe białko choroby Huntingtona Badacze
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Budowa rybosomu Translacja
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Budowa rybosomu Translacja
użytkownika 1 Jak wybrać temat pracy 2 Spis treści 3 Część pierwsza problematyka 4 Część druga stosowane metody 5 Część trzecia propozycja rozwiązania
 1 Jak wybrać temat pracy 2 Spis treści 3 Część pierwsza problematyka 4 Część druga stosowane metody 5 Część trzecia propozycja rozwiązania 6 Część czwarta dokumentacja techniczna i dokumentacja użytkownika
1 Jak wybrać temat pracy 2 Spis treści 3 Część pierwsza problematyka 4 Część druga stosowane metody 5 Część trzecia propozycja rozwiązania 6 Część czwarta dokumentacja techniczna i dokumentacja użytkownika
Katarzyna Wojewoda-Buraczyńska Koncepcja multicentryczności prawa a derywacyjne argumenty systemowe. Studenckie Zeszyty Naukowe 9/13, 84-87
 Katarzyna Wojewoda-Buraczyńska Koncepcja multicentryczności prawa a derywacyjne argumenty systemowe Studenckie Zeszyty Naukowe 9/13, 84-87 2006 Katarzyna Wojewoda-Buraczyńska Koncepcja multicentryczności
Katarzyna Wojewoda-Buraczyńska Koncepcja multicentryczności prawa a derywacyjne argumenty systemowe Studenckie Zeszyty Naukowe 9/13, 84-87 2006 Katarzyna Wojewoda-Buraczyńska Koncepcja multicentryczności
166 Wstęp do statystyki matematycznej
 166 Wstęp do statystyki matematycznej Etap trzeci realizacji procesu analizy danych statystycznych w zasadzie powinien rozwiązać nasz zasadniczy problem związany z identyfikacją cechy populacji generalnej
166 Wstęp do statystyki matematycznej Etap trzeci realizacji procesu analizy danych statystycznych w zasadzie powinien rozwiązać nasz zasadniczy problem związany z identyfikacją cechy populacji generalnej
Kamila Muraszkowska Znaczenie wąskich gardeł w sieciach białkowych. źródło: (3)
") Kamila Muraszkowska Znaczenie wąskich gardeł w sieciach białkowych źródło: (3) Interakcje białko-białko Ze względu na zadanie: strukturalne lub funkcjonalne. Ze względu na właściwości fizyczne: stałe lub
Kamila Muraszkowska Znaczenie wąskich gardeł w sieciach białkowych źródło: (3) Interakcje białko-białko Ze względu na zadanie: strukturalne lub funkcjonalne. Ze względu na właściwości fizyczne: stałe lub
Pamiętając o komplementarności zasad azotowych, dopisz sekwencję nukleotydów brakującej nici DNA. A C C G T G C C A A T C G A...
 1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
dr hab. Mikołaj Olejniczak, prof. UAM Zakład Biochemii 16 grudnia 2018, Poznań
 dr hab. Mikołaj Olejniczak, prof. UAM Zakład Biochemii 16 grudnia 2018, Poznań Recenzja rozprawy doktorskiej mgr Michała Rażewa zatytułowanej Badania strukturalne drożdżowego degradosomu mitochondrialnego
dr hab. Mikołaj Olejniczak, prof. UAM Zakład Biochemii 16 grudnia 2018, Poznań Recenzja rozprawy doktorskiej mgr Michała Rażewa zatytułowanej Badania strukturalne drożdżowego degradosomu mitochondrialnego
Podstawy mikrobiologii. Wirusy bezkomórkowe formy materii oŝywionej
 Podstawy mikrobiologii Wykład 3 Wirusy bezkomórkowe formy materii oŝywionej Budowa wirusów Wirusy nie mają budowy komórkowej, zatem pod względem biologicznym nie są organizmami Ŝywymi! Są to twory nukleinowo
Podstawy mikrobiologii Wykład 3 Wirusy bezkomórkowe formy materii oŝywionej Budowa wirusów Wirusy nie mają budowy komórkowej, zatem pod względem biologicznym nie są organizmami Ŝywymi! Są to twory nukleinowo
WPROWADZENIE DO GENETYKI MOLEKULARNEJ
 WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
WPROWADZENIE DO GENETYKI MOLEKULARNEJ
 WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
Chemiczne składniki komórek
 Chemiczne składniki komórek Pierwiastki chemiczne w komórkach: - makroelementy (pierwiastki biogenne) H, O, C, N, S, P Ca, Mg, K, Na, Cl >1% suchej masy - mikroelementy Fe, Cu, Mn, Mo, B, Zn, Co, J, F
Chemiczne składniki komórek Pierwiastki chemiczne w komórkach: - makroelementy (pierwiastki biogenne) H, O, C, N, S, P Ca, Mg, K, Na, Cl >1% suchej masy - mikroelementy Fe, Cu, Mn, Mo, B, Zn, Co, J, F
TEORIA KOMÓRKI (dlaczego istnieją osobniki?)
") Wstęp do biologii 2. TEORIA KOMÓRKI (dlaczego istnieją osobniki?) Jerzy Dzik Instytut Paleobiologii PAN Instytut Zoologii UW 2017 WSPÓLNE WŁAŚCIWOŚCI dzisiejszych organizmów procesy życiowe katalizowane
Wstęp do biologii 2. TEORIA KOMÓRKI (dlaczego istnieją osobniki?) Jerzy Dzik Instytut Paleobiologii PAN Instytut Zoologii UW 2017 WSPÓLNE WŁAŚCIWOŚCI dzisiejszych organizmów procesy życiowe katalizowane
dr Anna Mazur Wyższa Szkoła Promocji Intuicja a systemy przekonań
 dr Anna Mazur Wyższa Szkoła Promocji Intuicja a systemy przekonań Systemy przekonań Dlaczego mądrzy ludzie podejmują głupie decyzje? Odpowiedzialne są nasze przekonania. Przekonania, które składają się
dr Anna Mazur Wyższa Szkoła Promocji Intuicja a systemy przekonań Systemy przekonań Dlaczego mądrzy ludzie podejmują głupie decyzje? Odpowiedzialne są nasze przekonania. Przekonania, które składają się
znajdowały się różne instrukcje) to tak naprawdę definicja funkcji main.
 to tak naprawdę definicja funkcji main.") Część XVI C++ Funkcje Jeśli nasz program rozrósł się już do kilkudziesięciu linijek, warto pomyśleć o jego podziale na mniejsze części. Poznajmy więc funkcje. Szybko się przekonamy, że funkcja to bardzo
Część XVI C++ Funkcje Jeśli nasz program rozrósł się już do kilkudziesięciu linijek, warto pomyśleć o jego podziale na mniejsze części. Poznajmy więc funkcje. Szybko się przekonamy, że funkcja to bardzo
Wprowadzenie do biologii molekularnej.
 Wprowadzenie do biologii molekularnej. Materiały dydaktyczne współfinansowane ze środków Unii Europejskiej w ramach Europejskiego Funduszu Społecznego. Biologia molekularna zajmuje się badaniem biologicznych
Wprowadzenie do biologii molekularnej. Materiały dydaktyczne współfinansowane ze środków Unii Europejskiej w ramach Europejskiego Funduszu Społecznego. Biologia molekularna zajmuje się badaniem biologicznych
Jak obracać trójkąt, by otrzymać bryłę o największej. objętości?
 Jak obracać trójkąt, by otrzymać bryłę o największej objętości? Czas trwania zajęć: 40 minut Kontekst w jakim wprowadzono doświadczenie: Trójkąt o bokach długości: cm, 4 cm, 5 cm obrócono o kąt 60 o w
Jak obracać trójkąt, by otrzymać bryłę o największej objętości? Czas trwania zajęć: 40 minut Kontekst w jakim wprowadzono doświadczenie: Trójkąt o bokach długości: cm, 4 cm, 5 cm obrócono o kąt 60 o w
W. Guzicki Próbna matura, grudzień 2014 r. poziom rozszerzony 1
 W. Guzicki Próbna matura, grudzień 01 r. poziom rozszerzony 1 Próbna matura rozszerzona (jesień 01 r.) Zadanie 18 kilka innych rozwiązań Wojciech Guzicki Zadanie 18. Okno na poddaszu ma mieć kształt trapezu
W. Guzicki Próbna matura, grudzień 01 r. poziom rozszerzony 1 Próbna matura rozszerzona (jesień 01 r.) Zadanie 18 kilka innych rozwiązań Wojciech Guzicki Zadanie 18. Okno na poddaszu ma mieć kształt trapezu
ETAPY PROCESU BADAWCZEGO. wg Babińskiego
 ETAPY PROCESU BADAWCZEGO wg Babińskiego NA ZACHĘTĘ Ludowe porzekadło mówi: CIEKAKAWOŚĆ TO PIERWSZY STOPIEŃ DO PIEKŁA. ale BEZ CIEKAWOŚCI I CHĘCI POZNANIA NIE MA Nauki Badań Rozwoju I jeszcze kilku ciekawych
ETAPY PROCESU BADAWCZEGO wg Babińskiego NA ZACHĘTĘ Ludowe porzekadło mówi: CIEKAKAWOŚĆ TO PIERWSZY STOPIEŃ DO PIEKŁA. ale BEZ CIEKAWOŚCI I CHĘCI POZNANIA NIE MA Nauki Badań Rozwoju I jeszcze kilku ciekawych
Dominika Stelmach Gr. 10B2
 Dominika Stelmach Gr. 10B2 Czym jest DNA? Wielkocząsteczkowy organiczny związek chemiczny z grupy kwasów nukleinowych Zawiera kwas deoksyrybonukleoinowy U organizmów eukariotycznych zlokalizowany w jądrze
Dominika Stelmach Gr. 10B2 Czym jest DNA? Wielkocząsteczkowy organiczny związek chemiczny z grupy kwasów nukleinowych Zawiera kwas deoksyrybonukleoinowy U organizmów eukariotycznych zlokalizowany w jądrze
Inżynieria genetyczna- 6 ECTS. Inżynieria genetyczna. Podstawowe pojęcia Część II Klonowanie ekspresyjne Od genu do białka
 Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Projektowanie systemu krok po kroku
 Rozdział jedenast y Projektowanie systemu krok po kroku Projektowanie systemu transakcyjnego jest ciągłym szeregiem wzajemnie powiązanych decyzji, z których każda oferuje pewien zysk i pewien koszt. Twórca
Rozdział jedenast y Projektowanie systemu krok po kroku Projektowanie systemu transakcyjnego jest ciągłym szeregiem wzajemnie powiązanych decyzji, z których każda oferuje pewien zysk i pewien koszt. Twórca
DNA musi współdziałać z białkami!
 DNA musi współdziałać z białkami! Specyficzność oddziaływań między DNA a białkami wiążącymi DNA zależy od: zmian konformacyjnych wzdłuż cząsteczki DNA zróżnicowania struktury DNA wynikającego z sekwencji
DNA musi współdziałać z białkami! Specyficzność oddziaływań między DNA a białkami wiążącymi DNA zależy od: zmian konformacyjnych wzdłuż cząsteczki DNA zróżnicowania struktury DNA wynikającego z sekwencji
OCENA Rozprawy doktorskiej mgr Aksany Varabyovej Biogeneza dysmutazy ponadtlenkowej 1 w mitochondrialnej przestrzeni międzybłonowej
 prof. dr hab. Barbara Zabłocka Pracownia Biologii Molekularnej Instytut Medycyny Doświadczalnej i Klinicznej im. M. Mossakowskiego PAN ul. Pawińskiego 5, 02-106 Warszawa tel: 22-60 86 486 e-mail: bzablocka@imdik.pan.pl
prof. dr hab. Barbara Zabłocka Pracownia Biologii Molekularnej Instytut Medycyny Doświadczalnej i Klinicznej im. M. Mossakowskiego PAN ul. Pawińskiego 5, 02-106 Warszawa tel: 22-60 86 486 e-mail: bzablocka@imdik.pan.pl
Matematyk Ci powie, co łączy Eugeniusza Oniegina i gry hazardowe
 Matematyk Ci powie, co łączy Eugeniusza Oniegina i gry hazardowe Empik każdego inspiruje inaczej Aleksander Puszkin (1799 1837) Andrey (Andrei) Andreyevich Markov (1856 1922) Wśród 20 tysięcy początkowych
Matematyk Ci powie, co łączy Eugeniusza Oniegina i gry hazardowe Empik każdego inspiruje inaczej Aleksander Puszkin (1799 1837) Andrey (Andrei) Andreyevich Markov (1856 1922) Wśród 20 tysięcy początkowych
ROZPORZĄDZENIE KOMISJI (UE) / z dnia r.
 / z dnia r.") KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
PRZEKROJE RYSUNKOWE CZ.1 PRZEKROJE PROSTE. Opracował : Robert Urbanik Zespół Szkół Mechanicznych w Opolu
 PRZEKROJE RYSUNKOWE CZ.1 PRZEKROJE PROSTE Opracował : Robert Urbanik Zespół Szkół Mechanicznych w Opolu IDEA PRZEKROJU stosujemy, aby odzwierciedlić wewnętrzne, niewidoczne z zewnątrz, kształty przedmiotu.
PRZEKROJE RYSUNKOWE CZ.1 PRZEKROJE PROSTE Opracował : Robert Urbanik Zespół Szkół Mechanicznych w Opolu IDEA PRZEKROJU stosujemy, aby odzwierciedlić wewnętrzne, niewidoczne z zewnątrz, kształty przedmiotu.
SCENARIUSZ LEKCJI. TEMAT LEKCJI: Podstawowe techniki inżynierii genetycznej. Streszczenie
 SCENARIUSZ LEKCJI OPRACOWANY W RAMACH PROJEKTU: INFORMATYKA MÓJ SPOSÓB NA POZNANIE I OPISANIE ŚWIATA. PROGRAM NAUCZANIA INFORMATYKI Z ELEMENTAMI PRZEDMIOTÓW MATEMATYCZNO-PRZYRODNICZYCH Autorzy scenariusza:
SCENARIUSZ LEKCJI OPRACOWANY W RAMACH PROJEKTU: INFORMATYKA MÓJ SPOSÓB NA POZNANIE I OPISANIE ŚWIATA. PROGRAM NAUCZANIA INFORMATYKI Z ELEMENTAMI PRZEDMIOTÓW MATEMATYCZNO-PRZYRODNICZYCH Autorzy scenariusza:
plezjomorfie: podobieństwa dziedziczone po dalszych przodkach (c. atawistyczna)
") Podobieństwa pomiędzy organizmami - cechy homologiczne: podobieństwa wynikające z dziedziczenia - apomorfie: podobieństwa dziedziczone po najbliższym przodku lub pojawiająca się de novo (c. ewolucyjnie
Podobieństwa pomiędzy organizmami - cechy homologiczne: podobieństwa wynikające z dziedziczenia - apomorfie: podobieństwa dziedziczone po najbliższym przodku lub pojawiająca się de novo (c. ewolucyjnie
Trajektoria rzuconego ukośnie granatu w układzie odniesienia skręcającego samolotu
 Politechnika Łódzka FTIMS Kierunek: Informatyka rok akademicki: 2009/2010 sem. 3. grupa II Termin: 10 XI 2009 Zadanie: Trajektoria rzuconego ukośnie granatu w układzie odniesienia skręcającego samolotu
Politechnika Łódzka FTIMS Kierunek: Informatyka rok akademicki: 2009/2010 sem. 3. grupa II Termin: 10 XI 2009 Zadanie: Trajektoria rzuconego ukośnie granatu w układzie odniesienia skręcającego samolotu
Pętle. Dodał Administrator niedziela, 14 marzec :27
 Pętlami nazywamy konstrukcje języka, które pozwalają na wielokrotne wykonywanie powtarzających się instrukcji. Przykładowo, jeśli trzeba 10 razy wyświetlić na ekranie pewien napis, to można wykorzystać
Pętlami nazywamy konstrukcje języka, które pozwalają na wielokrotne wykonywanie powtarzających się instrukcji. Przykładowo, jeśli trzeba 10 razy wyświetlić na ekranie pewien napis, to można wykorzystać
POLIMERAZY DNA- PROCARYOTA
 Enzymy DNA-zależne, katalizujące syntezę DNA wykazują aktywność polimerazy zawsze w kierunku 5 3 wykazują aktywność polimerazy zawsze wobec jednoniciowej cząsteczki DNA do utworzenia kompleksu z ssdna
Enzymy DNA-zależne, katalizujące syntezę DNA wykazują aktywność polimerazy zawsze w kierunku 5 3 wykazują aktywność polimerazy zawsze wobec jednoniciowej cząsteczki DNA do utworzenia kompleksu z ssdna
Uniwersytet Zielonogórski Instytut Sterowania i Systemów Informatycznych. Ćwiczenie 3 stos Laboratorium Metod i Języków Programowania
 Uniwersytet Zielonogórski Instytut Sterowania i Systemów Informatycznych Ćwiczenie 3 stos Laboratorium Metod i Języków Programowania Celem ćwiczenia jest zapoznanie studentów z najprostszą dynamiczną strukturą
Uniwersytet Zielonogórski Instytut Sterowania i Systemów Informatycznych Ćwiczenie 3 stos Laboratorium Metod i Języków Programowania Celem ćwiczenia jest zapoznanie studentów z najprostszą dynamiczną strukturą