Podstawy genetyki populacji. Genetyka mendlowska i ewolucja.
|
|
|
- Kornelia Zawadzka
- 6 lat temu
- Przeglądów:
Transkrypt
1 Podstawy genetyki populacji Genetyka mendlowska i ewolucja.


2 Podręczniki



3 Dla zainteresowanych
4 Ewolucja biologiczna Znaczenie ogólne: proces zmian informacji genetycznej organizmów (częstości i rodzaju alleli w populacji), które to zmiany są przekazywane z pokolenia na pokolenie dotyczy populacji, nie pojedynczego osobnika dotyczy zmian dziedziczonych Dziedziczenie z modyfikacją (descent with modification - Darwin)
5 Ewolucja biologiczna Zjawisko (fakt) Teoria ewolucji Historia zmian ewolucyjnych
6 Syntetyczna teoria ewolucji Pierwsza synteza: połączenie teorii ewolucji Darwina z genetyką mendlowską na poziomie populacji W naturalnych populacjach występują różne allele genów Częstość cech fenotypowych w populacji zależy od częstości alleli i genotypów Ewolucja jako zmiana częstości alleli w populacji z pokolenia na pokolenie

7 Populacja Grupa krzyżujących się ze sobą osobników oraz ich potomstwo Zbiór wszystkich alleli populacji pula genowa
8 Najprostszy model Populacja N organizmów diploidalnych Rozważany jeden A gen o dwóch allelach A1 i A2 Częstości alleli, odpowiednio p i q p + q = 1
9 Populacja w stanie równowagi Liczebność populacji bardzo duża (N ~ ) Całkowicie losowe krzyżowanie (panmiksja) Sukces reprodukcyjny nie zależy od genotypu genu A Brak migracji Nie zachodzą mutacje zmieniające A1 w A2 i vice versa
10 Równowaga Hardy ego-weinberga Jeżeli częstości alleli A1 i A2 to odpowiednio p i q to częstości genotypów A1A1 p 2 A1A2 pq + qp = 2pq A2A2 q 2
11 Równowaga Hardy ego-weinberga Częstości alleli w populacji w stanie równowagi się nie zmieniają Gamety A1 - wszystkie gamety homozygot A1A1 i połowa gamet heterozygot A1A2 W kolejnym pokoleniu: p = p pq 2 = p2 + pq q = 1 p p' = p 2 + p (1 p) = p 2 + p p 2 = p
12
13 Równowaga Hardy ego-weinberga W populacji będącej w równowadze H-W częstości alleli nie zmieniają się Nie przebiega ewolucja Mechanizmy zaburzające równowagę H-W mogą być mechanizmami ewolucji
14 Mechanizmy zmieniające częstość alleli Mutacje Dobór Migracje Dryf
15 Dobór Dostosowanie (w) prawdopodobieństwo odniesienia sukcesu reprodukcyjnego przez osobnika o danym genotypie A1A1 : w11 A1A2 : w12 A2A2 : w22 w = 1 s gdzie s to współczynnik selekcji Nie ma znaczenia, czy chodzi o prawdopodobieństwo przeżycia, czy o liczbę wyprodukowanych gamet, czy o kondycję potomstwa itp. walka o byt uproszczona i niekiedy myląca metafora
16 Silna i słaba selekcja - symulacje
17 Mutacje i dobór Mutacje stają się istotną siłą w ewolucji gdy: działa dobór naturalny działa dryf genetyczny (populacje o skończonym N)
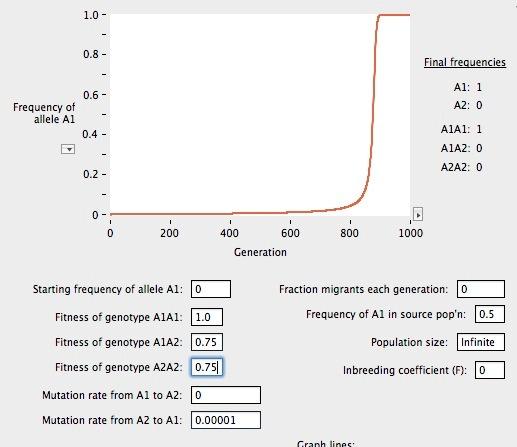
18
19 Równowaga mutacje-selekcja Większość mutacji obniża dostosowanie, dobór je eliminuje Wytwarza się równowaga, utrzymująca w populacji pulę allelu o szkodliwym działaniu Dla allelu recesywnego ˆq = µ s Dla dominującego allelu letalnego ˆq = µ
20 Dryf genetyczny a ewolucja Dobór naturalny nie jest jedynym mechanizmem kształtującym zmiany ewolucyjne Losowe procesy w populacjach o skończonej liczebności dryf genetyczny
21
22 Dryf genetyczny W populacjach o skończonej liczebności może dochodzić do zmian częstości alleli nawet jeżeli nie działa na nie dobór Nowy allel (mutacja) może się utrwalić w populacji nawet bez selekcji częściowo (polimorfizm) całkowicie
23 Dryf a wielkość populacji Efekty dryfu genetycznego są wyraźniejsze w populacjach o mniejszej wielkości Z czasem dryf doprowadzi do utraty jednego z alleli i utrwalenia drugiego utrata heterozygotyczności
24
25 Utrata heterozygotyczności Przy braku działania doboru dryf doprowadzi do utraty jednego allelu i utrwalenia (fiksacji) drugiego Może powodować powstanie populacji odmiennych genetycznie, bez udziału doboru
26 Utrata heterozygotyczności H = H 1 1 t 0 2N t czas półtrwania heterozygotycznności: H = 1 t 2 H przy t = 2N ln 1 0 1,39N 2 S. Wright, 1931
27 Efektywna wielkość populacji We wszystkich modelach zakładaliśmy panmiksję jednakowe prawdopodobieństwo wydania potomstwa przez każdego osobnika Rzeczywiste populacje nie spełniają tego warunku nierównomierne stosunki płci (haremy) zróżnicowanie sukcesu reprodukcyjnego
28 Efektywna wielkość populacji Efektywna wielkość populacji Ne jest to liczebność idealnej populacji panmiktycznej, w której tempo dryfu byłoby takie same, jak w badanej populacji o rzeczywistej liczebności N We wszystkich dotychczasowych rozważaniach podając N tak naprawdę mieliśmy na myśli Ne
29 Utrwalenie allelu Prawdopodobieństwo utrwalenia konkretnego allelu W populacji N osobników diploidalnych jest 2N alleli Utrwalenie oznacza, że wszystkie allele obecne w populacji pochodzą od jednego Prawdopodobieństwo tego jest 1/2N Jeżeli częstość allelu jest p, to wyjściowo jest 2Np kopii Czyli prawdopodobieństwo utrwalenia wynosi: 2Np 1/2N = p
30 Dryf i mutacje Mutacja powoduje powstanie nowego allelu Przy założeniu braku doboru (neutralność) Prawdopodobieństwo, że nowy allel się utrwali wynosi 1/2N Utrwalanie się kolejnych mutacji powoduje ewolucję populacji ewolucja neutralna
31 Tempo ewolucji neutralnej Prawdopodobieństwo utrwalenia mutacji neutralnej: ½N Prawdopodobieństwo powstania zmutowanego allelu: 2Nµ (µ - tempo mutacji) Prawdopodobieństwo powstania i utrwalenia się zmutowanego allelu (tempo ewolucji neutralnej)
32 Czas i częstość utrwalania alleli neutralnych Tempo ewolucji neutralnej odpowiada częstości mutacji Czas od powstania do utrwalenia mutacji średnio 4N (2N u haploidów)
33 Ewolucja neutralna Dryf zmniejsza różnorodność alleli (prowadzi do utrwalania jednego z alleli) Mutacje powodują powstawanie nowych alleli Dzięki temu różnorodność zostanie zachowana, ale skład konkretnych alleli się będzie zmieniał
34 Dryf i dobór Dryf może doprowadzić do utraty allelu korzystnego, albo do utrwalenia allelu niekorzystnego Równowaga między dryfem a doborem zależy od wielkości populacji i siły (współczynnika) selekcji Prosty model (kodominacja) A1A2 A1A2 A2A2 w 1 1+s 1+2s
35 Dryf i dobór Prosty model (kodominacja) A1A2 A1A2 A2A2 w 1 1+s 1+2s Model nie jest trywialny do wyprowadzenia (Kimura 1962) Rezultat:
36 Dryf i dobór Gdy s 0 to P q (prawdopodobieństwo utrwalenia allelu neutralnego jest równe jego częstości) (Dla nowej mutacji q=1/2ne) P = 1 e 4 N esq 1 e 4 N es
37 Dryf i dobór allele korzystne Jeżeli s > 0 i N jest duże to P 2s 98% mutacji o s = 0,01 się nie utrwali (Dla nowej mutacji q=1/2ne) P = 1 e 4 N esq 1 e 4 N es
38 Mutacja nieznacznie korzystna (Dla nowej mutacji q=1/2ne)
39 Mutacja wyraźnie korzystna (Dla nowej mutacji q=1/2ne) Efekt praktycznie nie zależy od Ne
40 Mutacja nieznacznie niekorzystna (Dla nowej mutacji q=1/2ne)
41 Mutacja bardzo nieznacznie niekorzystna (Dla nowej mutacji q=1/2ne)
42 Dlaczego mała populacja to zagrożenie Dla małych Ne bardzo wyraźnie spada skuteczność eliminacji alleli niekorzystnych (doboru oczyszczającego) Za to wzrost skuteczności utrwalania alleli korzystnych (doboru dodatniego) nie jest tak istotny Szczególnie krytyczne dla alleli w niewielkim stopniu niekorzystnych
43 Dryf i dobór Tempo utrwalania mutacji neutralnych Tempo utrwalania mutacji korzystnych 2Nµ 1 2N = µ
44 Dryf i dobór - podsumowanie Większość mutacji (korzystnych, neutralnych i niekorzystnych) nie utrwali się w populacji Gdy dobór przeciwko allelowi niekorzystnemu jest nieznaczny mutacja szkodliwa jest efektywnie neutralna zostanie utrwalona z prawdopodobieństwem takim, jak neutralna Dobór jest nieznaczny gdy: s 1 4N e
45 Dryf i dobór równowaga Gdy Ne jest duże, mutacje szkodliwe są skutecznie usuwane Gdy Ne jest małe, dryf może prowadzić do akumulacji mutacji szkodliwych! Nawet gdy Ne jest duże, wiele mutacji korzystnych jest traconych, jeżeli s nie jest bardzo duże
46 Druga synteza - ewolucja molekularna Ewolucja jako zmiany sekwencji DNA (i białek) Mechanizmy ewolucji molekularnej źródła zmienności - mutacje i rearanżacje DNA mechanizmy kształtujące zmienność - dobór i dryf Początki - lata 60. i 70. XX w. (sekwencjonowanie DNA , wcześniej sekwencje peptydowe)
47 Mutacje Podstawienia (substytucje) Niewielkie delecje i insercje niewielkie tzn. wpływające na sekwencję 1-2 genów
48 Substytucje Tranzycje zachodzą w naturze częściej od transwersji mimo tego, że możliwych transwersji jest więcej stosunek ts/tv od ~2 (ndna) do ~15 (mtdna człowieka) wyjątek mtdna roślin duże różnice ts/tv u różnych grup organizmów
49 Tranzycje i transwersje Dlaczego tranzycje są częstsze? Wyjaśnienia mechanistyczne mechanizmy powstawania i naprawy mutacji Tranzycje powstają w wyniku m. in.: przejść tautomerycznych deaminacji (np. oksydacyjnej) Tranzycje w mniejszym stopniu zaburzają strukturę podwójnej helisy podczas replikacji mniejsza wydajność naprawy przez system MMR
50 Modele ewolucji sekwencji Badając ewolucję nie dysponujemy z reguły sekwencją przodka Liczbę mutacji musimy oszacować na podstawie różnic między sekwencjami współczesnymi Konieczne jest uwzględnienie wielokrotnych mutacji w tej samej pozycji, zwłaszcza dla bardziej odległych sekwencji
51 Problem obliczania odległości (liczby zdarzeń mutacyjnych) ACGGTGC C A GCGGTGA
52 Modele ewolucji sekwencji Modele Markova stan w pokoleniu n +1 zależy tylko od stanu w pokoleniu n i reguł przekształcenia (macierz prawdopodobieństw zmiany stanów) Modele o różnym stopniu skomplikowania Mogą uwzględniać: mutacje wielokrotne w tej samej pozycji (poprawka Poissona) różne prawdopodobieństwa zmian nukleotydowych (lub białkowych) różne prawdopodobieństwo mutacji w różnych pozycjach sekwencji różne częstości nukleotydów
53 Modele ewolucji DNA model Jukesa-Cantora A C G T A 1-3α α α α C α 1-3α α α G α α 1-3α α T α α α 1-3α D JC = 3 4 ln(1 4 3 D)
54 Inne modele Kimura (K80, dwuparametrowy) - różne prawdopodobieństwo tranzycji i transwersji Felsenstein (F81), Hasegawa-Kishino-Yano (HKY85) - różne częstości nukleotydów (F81) + różne prawd. tranzycji i transwersji (HKY85) GTR (General Time Reversible, Tavare 86)
55 Ewolucja sekwencji aminokwasowych Trudno stworzyć model analityczny złożoność kodu aminokwasy o różnych właściwościach - konieczna miara niepodobieństwa Stosuje się empirycznie uzyskiwane macierze prawdopodobieństwa zmiany danego aminokwasu w inny
56 Tempo zmian sekwencji białka PAM - utrwalone mutacje punktowe/100 pozycji (od Point Accepted Mutation) Granica istotności Różnice sekwencji 80% 60% 40% 20% PAM
57 Istotność podobieństwa
 Na podstawie globalnych")



 Margaret O.")
58 Porównywanie białek - macierze Macierze Dayhoff (PAM) Na podstawie globalnych porównań sekwencji różniących się o 1PAM ustalono prawdopodobieństwo zmiany każdego aminokwasu w inny macierz PAM-1 Ekstrapolacja dla sekwencji bardziej odległych - mnożenie macierzy PAM-1 przez samą siebie odpowiednią liczbę razy macierze PAM-20, PAM-40, PAM-250 itp. (proces Markova) Margaret O. Dayhoff ( )

59 Porównywanie białek - macierze Macierze BLOSUM Na podstawie prawdopodobieństwa zmiany każdego aminokwasu w inny w bloku lokalnego przyrównania sekwencji o n% identycznych aminokwasów (BLOSUM62-62% identycznych aa itp.)
60 Mutacje i dobór naturalny Efekty działania mutacji obserwujemy pośrednio różnice sekwencji między populacjami (gatunkami) polimorfizm sekwencji w obrębie populacji Na allele wytworzone przez mutacje może działać dobór Za zmiany częstości powstających alleli może odpowiadać dryf genetyczny Obserwujemy mutacje utrwalone całkowicie lub częściowo (polimorfizmy) w puli genowej
61 Podstawowe pytanie ewolucji molekularnej Jaka jest rola dryfu i doboru w wyjaśnieniu obserwowanego zróżnicowania sekwencji? wewnątrzpopulacyjnego (polimorfizmy) międzygatunkowego Pytanie dotyczy zróżnicowania ilościowego! Nikt nie podaje w wątpliwość tego, że adaptacje w ewolucji powstają dzięki działaniu doboru!
62 Dobor czy dryf? Selekcjonizm większość utrwalonych mutacji została wyselekcjonowana przed dobór większość polimorfizmów jest utrzymywana przez dobór dobór równoważący, naddominacja, dobór zależny od częstości Neutralizm (Kimura, 1968) większość utrwalonych mutacji została utrwalona przez dryf za większość polimorfizmów odpowiada dryf mutacje utrwalane przez dobór są rzadkie, nie mają wpływu na ilościową analizę zmienności molekularnej
63 Mutacje i dobór niekorzystne (szkodliwe) s < 0 eliminowane przez dobór (oczyszczający/negatywny) neutralne s 0 (a konkretniej, s 1/4N) utrwalane przez dryf korzystne s > 0 utrwalane przez dobór (z udziałem dryfu dla niewielkich s)
64 Selekcjonizm i neutralizm Selekcjonizm: większość mutacji jest niekorzystna lub korzystna większość utrwalonych mutacji jest korzystna mutacje neutralne są rzadkie (nie częstsze od korzystnych) dobór jest głównym mechanizmem kształtującym zmienność sekwencji Neutralizm większość mutacji jest niekorzystna lub neutralna większość utrwalonych mutacji jest neutralna dryf jest głównym mechanizmem kształtującym zmienność sekwencji mutacje korzystne są rzadkie (znacznie rzadsze od neutralnych)

65 Selekcjonizm i neutralizm selekcjonizm neutralizm pan-neutralizm Neutralizm nie oznacza pan-neutralizmu, czyli negowania znaczenia selekcyjnego mutacji!
66 Przesłanki teorii neutralnej Tempo zmian sekwencji i polimorfizm są zbyt duże, by dały się wyjaśnić doborem Stałe tempo ewolucji molekularnej (zegar molekularny) Sekwencje o mniejszym znaczeniu funkcjonalnym (pseudogeny, mniej istotne obszary białek) ewoluują szybciej, niż obszary kluczowe dla funkcji

67 Stałe tempo ewolucji molekularnej Wiele sekwencji ewoluuje w stałym tempie Tempo to jest różne dla różnych sekwencji, ale stałe w czasie ewolucji dla danej sekwencji Tzw. zegar molekularny Różnice sekwencji globin kręgowców
68 Tempo ewolucji i dryf Neutralny dryf jest procesem losowym, ale jego tempo będzie stałe w odpowiednio długim czasie Zależy tylko od częstości mutacji (jedna zmiana na 1/µ pokoleń) Dla doboru stałe tempo zmian oznacza stałe tempo zmian środowiska Tempo zmian adaptacyjnych nie wydaje się być stałe
69 Zegar molekularny - problem W modelu neutralnym tempo utrwalania mutacji: 2Nµ 1 2N = µ Powinno być stałe w przeliczeniu na pokolenie Czas generacji jest różny u różnych organizmów Czyli nie powinna być obserwowana stałość tempa w czasie rzeczywistym A często jest (w tych sekwencjach, które zachowują zegar)
70 Problem czasu generacji Czas generacji różnych organizmów jest istotnie różny Dlaczego nie wpływa to na tempo utrwalania mutacji? ~0,03 pokolenia/rok ~3 pokolenia/rok
71 Zmiany prawie neutralne Model Kimury dotyczy zmian neutralnych (s = 0), takie nie są (w sekwencji białek) częste Mutacje zachowują się jak neutralne gdy spełnione jest: s 1 4N e Mutacje o niewielkim współczynniku doboru s będą zachowywały się jak neutralne w małych populacjach, a w większych populacjach będą podlegały doborowi
72 Zmiany prawie neutralne Istnieje odwrotna korelacja między czasem generacji a wielkością populacji
73 Zmiany prawie neutralne ~0,03 pokolenia/rok s 1 4N e ~3 pokolenia/rok Długi czas generacji Krótki czas generacji Mniej mutacji na rok Więcej mutacji na rok Populacja nieliczna (małe Ne) Populacja liczna (duże Ne) Więcej mutacji zachowuje się jak neutralne i utrwala przez dryf Więcej mutacji podlega doborowi (i jest eliminowane przez dobór oczyszczający) Efekty czasu generacji i wielkości populacji się znosząc, dając stałe tempo w czasie (Ohta & Kimura, 1971).
74 Zegar molekularny Dla sekwencji białek i zmian niesynonimicznych w DNA zmiany jednostajne w czasie Na poziomie DNA, dla mutacji synonimicznych pseudogenów niektórych sekwencji niekodujących tempo ewolucji zależy od czasu generacji
 ewoluują szybciej, niż obszary kluczowe dla funkcji Konserwacja sekwencji świadczy o jej")
75 Tempo ewolucji sekwencji a funkcja Głównym czynnikiem determinującym ilościową zmienność sekwencji jest dobór negatywny (oczyszczający) Sekwencje o mniejszym znaczeniu funkcjonalnym (pseudogeny, mniej istotne obszary białek) ewoluują szybciej, niż obszary kluczowe dla funkcji Konserwacja sekwencji świadczy o jej funkcji!
76 Tempo zmian Białka zaangażowane w podstawowe funkcje komórki ewoluują wolniej. W sekwencji białka obszary kluczowe dla funkcji ewoluują wolniej. Jednostka: PAM/10 8 lat Jednostka czasu ewolucyjnego: ile lat (w milionach, 10 6 ) potrzeba do utrwalenia 1 mutacji/100 aa (1 PAM)


77 Abraham Wald Pionier badań operacyjnych (teoria decyzji) Prace dla Center for Naval Analyses podczas II w. ś. Analiza rozmieszczenia przestrzelin w uszkodzonych samolotach oryginalny plan: wzmocnić pancerz w miejscach, gdzie obserwuje się najwięcej przestrzelin analiza Walda: wzmocnić tam, gdzie nie obserwuje się przestrzelin (samoloty tam trafione nie wróciły)

78 Zachowawczość sekwencji Obszary sekwencji białkowych najwolniej się zmieniające są zwykle kluczowe dla funkcji
79 Tempo zmian Czynnikiem decydującym o tempie zmian jest dobór oczyszczający (negatywny) w ważniejszych sekwencjach więcej zmian będzie niekorzystnych (eliminacja przez dobór) w mniej istotnych sekwencjach więcej zmian będzie neutralnych (utrwalanie przez dryf) zmiany bez znaczenia dla funkcji będą neutralne pseudogeny niekodujące obszary międzygenowe? podstawienia synonimiczne?

80 Spór wokół ENCODE ENCODE - projekt opisujący sekwencje w genomie (Encyclopedia of DNA Elements) Wiele sekwencji międzygenowych, niekodujących ulega transkrypcji 80% genomu funkcjonalne czy istnieje śmieciowy DNA? Czy to znaczy, że są funkcjonalne? Jeżeli nie ma śladów działania doboru - nie ma funkcji! Ślady działania doboru: 2-15% całego genomu
81 Status neutralizmu Wyjaśnia wiele zjawisk obserwowanych w ewolucji molekularnej wysoki polimorfizm sekwencji DNA i białek zegar molekularny ale jest wiele odstępstw, nie istnieje globalny zegar prawdziwy dla wszystkich gałęzi drzewa życia wolniejsza ewolucja sekwencji o kluczowym znaczeniu to też można wyjaśnić modelem, w którym większość mutacji jest albo niekorzystna, albo korzystna, ale niekorzystnych jest więcej Jest bardzo przydatny jako hipoteza zerowa do badania doboru naturalnego na poziomie sekwencji!
82 Status neutralizmu Dryf genetyczny ma w ewolucji molekularnej bardzo znaczącą, ale nie wyłączną rolę znaczne obszary genomu ewoluują w sposób bliski neutralnemu
83 Badanie doboru Założenie: mutacje synonimiczne są neutralne, sekwencje porównywane są parami Ka (dn) liczba mutacji niesynonimicznych na liczbę możliwych miejsc niesynonimicznych Ks (ds) liczba mutacji synonimicznych na liczbę możliwych miejsc synonimicznych Stosunek Ka/Ks (ω) jest miarą działania doboru
84 Czy zmiany synonimiczne są neutralne Kodony synonimiczne nie są równocenne Zmiana kodonu częstego na rzadki może wpłynąć na poziom ekspresji i kinetykę translacji

85 Czy zmiany synonimiczne są neutralne?
86 Epistaza W genetyce populacji termin oznacza dowolne oddziaływania genetyczne (w genetyce klasycznej tylko jeden rodzaj oddziaływania) Epistaza pozytywna - dostosowanie podwójnego mutanta większe, niż oczekiwane negatywna - dostosowanie podwójnego mutanta mniejsze, niż oczekiwane epistaza znaku - np. dwie mutacje szkodliwe (pojedynczo) razem są korzystne model wartości oczekiwanej: addytywny lub (częściej) multiplikatywny
 - trajektoria ewolucyjna zależy od kolejności zdarzeń T.")
87 Epistaza i ewolucja Przy braku epistazy nie ma znaczenia kolejność nabywania mutacji Przy epistazie efekt danej mutacji zależy od tła genetycznego (innych mutacji) - trajektoria ewolucyjna zależy od kolejności zdarzeń T. Shafee, wikimedia commons
88 Epistaza i płeć W populacjach rozmnażających się płciowo epistaza negatywna pomaga usuwać mutacje szkodliwe dzięki ich synergii: hipoteza deterministyczna mutacyjna (A. Kondrashov, 1988) inna hipoteza - płeć ułatwia powstanie korzystnych kombinacji niezależnych mutacji
89 Teoria informacji Zmienna losowa X obiekt mogący przyjąć skończoną liczbę stanów x1,,xn, z określonymi prawdopodobieństwami p1,,pn Przykład x1; p1=0,5 x2; p2=0,5
90 Informacja ujęcie matematyczne Entropia miara niepewności dotyczącej stanu zmiennej losowej N H(X) = p log p i i i=1 Podstawa logarytmu definiuje jednostkę log2 bity ln naty logn mery
91 Informacja ujęcie matematyczne W przypadku monety N H(X) = p log p i i i=1 x1; p1 = 0,5 x2; p2 = 0,5 H(X) = - [0,5*log2(0,5)+0,5*log2(0,5)] = -[0,5*(-1)+0,5*(-1)] = 1 bit Informacja zmniejszenie niepewności Znajomość wyniku rzutu monetą to 1 bit informacji Maksymalna informacja, którą może nieść układ jest równa jego entropii
92 Teoria informacji Informacja musi mieć reprezentację fizyczną (moneta, bity w pamięci komputerowej, nukleotydy w DNA, neurony) Informacja musi mieć kontekst Dowolny układ symboli jest potencjalną informacją (posiada entropię), ale dopiero związek ze światem fizycznym nadaje mu status informacji


93 Maksimum entropii Entropia jest największa wtedy, gdy p1=p2= pn największa niepewność
94 Teoria informacji w biologii W układach biologicznych informacja zapisana jest w DNA Jedna pozycja nukleotydowa, gdzie każdy z nukleotydów może występować z jednakowym prawdopodobieństwem: H(X) = 2 bity (1 mer) Jest to maksymalna entropia dla tej pozycji W rzeczywistości prawdopodobieństwo wystąpienia danego nukleotydu w danej pozycji nie zawsze wynosi 0,25, jeżeli na sekwencję działa dobór
95 O czym ta informacja? Żeby określić właściwą zawartość informacji i odróżnić ją od redundancji, trzeba wiedzieć czego dotyczy Każda populacja żyje w określonym środowisku Mutacje wywołują zmienność Środowisko dokonuje selekcji Informacja płynie ze środowiska do genomu (genomów)
96 Ewolucja a informacja Informacja w biologii jest więc ściśle powiązana z ewolucją. Tylko analiza ewolucyjna pozwala na określenie informacji w biologii.
97 Informacja a ewolucja W rzeczywistości prawdopodobieństwo wystąpienia danego nukleotydu w danej pozycji nie zawsze wynosi 0,25, jeżeli na sekwencję działa dobór W danym środowisku prawdopodobieństwa wystąpienia w sekwencji określonego nukleotydu są różne (np. C w 78% w danej pozycji, itp.) Na tej podstawie obliczamy H(X) Różnica między Hmax a H(X) to miara informacji o środowisku zawartej w tej pozycji
98 Informacja w sekwencji białka Maksymalna entropia pozycji jeżeli nie ma żadnych ograniczeń: H(Xi) = Hmax = log2(20) 4,32 bity W rzeczywistości aminokwasy w danej pozycji nie występują losowo ogranicza to dobór naturalny, czyli środowisko Sekwencje DNA i białek zawierają informację o środowisku (szeroko pojmowanym)
99 Informacja w sekwencji białka Stan środowiska to zmienna losowa E o stanach ej o określonym prawdopodobieństwie. Środowisko ogranicza występowanie w danej pozycji konkretnych aminokwasów (np. w danej grupie w 80% w danej pozycji Trp, itp.). Na tej podstawie można obliczyć obserwowaną entropię danej pozycji, kształtowaną przez środowisko (poprzez dobór): H(Xi E = ej) Różnica Hmax - H(Xi E = ej) to informacja o środowisku zapisana w pozycji i sekwencji.
100 Profil entropii białka Przykład: 57 aminokwasów homeodomeny gryzoni, na podstawie porównania 810 sekwencji Adami, C. (2012). The use of information theory in evolutionary biology. Ann N Y Acad Sci 1256,
101 Całkowita zawartość informacji Całkowitą entropię (i całkowitą zawartość informacji) uzyskamy sumując entropię dla wszystkich pozycji 57 i=1 I gryzonie = 57 H(X i ) I gryzonie = 25, 29 ± 0, 09 merów 109 bitów
102 Entropia homeodomeny w ewolucji Adami, C. (2012). The use of information theory in evolutionary biology. Ann N Y Acad Sci 1256,

103 Gdyby nie było doboru Schneider TD, 2000, Evolution of Biological Information, Nucleic Acids Res. 28:
104 Skąd pochodzi informacja genetyczna Informację genetyczną zapisuje środowisko poprzez proces doboru naturalnego, równoważąc utratę informacji na skutek błędów replikacji
105 Innowacje ewolucyjne w genomie

106 Skąd biorą się nowe funkcje (geny) Mutacje mogą zmienić funkcję genu, ale zwykle z utratą funkcji dotychczasowej Prawdopodobieństwo powstania nowego genu de novo (np. z sekwencji niekodującej) jest małe Rozwiązanie - duplikacje Susumu Ohno ( )
107 Duplikacje T.A. Brown. Genomy III, PWN 2009
108 Liczba genów wzrastała w historii życia
109 Ewolucja globin
110 Paralogi i ortologi Paralogi geny homologiczne w tym samym genomie, powstałe w wyniku duplikacji genu - np. α-globina i β-globina człowieka Ortologi geny homologiczne powstałe w wyniku specjacji, pochodzące od genu u wspólnego przodka np. α-globina człowieka i α-globina myszy
111 Ewolucja genów opsyn


112 Ewolucja widzenia barw
113 Geny HOX regulatory rozwoju
114 Duplikacje całych genomów Zmianie może ulec liczba chromosomów Podwojeniu może ulec cały genom Hipoteza 2R (hipoteza Ohno) podwojenie genomu na początku ewolucji kręgowców 2 rundy podwojenia np. geny Hox

115 Białka składają się z domen T.A. Brown. Genomy III, PWN 2009 Tasowanie domen kombinatoryka w białkach.
116 Wspólne motywy w różnych genach Możemy stawiać hipotezy dotyczące funkcji nieznanych białek na podstawie motywów znajdowanych w sekwencji. Podstawa większości współczesnych badań biochemicznych!!
117 Ewolucyjna zmienność genomów U Eukaryota skład genomu zmienia się powoli większość genów człowieka (>95%) ma odpowiedniki w genomie myszy, żaby itp. za różnice odpowiadają subtelne zmiany regulacji i współdziałania genów U Prokaryota (bakterie, archeony) duże róznice w zestawie genów nawet u blisko spokrewnionych organizmów



118 Ewolucyjne klocki Złożone sieci współzależności złożoność budowana przez oddziaływania i kombinacje, a nie liczbę elementów składowych Nowe elementy przez duplikację istniejących
119 Początki ewolucji Od abiogenezy do komórek eukariotycznych
120 Czym jest życie? metabolizm + informacja (replikacja)
121 (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja Powstanie życia z materii nieożywionej nie było zjawiskiem ewolucyjnym trudności z wyjaśnieniem abiogenezy nie mogą być traktowane jako zarzut wobec teorii ewolucji właściwe dziedziny: fizyka (teoria złożoności, teoria samoorganizacji, termodynamika) chemia planetologia
122 Czas i scena Istnieją też koncepcje umieszczające część z tych etapów poza Ziemią
123 Kluczowe etapy Etap chemiczny - prebiotyczny: początek >4,2 mld. lat temu Etap progenota - powstanie replikacji i podstawowych procesów biologicznych, początek ewolucji biologicznej: 4,2-3,8 mld. lat temu Etap komórkowy: od ~3,8 mld lat temu (najstarsze ślady kopalne) LUCA: ~3,6 mld. lat temu Eukarionty: ~2 mld. lat temu
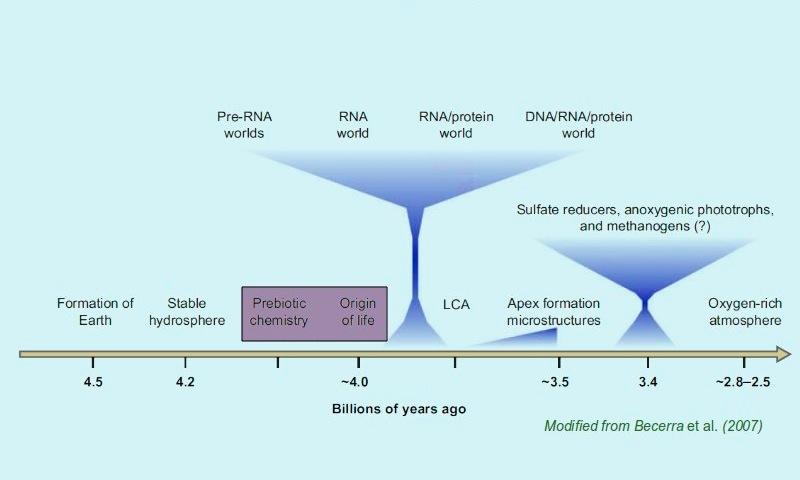
124 Prehistoria życia



125 Etap chemiczny Nie ma charakteru ewolucji biologicznej! Wiele eksperymentów odtwarzających pierwotną atmosferę i zachodzące w niej reakcje atmosfera beztlenowa, bogata w azot, dwutlenek węgla, związki siarki koncepcja bulionu koncepcja hydrotermalna Związki organiczne znajdowane w materiale pozaziemskim (meteoryty, komety) pokazują możliwość abiotycznego powstawania niezbędnych związków Ligacja polinukleotydów na katalizatorze glinokrzeminanowym
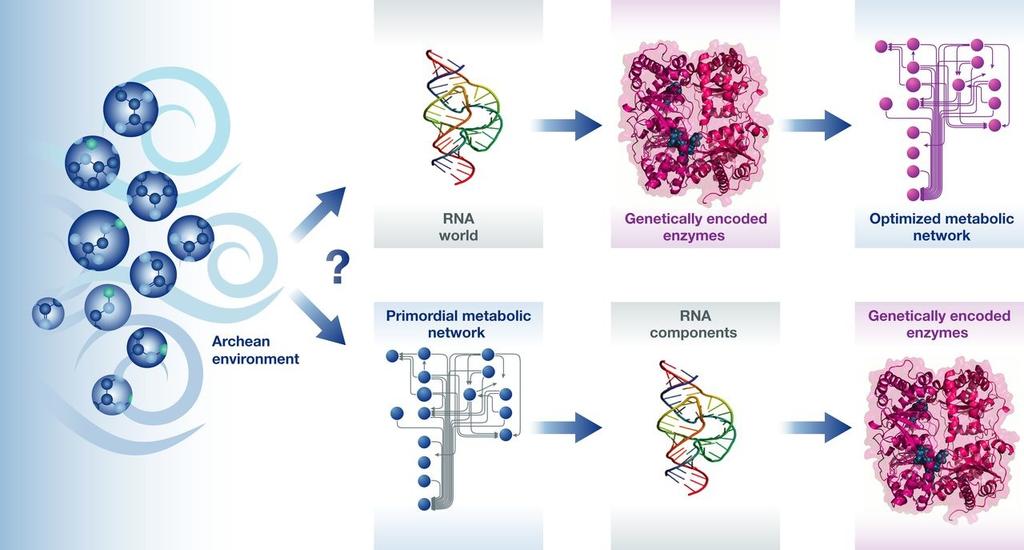
126 Prebiotyczna biochemia?
127 Prebiotyczna biochemia? Cząsteczki niezbędne do syntezy RNA mogły powstać w pierwotnych sieciach metabolicznych
128 Etap progenota Powstanie informacji kluczowe powstanie zdolności (samo)replikacji powiązanie genotypu z fenotypem możliwość działania doboru progenota Powstanie metabolizmu kluczowe powstanie samoorganizującej się sieci metabolicznej powielanie struktury nie na zasadzie replikacji matrycowej replikacja wynaleziona później
 Zależny od informacji genetycznej (kodowane enzymy)")
129 Co było najpierw? Metabolizm (Oparin, Dyson) Zależny od informacji genetycznej (kodowane enzymy) Replikacja (Eigen) Zależna od metabolizmu (enzymy replikujące DNA)
 jak i cząsteczki wyrażającej tę informację (enzymu) Można wyobrazić sobie życie oparte na RNA jako jedynej")
130 Świat RNA Odkrycie enzymatycznej aktywności RNA (rybozymy), 1982 RNA przyjmuje różne struktury przestrzenne, jest bardziej od DNA reaktywny chemicznie Może zatem pełnić rolę zarówno nośnika informacji genetycznej (podlegać replikacji) jak i cząsteczki wyrażającej tę informację (enzymu) Można wyobrazić sobie życie oparte na RNA jako jedynej makrocząsteczce
131 Rybozymy w naturze Cięcie i obróbka RNA introny grupy I i II (autokatalityczne) bakteryjna RNaza P spliceosom rybozymy wirusów roślinnych Synteza białek aktywność peptydylotransferazy rybosomu Wiele innych aktywności uzyskanych w laboratorium

132 Rybozym zdolny do syntezy RNA Wyselekcjonowany w laboratorium Wciąż nie autonomiczna replikaza
133 Czy zbadaliśmy wszystkie możliwe rybozymy? Długość cząsteczki (nt) Liczba możliwych kombinacji (4 N ) 50 1, , Liczba atomów na Ziemi: Liczba atomów we Wszechświecie:
134 Autokatalityczna ligaza RNA 2002 by National Academy of Sciences Paul N, Joyce G F PNAS 2002;99:
135 Paul N, Joyce G F PNAS 2002;99: Autokatalityczna ligaza RNA 2002 by National Academy of Sciences
136 Problemy świata RNA Ograniczona zdolność magazynowania informacji w pojedynczym replikatorze (ilość informacji możliwej do zakodowania jest odwrotnie proporcjonalna do częstości błędów replikacji granica Eigena) Rozwiązanie sieci współdziałających replikatorów (hipercykle) Samolubne RNA w sieci replikatorów rozwiązanie wydzielenie hipercyklu błoną i specjalizacja

137 Kooperacja czy samolubność
138 Kooperacja i samoorganizacja Nature, 2012 Nov 1;491(7422):72-7 Samoorganizujące się sieci rybozymów (2012) - potwierdzone doświadczalnie
139 Samoorganizacja sieci RNA Rybozym pochodzący z intronu Azoarcus (Proteobacteria) Pofragmentowany ma zdolność do autokatalitycznej ligacji fragmentów Stworzono warianty o różnej sekwencji i podobnych właściwościach Na ich podstawie stworzono warianty zdolne do ligowania innych wariantów
140 Rybozym Azoarcus
141 Samoorganizacja sieci RNA Rybozymy zdolne do katalizy ligacji innych wariantów tworzą cykle autokatalizy Cykl jest wydajniejszy od pojedynczych samolubnych rybozymów N Vaidya et al. Nature Nature, 2012 Nov 1;491(7422):72-7 doi: /nature11549
142 Kooperacja jest skuteczniejsza N Vaidya et al. Nature Nature, 2012 Nov 1;491(7422):72-7 doi: /nature11549
143 Zwiększenie liczby fragmentów Tworzą się sieci, których większość wymaga kooperacji. Złożoność w ewolucji in vitro przyrasta. N Vaidya et al. Nature Nature, 2012 Nov 1;491(7422):72-7 doi: /nature11549

 za mało kopii - replikacja się nie")
144 Ewolucja kooperacji Ewolucja kooperacji potwierdzona eksperymentalnie 2 RNA kodujące enzymy: replikaza i kinaza CDP Na skalę populacyjną wymaga enkapsulacji w pęcherzykach błonowych Zależy od liczby kopii RNA za dużo kopii - wygrywają pasożytnicze RNA (B) za mało kopii - replikacja się nie utrzymuje

145 Powstanie błon pierwsze prakomórki Samoorganizacja lipidów amfipatycznych w struktury mogące otaczać prakomórki Takie lipidy mogły powstawać w warunkach prebiotycznych, a nawet w kosmosie Wyodrębnienie prakomórek błoną nastąpiło wcześnie w ewolucji Deamer et al. (2002), Astrobiology 2(4)
146 Od świata RNA do świata RNP Oddziaływania RNA aminokwasy (pratrna) rybozymy syntetyzujące uniwersalny rybosom pojawił się później

147 Aminoacylacja RNA przez prosty rybozym
148 Powstanie DNA Reduktaza rybonukleotydów Enzym obecny we wszystkich gałęziach drzewa życia bardzo stary
149 Podsumowanie
150 Hipotezy alternatywne Przesunięcie niektórych etapów prehistorii życia poza Ziemię kosmiczne pochodzenie prostych cząsteczek organicznych kosmiczne pochodzenie życia - panspermia

151 Panspermia Pierwsze cząsteczki biologiczne, a nawet organizmy nie powstały na Ziemi Cząsteczki organiczne, aminokwasy w materiale kosmicznym Problem ustalenia warunków początkowych Jeżeli nie wiemy, gdzie powstawało życie, nie mamy możliwości formułowania hipotez Panspermia ukierunowana życie celowo zasiane na Ziemi przez inną cywilizację (Orgel & Crick, 1973)
152 LUCA
153 Jaki był LUCA? Na podstawie zestawu genów obecnych we wszystkich gałęziach drzewa życia problem horyzontalny transfer genów (przeszacowanie zestawu genów pierwotnych) Organizm o budowie komórkowej zbliżony do współczesnych prokariontów Poprzedzał rozejście się linii Bacteria, Archaea i Eukarya

154 Rekonstrukcja genomu LUCA Geny obecne we wszystkich liniach Bacteria i Archaea - ok. 30 Geny obecne w obu domenach: ~11 000, ale niewykluczony transfer horyzontalny Geny obecne w co najmniej 2 liniach każdej z domen i monofiletyczne w obu domenach: 355
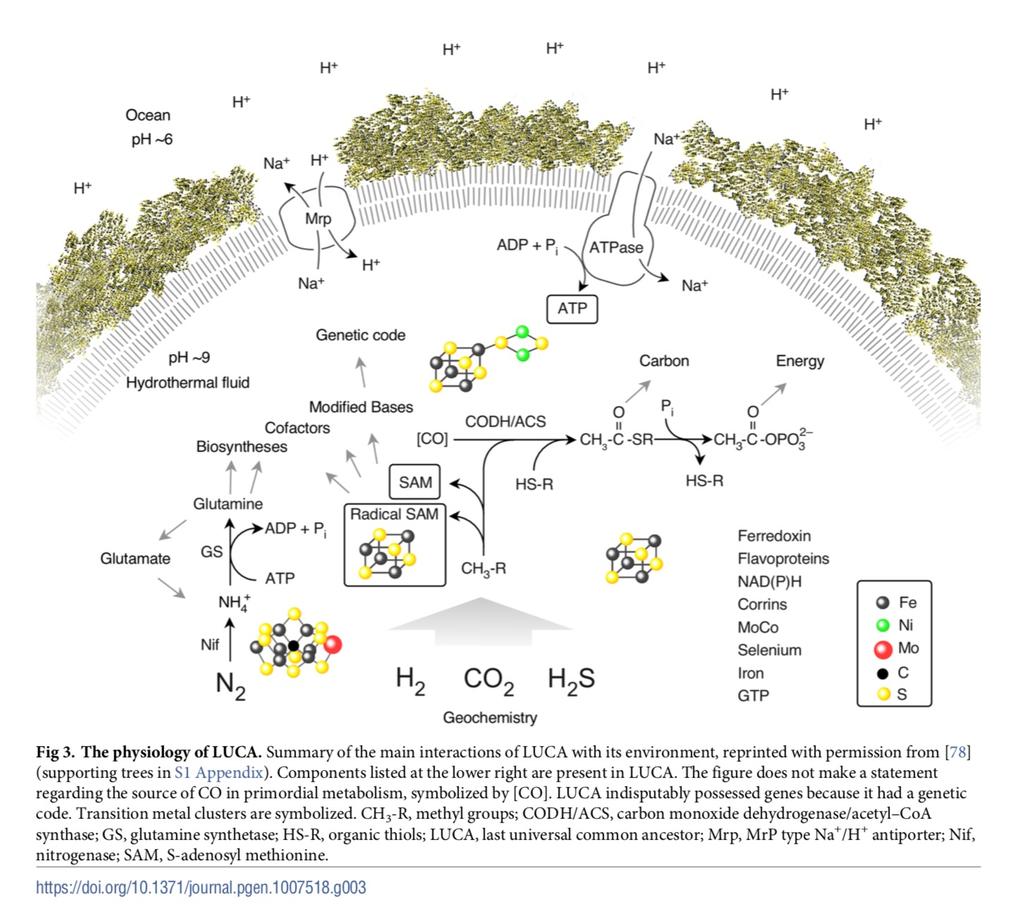
155 Fizjologia LUCA
156 Co potrafił LUCA transkrypcja, translacja (kod odpowiadający współczesnemu) chemoautotrof, metabolizm energetyczny oparty na ATP synteza długich łańcuchów DNA białka błonowe ok. 600 genów Ale nie wszyscy się zgadzają: prosty, niewiele genów, bez współczesnego systemu replikacji genomu, genom RNA (Koonin 2003)

157 Od LUCA do trzech domen Komórki eukariotyczne prezentują wyższy poziom złożoności Komórki eukariotyczne są znacznie większe: µm vs µm, co daje ~1000 x różnicę objętości

158 Po co nam mitochondria Systemy wytwarzające energię znajdują się na powierzchni błony Przy zwiększaniu rozmiarów powierzchnia wzrasta wolniej niż objętość Wydajna produkcja energii w większych komórkach wymaga dodatkowej powierzchni błon Mitochondria dały zasoby energii dla zwiększenia złożoności u Eukaryota


")
159 Endosymbioza Mitochondria i chloroplasty mają pochodzenie endosymbiotyczne od Bacteria mitochondria: monofiletyczne, spokrewnione z α-proteobacteria chloroplasty - polifiletyczne, symbiozy pierwotne i wtórne, spokrewnione z Cyanobacteria Konstanty Miereżkowski ( ) Lynn Margulis ( )


160 Prokaryota nie są jedną grupą Carl Woese ( )
161 Drzewo życia - wersja (bardzo) uproszczona (Doolittle, 1999)
162 Hipoteza Archezoa Lata 90. Gospodarzem endosymbiozy była komórka typu eukariotycznego (z jądrem), pozbawiona mitochondriów Współczesne eukarionty pozbawione mitochondriów - Archezoa- są jej potomkami Embley and Martin. Nature 2006


 jeżeli oddychają")

163 Archezoa Wszystkie współczesne Eukaryota, nawet pozbawione mitochondriów: posiadają geny pochodzące od symbionta mitochondrium hydrogenosom mitosom posiadają zredukowane organella pochodzące od mitochondriów (z jednym znanym wyjątkiem) jeżeli oddychają tlenowo, to mają DNA w mitochondriach Wspólna krytyczna funkcja pochodzenia endosymbiotycznego - synteza grup FeS

164 Współczesny obraz eukariogenezy Nie było etapu Archezoa - eukarionta pierwotnie pozbawionego mitochondrium Symbioza gospodarza - Archeona i symbionta - α-proteobakterii Struktury komórki eukariotycznej powstały później Problemem tej teorii długo była kwestia pochodzenia typowo eukariotycznych funkcji - cytoszkieletu, pęcherzyków błonowych, jądra itp. i identyfikacja gospodarza

 Najbliżej (wtedy) spokrewniona z Eukaryota Posiada geny")

165 Gospodarz endosymbiozy był archeonem Lokiarcheota - monofiletyczna grupa Archaea, odkryta w badaniach metagenomicznych (2015 r.) Najbliżej (wtedy) spokrewniona z Eukaryota Posiada geny kodujące białka umożliwiające tworzenie złożonych struktur błonowych Tak mógł wyglądać gospodarz endosymbiozy, która dała początek Eukaryota W 2017 opisano więcej typów Archaea z tej gałęzi
166 Archaea i początki eukariontów a - drzewo Woese (1977) z modyfikacjami: 3 domeny b - dwie domeny c, d - po odkryciu nowych grup Archaea dzięki metagenomice TACK - Thaumarchaeota, Aigarchaeota, Crenarchaeota, Korarchaeota DPANN - Diapherotrites, Parvarchaeota, Aenigmarchaeota, Nanoarchaeota, Nanohaloarchaeota

167 Powstanie eukariontów Geny symbionta nie są jedynymi genami Bacteria nabytymi przez gospodarza Białka mitochondrialne stanowią monofiletyczną grupę, co sugeruje, że zostały nabyte po wcześniejszych transferach FECA - First Eukaryotic Common Ancestor LECA - Last Eukaryotic Common Ancestor

168 Pochodzenie typowo eukariotycznych funkcji u FECA
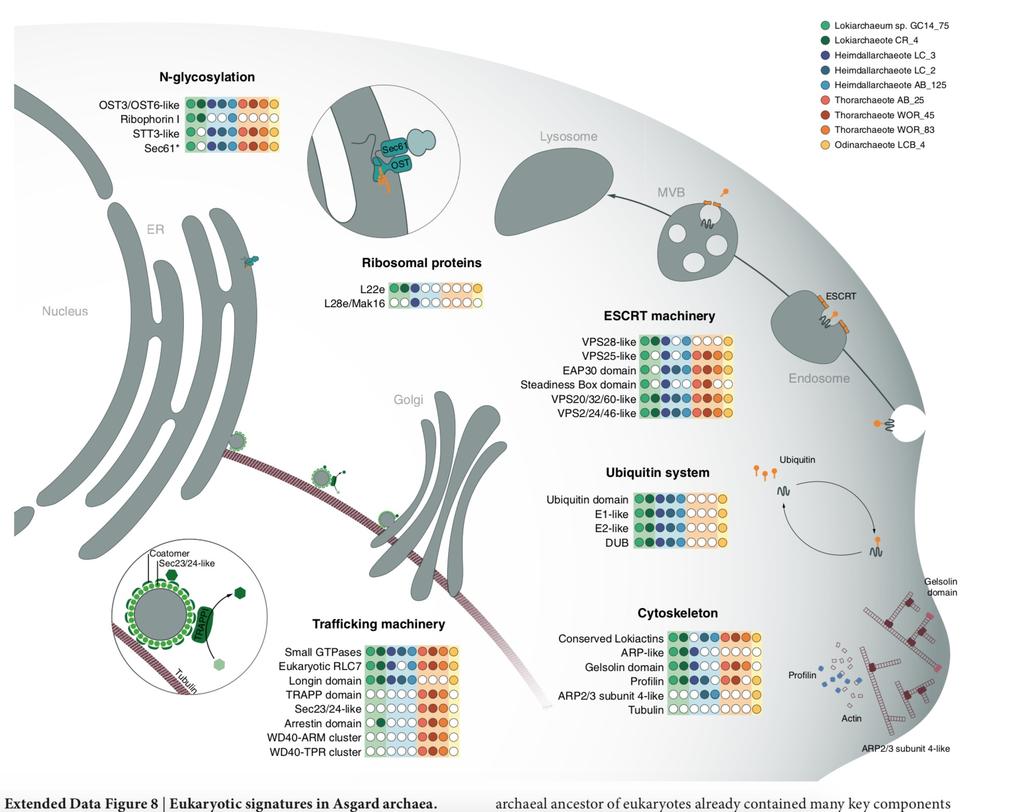
169

170 Gospodarz symbiozy FECA był najbliższym krewnym współczesnych Archaea z grupy Asgard Chyba, że znajdziemy jeszcze bliższą grupę
 translacja")
171 Pochodzenie jądra i eukariotycznego typu ekspresji genów Prokarionty - translacja sprzężona z transkrypcją w czasie i przestrzeni Eukarionty - transkrypcja i translacja rozdzielone jądro - transkrypcja i obróbka RNA eksport mrna z jądra po obróbce i kontroli jakości (RNA surveillance) translacja - cytoplazma
172 Introny Geny wszystkich eukariontów zawierają introny liczba i gęstość są różne Wszystkie eukarionty mają spliceosom (Collins L, Penny D. Complex spliceosomal organization ancestral to extant eukaryotes. Mol Biol Evol. 2005, 22: ) Introny eukariotyczne pochodzą od intronów grupy II Rogozin et al. Biol. Direct 2012
173 Introny prokariotyczne Powszechne, ale rzadkie grupa II - zasadniczo tylko Bacteria Przeważnie w genach kodujących funkcjonalne RNA, nie mrna Inny mechanizm - przeważnie autokatalityczne introny są rybozymami zdolnymi do samowycinania ruchome elementy genetyczne: transpozycja i retrotranspozycja (grupa II) Introny typu prokariotycznego obecne w genomach organellarnych wielu grup
174 Pochodzenie intronów - hipotezy Introny wcześnie (introns early) Introny (typu eukariotycznego) były powszechne, bakterie utraciły większość jako adaptację do szybkich podziałów Hipoteza tasowania eksonów (exon shuffling) - eksony odpowiadają podstawowym domenom białek, introny ułatwiają kombinatoryczną ewolucję nowych białek Introny późno (introns late) Introny powstały po powstaniu głównych gałęzi drzewa życia, rozprzestrzeniają się w HGT
175 Introny a jądro Hipoteza - jądro powstało jako system obrony przed inwazją intronów od symbionta mitochondrialnego Ekspansja intronów grupy II symbionta (gospodarz nie miał systemów kontroli) Składanie (splicing) jest wolniejsze od translacji Rozdzielenie translacji od składania - obrona przed zaburzeniem translacji przez sekwencje intronowe Introny istniały od początku, ale po powstaniu eukariontów gwałtownie się rozprzestrzeniły





176 Najpierw mitochondria, potem jądro? Czy w strukturze jądra są białka kodowane przez geny pochodzące od endosymbionta? Tak: Elementy kompleksu poru jądrowego:

177 Energetyka złożoności Budżet energetyczny komórki prokariotycznej: ~0,5 pw (1 pw = W) Budżet energetyczny komórki eukariotycznej: ~2300 pw Udział w budżecie energetycznym: replikacja: ~2% ekspresja genów i synteza białek: ~75% Znacznie więcej energii/gen u eukariontów!! Jedyna droga zwiększania złożoności
Badanie doboru naturalnego na poziomie molekularnym
 Badanie doboru naturalnego na poziomie molekularnym Podstawy ewolucji molekulanej Jak ewoluują sekwencje Zmiany genetyczne w ewolucji Mutacje tworzą nowe allele genów Inwersje zmieniają układ genów na
Badanie doboru naturalnego na poziomie molekularnym Podstawy ewolucji molekulanej Jak ewoluują sekwencje Zmiany genetyczne w ewolucji Mutacje tworzą nowe allele genów Inwersje zmieniają układ genów na
Podstawy ewolucji molekularnej. Ewolucja sekwencji DNA i białek
 Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Zmiany genetyczne w ewolucji Mutacje tworzą nowe allele genów Inwersje zmieniają układ genów na chromosomach mogą uniemożliwić rekombinację
Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Zmiany genetyczne w ewolucji Mutacje tworzą nowe allele genów Inwersje zmieniają układ genów na chromosomach mogą uniemożliwić rekombinację
Podstawy ewolucji molekularnej. Ewolucja sekwencji DNA i białek
 Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Egzamin: 29.01.2018 16:00, sala 9B Pierwsza synteza Ewolucja jako zmiany częstości alleli w populacji Mutacje jako źródło nowych alleli Dobór
Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Egzamin: 29.01.2018 16:00, sala 9B Pierwsza synteza Ewolucja jako zmiany częstości alleli w populacji Mutacje jako źródło nowych alleli Dobór
Podstawy ewolucji molekularnej. Ewolucja sekwencji DNA i białek
 Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Zmiany genetyczne w ewolucji } Mutacje } tworzą nowe allele genów } Inwersje } zmieniają układ genów na chromosomach } mogą uniemożliwić rekombinację
Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Zmiany genetyczne w ewolucji } Mutacje } tworzą nowe allele genów } Inwersje } zmieniają układ genów na chromosomach } mogą uniemożliwić rekombinację
Zmienność ewolucyjna. Ewolucja molekularna
 Zmienność ewolucyjna Ewolucja molekularna Mechanizmy ewolucji Generujące zmienność mutacje rearanżacje genomu horyzontalny transfer genów! Działające na warianty wytworzone przez zmienność dobór naturalny
Zmienność ewolucyjna Ewolucja molekularna Mechanizmy ewolucji Generujące zmienność mutacje rearanżacje genomu horyzontalny transfer genów! Działające na warianty wytworzone przez zmienność dobór naturalny
Podstawy genetyki populacji. Genetyka mendlowska i ewolucja. Dobór i dryf.
 Podstawy genetyki populacji Genetyka mendlowska i ewolucja. Dobór i dryf. Dryf genetyczny W populacjach o skończonej liczebności może dochodzić do zmian częstości alleli nawet jeżeli nie działa na nie
Podstawy genetyki populacji Genetyka mendlowska i ewolucja. Dobór i dryf. Dryf genetyczny W populacjach o skończonej liczebności może dochodzić do zmian częstości alleli nawet jeżeli nie działa na nie
Mechanizmy zmienności ewolucyjnej. Podstawy ewolucji molekularnej.
 Mechanizmy zmienności ewolucyjnej Podstawy ewolucji molekularnej. Mechanizmy ewolucji } Generujące zmienność } mutacje } rearanżacje genomu } horyzontalny transfer genów } Działające na warianty wytworzone
Mechanizmy zmienności ewolucyjnej Podstawy ewolucji molekularnej. Mechanizmy ewolucji } Generujące zmienność } mutacje } rearanżacje genomu } horyzontalny transfer genów } Działające na warianty wytworzone
Podstawy teorii ewolucji. Informacja i ewolucja
 Podstawy teorii ewolucji Informacja i ewolucja Podręczniki 2 Dla zainteresowanych http://wps.prenhall.com/esm_freeman_evol_4/ 3 Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego
Podstawy teorii ewolucji Informacja i ewolucja Podręczniki 2 Dla zainteresowanych http://wps.prenhall.com/esm_freeman_evol_4/ 3 Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego
Podstawy ewolucji molekularnej. Ewolucja sekwencji DNA i białek
 Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Podręczniki Populacja Grupa krzyżujących się ze sobą osobników oraz ich potomstwo Zbiór wszystkich alleli populacji pula genowa Najprostszy
Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Podręczniki Populacja Grupa krzyżujących się ze sobą osobników oraz ich potomstwo Zbiór wszystkich alleli populacji pula genowa Najprostszy
Podstawy teorii ewolucji. Informacja i ewolucja
 Podstawy teorii ewolucji Informacja i ewolucja Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Podstawy teorii ewolucji Informacja i ewolucja Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Podstawy genetyki populacji. Genetyka mendlowska i ewolucja. Dobór i dryf.
 Podstawy genetyki populacji Genetyka mendlowska i ewolucja. Dobór i dryf. Dryf genetyczny W populacjach o skończonej liczebności może dochodzić do zmian częstości alleli nawet jeżeli nie działa na nie
Podstawy genetyki populacji Genetyka mendlowska i ewolucja. Dobór i dryf. Dryf genetyczny W populacjach o skończonej liczebności może dochodzić do zmian częstości alleli nawet jeżeli nie działa na nie
Podstawy genetyki populacji. Genetyka mendlowska i ewolucja
 Podstawy genetyki populacji Genetyka mendlowska i ewolucja Informacja ujęcie matematyczne Entropia miara niepewności dotyczącej stanu zmiennej losowej N H(X) = p log p i i i=1 Podstawa logarytmu definiuje
Podstawy genetyki populacji Genetyka mendlowska i ewolucja Informacja ujęcie matematyczne Entropia miara niepewności dotyczącej stanu zmiennej losowej N H(X) = p log p i i i=1 Podstawa logarytmu definiuje
Historia informacji genetycznej. Jak ewolucja tworzy nową informację (z ma ą dygresją).
.") Historia informacji genetycznej. Jak ewolucja tworzy nową informację (z ma ą dygresją). Czym jest życie? metabolizm + informacja (replikacja) 2 Cząsteczki organiczne mog y powstać w atmosferze pierwotnej
Historia informacji genetycznej. Jak ewolucja tworzy nową informację (z ma ą dygresją). Czym jest życie? metabolizm + informacja (replikacja) 2 Cząsteczki organiczne mog y powstać w atmosferze pierwotnej
Elementy teorii informacji w ewolucji
 Elementy teorii informacji w ewolucji Teoria informacji Zmienna losowa X obiekt mogący przyjąć skończoną liczbę stanów x1,,xn, z określonymi prawdopodobieństwami p1,,pn Przykład x1; p1=0,5 x2; p2=0,5 Informacja
Elementy teorii informacji w ewolucji Teoria informacji Zmienna losowa X obiekt mogący przyjąć skończoną liczbę stanów x1,,xn, z określonymi prawdopodobieństwami p1,,pn Przykład x1; p1=0,5 x2; p2=0,5 Informacja
Powstanie i ewolucja informacji genetycznej
 Powstanie i ewolucja informacji genetycznej Literatura Brown, r. 18 Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja Powstanie
Powstanie i ewolucja informacji genetycznej Literatura Brown, r. 18 Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja Powstanie
Powstanie i ewolucja informacji genetycznej
 Powstanie i ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja Powstanie życia z materii nieożywionej
Powstanie i ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja Powstanie życia z materii nieożywionej
Podstawy teorii ewolucji. Informacja i ewolucja
 Podstawy teorii ewolucji Informacja i ewolucja Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Podstawy teorii ewolucji Informacja i ewolucja Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Powstanie i ewolucja informacji genetycznej
 Powstanie i ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych! Życie = ewolucja! Powstanie życia z materii
Powstanie i ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych! Życie = ewolucja! Powstanie życia z materii
Powstanie życia i dzieje informacji genetycznej. Oraz (bardzo) krótka historia życia.
 krótka historia życia.") Powstanie życia i dzieje informacji genetycznej Oraz (bardzo) krótka historia życia. Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie
Powstanie życia i dzieje informacji genetycznej Oraz (bardzo) krótka historia życia. Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie
Powstanie i ewolucja informacji genetycznej
 Powstanie i ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) 2 Cząsteczki organiczne mogły powstać w atmosferze pierwotnej Ziemi Oparin, Haldane Miller, 1953 3 (A)biogeneza
Powstanie i ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) 2 Cząsteczki organiczne mogły powstać w atmosferze pierwotnej Ziemi Oparin, Haldane Miller, 1953 3 (A)biogeneza
Powstanie życia i dzieje informacji genetycznej. Oraz (bardzo) krótka historia życia.
 krótka historia życia.") Powstanie życia i dzieje informacji genetycznej Oraz (bardzo) krótka historia życia. Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie
Powstanie życia i dzieje informacji genetycznej Oraz (bardzo) krótka historia życia. Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie
Zmienność ewolucyjna. Ewolucja molekularna
 Zmienność ewolucyjna Ewolucja molekularna Mechanizmy ewolucji Generujące zmienność mutacje rearanżacje genomu horyzontalny transfer genów Działające na warianty wytworzone przez zmienność dobór naturalny
Zmienność ewolucyjna Ewolucja molekularna Mechanizmy ewolucji Generujące zmienność mutacje rearanżacje genomu horyzontalny transfer genów Działające na warianty wytworzone przez zmienność dobór naturalny
Podstawy genetyki populacji. Genetyka mendlowska i ewolucja
 Podstawy genetyki populacji Genetyka mendlowska i ewolucja Syntetyczna teoria ewolucji } Pierwsza synteza: połączenie teorii ewolucji Darwina z genetyką mendlowską na poziomie populacji } W naturalnych
Podstawy genetyki populacji Genetyka mendlowska i ewolucja Syntetyczna teoria ewolucji } Pierwsza synteza: połączenie teorii ewolucji Darwina z genetyką mendlowską na poziomie populacji } W naturalnych
Teoria ewolucji. Podstawowe pojęcia. Wspólne pochodzenie.
 Teoria ewolucji Podstawowe pojęcia. Wspólne pochodzenie. Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Teoria ewolucji Podstawowe pojęcia. Wspólne pochodzenie. Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Zmienność ewolucyjna. Ewolucja molekularna
 Zmienność ewolucyjna Ewolucja molekularna Mechanizmy ewolucji Generujące zmienność mutacje rearanżacje genomu horyzontalny transfer genów Działające na warianty wytworzone przez zmienność dobór naturalny
Zmienność ewolucyjna Ewolucja molekularna Mechanizmy ewolucji Generujące zmienność mutacje rearanżacje genomu horyzontalny transfer genów Działające na warianty wytworzone przez zmienność dobór naturalny
Ewolucja informacji genetycznej
 1 Ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) Cząsteczki organiczne mog y powstać w atmosferze pierwotnej Ziemi Oparin, Haldane Miller, 1953 Co by o najpierw?
1 Ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) Cząsteczki organiczne mog y powstać w atmosferze pierwotnej Ziemi Oparin, Haldane Miller, 1953 Co by o najpierw?
Powstanie i najwcześniejsze dzieje życia. Od abiogenezy do LUCA
 Powstanie i najwcześniejsze dzieje życia Od abiogenezy do LUCA Czym jest życie? metabolizm + informacja (replikacja) 2 (A)biogeneza } Ewolucja jest właściwością organizmów żywych } Życie = ewolucja } Powstanie
Powstanie i najwcześniejsze dzieje życia Od abiogenezy do LUCA Czym jest życie? metabolizm + informacja (replikacja) 2 (A)biogeneza } Ewolucja jest właściwością organizmów żywych } Życie = ewolucja } Powstanie
Podstawy teorii ewolucji. Informacja i ewolucja
 Podstawy teorii ewolucji Informacja i ewolucja Podręczniki Dla zainteresowanych http://wps.prenhall.com/esm_freeman_evol_4/ Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego
Podstawy teorii ewolucji Informacja i ewolucja Podręczniki Dla zainteresowanych http://wps.prenhall.com/esm_freeman_evol_4/ Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego
Powstanie i dzieje informacji genetycznej. Oraz (bardzo) krótka historia życia.
 krótka historia życia.") Powstanie i dzieje informacji genetycznej. Oraz (bardzo) krótka historia życia. Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja
Powstanie i dzieje informacji genetycznej. Oraz (bardzo) krótka historia życia. Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja
Powstanie i dzieje informacji genetycznej. Oraz (bardzo) krótka historia życia.
 krótka historia życia.") Powstanie i dzieje informacji genetycznej Oraz (bardzo) krótka historia życia. Czym jest życie? metabolizm + informacja (replikacja)!2 (A)biogeneza } Ewolucja jest właściwością organizmów żywych!3! } Życie
Powstanie i dzieje informacji genetycznej Oraz (bardzo) krótka historia życia. Czym jest życie? metabolizm + informacja (replikacja)!2 (A)biogeneza } Ewolucja jest właściwością organizmów żywych!3! } Życie
Powstanie i najwcześniejsze dzieje życia. Od abiogenezy do LUCA
 Powstanie i najwcześniejsze dzieje życia Od abiogenezy do LUCA Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja Powstanie
Powstanie i najwcześniejsze dzieje życia Od abiogenezy do LUCA Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja Powstanie
Powstanie i najwcześniejsze dzieje życia. Od abiogenezy do LUCA
 Powstanie i najwcześniejsze dzieje życia Od abiogenezy do LUCA Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja Powstanie
Powstanie i najwcześniejsze dzieje życia Od abiogenezy do LUCA Czym jest życie? metabolizm + informacja (replikacja) (A)biogeneza Ewolucja jest właściwością organizmów żywych Życie = ewolucja Powstanie
ZARZĄDZANIE POPULACJAMI ZWIERZĄT
 ZARZĄDZANIE POPULACJAMI ZWIERZĄT Ćwiczenia 1 mgr Magda Kaczmarek-Okrój magda_kaczmarek_okroj@sggw.pl 1 ZAGADNIENIA struktura genetyczna populacji obliczanie frekwencji genotypów obliczanie frekwencji alleli
ZARZĄDZANIE POPULACJAMI ZWIERZĄT Ćwiczenia 1 mgr Magda Kaczmarek-Okrój magda_kaczmarek_okroj@sggw.pl 1 ZAGADNIENIA struktura genetyczna populacji obliczanie frekwencji genotypów obliczanie frekwencji alleli
Ewolucja informacji genetycznej
 1 Ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) Cząsteczki organiczne mogły powstać w atmosferze pierwotnej Ziemi Oparin, Haldane Miller, 1953 Co było najpierw?
1 Ewolucja informacji genetycznej Czym jest życie? metabolizm + informacja (replikacja) Cząsteczki organiczne mogły powstać w atmosferze pierwotnej Ziemi Oparin, Haldane Miller, 1953 Co było najpierw?
Teoria ewolucji. Losy gatunków: specjacja i wymieranie. Podstawy ewolucji molekularnej
 Teoria ewolucji. Losy gatunków: specjacja i wymieranie. Podstawy ewolucji molekularnej Specjacja } Pojawienie się bariery reprodukcyjnej między populacjami dające początek gatunkom } Specjacja allopatryczna
Teoria ewolucji. Losy gatunków: specjacja i wymieranie. Podstawy ewolucji molekularnej Specjacja } Pojawienie się bariery reprodukcyjnej między populacjami dające początek gatunkom } Specjacja allopatryczna
Teoria ewolucji. Podstawowe pojęcia. Wspólne pochodzenie.
 Teoria ewolucji Podstawowe pojęcia. Wspólne pochodzenie. Ewolucja Znaczenie ogólne: zmiany zachodzące stopniowo w czasie W biologii ewolucja biologiczna W astronomii i kosmologii ewolucja gwiazd i wszechświata
Teoria ewolucji Podstawowe pojęcia. Wspólne pochodzenie. Ewolucja Znaczenie ogólne: zmiany zachodzące stopniowo w czasie W biologii ewolucja biologiczna W astronomii i kosmologii ewolucja gwiazd i wszechświata
Konspekt do zajęć z przedmiotu Genetyka dla kierunku Położnictwo dr Anna Skorczyk-Werner Katedra i Zakład Genetyki Medycznej
 Seminarium 1 część 1 Konspekt do zajęć z przedmiotu Genetyka dla kierunku Położnictwo dr Anna Skorczyk-Werner Katedra i Zakład Genetyki Medycznej Genom człowieka Genomem nazywamy całkowitą ilość DNA jaka
Seminarium 1 część 1 Konspekt do zajęć z przedmiotu Genetyka dla kierunku Położnictwo dr Anna Skorczyk-Werner Katedra i Zakład Genetyki Medycznej Genom człowieka Genomem nazywamy całkowitą ilość DNA jaka
GENETYKA POPULACJI. Ćwiczenia 1 Biologia I MGR /
 GENETYKA POPULACJI Ćwiczenia 1 Biologia I MGR 1 ZAGADNIENIA struktura genetyczna populacji obliczanie frekwencji genotypów obliczanie frekwencji alleli przewidywanie struktury następnego pokolenia przy
GENETYKA POPULACJI Ćwiczenia 1 Biologia I MGR 1 ZAGADNIENIA struktura genetyczna populacji obliczanie frekwencji genotypów obliczanie frekwencji alleli przewidywanie struktury następnego pokolenia przy
Teoria ewolucji. Podstawy wspólne pochodzenie.
 Teoria ewolucji. Podstawy wspólne pochodzenie. Ewolucja biologiczna } Znaczenie ogólne: } proces zmian informacji genetycznej (częstości i rodzaju alleli), } które to zmiany są przekazywane z pokolenia
Teoria ewolucji. Podstawy wspólne pochodzenie. Ewolucja biologiczna } Znaczenie ogólne: } proces zmian informacji genetycznej (częstości i rodzaju alleli), } które to zmiany są przekazywane z pokolenia
plezjomorfie: podobieństwa dziedziczone po dalszych przodkach (c. atawistyczna)
") Podobieństwa pomiędzy organizmami - cechy homologiczne: podobieństwa wynikające z dziedziczenia - apomorfie: podobieństwa dziedziczone po najbliższym przodku lub pojawiająca się de novo (c. ewolucyjnie
Podobieństwa pomiędzy organizmami - cechy homologiczne: podobieństwa wynikające z dziedziczenia - apomorfie: podobieństwa dziedziczone po najbliższym przodku lub pojawiająca się de novo (c. ewolucyjnie
Ewolucjonizm NEODARWINIZM. Dr Jacek Francikowski Uniwersyteckie Towarzystwo Naukowe Uniwersytet Śląski w Katowicach
 Ewolucjonizm NEODARWINIZM Dr Jacek Francikowski Uniwersyteckie Towarzystwo Naukowe Uniwersytet Śląski w Katowicach Główne paradygmaty biologii Wspólne początki życia Komórka jako podstawowo jednostka funkcjonalna
Ewolucjonizm NEODARWINIZM Dr Jacek Francikowski Uniwersyteckie Towarzystwo Naukowe Uniwersytet Śląski w Katowicach Główne paradygmaty biologii Wspólne początki życia Komórka jako podstawowo jednostka funkcjonalna
Bliskie Spotkanie z Biologią. Genetyka populacji
 Bliskie Spotkanie z Biologią Genetyka populacji Plan wykładu 1) Częstości alleli i genotypów w populacji 2) Prawo Hardy ego-weinberga 3) Dryf genetyczny 4) Efekt założyciela i efekt wąskiego gardła 5)
Bliskie Spotkanie z Biologią Genetyka populacji Plan wykładu 1) Częstości alleli i genotypów w populacji 2) Prawo Hardy ego-weinberga 3) Dryf genetyczny 4) Efekt założyciela i efekt wąskiego gardła 5)
Modelowanie ewolucji. Dobór i dryf genetyczny
 Modelowanie ewolucji Dobór i dryf genetyczny Syntetyczna teoria ewolucji Pierwsza synteza: połączenie teorii ewolucji Darwina z genetyką mendlowską na poziomie populacji W naturalnych populacjach występują
Modelowanie ewolucji Dobór i dryf genetyczny Syntetyczna teoria ewolucji Pierwsza synteza: połączenie teorii ewolucji Darwina z genetyką mendlowską na poziomie populacji W naturalnych populacjach występują
Podstawy genetyki populacji. Populacje o skończonej liczebności. Dryf. Modele wielogenowe.
 Podstawy genetyki populacji Populacje o skończonej liczebności. Dryf. Modele wielogenowe. Dryf genetyczny a ewolucja } Dobór naturalny nie jest jedynym mechanizmem kształtującym zmiany ewolucyjne } Losowe
Podstawy genetyki populacji Populacje o skończonej liczebności. Dryf. Modele wielogenowe. Dryf genetyczny a ewolucja } Dobór naturalny nie jest jedynym mechanizmem kształtującym zmiany ewolucyjne } Losowe
Zmienność ewolucyjna. Ewolucja molekularna
 Zmienność ewolucyjna Ewolucja molekularna Podobieństwo i homologia Homologia: podobieństwo wynikające ze wspólnego pochodzenia ewolucyjnego cecha odziedziczona od wspólnego przodka vs. homoplazja podobieństwo
Zmienność ewolucyjna Ewolucja molekularna Podobieństwo i homologia Homologia: podobieństwo wynikające ze wspólnego pochodzenia ewolucyjnego cecha odziedziczona od wspólnego przodka vs. homoplazja podobieństwo
Podstawy biologii. Informacja genetyczna. Co to jest ewolucja.
 Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Materiał genetyczny Materiałem genetycznym są kwasy nukleinowe Materiałem genetycznym organizmów komórkowych jest kwas deoksyrybonukleinowy
Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Materiał genetyczny Materiałem genetycznym są kwasy nukleinowe Materiałem genetycznym organizmów komórkowych jest kwas deoksyrybonukleinowy
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Choroby genetyczne o złożonym
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Choroby genetyczne o złożonym
Dobór naturalny. Ewolucjonizm i eugenika
 Dobór naturalny Ewolucjonizm i eugenika Silna i słaba selekcja - symulacje W cieniu eugeniki Początki - XIX w. (Francis Galton) XX w. - eugenika totalitarna Poprawa jakości gatunku ludzkiego poprzez kierowanie
Dobór naturalny Ewolucjonizm i eugenika Silna i słaba selekcja - symulacje W cieniu eugeniki Początki - XIX w. (Francis Galton) XX w. - eugenika totalitarna Poprawa jakości gatunku ludzkiego poprzez kierowanie
Ekologia molekularna. wykład 6
 Ekologia molekularna wykład 6 Tempo mutacji Tempo błędu polimerazy: 10-4 pomyłka polimerazy 10-8 po naprawie błędów Faktyczne tempo mutacji: 10-9/zasadę/pokolenie W genomie człowieka jest 3 x 109 zasad
Ekologia molekularna wykład 6 Tempo mutacji Tempo błędu polimerazy: 10-4 pomyłka polimerazy 10-8 po naprawie błędów Faktyczne tempo mutacji: 10-9/zasadę/pokolenie W genomie człowieka jest 3 x 109 zasad
Podstawy genetyki populacji. Genetyka mendlowska i ewolucja.
 Podstawy genetyki populacji Genetyka mendlowska i ewolucja. Syntetyczna teoria ewolucji Pierwsza synteza: połączenie teorii ewolucji Darwina z genetyką mendlowską na poziomie populacji W naturalnych populacjach
Podstawy genetyki populacji Genetyka mendlowska i ewolucja. Syntetyczna teoria ewolucji Pierwsza synteza: połączenie teorii ewolucji Darwina z genetyką mendlowską na poziomie populacji W naturalnych populacjach
Analizy DNA in silico - czyli czego można szukać i co można znaleźć w sekwencjach nukleotydowych???
 Analizy DNA in silico - czyli czego można szukać i co można znaleźć w sekwencjach nukleotydowych??? Alfabet kwasów nukleinowych jest stosunkowo ubogi!!! Dla sekwencji DNA (RNA) stosuje się zasadniczo*
Analizy DNA in silico - czyli czego można szukać i co można znaleźć w sekwencjach nukleotydowych??? Alfabet kwasów nukleinowych jest stosunkowo ubogi!!! Dla sekwencji DNA (RNA) stosuje się zasadniczo*
Początki ewolucji. Historia komórek eukariotycznych i ich symbiontów
 Początki ewolucji Historia komórek eukariotycznych i ich symbiontów Współczesny obraz eukariogenezy Symbioza gospodarza - Archeona i symbionta - α-proteobakterii Powstanie jądra - ochrona przed inwazyjnymi
Początki ewolucji Historia komórek eukariotycznych i ich symbiontów Współczesny obraz eukariogenezy Symbioza gospodarza - Archeona i symbionta - α-proteobakterii Powstanie jądra - ochrona przed inwazyjnymi
ZARZĄDZANIE POPULACJAMI ZWIERZĄT 1. RÓWNOWAGA GENETYCZNA POPULACJI. Prowadzący: dr Wioleta Drobik Katedra Genetyki i Ogólnej Hodowli Zwierząt
 ZARZĄDZANIE POPULACJAMI ZWIERZĄT 1. RÓWNOWAGA GENETYCZNA POPULACJI Fot. W. Wołkow Prowadzący: dr Wioleta Drobik Katedra Genetyki i Ogólnej Hodowli Zwierząt POPULACJA Zbiór organizmów żywych, które łączy
ZARZĄDZANIE POPULACJAMI ZWIERZĄT 1. RÓWNOWAGA GENETYCZNA POPULACJI Fot. W. Wołkow Prowadzący: dr Wioleta Drobik Katedra Genetyki i Ogólnej Hodowli Zwierząt POPULACJA Zbiór organizmów żywych, które łączy
Podstawy biologii. Informacja, struktura i metabolizm.
 Podstawy biologii Informacja, struktura i metabolizm. Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Podstawy biologii Informacja, struktura i metabolizm. Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Mitochondrialna Ewa;
 Mitochondrialna Ewa; jej sprzymierzeńcy i wrogowie Lien Dybczyńska Zakład genetyki, Uniwersytet Warszawski 01.05.2004 Milion lat temu Ale co dalej??? I wtedy wkracza biologia molekularna Analiza różnic
Mitochondrialna Ewa; jej sprzymierzeńcy i wrogowie Lien Dybczyńska Zakład genetyki, Uniwersytet Warszawski 01.05.2004 Milion lat temu Ale co dalej??? I wtedy wkracza biologia molekularna Analiza różnic
Podstawy biologii. Informacja genetyczna. Co to jest ewolucja.
 Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Zarys biologii molekularnej genu Podstawowe procesy genetyczne Replikacja powielanie informacji Ekspresja wyrażanie (realizowanie funkcji)
Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Zarys biologii molekularnej genu Podstawowe procesy genetyczne Replikacja powielanie informacji Ekspresja wyrażanie (realizowanie funkcji)
Biologiczne podstawy ewolucji. Informacja genetyczna. Co to jest ewolucja.
 Biologiczne podstawy ewolucji. Informacja genetyczna. Co to jest ewolucja. Historia } Selekcja w hodowli zwierząt, co najmniej 10 000 lat temu } Sztuczne zapłodnienie (np. drzewa daktylowe) 1000 lat temu
Biologiczne podstawy ewolucji. Informacja genetyczna. Co to jest ewolucja. Historia } Selekcja w hodowli zwierząt, co najmniej 10 000 lat temu } Sztuczne zapłodnienie (np. drzewa daktylowe) 1000 lat temu
Dobór naturalny w sekwencjach genów. Metody analizy
 Dobór naturalny w sekwencjach genów Metody analizy Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Zmiany genetyczne w ewolucji } Mutacje } tworzą nowe allele genów } Inwersje } zmieniają
Dobór naturalny w sekwencjach genów Metody analizy Podstawy ewolucji molekularnej Ewolucja sekwencji DNA i białek Zmiany genetyczne w ewolucji } Mutacje } tworzą nowe allele genów } Inwersje } zmieniają
Podstawy biologii. Informacja, struktura i metabolizm.
 Podstawy biologii Informacja, struktura i metabolizm. Czym jest życie? Struktura Metabolizm Informacja (replikacja) Właściwości emergentne System jako całość ma właściwości nie będące sumą właściwości
Podstawy biologii Informacja, struktura i metabolizm. Czym jest życie? Struktura Metabolizm Informacja (replikacja) Właściwości emergentne System jako całość ma właściwości nie będące sumą właściwości
Biologia medyczna, materiały dla studentów
 Jaka tam ewolucja. Zanim trafię na jednego myślącego, muszę stoczyć bitwę zdziewięcioma orangutanami Carlos Ruis Zafon Wierzbownica drobnokwiatowa Fitosterole, garbniki, flawonoidy Właściwości przeciwzapalne,
Jaka tam ewolucja. Zanim trafię na jednego myślącego, muszę stoczyć bitwę zdziewięcioma orangutanami Carlos Ruis Zafon Wierzbownica drobnokwiatowa Fitosterole, garbniki, flawonoidy Właściwości przeciwzapalne,
października 2013: Elementarz biologii molekularnej. Wykład nr 2 BIOINFORMATYKA rok II
 10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
NaCoBeZu klasa 8 Dział Temat nacobezu programu I. Genetyka 1. Czym jest genetyka? 2. Nośnik informacji genetycznej DNA 3. Podziały komórkowe
 NaCoBeZu klasa 8 Dział programu Temat nacobezu I. Genetyka 1. Czym jest genetyka? wymieniam zakres badao genetyki rozróżniam cechy dziedziczne i niedziedziczne wskazuję cechy indywidualne i gatunkowe omawiam
NaCoBeZu klasa 8 Dział programu Temat nacobezu I. Genetyka 1. Czym jest genetyka? wymieniam zakres badao genetyki rozróżniam cechy dziedziczne i niedziedziczne wskazuję cechy indywidualne i gatunkowe omawiam
Podstawy genetyki populacji. Genetyka mendlowska i ewolucja
 Podstawy genetyki populacji Genetyka mendlowska i ewolucja Syntetyczna teoria ewolucji } Pierwsza synteza: połączenie teorii ewolucji Darwina z genetyką mendlowską na poziomie populacji } W naturalnych
Podstawy genetyki populacji Genetyka mendlowska i ewolucja Syntetyczna teoria ewolucji } Pierwsza synteza: połączenie teorii ewolucji Darwina z genetyką mendlowską na poziomie populacji } W naturalnych
Składniki jądrowego genomu człowieka
 Składniki jądrowego genomu człowieka Genom człowieka 3 000 Mpz (3x10 9, 100 cm) Geny i sekwencje związane z genami (900 Mpz, 30% g. jądrowego) DNA pozagenowy (2100 Mpz, 70%) DNA kodujący (90 Mpz ~ ok.
Składniki jądrowego genomu człowieka Genom człowieka 3 000 Mpz (3x10 9, 100 cm) Geny i sekwencje związane z genami (900 Mpz, 30% g. jądrowego) DNA pozagenowy (2100 Mpz, 70%) DNA kodujący (90 Mpz ~ ok.
1 Genetykapopulacyjna
 1 Genetykapopulacyjna Genetyka populacyjna zajmuje się badaniem częstości występowania poszczególnych alleli oraz genotypów w populacji. Bada także zmiany tych częstości spowodowane doborem naturalnym
1 Genetykapopulacyjna Genetyka populacyjna zajmuje się badaniem częstości występowania poszczególnych alleli oraz genotypów w populacji. Bada także zmiany tych częstości spowodowane doborem naturalnym
GENETYCZNE PODSTAWY ZMIENNOŚCI ORGANIZMÓW ZASADY DZIEDZICZENIA CECH PODSTAWY GENETYKI POPULACYJNEJ
 GENETYCZNE PODSTAWY ZMIENNOŚCI ORGANIZMÓW ZASADY DZIEDZICZENIA CECH PODSTAWY GENETYKI POPULACYJNEJ ZMIENNOŚĆ - występowanie dziedzicznych i niedziedzicznych różnic między osobnikami należącymi do tej samej
GENETYCZNE PODSTAWY ZMIENNOŚCI ORGANIZMÓW ZASADY DZIEDZICZENIA CECH PODSTAWY GENETYKI POPULACYJNEJ ZMIENNOŚĆ - występowanie dziedzicznych i niedziedzicznych różnic między osobnikami należącymi do tej samej
Biologiczne podstawy ewolucji. Informacja genetyczna. Co to jest ewolucja.
 Biologiczne podstawy ewolucji. Informacja genetyczna. Co to jest ewolucja. Historia } Selekcja w hodowli zwierząt, co najmniej 10 000 lat temu } Sztuczne zapłodnienie (np. drzewa daktylowe) 1000 lat temu
Biologiczne podstawy ewolucji. Informacja genetyczna. Co to jest ewolucja. Historia } Selekcja w hodowli zwierząt, co najmniej 10 000 lat temu } Sztuczne zapłodnienie (np. drzewa daktylowe) 1000 lat temu
Porównywanie i dopasowywanie sekwencji
 Porównywanie i dopasowywanie sekwencji Związek bioinformatyki z ewolucją Wraz ze wzrostem dostępności sekwencji DNA i białek pojawiła się nowa możliwość śledzenia ewolucji na poziomie molekularnym Ewolucja
Porównywanie i dopasowywanie sekwencji Związek bioinformatyki z ewolucją Wraz ze wzrostem dostępności sekwencji DNA i białek pojawiła się nowa możliwość śledzenia ewolucji na poziomie molekularnym Ewolucja
Wprowadzenie. DNA i białka. W uproszczeniu: program działania żywego organizmu zapisany jest w nici DNA i wykonuje się na maszynie białkowej.
 Wprowadzenie DNA i białka W uproszczeniu: program działania żywego organizmu zapisany jest w nici DNA i wykonuje się na maszynie białkowej. Białka: łańcuchy złożone z aminokwasów (kilkadziesiąt kilkadziesiąt
Wprowadzenie DNA i białka W uproszczeniu: program działania żywego organizmu zapisany jest w nici DNA i wykonuje się na maszynie białkowej. Białka: łańcuchy złożone z aminokwasów (kilkadziesiąt kilkadziesiąt
o cechach dziedziczonych decyduje środowisko, a gatunki mogą łatwo i spontanicznie przechodzić jedne w drugie
 Iwan Miczurin (1855-1935) Trofim Denisowicz Łysenko (1898-1976) przy interwencji człowieka możliwe jest zmuszenie każdej formy zwierzęcia lub rośliny do znacznie szybszych zmian, w kierunku pożądanym przez
Iwan Miczurin (1855-1935) Trofim Denisowicz Łysenko (1898-1976) przy interwencji człowieka możliwe jest zmuszenie każdej formy zwierzęcia lub rośliny do znacznie szybszych zmian, w kierunku pożądanym przez
Pamiętając o komplementarności zasad azotowych, dopisz sekwencję nukleotydów brakującej nici DNA. A C C G T G C C A A T C G A...
 1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
2. CZYNNIKI ZABURZAJĄCE RÓWNOWAGĘ GENETYCZNĄ
 ZARZĄDZANIE POPULACJAMI ZWIERZĄT 2. CZYNNIKI ZABURZAJĄCE RÓWNOWAGĘ GENETYCZNĄ POPULACJI Fot. W. Wołkow Prowadzący: dr Wioleta Drobik Katedra Genetyki i Ogólnej Hodowli Zwierząt MIGRACJE Zmiana frekwencji
ZARZĄDZANIE POPULACJAMI ZWIERZĄT 2. CZYNNIKI ZABURZAJĄCE RÓWNOWAGĘ GENETYCZNĄ POPULACJI Fot. W. Wołkow Prowadzący: dr Wioleta Drobik Katedra Genetyki i Ogólnej Hodowli Zwierząt MIGRACJE Zmiana frekwencji
Dopasowanie sekwencji (sequence alignment)
") Co to jest alignment? Dopasowanie sekwencji (sequence alignment) Alignment jest sposobem dopasowania struktur pierwszorzędowych DNA, RNA lub białek do zidentyfikowanych regionów w celu określenia podobieństwa;
Co to jest alignment? Dopasowanie sekwencji (sequence alignment) Alignment jest sposobem dopasowania struktur pierwszorzędowych DNA, RNA lub białek do zidentyfikowanych regionów w celu określenia podobieństwa;
Plan wykładów z genetyki ogólnej
 Plan wykładów z genetyki ogólnej 01 Metody genetyki klasycznej 02 Metody analizy DNA 03 Metody analizy genomu 04 Genomy prokariontów 05 Genomy eukariontów 06 Zmienność genomów w populacjach 07 Genomy a
Plan wykładów z genetyki ogólnej 01 Metody genetyki klasycznej 02 Metody analizy DNA 03 Metody analizy genomu 04 Genomy prokariontów 05 Genomy eukariontów 06 Zmienność genomów w populacjach 07 Genomy a
Nuttall przeprowadził testy precypitacyjne białek surowicy, aby wykazać związek filogenetyczny między różnymi grupami zwierząt.
 1904 Nuttall przeprowadził testy precypitacyjne białek surowicy, aby wykazać związek filogenetyczny między różnymi grupami zwierząt. M. Prakash 2007.Encyclopaedia of Gene Evolution Vol. 2, Molecular Genetics,
1904 Nuttall przeprowadził testy precypitacyjne białek surowicy, aby wykazać związek filogenetyczny między różnymi grupami zwierząt. M. Prakash 2007.Encyclopaedia of Gene Evolution Vol. 2, Molecular Genetics,
PORÓWNYWANIE POPULACJI POD WZGLĘDEM STRUKTURY
 PORÓWNYWANIE POPULACJI POD WZGLĘDEM STRUKTURY obliczanie dystansu dzielącego grupy (subpopulacje) wyrażonego za pomocą indeksu F Wrighta (fixation index) w modelu jednego locus 1 Ćwiczenia III Mgr Kaczmarek-Okrój
PORÓWNYWANIE POPULACJI POD WZGLĘDEM STRUKTURY obliczanie dystansu dzielącego grupy (subpopulacje) wyrażonego za pomocą indeksu F Wrighta (fixation index) w modelu jednego locus 1 Ćwiczenia III Mgr Kaczmarek-Okrój
Ekologia wyk. 1. wiedza z zakresu zarówno matematyki, biologii, fizyki, chemii, rozumienia modeli matematycznych
 Ekologia wyk. 1 wiedza z zakresu zarówno matematyki, biologii, fizyki, chemii, rozumienia modeli matematycznych Ochrona środowiska Ekologia jako dziedzina nauki jest nauką o zależnościach decydujących
Ekologia wyk. 1 wiedza z zakresu zarówno matematyki, biologii, fizyki, chemii, rozumienia modeli matematycznych Ochrona środowiska Ekologia jako dziedzina nauki jest nauką o zależnościach decydujących
Podstawy genetyki populacji. Genetyka mendlowska i ewolucja. Dobór i dryf.
 Podstawy genetyki populacji Genetyka mendlowska i ewolucja. Dobór i dryf. Równowaga Hardy ego-weinberga W populacji będącej w równowadze H-W częstości alleli nie zmieniają się Nie przebiega ewolucja Mechanizmy
Podstawy genetyki populacji Genetyka mendlowska i ewolucja. Dobór i dryf. Równowaga Hardy ego-weinberga W populacji będącej w równowadze H-W częstości alleli nie zmieniają się Nie przebiega ewolucja Mechanizmy
Podstawy biologii. Informacja genetyczna. Co to jest ewolucja.
 Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Historia } Selekcja w hodowli zwierząt, co najmniej 10 000 lat temu } Sztuczne zapłodnienie (np. drzewa daktylowe) 1000 lat temu } Podobne
Podstawy biologii Informacja genetyczna. Co to jest ewolucja. Historia } Selekcja w hodowli zwierząt, co najmniej 10 000 lat temu } Sztuczne zapłodnienie (np. drzewa daktylowe) 1000 lat temu } Podobne
Rozkład materiału z biologii dla klasy III AD. 7 godz / tyg rok szkolny 2016/17
 Rozkład materiału z biologii dla klasy III AD zakres rozszerzony LO 7 godz / tyg rok szkolny 2016/17 Biologia na czasie 2 zakres rozszerzony nr dopuszczenia 564/2/2012 Biologia na czasie 3 zakres rozszerzony
Rozkład materiału z biologii dla klasy III AD zakres rozszerzony LO 7 godz / tyg rok szkolny 2016/17 Biologia na czasie 2 zakres rozszerzony nr dopuszczenia 564/2/2012 Biologia na czasie 3 zakres rozszerzony
Wykład 14 Biosynteza białek
 BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
GENETYKA POPULACJI. Ćwiczenia 3 Biologia I MGR
 GENETYKA POPULACJI Ćwiczenia 3 Biologia I MGR Heterozygotyczność Rozpatrując różnorodność genetyczną w populacjach o układzie hierarchicznym zauważamy, że najwyższy poziom heterozygotyczności zawsze występuje
GENETYKA POPULACJI Ćwiczenia 3 Biologia I MGR Heterozygotyczność Rozpatrując różnorodność genetyczną w populacjach o układzie hierarchicznym zauważamy, że najwyższy poziom heterozygotyczności zawsze występuje
1 Podstawowe pojęcia z zakresu genetyki. 2 Podstawowy model dziedziczenia
 Rachunek Prawdopodobieństwa MAP8 Wydział Matematyki, Matematyka Stosowana Projekt - zastosowania rachunku prawdopodobieństwa w genetyce Opracowanie: Antonina Urbaniak Podstawowe pojęcia z zakresu genetyki
Rachunek Prawdopodobieństwa MAP8 Wydział Matematyki, Matematyka Stosowana Projekt - zastosowania rachunku prawdopodobieństwa w genetyce Opracowanie: Antonina Urbaniak Podstawowe pojęcia z zakresu genetyki
ZARZĄDZANIE POPULACJAMI ZWIERZĄT DRYF GENETYCZNY EFEKTYWNA WIELKOŚĆ POPULACJI PRZYROST INBREDU
 ZARZĄDZANIE POPULACJAMI ZWIERZĄT DRYF GENETYCZNY EFEKTYWNA WIELKOŚĆ POPULACJI PRZYROST INBREDU DRYF GENETYCZNY ) Każdy żywy organizm wytwarza więcej gamet, niż zdolne jest przetrwać (Darwin). 2) Przypadek
ZARZĄDZANIE POPULACJAMI ZWIERZĄT DRYF GENETYCZNY EFEKTYWNA WIELKOŚĆ POPULACJI PRZYROST INBREDU DRYF GENETYCZNY ) Każdy żywy organizm wytwarza więcej gamet, niż zdolne jest przetrwać (Darwin). 2) Przypadek
WPROWADZENIE DO GENETYKI MOLEKULARNEJ
 WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
Ewolucja człowieka. Ślady w ziemi i ślady w genach
 Ewolucja człowieka Ślady w ziemi i ślady w genach!1 !2 !3 Nie taka prosta historia } mtdna współczesnych Europejczyków różni się od mtdna Europejczyków sprzed 10 000 lat } fala migracji neolitycznej (~
Ewolucja człowieka Ślady w ziemi i ślady w genach!1 !2 !3 Nie taka prosta historia } mtdna współczesnych Europejczyków różni się od mtdna Europejczyków sprzed 10 000 lat } fala migracji neolitycznej (~
Wykład Bioinformatyka 2012-09-24. Bioinformatyka. Wykład 7. E. Banachowicz. Zakład Biofizyki Molekularnej IF UAM. Ewolucyjne podstawy Bioinformatyki
 Bioinformatyka Wykład 7 E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas 1 Plan Bioinformatyka Ewolucyjne podstawy Bioinformatyki Filogenetyka Bioinformatyczne narzędzia
Bioinformatyka Wykład 7 E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas 1 Plan Bioinformatyka Ewolucyjne podstawy Bioinformatyki Filogenetyka Bioinformatyczne narzędzia
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Budowa rybosomu Translacja
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Budowa rybosomu Translacja
Imię i nazwisko...kl...
 Gimnazjum nr 4 im. Ojca Świętego Jana Pawła II we Wrocławiu SPRAWDZIAN GENETYKA GR. A Imię i nazwisko...kl.... 1. Nauka o regułach i mechanizmach dziedziczenia to: (0-1pkt) a) cytologia b) biochemia c)
Gimnazjum nr 4 im. Ojca Świętego Jana Pawła II we Wrocławiu SPRAWDZIAN GENETYKA GR. A Imię i nazwisko...kl.... 1. Nauka o regułach i mechanizmach dziedziczenia to: (0-1pkt) a) cytologia b) biochemia c)
Szczegółowy harmonogram ćwiczeń Biologia medyczna w Zakładzie Biologii w roku akademickim 2017/2018 Analityka Medyczna I rok
 Szczegółowy harmonogram ćwiczeń Biologia medyczna w Zakładzie Biologii w roku akademickim 2017/2018 Analityka Medyczna I rok Przedmiot Wykłady Ćwiczenia Poniedziałek 8.00 10.15 grupa V Wtorek 11.00 13.15
Szczegółowy harmonogram ćwiczeń Biologia medyczna w Zakładzie Biologii w roku akademickim 2017/2018 Analityka Medyczna I rok Przedmiot Wykłady Ćwiczenia Poniedziałek 8.00 10.15 grupa V Wtorek 11.00 13.15
Selekcja, dobór hodowlany. ESPZiWP
 Selekcja, dobór hodowlany ESPZiWP Celem pracy hodowlanej jest genetyczne doskonalenie zwierząt w wyznaczonym kierunku. Trudno jest doskonalić zwierzęta już urodzone, ale można doskonalić populację w ten
Selekcja, dobór hodowlany ESPZiWP Celem pracy hodowlanej jest genetyczne doskonalenie zwierząt w wyznaczonym kierunku. Trudno jest doskonalić zwierzęta już urodzone, ale można doskonalić populację w ten
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii
 Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Dobór naturalny i dryf
 Dobór naturalny i dryf Efekty działania doboru Adaptacje Dostosowania do środowiska Egzaptacje Cechy, ktorych obecna funkcja jest inna niż pierwotna Np. pióra ptaków (kiedyś do zatrzymywania ciepła, obecnie
Dobór naturalny i dryf Efekty działania doboru Adaptacje Dostosowania do środowiska Egzaptacje Cechy, ktorych obecna funkcja jest inna niż pierwotna Np. pióra ptaków (kiedyś do zatrzymywania ciepła, obecnie
Konkurs szkolny Mistrz genetyki etap II
 onkurs szkolny istrz genetyki etap II 1.W D pewnego pierwotniaka tymina stanowi 28 % wszystkich zasad azotowych. blicz i zapisz, jaka jest zawartość procentowa każdej z pozostałych zasad w D tego pierwotniaka.
onkurs szkolny istrz genetyki etap II 1.W D pewnego pierwotniaka tymina stanowi 28 % wszystkich zasad azotowych. blicz i zapisz, jaka jest zawartość procentowa każdej z pozostałych zasad w D tego pierwotniaka.
Stochastyczna dynamika z opóźnieniem czasowym w grach ewolucyjnych oraz modelach ekspresji i regulacji genów
 Stochastyczna dynamika z opóźnieniem czasowym w grach ewolucyjnych oraz modelach ekspresji i regulacji genów Jacek Miękisz Instytut Matematyki Stosowanej i Mechaniki Uniwersytet Warszawski Warszawa 14
Stochastyczna dynamika z opóźnieniem czasowym w grach ewolucyjnych oraz modelach ekspresji i regulacji genów Jacek Miękisz Instytut Matematyki Stosowanej i Mechaniki Uniwersytet Warszawski Warszawa 14
PRZEDMIOTOWY SYSTEM OCENIANIA BIOLOGIA POZIOM ROZSZERZONY Opracowany w oparciu o program DKOS /02 KLASA III
 PRZEDMIOTOWY SYSTEM OCENIANIA BIOLOGIA POZIOM ROZSZERZONY Opracowany w oparciu o program DKOS 4015 5/02 ZAKRES WYMAGAŃ NA POSZCZEGÓLNE STOPNIE KLASA III DZIAŁ PROGRAMOWY I. Informacja genetyczna II. Przekazywanie
PRZEDMIOTOWY SYSTEM OCENIANIA BIOLOGIA POZIOM ROZSZERZONY Opracowany w oparciu o program DKOS 4015 5/02 ZAKRES WYMAGAŃ NA POSZCZEGÓLNE STOPNIE KLASA III DZIAŁ PROGRAMOWY I. Informacja genetyczna II. Przekazywanie
Geny i działania na nich
 Metody bioinformatyki Geny i działania na nich prof. dr hab. Jan Mulawka Trzy królestwa w biologii Prokaryota organizmy, których komórki nie zawierają jądra, np. bakterie Eukaryota - organizmy, których
Metody bioinformatyki Geny i działania na nich prof. dr hab. Jan Mulawka Trzy królestwa w biologii Prokaryota organizmy, których komórki nie zawierają jądra, np. bakterie Eukaryota - organizmy, których
Podstawy teorii ewolucji. Informacja i ewolucja
 Podstawy teorii ewolucji Informacja i ewolucja Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
Podstawy teorii ewolucji Informacja i ewolucja Informacje Kontakt: Paweł Golik Instytut Genetyki i Biotechnologii, Pawińskiego 5A pgolik@igib.uw.edu.pl Informacje, materiały: http://www.igib.uw.edu.pl/
ocena dopuszczająca ocena dostateczna ocena dobra ocena bardzo dobra Dział VII. EKOLOGIA NAUKA O ŚRODOWISKU
 Dział VII. EKOLOGIA NAUKA O ŚRODOWISKU wyróżnia elementy żywe i nieożywione w obserwowanym ekosystemie oblicza zagęszczenie wybranej rośliny na badanym terenie określa znaczenie wiedzy ekologicznej w życiu
Dział VII. EKOLOGIA NAUKA O ŚRODOWISKU wyróżnia elementy żywe i nieożywione w obserwowanym ekosystemie oblicza zagęszczenie wybranej rośliny na badanym terenie określa znaczenie wiedzy ekologicznej w życiu
Informacje dotyczące pracy kontrolnej
 Informacje dotyczące pracy kontrolnej Słuchacze, którzy z przyczyn usprawiedliwionych nie przystąpili do pracy kontrolnej lub otrzymali z niej ocenę negatywną zobowiązani są do dnia 06 grudnia 2015 r.
Informacje dotyczące pracy kontrolnej Słuchacze, którzy z przyczyn usprawiedliwionych nie przystąpili do pracy kontrolnej lub otrzymali z niej ocenę negatywną zobowiązani są do dnia 06 grudnia 2015 r.