Algorytmy Genetyczne (AG) dr inż. Tomasz Białaszewski
|
|
|
- Antoni Biernacki
- 7 lat temu
- Przeglądów:
Transkrypt
1 Algorytmy Genetyczne (AG) dr inż. Tomasz Białaszewski
2 Biologiczne tło AG Genetyka - badanie dziedziczności i zmienności organizmów Początek badań - XIX w. (Gregor Johann Mendel) Gen (z gr. genos - ród) - abstrakcyjna jednostki dziedziczności Allel różne odmiany genu
3 Biologiczne tło AG Chromosom (z gr. chromo barwa, soma ciało) struktury wewnątrz jąder komórek roślin i zwierząt zawierające geny Wszystkie komórki (z wyjątkiem rozrodczych) danego gatunku posiadają tę samą liczbę chromosomów Diplont organizm posiadający pary homologiczne chromosomów U organizmów płciowych dwa niesparowane chromosomy nazywane (X i Y) określają płeć
: Adenina (A) Cytozyna (C) Guanina (G) Tymina (T) DNA - struktura podwójnej helisy (taśmy) (James")
4 Biologiczne tło AG Chromosom kwas dezoksyrybonukleinowy (DNA Friedrich Miescher, 1869). Podstawowe elementy DNA (nukleotydy): Adenina (A) Cytozyna (C) Guanina (G) Tymina (T) DNA - struktura podwójnej helisy (taśmy) (James Watson i Francis Crick 1953) komplementarne nici
5 Biologiczne tło AG Replikacja - przenoszenie i kopiowanie genów (DNA) Synteza informacji w genach: Transkrypcja przepisanie genów do kwasu rybonukleinowego (RNA) Translacja - tłumaczenie fragmentów RNA na odpowiednie białka
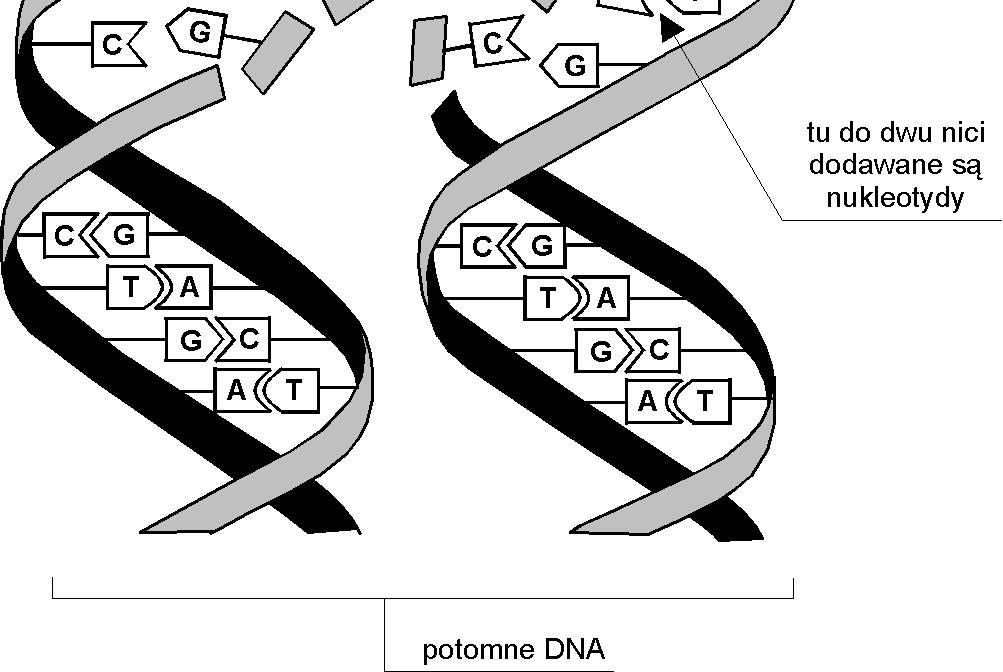
6 Replikacja DNA
7 Synteza RNA - transkrypcja
8 Aminokwasowa interpretacja kodonów nici informacyjnej mrna trzeci element kodonu A C G U AA Lizyna (Lys) Asparagina (Asn) Lizyna (Lys) Asparagina (Asn) AC Treonina (Thr) Treonina (Thr) Treonina (Thr) Treonina (Thr) AG Arginina (Arg) Seryna (Ser) Arginina (Arg) Seryna (Ser) AU Izoleucyna (Ile) Izoleucyna (Ile) Metionina (Met) - kodon startowy Izoleucyna (Ile) CA Glutamina (Gln) Histydyna (His) Glutamina (Gln) Histydyna (His) CC Prolina (Pro) Prolina (Pro) Prolina (Pro) Prolina (Pro) CG Arginina (Arg) Arginina (Arg) Arginina (Arg) Arginina (Arg) CU Leucyna (Leu) Leucyna (Leu) Leucyna (Leu) Leucyna (Leu) GA Kwas glutaminowy Kwas asparaginowy Kwas glutaminowy Kwas asparaginowy (Glu) (Asp) (Glu) (Asp) GC Alanina (Ala) Alanina (Ala) Alanina (Ala) Alanina (Ala) GG Glicyna (Gly) Glicyna (Gly) Glicyna (Gly) Glicyna (Gly) GU Walina (Val) Walina (Val) Walina (Val) Walina (Val) UA kodon stopowy Fenyloalanina (Phe) kodon stopowy Fenyloalanina (Phe) UC Seryna (Ser) Seryna (Ser) Seryna (Ser) Seryna (Ser) UG kodon stopowy Cysteina (Cys) Tryptofan (Trp) Cysteina (Cys) UU Leucyna (Leu) Fenyloalanina (Phe) Leucyna (Leu) Fenyloalanina (Phe)
9 Synteza białaka - translacaja mrna
10 Biologiczne tło AG Rekombinacja (krzyżowanie): poprzedza proces mejozy (tworzenie komórek rozrodczych) wymiana informacji genetycznej zawartej w chromosomach poprzez wymianę odcinków DNA między chromosomami z par homologicznych tworzenie nowych kombinacji genetycznych, czyli sekwencji nukleotydowych


11 Rekombinacja dla pary łańcuchów DNA z chromosomów homologicznych
12 Biologiczne tło AG W genetyce rozróżnia się cztery typy mutacji: tranzycja transwersja delecja insercja
13 Rodzaje mutacji
14 Wprowadzenie Metody optymalizacji (poszukiwań) można podzielić na trzy klasy: analityczne stochastyczne enumeracyjne
15 Metody poszukiwań
16 Wprowadzenie Podstawowe cechy algorytmów genetycznych: prowadzenie poszukiwań w wielu punktach parametry zadania optymalizacji posiadają postać zakodowaną do wyboru i tworzenia nowych rozwiązań stosuje się reguły ewolucyjno-probabilistyczne wykorzystanie do poszukiwań łatwo obliczalnej wartości funkcji celu wolne są od ograniczeń nakładanych na przestrzeń poszukiwań (np. ciągłość, istnienie pochodnych oraz unimodalność funkcji celu) uniwersalność i prostota implementacyjna
17 Cykl ewolucyjny AG Populacja osobników fenotyp SELEKCJA IE PODSTAWIENI OPERACJE GENETYCZNE Pula rodzicielska (rodzice) stopień przystosowania Funkcja celu Subpopulacja (potomkowie) fenotyp
18 Model populacji osobników
19 Model chromosomu w AG
20 Podstawowe oznaczenia Funkcja przystosowania x R n f ( x) 0 Genotyp osobnika haploidalnego (chromosom) v i [ ] T v v K v, i = 1,2,, N, ℵ = n 1i 2i n K i Populacja osobników haploidalnych V = [ ] v 1 v 2 K v N
21 Podstawowe oznaczenia Fenotyp osobnika haploidalnego x i = [ ] T n x x x R 1i 2 K i ni Stopień przystosowania osobnika haploidalnego f (x i ) R m
22 Podstawowe oznaczenia Genotyp osobnika diploidalnego gdzie v [ ] T v, i = 1,2, N di = 1 v, i 2 K i v v = [ v v K v ] T 1i 11i 21i n1i = [ v v K v ] T 2i 12i 22i n2i
23 Podstawowe oznaczenia Populacja osobników diploidalnych V d = [ ] v d1 v d 2 K v dn Fenotyp osobnika diploidalnego x i = [ ] T n x x x R Stopień przystosowania osobnika diploidalnego f 1i 2 K i (x i ) R m ni
24 Kodowanie informacji Rodzaje kodowania (reprezentacji parametrów): bialleliczne Gray a logarytmiczne trialleliczne multialleliczne wielopoziomowe
25 Kodowanie informacji Bialleliczne - odcinki (współrzędne ) chromosomu osobnika są skończonymi sekwencjami genów o allelach ze zbioru {0, 1} v i = [ ] T Gray a kodowanie bialleliczne (sąsiednie punkty w przestrzeni poszukiwań różnią się tylko jednym bitem w przestrzeni zakodowanych parametrów
26 Kodowanie informacji Logarytmiczne kodowanie bialleliczne zmniejszenie długości chromosomów dla ogromnych oraz wielowymiarowych przestrzeni poszukiwań bit najstarszy jest bitem znaku funkcji wykładniczej, zaś bit po nim następujący jest bitem znaku wykładnika funkcji wykładniczej pozostałe geny reprezentują wykładnik funkcji wykładniczej
27 Kodowanie informacji Przykład 1. Chromosom z kodowaniem logarytmicznym v i = [ ] T v i = 0 ( 1) 1 ( ) 0 ( 1) 0 (011011) ( 1) e ( 1) e T
28 Kodowanie informacji Przykład 2. Dla długości chromosomu m =10 i kodowania logarytmicznego można zakodować liczby rzeczywiste z przedziału W przypadku kodowania biallelicznego zakres wynosi [ 0,1023] [ ] e, e
29 Kodowanie informacji Trialleliczne - sekwencja genów, których allele należą do {-1, 0, 1} v di = T vd = v d Multialleliczne (zmiennopozycyjne) - naturalny sposób kodowania parametrów należących do dziedziny określonej w zbiorze liczb rzeczywistych T v i = x i R n
30 Kodowanie informacji Całkowitoliczbowe stosowane w zadaniach optymalizacji kombinatorycznej v i = [ ] T Wielopoziomowe - dla problemów parametrycznej i strukturalnej optymalizacji. Genotyp osobnika składa się z dwóch części: 7 sekwencji genów sterujących (strukturalnych) sekwencji genów kodujących parametry zadania optymalizacji (parametryczne)
31 Kodowanie informacji Przykładowe kodowanie dwupoziomowe
32 Dekodowanie informacji Dla kodowania binarnego gdzie x ki mk x, x ki = x k + x 2 k mk x k 1 mk j = 0 v kij - k-ta współrzędna wektora parametrów i-tego osobnika - długość sekwencji kodującej [ ] R k x k v k i 2 - zakres poszukiwań j x ki
33 Dekodowanie informacji Dla kodowania triallelicznego (dwie fazy): 1. Dominacja genetyczna 2. Liniowe odwzorowanie x ki = x k + x 2 k mk x k 1 mk j= 0 v kij 2 j
34 Ocena przystosowania Funkcja celu jest głównym źródłem oceny stopnia przystosowania każdego osobnika W algorytmach ewolucyjnych wymaga się, aby funkcja celu była funkcją przystosowania (często niespełniony warunek)
35 Ocena przystosowania Przekształcenie funkcji kosztu do funkcji przystosowania: u(x) R f ( x) = Cmax u( x) gdzie: C max - współczynnik nie mniejszy niż maksymalna wartość funkcji uzyskana w aktualnej populacji
36 Ocena przystosowania Przekształcenie funkcji zysku do funkcji przystosowania: g(x) R gdzie: f ( x) = g( x) C min C min - współczynnik nie większy niż minimalna wartość funkcji uzyskana w aktualnej populacji
37 Selekcja osobników Naśladuje mechanizm przeżycia w naturze Osobnik o najwyższym stopniu przystosowaniu uzyska liczne potomstwo, tj. pomnoży swój materiał genetyczny Osobniki o najwyższym stopniu przystosowaniu podnoszą w ten sposób prawdopodobieństwo swojego przeniknięcia ( przeżycia ) do następnego pokolenia Osobniki o najniższym stopniu przystosowania powinny być eliminowane z opisywanego procesu prokreacji
38 Selekcja osobników Najczęściej stosowane metody selekcji: metoda proporcjonalna metoda ze stochastycznym doborem resztowym metoda turniejowa metoda rangowa metoda progowa
39 Nacisk selektywny Naturalna tendencja do poprawiania średniego przystosowania populacji podczas cyklu ewolucyjnego Algorytm genetyczny posiada tym większy nacisk selektywny, im większa jest wartość oczekiwana liczby kopii lepszego osobnika w porównaniu do osobnika gorszego
40 Metoda proporcjonalna Konstruowanie tzw. ruletki, której sektory kątowe są proporcjonalne do wartości funkcji przystosowania osobników N krotne (N liczba osobników w populacji) ustalenie pozycji zakręcanego koła ruletki Lepiej przystosowane osobniki to proporcjonalnie większe sektory kątowe na tarczy ruletki (większa szansa na wprowadzenie większej liczby potomków do następnego pokolenia populacji
41 Metoda proporcjonalna 1. Wyznacz prawdopodobieństwa wyboru osobników z populacji w postaci względnego przystosowania p N s ( xi ) = f ( xi ) / f ( xi ), i = 1,2, K, i= 1 2. Wyznacz dystrybuantę dla ustalonej sekwencji osobników q( x ( x = i i) p s j= 1 j ) N
42 Metoda proporcjonalna 3. N-krotnie zakręć kołem ruletki poprzez: wygenerowanie losowej liczby r [ 0,1] ustalenie pierwszego napotkanego osobnika w populacji, dla którego spełniony jest warunek r q( x i )
43 Metoda proporcjonalna Przykład 3. Funkcja kryterialna 1, x2, x3) = 1300 określona w dziedzinie f ( x x x x x 1 [ 0,15] x 2 [ 0,7] x 3 [ 0,31] Populacja składa się z 4 osobników haploidalnych v i [ ] T v v v, 1,2,3, 4 = i = i i i odcinki chromosomu kodowane binarnie odpowiednio na 4, 3 i 5 bitach 2 3
44 Metoda proporcjonalna Genotyp, fenotyp i przystosowanie osobnika Lp. genotyp fenotyp przystosowanie bezwzględne względne dystrybuanta 1 [ ] T [6 6 31] T [ ] T [ ] T [ ] T [7 3 18] T [ ] T [4 2 8] T i = 1 f ( xi ) = 3091


45 Metoda proporcjonalna Koło ruletki dla rozważanego przykładu
46 Metoda stochastycznego doboru resztowego 1. Wyznacz oczekiwaną liczbę kopii osobników w populacji N i i i, i= 1 e ( x ) = N f ( x )/ f ( x ), i = 1,2, K, N 2. Skopiuj osobników do puli rodzicielskiej na podstawie całkowitych częściach liczby e( x i ), ~ oraz określ liczbę wakatów N = N N int
47 Metoda stochastycznego doboru resztowego 3. Wyznacz dystrybuantę osobników wg resztowej części e( x i ) i { e( x ) e( } q ( x ) x ) = i j j j = krotnie zakręć kołem ruletki poprzez: N ~ wygenerowanie losowej liczby ustalenie pierwszego napotkanego osobnika spełniającego warunek r q( x q( x i N ) ) [ ] r 0,1
48 Metoda stochastycznego doboru Dla przykładu 3 resztowego v ) i f ( x i ( i ) x q x ) e x e ) e( x ) ( i i ( i ( x q( q i x 4 ) ) p s e( x = i ) q( x 4 e( x ) i ) v 1 v 2 v 3 v i = 1 f ( xi ) = 3091


49 Metoda stochastycznego doboru resztowego Koło ruletki dla rozważanego przykładu
50 Metoda turniejowa Sposób selekcji został skojarzony z procesem rywalizacji dwóch osobników, z których wygrywa lepiej przystosowany Liczba osobników uczestniczących w turnieju decyduje o tzw. nacisku selekcyjnym (ang. selection pressure) Jeżeli liczba uczestników turnieju jest duża, wówczas słabsze osobniki mają mniejszą szansę na wybranie do puli rodzicielskiej
51 Metoda turniejowa 1. Wybierz losowo z rozkładem równiomiernym r osobników z populacji N osobników (r<n) 2. Przeprowadź turniej dla r-osobników poprzez: wyznaczenie zwycięzcy spełniającego warunek f ( xi ) f ( x j ) i j i skopiowanie zwycięzcy turnieju do puli rodzicielskiej 3. powróć do kroku 1 i powtórz cały cykl N 1 krotnie
52 Metoda turniejowa Proces wyboru osobników może być przeprowadzany w dwojaki sposób poprzez: losowanie ze zwracaniem losowanie bez zwracania Gdy warunek zwycięzcy nierozstrzygnięty wówczas zwycięzca jest wybierany losowo Jeżeli grupa turniejowa składa się z dwóch osobników to selekcja nazywana jest turniejem binarnym
53 Metoda rangowa Stopień przystosowania jest reprezentowany przez rangę, która nadawana jest w procesie sortowania osobników w populacji Sortowanie polega na uszeregowaniu osobników w populacji według nierosnących wartości funkcji przystosowania Dwa sposoby przypisywania rang: ranga jest równa indeksowi (pozycji) w uszeregowanej populacji ranga identyczna dla osobników identycznych
54 Metoda rangowa 1. Posortuj osobniki w populacji według nierosnących wartości funkcji przystosowania 2. Nadaj każdemu osobnikowi rangę r( x i przypisanie kolejnego indeksu (pozycji) w uszeregowanej populacji albo przypisanie jednakowej pozycji dla osobników o identycznym stopniu przystosowania )
55 Metoda rangowa 3. Wyznacz prawdopodobieństwa wyboru osobników wg odwzorowania: liniowego p ( x lub nieliniowego s i ) r ( x = ) a + k 1 i r = max gdzie p ( x ) = a + k rmax r( x ) s r i max = max i ( ) b r( x ) { } i i
56 Metoda rangowa a, k, b parametry dobieranie arbitralnie aby spełnione były następujące warunki: N i=1 p ( x ) = 1 0 p ( x ) 1, s jeżeli i 4. Na podstawie prawdopodobieństw wyboru przeprowadź selekcje według metody proporcjonalnej lub metody ze stochastycznym doborem resztowym s i r( xi ) r( x j ) to ps ( xi ) ps ( x j )
57 Metoda progowa Sortowanie osobników i nadawanie rang Prawdopodobieństwo wyboru osobnika gdzie γ p s ( x i ) 1 = γ 0 dla dla 0 r ( x r( x i ) γ > γ - parametr sterujący naciskiem selektywnym Zmniejszanie wartości γ zwiększa nacisk selektywny i )
58 Operacje genetyczne Operacje genetyczne wykonywane na osobnikach rodzicielskich: krzyżowanie oraz mutacja. Chromosom osobnika traktowany jest jako pojedyncza sekwencja kodowa Dla kodowania multiallelicznego operacje genetyczne wykonuje się odrębnie dla poszczególnych współrzędnych (genów)
59 Krzyżowanie Krzyżowanie (ang. crossover) analogiczna do rekombinacji pomiędzy nićmi DNA chromosomów z par homologicznych W GA krzyżowanie polega na wymianie materiału genetycznego (części chromosomu) między dwoma rodzicami W wyniku tego uzyskuje się dwóch potomków stanowiących nowe rozwiązania
60 Krzyżowanie Krzyżowanie przebiega w dwóch etapach: 1. Kojarzenie w sposób losowy osobników z puli rodzicielskiej w pary 2. Każda para rodziców przechodzi proces wymiany materiału genetycznego (tj. wymiany fragmentów sekwencji kodowych pomiędzy chromosomami osobników rodzicielskich) Krzyżowanie zachodzi z pewnym prawdopodobieństwem p c [0.6, 1] nazywanym prawdopodobieństwem krzyżowania
61 Podstawowe sposoby krzyżowania Najczęściej stosowanym sposobem krzyżowania jest: krzyżowanie jednopunktowe, krzyżowanie wielopunktowe (zwykle dwulub trójpunktowe) oraz krzyżowanie jednorodne.

62 Podstawowe sposoby krzyżowania jednopunktowe trójpunktowe
63 Podstawowe sposoby krzyżowania Jednopunktowe dla osobników diploidalnych
64 Podstawowe sposoby krzyżowania Przykład 4. Krzyżowanie jednopunktowe dla osobników kodowanych binarnie v r v s = = [ ] T [ ] T Punkt krzyżowania pomiędzy 10 a 11 bitem v r v s ' = [ ] T ' = [ ] T
65 Podstawowe sposoby krzyżowania Krzyżowanie równomierne (jednolite) (ang. uniform crossover) Przeprowadzane wg binarnego wzorca krzyżowania generowanego losowo dla każdej pary rodzicielskich osobników Allele równe 1 z takiego wzorca decydują o wymianie pomiędzy rodzicami genów z pozycji im odpowiadającym
66 Podstawowe sposoby krzyżowania Przykład 5. Krzyżowanie jednorodne v r v s = = [ ] T [ ] T Wylosowany wzorzec krzyżowania w = Uzyskani potomkowie v r v s [ ] T ' = [ ] T ' = [ ] T
67 Podstawowe sposoby krzyżowania Dla reprezentacji zmiennopozycyjnej stosuje się: krzyżowanie arytmetyczne lub krzyżowanie mieszane Utworzeni potomkowie stanowią rozwiązania dopuszczalne, tzn. należą do dziedziny określonej na podstawie zakresu parametrów i innych liniowych ograniczeń nałożonych na te parametry
68 Podstawowe sposoby krzyżowania Krzyżowanie arytmetyczne - unormowana liniowa kombinacji dwóch wektorów v r = [ v v K v ] T 1r 2r nr Potomkowie wyznaczani z poniższych formuł ' r gdzie v s = [ v v K v ] T 1s 2s ns ' = avr + (1 a vs v s = avs + (1 a) vr v ) a [0,1], - wybierana losowo dla pary rodziców
69 v v ' r ' s Podstawowe sposoby krzyżowania Krzyżowanie mieszane (proste) - połączenie krzyżowania jednopunktowego z krzyżowaniem arytmetycznym v v r s = = [ v ] 1 v2 K v T r r nr [ v v K v ] T 1s 2s ns Potomkowie wyznaczani z poniższych formuł [ v K v av + (1 a) v av + (1 a) v ] T = 1r kr ( k+ 1) s ( k+ 1) K r [ v K v av + (1 a) v av + (1 a) v ] T = 1s ks ( k+ 1) r ( k+ 1) K s ns nr nr ns
70 Podstawowe sposoby krzyżowania Krzyżowanie wieloosobnicze - do utworzenia potomków wybieranych jest losowo więcej niż dwóch rodziców Dla reprezentacji fenotypowej gdzie a j v - losowe liczby z rozkładem równomiernym, przy warunku ' = k i a j j= 1 k j v j =1 a =1 j
71 Podstawowe sposoby krzyżowania punkty krzyżowania Trójosobnicze dwu upunktowe rodzic v 1 rodzic v 2 rodzic v 3 ' potomek v 1 ' potomek v 2 ' potomek v 3
72 Podstawowe sposoby krzyżowania Dla kodowania całkowitoliczbowego stosuje się zwykle krzyżowanie: z częściowym odwzorowaniem (ang. partialy-mapped crossover, PMX) z zachowaniem porządku (ang. order crossover, OX) cykliczne (ang. cycle crossover, CX)
73 Podstawowe sposoby krzyżowania Krzyżowanie PMX polega na wylosowaniu dwóch punktów krzyżowania dla wybranej losowo pary rodziców Wybrana sekwencja genów ulega zamianie Sekwencje pozostałych genów są tak dobierane, aby zachować porządek genów i ich pozycje w obu powstających potomkach
74 Podstawowe sposoby krzyżowania Przykład 6. Krzyżowanie PMX v r v s = = [ ] T [ ] T Wymiana sekwencji genów pomiędzy punktami krzyżowań Pozostałe pozycje nieokreślone 5 v ' r =?? ??? 4 [ ] T v ' s =?? ??? [ ] T
75 Podstawowe sposoby krzyżowania Uzupełnianie pozycji nieokreślonych genami, dla których nie występuje konflikt Niedopuszczalne powtórzenia alleli w osobniku potomnym (każde rozwiązanie jest permutacją 9-elementowego ciągu cyfr od 1 do 9 v ' r =? ? [ ] T v ' s = 1? ? 2 3 [ ] T
76 Podstawowe sposoby krzyżowania Wypełnianie pozycji nieokreślonych wg ciągu odwzorowań wymiany sekwencji genów (określony na podstawie punktów krzyżowań) 8 7, 7 5, 5 4, 9 6 Ostatecznie uzyskani potomkowie v ' r = v ' s = [ ] T [ ] T
77 Podstawowe sposoby krzyżowania Krzyżowanie OX polega na skopiowaniu losowych sekwencji genów ustalonych na podstawie dwóch punktów krzyżowań Nieokreślone pozycje w tworzonych nowych rozwiązaniach wyznaczane są na podstawie wzajemnego uporządkowania genów jednego z osobników rodzicielskich
78 Podstawowe sposoby krzyżowania Przykład 7. Krzyżowanie OX v r v s = = [ ] T [ ] T Kopiowanie sekwencji genów pomiędzy punktami krzyżowań Pozostałe pozycje nieokreślone v ' r =?? ??? 4 [ ] T v ' s =?? ??? [ ] T
79 Podstawowe sposoby krzyżowania Tworzenie potomka v r wg sekwencji genów chromosomu v s począwszy od drugiego punktu krzyżowania ~ = v s Usunięcie z chromosomu v ~ s genów o allelach ' powtarzających się dla potomka ~ = v s ' [ ] T v r [ ] T
80 Podstawowe sposoby krzyżowania Uzyskany ciąg genów z v ~ s umieszczany w ' potomku v r począwszy od pozycji określonej drugi punktem krzyżowania v ' r = [ ] T ' v s Potomek określany podobnie ~ = v r ~ = [ ] T [ ] T v r v ' s = [ ] T 7 5 9
81 Podstawowe sposoby krzyżowania Krzyżowanie CX w odróżnieniu od PMX i OX nie wykorzystuje punków krzyżowania do generowania potomków Operacja ta polega na odpowiednich kopiowaniu ciągów genów z wybranego rodzica według pozycji określonych drugi rodzicem
82 Podstawowe sposoby krzyżowania Przykład 8. Krzyżowanie CX v r v s = = [ ] T [ ] T Rozpoczęcie tworzenia potomka poprzez skopiowania pierwszego genu z rodzica ' v r v ' r = 4???????? [ ] T v r
83 Podstawowe sposoby krzyżowania Pozycja skopiowanego genu wyznacza w rodzicu v s następny gen jaki należy skopiować do potomka ' v r zgodnie z jego pozycją określoną w rodzicu v r v ' r = 4????? 1?? [ ] T v ' r = 4? 8??? 1?? [ ] T Powyższa reguła realizowana do momentu uzyskania tzw. zamkniętej sekwencji genów v ' r = 4? 8 7 5? 1?? [ ] T
84 Podstawowe sposoby krzyżowania Kopiowanie genów z rodzica v s określonych ' pozycjami nieokreślonymi potomka v ' r = v r [ ] T Wyznaczenie drugiego potomka v ' s = 1???????? v ' s = 1??? 4???? v ' s = 1? 7 5 4? 8?? v ' s = [ ] T [ ] T [ ] T [ ] T
85 Mutacja Mutacja - przypadkowe zaburzenia w nowo powstałych rozwiązaniach (potomkach) Ze względu na obiekt modyfikacji, mutacje można podzielić na dwie grupy: mutacje genotypowe (genetyczne) oraz mutacje fenotypowe (parametryczne) Mutacja - zmiana alleli genów z założonym prawdopodobieństwem p m całej populacji osobników
86 Mutacja genotypowa Mutacja genotypowa analogiczna do istniejącego mechanizmu tranzycji występującego przy mutacji nici DNA Dla kodowania binarnego - przypadkowa zmiana allelu genu w chromosomie na przeciwny (zanegowanie wybranego bitu kodu binarnego

87 Mutacja genotypowa Przykład mutacji genotypowej dla kodu binarnego
88 Mutacja genotypowa Schemat mutacji genotypowej dla kodu triallelicznego
89 Mutacja fenotypowa Mutacja fenotypowa wprowadza zmianę wybranych parametrów Zmutowane osobniki są rozwiązaniami dopuszczalnymi (należą do dziedziny parametrów i liniowych ograniczeń na te parametry) Sposoby modyfikacji wartości parametru polegające na zmianie wartościowo-równomiernej wartościowo-nierównomiernej
90 Mutacja fenotypowa Zmiana wartościowo-równomierna - nadawanie wybranemu parametrowi v kq osobnika v q losowej wartości z przedziału określającego dziedzinę mutowanego k-tego parametru z równomiernym rozkładem prawdopodobieństwa Potomek o nowym fenotypie: v ' q = [ ' ] v v K v K v T 1 1 k n
91 Mutacja fenotypowa W zadaniach z liniowymi ograniczeniami dziedzina mutowanego parametru musi być określana wg narzuconych ograniczeń przy uwzględnieniu niezmienionych, pozostałych wartości parametrów w fenotypie Przymiotnik równomierna oznacza, że działanie mutacji jest niezależne od numeru iteracji algorytmu
92 Mutacja fenotypowa Zmiana wartościowo-nierównomierna pozwala na dokładne dostrojenie parametrów Stochastyczne zaburzenie poprzednio ustalonej wartości wybranego parametru z zastosowaniem zawężającego się wraz ze wzrostem numeru iteracji dopuszczalnego przedziału
93 Mutacja fenotypowa Mutowany parametr v osobnika k q vq v ' kq = v v k k gdzie b + ( t, vk v ) (A) q kq t (B), ( t, v) = r v 1 ( t, v k v k ) q q t max [ v, ] R t k v k, t max r [ 0,1] - dziedzina parametru - kolejne, ostatnie pokolenie - współczynnik niejednorodności (b=2) - losowa z rozkładem równomiernym b
94 Mutacja Inne mechanizmy mutacji stosowane dla genotypowej jak i fenotypowej struktury osobnika: inwersja (ang. inversion) wstawianie (ang. insertion) przenoszenie (ang. displacement) wzajemna wymiana (ang. reciprocal exchange)
95 Mutacja Inwersja - wylosowanie w chromosomie podciągu kodowego (sekwencji genów między dwoma losowo wybranymi pozycjami) i odwróceniu kolejności pozycji allei Wstawianie - wylosowanie pojedynczego genu i umiejscowieniu go w losowo wybranej pozycji w chromosomie osobnika
96 Mutacja Przenoszenie wylosowywany podciąg kodowy wstawiany jest w losowo wybraną pozycje w chromosomie (uogólnienie mechanizmu wstawiania) Wzajemna wymiana wylosowanie k-par genów do zamiany pozycji w chromosomie
97 Mutacja osobnik Schematy operacji genetycznych: a) inwersji b) wstawiania c) przenoszenia d) wzajemnej wymiany osobnik zmutowany osobnik osobnik zmutowany osobnik osobnik zmutowany (a) (b) (c) osobnik osobnik zmutowany (d)
98 Strategie podstawień Proces zastępowania populacji uzyskanych w kolejnych pokoleniach Stosowane strategie podstawień reprodukcja pełna reprodukcja częściowa reprodukcja według czasu życia osobników
99 Strategie podstawień Strategia z reprodukcją pełną - nowo powstała populacja o liczbie osobników N zastępuje całą poprzednią populację (najprostszy mechanizm) Strategia z częściową reprodukcją - zastąpienie wybranych osobników przez nowo powstałych potomków (możliwość posiadania wielopokoleniowego potomstwa)
100 Strategia z częściową reprodukcją Strategia z częściową reprodukcją realizowana na trzy sposoby poprzez: zastępowanie najgorszych osobników (elityzm) zastępowanie osobników najbardziej podobnych do potomnych (metoda ze ściskiem) zastępowanie losowo wybranych osobników
101 Strategia z częściową reprodukcją Usuwanie z populacji osobników z najniższym stopniem przystosowania i w ich miejsce wstawianie potomków lepiej przystosowanych Operacja ta polega na: łącznym sortowaniu według niemalejących stopni przystosowania rodziców i potomków wybraniu N-pierwszych najlepszych osobników Strategia elitarna może spowodować wystąpienie zjawiska przedwczesnej zbieżności (tworzenie elity)
102 Strategia z częściową reprodukcją Metoda ze ściskiem - usuwania osobników z największym podobieństwem w sensie: genotypowym (w sensie odległości Hamminga) fenotypowym (według normy euklidesowej) Potomek porównywany jest genotypowo lub fenotypowo z k -rodzicami wybieranymi losowo z rozkładem równomiernym ( k N ) Najbardziej podobny rodzic usuwany z populacji i zastępowany potomkiem Wartość k nazywana współczynnikiem ścisku lub stłoczenia osobników w populacji o rozmiarze N
103 Strategia z reprodukcją wg czasu życia Przeprowadzana jest na podstawie przypisanego osobnikom dodatkowego wskaźnika zwanego czasem życia Wskaźnik życia zmniejsza się w kolejnych pokoleniach Z populacji usuwane są osobniki, których czas życia jest mniejszy lub równy zeru Nowa populacja jest ustalana na podstawie wyboru najlepszych (według stopnia przystosowania) osobników rodzicielskich i potomnych
104 Strategia z reprodukcją wg czasu życia Przypisanie osobnikom dodatkowego wskaźnika - czasu życia Z populacji usuwane są osobniki o czasie życia mniejszym lub równy zeru Nowa populacja ustalana na podstawie wyboru najlepszych rodziców i potomków Sposoby strategii wg: stałego czasu życia osobnika zmiennego czasu życia
105 Strategia z reprodukcją wg czasu życia Reprodukcja według stałego czasu życia osobników - przypisanie wszystkim osobnikom pewnego arbitralnie dobranego maksymalnego czasu życia Reprodukcja według zmiennego czasu życia osobników: populacje w kolejnych pokoleniach mogą przyjmować różne rozmiary pula rodzicielska jest wyłaniania poprzez losowy wybór osobników z populacji z rozkładem równomiernym
106 Strategia z reprodukcją wg czasu życia Czas życia osobnika l( x i ) określany na podstawie jego stopnia przystosowania za pomocą trzech możliwych przekształceń: liniowego proporcjonalnego biliniowego
107 Strategia z reprodukcją wg czasu życia Stosowane oznaczenia: - minimalny czas życia l min l max l = avg f f f min avg max - maksymalny czas życia l max ( t) = + 2 l min min { - średni czas życia min t= t1, K, t2 i= 1,2, K, N 1 t ( t) = 2 t t ( t) = max f ( x N = = ( t i f xi 1 N 1 1 ) { max t= t1, K, t2 i= 1,2, K, N i f ( x i ) } ) }
108 Strategia z reprodukcją wg czasu życia Stosowane oznaczenia: - numery pokoleń wyznaczające okno czasowe t = t 1,t 2 t 2 t 1 Wartość wpływa na nacisk selektywny t Małe okno czasowe powoduje większy nacisk selektywny Duże okno czasowe wartość daje mniejszy nacisk selektywny
109 Strategia z reprodukcją wg czasu życia Przekształcenie za pomocą funkcji: liniowej l max l l( x min i) = l min + f ( x i f ( t) f ( t) proporcjonalnej max min lavg lmin l( xi ) = min lmax, lmin + f ( xi f ( t) f ( t) avg ) min )
110 Strategia z reprodukcją wg czasu życia Przekształcenie za pomocą funkcji: biliniowej l( x i ) = l l min avg + + f f avg max l avg l ( t) f lmax l ( t) f min min avg avg ( t) ( t) f f ( x i ( x i ), ), f f ( x i ( x i ) < ) f f avg avg ( t) ( t)
111 Strategia z reprodukcją wg czasu życia zmiana funkcja f min ( t) f avg ( t) f avg ( t) f max ( t) liniowa proporcjonalna biliniowa zwiększenie średniego czasu życia oraz osłabienie nacisku selektywnego brak wpływu brak wpływu zmniejszenie średniego czasu życia oraz zmniejszenie nacisku selektywnego zwiększenie średniego czasu życia oraz osłabienie nacisku selektywnego zwiększenie liczby osobników o maksymalnym czasie życia zmniejszenie liczby osobników o maksymalnym czasie życia brak wpływu osłabienie nacisku selektywnego dla osobników o stopniu przystosowania poniżej średniej zmniejszenie nacisku selektywnego dla osobników lepszych oraz zwiększenie nacisku selektywnego dla osobników gorszych zwiększenie nacisku selektywnego dla osobników lepszych oraz zmniejszenie nacisku selektywnego dla osobników gorszych zmniejszenie nacisku selektywnego dla osobników powyżej przeciętnej
112 Skalowanie przystosowania Zjawisko przedwczesnej zbieżności (ang. premature convergance) do suboptymalnego rozwiązania Metody skalowania przystosowania: liniowe z odchyleniem standardowym (sigma-obcinające) wykładnicze (potęgowe)
113 Skalowanie liniowe Zmodyfikowane przystosowanie osobników fs ( xi) = a f ( xi ) + Warunki dobierania parametrów a i b: średnie przystosowanie populacji jest niezmienne po skalowaniu zmodyfikowane maksymalne przystosowania osobników równe wielokrotności średniego przystosowania populacji przeskalowane wartości funkcji przystosowania nie mogą przyjmować wartości ujemnych. b
114 Schemat skalowania liniowego 1. Znajdź w populacji osobnika o maksymalnym i minimalnym przystosowaniu f max f min = = max i { {1,2, K, N } min i {1,2, K, N} f ( x 2. Wyznacz średnie przystosowanie populacji f i ( x 1 N favg = i = f i N 1 ( x ) i ) )
115 Schemat skalowania liniowego 3. Przeskaluj stopnie przystosowania osobników fs ( xi) = a f ( xi ) + b a i b współczynniki skalowania linowego wartości wyznaczane następująco: sprawdź warunek skalowania f min > c f avg c 1 (*) c - współczynnik zwielokrotnienia (liczba kopii) f max
116 Schemat skalowania liniowego jeżeli warunek skalowania (*) spełniony to a favg ( c 1) =, b = favg ( a f f max avg w przeciwnym przypadku favg a =, f f avg min b = af min + 1) W których pokoleniach cyklu GA są wybierane odpowiednie parametry a i b?
117 Skalowanie σ-obcinające Zmodyfikowane przystosowanie osobników f s ( x ) = f ( x ) + ( f c σ) i i avg gdzie σ = 1 N ( f ( 2 xi ) f avg ) N 1 i= c - liczba naturalna (zwykle od 1 do 5)
118 Skalowanie wykładnicze Zmodyfikowane przystosowanie osobników gdzie f ( x ) = f ( x ) s i k - liczba zwykle bliską jedności W zależności od problemu optymalizacji wartość k powinna być dobrana odpowiednio Należy zadbać by zmieniała się podczas cyklu ewolucyjnego i k ( k = 1.005)
119 Niszowanie Zachowanie gorzej przystosowanych osobników by populacja była różnorodna Pozwala utrzymać stałą liczbę istniejących gatunków zarówno: bardziej licznych (lepiej przystosowanych) mniej licznych (słabiej przystosowanych). Nisza w GA jest pewnym obszarem, w którym znajdują się osobniki bliskie parametrycznie (fenotypowo, geometrycznie), które powinny mieć także podobne stopnie przystosowania
120 Mechanizm niszowania Funkcja bliskości (ang. sharing function) δ ij gdzie = 1 x 0 x i i x P x j P j P = jeśli 0 jeśli ( x i x j ) T x i x j P < 1 x i x j 1 P 2 ( x { φ φ } = diag 1 2 K φ n i P x j ) φ k ( k = 1,2,..., n) - k-ta średnica hiperelipsoidy o środku w i-tym osobniku
121 Mechanizm niszowania k-ta średnica hiperelipsoidy gdzie φ k k =, k = 1,2, K, n ε k ε - niezerowym zakresem poszukiwań k-tego parametru - liczba części, na jaką dzieli się zakres k
122 Mechanizm niszowania
123 Mechanizm niszowania
124 Mechanizm niszowania Modyfikacja stopnia przystosowania lub rangi każdego osobnika znajdującego się we własnej niszy według następującego przepisu ~ f ( = i) f ( xi ) x = δ N j 1 ij ~ r( = i) r ( xi ) x = δ Suma w obu mianownikach wyraża efektywną liczbę osobników w niszy własnej o środku opisanym przez fenotyp rozważanego i-tego osobnika N j 1 ij
125 Mechanizm niszowania 7 x 2 7 f x f 1
126 Mechanizm niszowania 7 ~ f ~ f
127 Teoria schematów Metodologię procesu poszukiwań GA można interpretować w kategoriach schematów Schemat - wzorzec opisujący podzbiór sekwencji kodowych podobnych pod względem pozycji o ustalonych wartościach zwanych pozycjami ustalonymi Schemat składa się z cyfr kodu binarnego i dodatkowego znak # Symboli # oznacza nieustaloną wartość pozycji
128 Teoria schematów Przykład 9. Schematy S [ # # 0 1 ] T 1 = 1 Do schematu pasują cztery chromosomy v v v v S 1 [ ] T 1 = 1 [ ] T 2 = 1 [ ] T 3 = [ ] T 4 = 1
129 Teoria schematów Schemat posiada następujące dwie istotne własności: rząd rozpiętość Rzędem schematu nazywa się liczbę ustalonych pozycji, tzn. liczbę 0 i 1 w schemacie Rozpiętością schematu nazywa się odległość między skrajnymi ustalonymi pozycjami, tzn. genami o ustalonej wartości (allelu) 0 lub 1
130 Teoria schematów Liczba osobników pasujących do danego schematu S po selekcji metodą proporcjonalną gdzie ξ + = ξ t ( S t 1( S) t ( S) N N i = 1 ft F ) 1 j Ft ( S) = i = f t i j 1 ( x ) ( x i )
131 Teoria schematów - efekt selekcji Średni stopień przystosowania populacji Zatem 1 N favg ( t) = i = f t i N 1 ( x ) ξ t + 1( S) = ξ t ( S) F f t avg ( S) ( t) Schematy o przystosowaniu wyższym niż średnie przystosowanie populacji mają większą liczbę reprezentantów w następnym pokoleniu
132 Teoria schematów - efekt selekcji Jeżeli schemat S ma przystosowanie większe od średniego przystosowania populacji o pewną stałą ε F t ( S ) = f ( t ) + ε f ( t ) ξ t + avg avg 1( S) = (1 + ε) ξt ( S) Poczynając od 0 i zakładając, że ε nie zmienia się ξ t + 1( ) = ξ0( S) (1 + ε S ) t
133 Teoria schematów - efekt selekcji Przykład 9. Schematy i ich przystosowanie przed selekcją metodą proporcjonalną Lp. S i v i F t (S i ) ξ t (S i ) 1 [#### 0## #####] T v 3 =[ ] T v 4 =[ ] T [0### ### 0####] T v 4 =[ ] T [#### #10 #####] T v 1 =[ ] T v 4 =[ ] T
134 Teoria schematów - efekt selekcji Schematy po selekcji metodą proporcjonalną Lp. S i v i F t+1 (S i ) ξ t+1 (S i ) 1 [#### 0## #####] T v 3 =[ ] T v 4 =[ ] T [0### ### 0####] T v 4 =[ ] T [#### #10 #####] T v 4 =[ ] T
135 Teoria schematów - efekt krzyżowania Zakłada się krzyżowanie jednopunktowe dla osobników hapliodalnych kodowanych binarnie Prawdopodobieństwo destrukcji (utraty) p d (S) oraz przeżycia (zachowania) p d (S) schematu S gdzie δ(s) m δ( S) δ( S) p d ( S) = p d ( S) = 1 m 1 m 1 - rozpiętość schematu S - długość chromosomu pasującego do S
136 Teoria schematów - efekt krzyżowania Przykład 10. Krzyżowanie jednopunktowe pary rodzicielskiej dla wybranego punktu krzyżowania między bitem 5 a 6 [ ] T = v Rozważmy dwa schematy [ ] T = v [ ] T = v [ ] T # # # # # # 1 # # = S [ ] T 2 # # # # # # 1 0 # # # = S
137 Teoria schematów - efekt krzyżowania Schematy charakteryzują się następującymi rzędami i rozpiętościami: ρ( S ) = 3, δ( S1) ρ S ) = 2, δ( S ) 1 = ( 1 1 = Potomkowie po krzyżowaniu jednopunktowym v v [ ] T 1 ' = 0 [ ] T 2 ' = 1 8 1
138 Teoria schematów - efekt krzyżowania Prawdopodobieństwa destrukcji i przeżycia schematów S 1 i S 2 wynoszą odpowiednio p p 8 ) =, 10 pd ( S 1 ) =, 10 pd ( S 2 ) = 10 9 ) 10 d ( S1 1 = d ( S2 2 = Wniosek: Większa szansa przeżycia schematów o małej rozpiętości
139 Teoria schematów - efekt krzyżowania Jeśli operacja krzyżowania jest wykonywana z prawdopodobieństwem krzyżowania 0<p c <1 Wówczas prawdopodobieństwo zniszczenia oraz zachowania schematu S ma postać 1 ) ( ) ( δ m S p S p c d 1 ) ( ) ( δ m S p S p c d
140 Teoria schematów - efekt mutacji Schemat S przeżywa mutację jeśli ustalone pozycje (geny o wartościach 0 lub 1) zostają zachowane Gen mutowany z prawdopodobieństwem mutacji p m Prawdopodobieństwo przeżycia schematu S gdzie ρ(s) ps m ( S) = (1 p m - rząd schematu S ) ρ( S)
141 Teoria schematów - efekt mutacji Aproksymacja prawdopodobieństwo przeżycia schematu S (dla małych wartości p m <<1) ps m ( S) 1 ρ( S) p m Prawdopodobieństwa przeżycia z przykładu 10 ps m ( S 1 ) 1 3 p m ps m ( S 2 ) 1 2 p m Wniosek: Schematy o małym rzędzie przetrwają mutację
142 Teoria schematów - twierdzenie Twierdzenie schematów ξ t + 1( S) ξ t ( S) F f t avg ( S) ( t) 1 p c δ( S) ρ( S) p m 1 Schematy o: małej rozpiętości niskiego rzędu z wyższym stopniem przystosowania uzyskują wykładniczo rosnącą liczbę reprezentantów w następnych pokoleniach m
143 Teoria schematów - twierdzenie Hipoteza: AG poszukuje rozwiązania optymalnego poprzez zachowywanie krótkich schematów niskiego rzędu o stopniu przystosowania wyższym od średniego Schematy takie nazwane są blokami budującymi Dowód oparty głównie na wynikach empirycznych uzyskanych dla szerokiego spektrum zastosowań GA
144 Teoria schematów - twierdzenie Przetwarzanie w toku genetycznej ewolucji m m co najmniej 2 i N 2 co najwyżej schematów (populacja o N osobnikach kodowanych binarnie o długości chromosomu m) Nie wszystkie schematy mają szansę na przetrwanie: operacja krzyżowania niszczy schematy o dużej rozpiętości mutacja niszczy schematy o wyższym rzędzie
145 Teoria schematów - twierdzenie Przy względnie długich chromosomach (2 m > N 2 ) efektywna liczba schematów przetwarzana w każdym pokoleniu przez AG wynosi N 3 Własność AG: ukryta równoległość (ang. implicit parallelism) AG działając na zbiorze N osobników dokonuje przetwarzania schematów N 3
146 Symulacja GA Maksymalizacja odwróconej funkcji Shekhel a - wilcze doły max ( 1, 2), = 1 ( 1 ) ( 2 ) g x x = + x x j j + x aij + x aij 1 A = K K
147 Symulacja GA W zastosowanym algorytmie genetycznym założono następujące parametry: dziedzina poszukiwań x,2 [ 50 osobników w populacji 24 bity na każdy parametr 1 dwupunktowe krzyżowanie z binarna mutacja z = 0.01 p m liczba pokoleń równa 50 65, p c 65] = 0.8



148 Symulacja GA Odwrócona funkcja Shekhel a - wilcze doły
149 Symulacja GA Przebiegi algorytmu genetycznego dla: a) maksymalnej i średniej wartości funkcji zysku b) poszukiwanych parametrów. 0 g(x 1,x 2 ) g max pokolenie pokolenie g(x 1,x 2 ) g avg pokolenie x x (a) (b)
150 Symulacja GA Maksymalizacja odwróconej funkcji skokowej 2 1, x 2) = 1 x 2 5 ( x ) + 0. max g( x + 1, x 2 ( ) 2 x [ ] [ ] Globalne maksimum w punkcie o wartości funkcji g( 0,0) = 0 x * * 1 x2 = 0 0
151 Symulacja GA W zastosowanym algorytmie genetycznym założono następujące parametry: dziedzina poszukiwań x1,2 [ 100, 100] 60 osobników w populacji 24 bity na każdy parametr dwupunktowe krzyżowanie z binarna mutacja z p m = 0.01 liczba pokoleń równa 50 p c = 0.8

152 Symulacja GA Odwrócona funkcja skokowa
153 Symulacja GA Przebiegi algorytmu genetycznego dla: a) maksymalnej i średniej wartości funkcji zysku b) poszukiwanych parametrów. 0 g(x 1,x 2 ) g max pokolenie pokolenie g(x 1,x 2 ) g avg pokolenie x x (a) (b)
154 Symulacja GA Maksymalizacja odwróconej funkcji Schaffer a max g ( x 1, x 2 ) = ( x 1 + x 2 ) 1 x 2 + x 1, x 2 [ sin ( 50( x + ) ) 1 ] Globalne maksimum w punkcie o wartości funkcji g( 0,0) = 0 [ * *] x x [ 0 0] 1 2 =



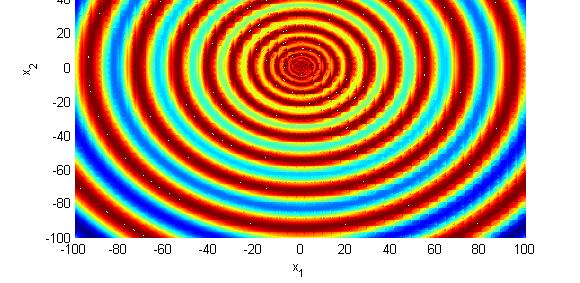
155 Symulacja GA Odwrócona funkcja Schaffer a
156 Symulacja GA Przebiegi algorytmu genetycznego dla: a) maksymalnej i średniej wartości funkcji zysku b) poszukiwanych parametrów. 0 g(x 1,x 2 ) g max pokolenie 0 pokolenie g(x 1,x 2 ) g avg pokolenie x x (a) (b)
157 Symulacja GA Maksymalizacja odwróconej funkcji Easom a max x 1, x 2 g( x 1, x 2 ) 1 2 ( 2 2) ( x π) ( π) = cos( x )cos( x )exp x 1 2 Globalne maksimum w punkcie o wartości funkcji g( π, π) = 1 [ * *] x 1 x = [ π π] 2


158 Symulacja GA Odwrócona funkcja Easom a
159 Symulacja GA Przebiegi algorytmu genetycznego dla: a) maksymalnej i średniej wartości funkcji zysku b) poszukiwanych parametrów. 1 g max g(x 1,x 2 ) pokolenie 1 g avg pokolenie g(x 1,x 2 ) pokolenie x x (a) (b)
160 Symulacja GA Maksymalizacja funkcji wielomodalnej 5 max g( x1, x2) = x 1, x 2 j = 1 gdzie ) ( x a ) + ( x a ) c j a11 = 4, a12 = 1, a13 = 8, a14 = 6, a15 = 7 a21 = 4, a22 = 1, a23 = 8, a24 = 6, a25 = j j c1 = 0.1, c2 = 0.2, c3 = 0.2, c4 = 0.4, c5 = 0.6
161 Symulacja GA Rozważana funkcja kryterialna posiada jedno globalne oraz cztery lokalne maksima: g( 4,4) g( 8,8) g( 1,1) g( 6,6) g( 7,3)
")
162 Symulacja GA Odwrócona funkcja Shekel a: a) wykres b) zrzutowana na płaszczyznę poszukiwanych parametrów x x 1 (a) (b)
163 Symulacja dla klasycznego GA Przebiegi algorytmu genetycznego dla: a) maksymalnej i średniej wartości funkcji zysku b) populacja osobników w 100 pokoleniu na tle płaszczyzny poszukiwanych parametrów g(x 1,x 2 ) 10 5 g max pokolenie 10 g avg x g(x 1,x 2 ) pokolenie (a) x 1 (b)
164 Symulacja dla GA z niszowaniem Przebiegi algorytmu genetycznego z zastosowanym niszowaniem dla: a) maksymalnej i średniej wartości funkcji zysku b) populacja osobników w 100 pokoleniu na tle płaszczyzny poszukiwanych parametrów 10 g(x 1,x 2 ) 10 5 g max gatunek gatunek pokolenie x g(x 1,x 2 ) pokolenie (a) g avg gatunek gatunek 1 gatunek x 1 (b)
165 Symulacja dla klasycznego GA (mały rozmiar populacji) Przebiegi algorytmu genetycznego dla: a) maksymalnej i średniej wartości funkcji zysku b) populacja osobników w 100 pokoleniu na tle płaszczyzny poszukiwanych parametrów 6 10 g(x 1,x 2 ) 5 4 g max pokolenie 6 x g(x 1,x 2 ) 4 2 g avg pokolenie (a) x 1 (b)
166 Symulacja dla GA z niszowaniem (mały rozmiar populacji) Przebiegi algorytmu genetycznego z zastosowanym niszowaniem dla: a) maksymalnej i średniej wartości funkcji zysku b) populacja osobników w 100 pokoleniu na tle płaszczyzny poszukiwanych parametrów g max 9 gatunek g(x 1,x 2 ) gatunek pokolenie x gatunek g(x 1,x 2 ) g avg gatunek gatunek pokolenie (a) x 1 (b)
167 Strategie Ewolucyjne (SE) SE powstały w wyniku badań optymalizacyjnych urządzeń mechanicznych metodą permutowania pewnego początkowego rozwiązania Rodzaje strategii ewolucyjnych: (1+1) (µ+λ) (µ, λ)
168 Strategia ewolucyjna (1+1) Przetwarzany jeden osobnik haploidalny z kodowaniem multiallelicznym Osobnik jest modyfikowany poprzez mutację fentypową z rozkładem normalnym Rodzic zastępowany potomkiem, jeżeli przystosowanie potomka jest lepsze Mutacja wykorzystuje mechanizm adaptacji jej zasięgu zwany regułą 1/5 sukcesów
169 Strategia ewolucyjna (1+1) Reguła 1/5 sukcesów: zwiększa zasięg mutacji, jeżeli pewna założona liczba sukcesów mutacji jest większa niż 1/5 ogólnej liczby wykonywanych mutacji w przeciwnym przypadku zasięg mutacji jest odpowiednio zmniejszony SE (1+1) podobna do algorytmu symulowanego wyżarzania, ale nie dopuszcza się akceptacji punktu gorszego
170 Schemat SE (1+1) 1. wygeneruj losowo z rozkładem równomiernym początkowego osobnika T [ ] n v = x = x1 x2 K x n R 2. wyznacz stopień przystosowania f (x) R 3. jeżeli liczba iteracji SE (1+1) osiągnęła wartość założoną to zakończ poszukiwania i wyznacz najlepiej przystosowanego osobnika; w przeciwnym przypadku przejdź do kroku 4
171 Schemat SE (1+1) 4. utwórz potomka poprzez operację mutacji każdego genu osobnika x ' j = x + r j ε gdzie x ' j zmodyfikowany j-ty gen osobnika ε liczba losowa wg rozkładu normalnego r zasięg mutacji (początkowa wartość r=1)
172 Schemat SE (1+1) 5. wyznacz stopień przystosowania potomka f (x') R f ( x') > f ( x) 6. jeżeli to zastąp rodzica potomkiem i zwiększ o jeden licznik sukcesów mutacji s (początkowa wartość s=0) s = s +1
173 Schemat SE (1+1) 7. zastosuj regułę 1/5 sukcesu s >1/5 jeżeli wszystkich sukcesów to zwiększ zasięg mutacji wg wzoru 1 r = (0.82) ε s =1/5 jeżeli wszystkich sukcesów to zasięg mutacji pozostaw bez zmian s <1/5 jeżeli wszystkich sukcesów to zmniejsz zasięg mutacji wg wzoru r = ε 8. powróć do kroku 3
174 Podsumowanie SE (1+1) Wartość 1/5 oraz pozostałe współczynniki modyfikacji zasięgu mutacji zostały wyznaczone eksperymentalnie Mimo zastosowanego mechanizmu adaptacji mutacji strategię ewolucyjną cechuje niewielka odporność na minima lokalne
175 Strategia Ewolucyjna (µ+λ) Uogólnienie SE (1+1) przetwarzana populacje µ osobników o strukturze diploidalnej gdzie i v [ T v ], i = 1,2, N di = 1 v K, i 2 = i [ ] T n x x x R v 1 = K i 1 i 2 i n i [ ] T n σ σ σ R v 2 = K 1 i 2 i n i
176 Strategia Ewolucyjna (µ+λ) Chromosom v = x jest wektorem 1i poszukiwanych parametrów v 2i Chromosom jest wektorem odchyleń standardowych dla każdego parametru (wykorzystywany przy operacji mutacji) i Selekcja osobników polega na wylosowaniu (ze zwracaniem) z rozkładem równomiernym λ osobników spośród µ osobników
177 Strategia Ewolucyjna (µ+λ) Rodzice poddawani są procesowi krzyżowania arytmetycznego oraz mutacji Potomkowie porównywani są ze z rodzicami Do dalszego cyklu strategii wybieranych jest tylko µ najlepszych osobników (elityzm)
178 Schemat SE (µ+ λ) 1. wygeneruj losowo z rozkładem równomiernym populację µ osobników V [ v v K v ] µ d = d1 d 2 dµ 2. wyznacz przystosowanie osobników f x ( i ) R 3. jeżeli liczba iteracji SE (µ+λ) osiągnęła wartość założoną to zakończ poszukiwania i wyznacz najlepiej przystosowanego osobnika; w przeciwnym przypadku przejdź do kroku 4
179 Schemat SE (µ+ λ) 4. utwórz pulę rodzicielską poprzez λ-losowań ze zwracaniem osobnika z populacji µ-osobników 5. utwórz z puli rodzicielskiej populację potomków poprzez: λ V d przeprowadzenie krzyżowania arytmetycznego dla wylosowanych par rodziców z populacji λ V d
180 Schemat SE (µ+ λ) przeprowadzenie dla v di mutacji v ' 2 ' i i gdzie = [ ( τζ+υζ τζ+υζ ] T n n σ e 1) ( ) K σ e R 1 i ( ) 2 1 τ = n υ = ( 2n ) 1 [ ' ' ] T n x + σ ε x + σ ε R v 1 = K εi 1 i 1 i 1 n i, ζ i, ζ - wartości wygenerowane losowo z rozkładem normalnym n i n i n
181 Schemat SE (µ+ λ) 6. wyznacz stopień przystosowania potomków f ( x i ') R 7. utwórz nową populacje µ najlepszych osobników spośród populacji λ V d µ V d 8. powróć do kroku 3
182 Podsumowanie SE (µ+ λ) Mutacja przeprowadzana jest w pierwszej kolejności dla chromosomu z odchyleniami standardowymi Zmodyfikowany wektor z odchyleniami standardowymi jest wykorzystywany przy mutacji chromosomu reprezentującego poszukiwane parametry zadania optymalizacji Mechanizm mutacji ma na celu zapewnienie samoczynnej adaptacji zasięgu mutacji
183 Podsumowanie SE (µ+ λ) Brak arbitralności przy doborze poszczególnych odchyleń standardowych (dobierane losowo) Efekt adaptacji jest wynikiem zachodzącego procesu selekcji wybierającego do dalszej ewolucji lepiej przystosowane osobniki SE (µ+ λ) pomimo większej odporności od (1+1) również narażona jest na zjawisko przedwczesnej zbieżności algorytmu
184 Strategia Ewolucyjna (µ, λ) Udoskonalona wersja strategii (µ+λ) Mniej narażona na zjawisko przedwczesnej zbieżności poprzez zrezygnowanie z mechanizmu tworzenia elity W odróżnieniu od (µ+λ) utworzenie nowej populacji µ najlepszych osobników odbywa się tylko dla populacji potomków V λ d
185 Podsumowanie Strategii Ewolucyjnych W strategiach ewolucyjnych (µ+λ) i (µ, λ) parametry algorytmu zwykle przyjmują następujące wartości: µ d V złożona z µ [50,100] osobników λ V 6 d zawierająca λ = 4 razy więcej osobników niż populacja µ V d
186 Programowanie ewolucyjne (PE) Problem stworzenia gramatyki nieznanego języka na podstawie zestawu symboli i wyrażeń syntaktycznie poprawnych Gramatyka modelowana za pomocą poszukiwanych w sposób ewolucyjny grafów złożonych ze stanów i funkcji przejść Proces tworzenia nowego grafu poprzez mutację jego struktury (dodanie/usunięcie stanu, zmianę funkcji wyjść lub przejść)
187 Programowanie ewolucyjne W późniejszych badaniach programowanie ewolucyjne przeszło w stronę strategii ewolucyjnych jako kolejnych metod optymalizacji numerycznej Podobnie jak w SE (µ+λ) i (µ, λ) osobnik w PE jest reprezentowany kodowaniem multiallelicznym (zminennopozycyjnym) W odróżnieniu od SE, każdy osobnik generuje dokładnie jednego potomka
188 Programowanie ewolucyjne Tworzenie potomków przeprowadzane wg mutacji fenotypowej w trzech wariantach: bez adaptacji adaptacja ze śledzeniem wartości funkcji przystosowania samoczynna adaptacja Struktura osobnika haploidalna lub diploidalna w zależności od stosowanej mutacji
189 PE mutacja bez adaptacji Modyfikacja osobnika haploidalnego (z pojedynczym chromosomem) wg wzoru ' i v [ T n x + σ ε x + σ ε ] R ' = x i = K i n i n n gdzie ε - wektor zmiennych losowych o n-wymiarowym nieskolerowanym rozkładzie normalnym σ - zasięg mutacji
190 PE mutacja adaptacyjna śledząca zmiany przystosowania Generowanie potomka haploidalnego v gdzie ' i ' i = x = x + ε β( M f ( x )) i i + γ R ε - wektor zmiennych losowych o n-wymiarowym nieskolerowanym rozkładzie normalnym γ, β - parametry PE (zwykle β=1, γ=0) M - oszacowanie wartości globalnego maksmimum n
191 PE mutacja z samoczynną adaptacją Stosowana dla osobników diploidalnych W pierwszej kolejności modyfikowany jest chromosom zawierający poszukiwane parametry zadania optymalizacji W drugi kroku zmieniany jest chromosom składający się z genów będącymi wartościami odchyleń standardowych poszczególnych parametrów chromosomu
192 PE mutacja z samoczynną adaptacją Potomek uzyskiwany jest wg wzorów gdzie v ' 2 j i ε j σ j ζ v [ x + σ ε x + σ ε ] T ' 1 = K n n i ' i 2 v2 = min{ σmin, ( σ1 + ζε1 )} j i - j-ty gen chromosomu - zmienna losowa o rozkładzie normalnym - j-te odchylenie standardowe - intensywność zmian σ n i 2 ' v 2i j
Algorytmy genetyczne. Paweł Cieśla. 8 stycznia 2009
 Algorytmy genetyczne Paweł Cieśla 8 stycznia 2009 Genetyka - nauka o dziedziczeniu cech pomiędzy pokoleniami. Geny są czynnikami, które decydują o wyglądzie, zachowaniu, rozmnażaniu każdego żywego organizmu.
Algorytmy genetyczne Paweł Cieśla 8 stycznia 2009 Genetyka - nauka o dziedziczeniu cech pomiędzy pokoleniami. Geny są czynnikami, które decydują o wyglądzie, zachowaniu, rozmnażaniu każdego żywego organizmu.
PLAN WYKŁADU OPTYMALIZACJA GLOBALNA OPERATOR KRZYŻOWANIA ETAPY KRZYŻOWANIA
 PLAN WYKŁADU Operator krzyżowania Operator mutacji Operator inwersji Sukcesja Przykłady symulacji AG Kodowanie - rodzaje OPTYMALIZACJA GLOBALNA Wykład 3 dr inż. Agnieszka Bołtuć OPERATOR KRZYŻOWANIA Wymiana
PLAN WYKŁADU Operator krzyżowania Operator mutacji Operator inwersji Sukcesja Przykłady symulacji AG Kodowanie - rodzaje OPTYMALIZACJA GLOBALNA Wykład 3 dr inż. Agnieszka Bołtuć OPERATOR KRZYŻOWANIA Wymiana
Algorytmy genetyczne. Dariusz Banasiak. Katedra Informatyki Technicznej Wydział Elektroniki
 Dariusz Banasiak Katedra Informatyki Technicznej Wydział Elektroniki Obliczenia ewolucyjne (EC evolutionary computing) lub algorytmy ewolucyjne (EA evolutionary algorithms) to ogólne określenia używane
Dariusz Banasiak Katedra Informatyki Technicznej Wydział Elektroniki Obliczenia ewolucyjne (EC evolutionary computing) lub algorytmy ewolucyjne (EA evolutionary algorithms) to ogólne określenia używane
Algorytm genetyczny (genetic algorithm)-
-") Optymalizacja W praktyce inżynierskiej często zachodzi potrzeba znalezienia parametrów, dla których system/urządzenie będzie działać w sposób optymalny. Klasyczne podejście do optymalizacji: sformułowanie
Optymalizacja W praktyce inżynierskiej często zachodzi potrzeba znalezienia parametrów, dla których system/urządzenie będzie działać w sposób optymalny. Klasyczne podejście do optymalizacji: sformułowanie
Algorytmy genetyczne
 Politechnika Łódzka Katedra Informatyki Stosowanej Algorytmy genetyczne Wykład 2 Przygotował i prowadzi: Dr inż. Piotr Urbanek Powtórzenie Pytania: Jaki mechanizm jest stosowany w naturze do takiego modyfikowania
Politechnika Łódzka Katedra Informatyki Stosowanej Algorytmy genetyczne Wykład 2 Przygotował i prowadzi: Dr inż. Piotr Urbanek Powtórzenie Pytania: Jaki mechanizm jest stosowany w naturze do takiego modyfikowania
Algorytmy genetyczne
 Politechnika Łódzka Katedra Informatyki Stosowanej Algorytmy genetyczne Wykład 2 Przygotował i prowadzi: Dr inż. Piotr Urbanek Powtórzenie Pytania: Jaki mechanizm jest stosowany w naturze do takiego modyfikowania
Politechnika Łódzka Katedra Informatyki Stosowanej Algorytmy genetyczne Wykład 2 Przygotował i prowadzi: Dr inż. Piotr Urbanek Powtórzenie Pytania: Jaki mechanizm jest stosowany w naturze do takiego modyfikowania
Algorytmy genetyczne w optymalizacji
 Algorytmy genetyczne w optymalizacji Literatura 1. David E. Goldberg, Algorytmy genetyczne i ich zastosowania, WNT, Warszawa 1998; 2. Zbigniew Michalewicz, Algorytmy genetyczne + struktury danych = programy
Algorytmy genetyczne w optymalizacji Literatura 1. David E. Goldberg, Algorytmy genetyczne i ich zastosowania, WNT, Warszawa 1998; 2. Zbigniew Michalewicz, Algorytmy genetyczne + struktury danych = programy
SCHEMAT ROZWIĄZANIA ZADANIA OPTYMALIZACJI PRZY POMOCY ALGORYTMU GENETYCZNEGO
 SCHEMAT ROZWIĄZANIA ZADANIA OPTYMALIZACJI PRZY POMOCY ALGORYTMU GENETYCZNEGO. Rzeczywistość (istniejąca lub projektowana).. Model fizyczny. 3. Model matematyczny (optymalizacyjny): a. Zmienne projektowania
SCHEMAT ROZWIĄZANIA ZADANIA OPTYMALIZACJI PRZY POMOCY ALGORYTMU GENETYCZNEGO. Rzeczywistość (istniejąca lub projektowana).. Model fizyczny. 3. Model matematyczny (optymalizacyjny): a. Zmienne projektowania
Algorytm Genetyczny. zastosowanie do procesów rozmieszczenia stacji raportujących w sieciach komórkowych
 Algorytm Genetyczny zastosowanie do procesów rozmieszczenia stacji raportujących w sieciach komórkowych Dlaczego Algorytmy Inspirowane Naturą? Rozwój nowych technologii: złożone problemy obliczeniowe w
Algorytm Genetyczny zastosowanie do procesów rozmieszczenia stacji raportujących w sieciach komórkowych Dlaczego Algorytmy Inspirowane Naturą? Rozwój nowych technologii: złożone problemy obliczeniowe w
Obliczenia ewolucyjne - plan wykładu
 Obliczenia ewolucyjne - plan wykładu Wprowadzenie Algorytmy genetyczne Programowanie genetyczne Programowanie ewolucyjne Strategie ewolucyjne Inne modele obliczeń ewolucyjnych Podsumowanie Ewolucja Ewolucja
Obliczenia ewolucyjne - plan wykładu Wprowadzenie Algorytmy genetyczne Programowanie genetyczne Programowanie ewolucyjne Strategie ewolucyjne Inne modele obliczeń ewolucyjnych Podsumowanie Ewolucja Ewolucja
Metody Rozmyte i Algorytmy Ewolucyjne
 mgr inż. Wydział Matematyczno-Przyrodniczy Szkoła Nauk Ścisłych Uniwersytet Kardynała Stefana Wyszyńskiego Podstawowe operatory genetyczne Plan wykładu Przypomnienie 1 Przypomnienie Metody generacji liczb
mgr inż. Wydział Matematyczno-Przyrodniczy Szkoła Nauk Ścisłych Uniwersytet Kardynała Stefana Wyszyńskiego Podstawowe operatory genetyczne Plan wykładu Przypomnienie 1 Przypomnienie Metody generacji liczb
Algorytmy genetyczne. Materiały do laboratorium PSI. Studia niestacjonarne
 Algorytmy genetyczne Materiały do laboratorium PSI Studia niestacjonarne Podstawowy algorytm genetyczny (PAG) Schemat blokowy algorytmu genetycznego Znaczenia, pochodzących z biologii i genetyki, pojęć
Algorytmy genetyczne Materiały do laboratorium PSI Studia niestacjonarne Podstawowy algorytm genetyczny (PAG) Schemat blokowy algorytmu genetycznego Znaczenia, pochodzących z biologii i genetyki, pojęć
Zadanie 5 - Algorytmy genetyczne (optymalizacja)
") Zadanie 5 - Algorytmy genetyczne (optymalizacja) Marcin Pietrzykowski mpietrzykowski@wi.zut.edu.pl wersja 1.0 1 Cel Celem zadania jest zapoznanie się z Algorytmami Genetycznymi w celu rozwiązywanie zadania
Zadanie 5 - Algorytmy genetyczne (optymalizacja) Marcin Pietrzykowski mpietrzykowski@wi.zut.edu.pl wersja 1.0 1 Cel Celem zadania jest zapoznanie się z Algorytmami Genetycznymi w celu rozwiązywanie zadania
Algorytmy genetyczne. Materiały do laboratorium PSI. Studia stacjonarne i niestacjonarne
 Algorytmy genetyczne Materiały do laboratorium PSI Studia stacjonarne i niestacjonarne Podstawowy algorytm genetyczny (PAG) Schemat blokowy algorytmu genetycznego Znaczenia, pochodzących z biologii i genetyki,
Algorytmy genetyczne Materiały do laboratorium PSI Studia stacjonarne i niestacjonarne Podstawowy algorytm genetyczny (PAG) Schemat blokowy algorytmu genetycznego Znaczenia, pochodzących z biologii i genetyki,
Równoważność algorytmów optymalizacji
 Równoważność algorytmów optymalizacji Reguła nie ma nic za darmo (ang. no free lunch theory): efektywność różnych typowych algorytmów szukania uśredniona po wszystkich możliwych problemach optymalizacyjnych
Równoważność algorytmów optymalizacji Reguła nie ma nic za darmo (ang. no free lunch theory): efektywność różnych typowych algorytmów szukania uśredniona po wszystkich możliwych problemach optymalizacyjnych
Inspiracje soft computing. Soft computing. Terminy genetyczne i ich odpowiedniki w algorytmach genetycznych. Elementarny algorytm genetyczny
 Soft computing Soft computing tym róŝni się od klasycznych obliczeń (hard computing), Ŝe jest odporny na brak precyzji i niepewność danych wejściowych. Obliczenia soft computing mają inspiracje ze świata
Soft computing Soft computing tym róŝni się od klasycznych obliczeń (hard computing), Ŝe jest odporny na brak precyzji i niepewność danych wejściowych. Obliczenia soft computing mają inspiracje ze świata
Algorytmy ewolucyjne. wprowadzenie
 Algorytmy ewolucyjne wprowadzenie Gracjan Wilczewski, www.mat.uni.torun.pl/~gracjan Toruń, 2005 Historia Podstawowy algorytm genetyczny został wprowadzony przez Johna Hollanda (Uniwersytet Michigan) i
Algorytmy ewolucyjne wprowadzenie Gracjan Wilczewski, www.mat.uni.torun.pl/~gracjan Toruń, 2005 Historia Podstawowy algorytm genetyczny został wprowadzony przez Johna Hollanda (Uniwersytet Michigan) i
Algorytmy ewolucyjne - algorytmy genetyczne. I. Karcz-Dulęba
 Algorytmy ewolucyjne - algorytmy genetyczne I. Karcz-Dulęba Algorytmy klasyczne a algorytmy ewolucyjne Przeszukiwanie przestrzeni przez jeden punkt bazowy Przeszukiwanie przestrzeni przez zbiór punktów
Algorytmy ewolucyjne - algorytmy genetyczne I. Karcz-Dulęba Algorytmy klasyczne a algorytmy ewolucyjne Przeszukiwanie przestrzeni przez jeden punkt bazowy Przeszukiwanie przestrzeni przez zbiór punktów
6. Klasyczny algorytm genetyczny. 1
 6. Klasyczny algorytm genetyczny. 1 Idea algorytmu genetycznego została zaczerpnięta z nauk przyrodniczych opisujących zjawiska doboru naturalnego i dziedziczenia. Mechanizmy te polegają na przetrwaniu
6. Klasyczny algorytm genetyczny. 1 Idea algorytmu genetycznego została zaczerpnięta z nauk przyrodniczych opisujących zjawiska doboru naturalnego i dziedziczenia. Mechanizmy te polegają na przetrwaniu
Algorytmy ewolucyjne NAZEWNICTWO
 Algorytmy ewolucyjne http://zajecia.jakubw.pl/nai NAZEWNICTWO Algorytmy ewolucyjne nazwa ogólna, obejmująca metody szczegółowe, jak np.: algorytmy genetyczne programowanie genetyczne strategie ewolucyjne
Algorytmy ewolucyjne http://zajecia.jakubw.pl/nai NAZEWNICTWO Algorytmy ewolucyjne nazwa ogólna, obejmująca metody szczegółowe, jak np.: algorytmy genetyczne programowanie genetyczne strategie ewolucyjne
Algorytmy genetyczne
 Algorytmy genetyczne Motto: Zamiast pracowicie poszukiwać najlepszego rozwiązania problemu informatycznego lepiej pozwolić, żeby komputer sam sobie to rozwiązanie wyhodował! Algorytmy genetyczne służą
Algorytmy genetyczne Motto: Zamiast pracowicie poszukiwać najlepszego rozwiązania problemu informatycznego lepiej pozwolić, żeby komputer sam sobie to rozwiązanie wyhodował! Algorytmy genetyczne służą
Algorytmy genetyczne (AG)
") Algorytmy genetyczne (AG) 1. Wprowadzenie do AG a) ewolucja darwinowska b) podstawowe definicje c) operatory genetyczne d) konstruowanie AG e) standardowy AG f) przykład rozwiązania g) naprawdę bardzo,
Algorytmy genetyczne (AG) 1. Wprowadzenie do AG a) ewolucja darwinowska b) podstawowe definicje c) operatory genetyczne d) konstruowanie AG e) standardowy AG f) przykład rozwiązania g) naprawdę bardzo,
Dobór parametrów algorytmu ewolucyjnego
 Dobór parametrów algorytmu ewolucyjnego 1 2 Wstęp Algorytm ewolucyjny posiada wiele parametrów. Przykładowo dla algorytmu genetycznego są to: prawdopodobieństwa stosowania operatorów mutacji i krzyżowania.
Dobór parametrów algorytmu ewolucyjnego 1 2 Wstęp Algorytm ewolucyjny posiada wiele parametrów. Przykładowo dla algorytmu genetycznego są to: prawdopodobieństwa stosowania operatorów mutacji i krzyżowania.
PLAN WYKŁADU OPTYMALIZACJA GLOBALNA HISTORIA NA CZYM BAZUJĄ AG
 PLAN WYKŁADU OPTYMALIZACJA GLOBALNA Wykład 2 dr inż. Agnieszka Bołtuć Historia Zadania Co odróżnia od klasycznych algorytmów Nazewnictwo Etapy Kodowanie, inicjalizacja, transformacja funkcji celu Selekcja
PLAN WYKŁADU OPTYMALIZACJA GLOBALNA Wykład 2 dr inż. Agnieszka Bołtuć Historia Zadania Co odróżnia od klasycznych algorytmów Nazewnictwo Etapy Kodowanie, inicjalizacja, transformacja funkcji celu Selekcja
Strategie ewolucyjne. Gnypowicz Damian Staniszczak Łukasz Woźniak Marek
 Strategie ewolucyjne Gnypowicz Damian Staniszczak Łukasz Woźniak Marek Strategie ewolucyjne, a algorytmy genetyczne Podobieństwa: Oba działają na populacjach rozwiązań Korzystają z zasad selecji i przetwarzania
Strategie ewolucyjne Gnypowicz Damian Staniszczak Łukasz Woźniak Marek Strategie ewolucyjne, a algorytmy genetyczne Podobieństwa: Oba działają na populacjach rozwiązań Korzystają z zasad selecji i przetwarzania
METODY HEURYSTYCZNE wykład 3
 SCHEMAT DZIAŁANIA AG: METODY HEURYSTYCZNE wykład 3 procedure Algorytm_genetyczny t:=0 wybierz populację początkową P(t) oceń P(t) while (not warunek_zakończenia) do t:=t+ wybierz P(t) z P(t-) (selekcja)
SCHEMAT DZIAŁANIA AG: METODY HEURYSTYCZNE wykład 3 procedure Algorytm_genetyczny t:=0 wybierz populację początkową P(t) oceń P(t) while (not warunek_zakończenia) do t:=t+ wybierz P(t) z P(t-) (selekcja)
Obliczenia Naturalne - Algorytmy genetyczne
 Literatura Obliczenia Naturalne - Algorytmy genetyczne Paweł Paduch Politechnika Świętokrzyska 20 marca 2014 Paweł Paduch Obliczenia Naturalne - Algorytmy genetyczne 1 z 45 Plan wykładu Literatura 1 Literatura
Literatura Obliczenia Naturalne - Algorytmy genetyczne Paweł Paduch Politechnika Świętokrzyska 20 marca 2014 Paweł Paduch Obliczenia Naturalne - Algorytmy genetyczne 1 z 45 Plan wykładu Literatura 1 Literatura
Algorytmy ewolucyjne. Łukasz Przybyłek Studenckie Koło Naukowe BRAINS
 Algorytmy ewolucyjne Łukasz Przybyłek Studenckie Koło Naukowe BRAINS 1 Wprowadzenie Algorytmy ewolucyjne ogólne algorytmy optymalizacji operujące na populacji rozwiązań, inspirowane biologicznymi zjawiskami,
Algorytmy ewolucyjne Łukasz Przybyłek Studenckie Koło Naukowe BRAINS 1 Wprowadzenie Algorytmy ewolucyjne ogólne algorytmy optymalizacji operujące na populacji rozwiązań, inspirowane biologicznymi zjawiskami,
METODY HEURYSTYCZNE wykład 3
 METODY HEURYSTYCZNE wykład 3 1 Przykład: Znaleźć max { f (x)=x 2 } dla wartości całkowitych x z zakresu 0-31. Populacja w chwili t: P(t)= {x t 1,...x t n} Założenia: - łańcuchy 5-bitowe (x=0,1,...,31);
METODY HEURYSTYCZNE wykład 3 1 Przykład: Znaleźć max { f (x)=x 2 } dla wartości całkowitych x z zakresu 0-31. Populacja w chwili t: P(t)= {x t 1,...x t n} Założenia: - łańcuchy 5-bitowe (x=0,1,...,31);
Algorytmy stochastyczne, wykład 01 Podstawowy algorytm genetyczny
 Algorytmy stochastyczne, wykład 01 J. Piersa Wydział Matematyki i Informatyki, Uniwersytet Mikołaja Kopernika 2014-02-21 In memoriam prof. dr hab. Tomasz Schreiber (1975-2010) 1 2 3 Różne Orientacyjny
Algorytmy stochastyczne, wykład 01 J. Piersa Wydział Matematyki i Informatyki, Uniwersytet Mikołaja Kopernika 2014-02-21 In memoriam prof. dr hab. Tomasz Schreiber (1975-2010) 1 2 3 Różne Orientacyjny
Algorytmy stochastyczne, wykład 02 Algorytmy genetyczne
 Algorytmy stochastyczne, wykład 02 Algorytmy genetyczne J. Piersa Wydział Matematyki i Informatyki, Uniwersytet Mikołaja Kopernika 2014-02-27 1 Mutacje algorytmu genetycznego 2 Dziedzina niewypukła abstrakcyjna
Algorytmy stochastyczne, wykład 02 Algorytmy genetyczne J. Piersa Wydział Matematyki i Informatyki, Uniwersytet Mikołaja Kopernika 2014-02-27 1 Mutacje algorytmu genetycznego 2 Dziedzina niewypukła abstrakcyjna
ALGORYTMY GENETYCZNE ćwiczenia
 ćwiczenia Wykorzystaj algorytmy genetyczne do wyznaczenia minimum globalnego funkcji testowej: 1. Wylosuj dwuwymiarową tablicę 100x2 liczb 8-bitowych z zakresu [-100; +100] reprezentujących inicjalną populację
ćwiczenia Wykorzystaj algorytmy genetyczne do wyznaczenia minimum globalnego funkcji testowej: 1. Wylosuj dwuwymiarową tablicę 100x2 liczb 8-bitowych z zakresu [-100; +100] reprezentujących inicjalną populację
ALGORYTMY EWOLUCYJNE W OPTYMALIZACJI JEDNOKRYTERIALNEJ
 ALGORYTMY EWOLUCYJNE W OPTYMALIZACJI JEDNOKRYTERIALNEJ Zalety: nie wprowadzają żadnych ograniczeń na sformułowanie problemu optymalizacyjnego. Funkcja celu może być wielowartościowa i nieciągła, obszar
ALGORYTMY EWOLUCYJNE W OPTYMALIZACJI JEDNOKRYTERIALNEJ Zalety: nie wprowadzają żadnych ograniczeń na sformułowanie problemu optymalizacyjnego. Funkcja celu może być wielowartościowa i nieciągła, obszar
Modyfikacje i ulepszenia standardowego algorytmu genetycznego
 Modyfikacje i ulepszenia standardowego algorytmu genetycznego 1 2 Przypomnienie: pseudokod SGA t=0; initialize(p 0 ); while(!termination_condition(p t )) { evaluate(p t ); T t =selection(p t ); O t =crossover(t
Modyfikacje i ulepszenia standardowego algorytmu genetycznego 1 2 Przypomnienie: pseudokod SGA t=0; initialize(p 0 ); while(!termination_condition(p t )) { evaluate(p t ); T t =selection(p t ); O t =crossover(t
LABORATORIUM 4: Algorytmy ewolucyjne cz. 2 wpływ operatorów krzyżowania i mutacji na skuteczność poszukiwań AE
 Instytut Mechaniki i Inżynierii Obliczeniowej Wydział Mechaniczny Technologiczny, Politechnika Śląska www.imio.polsl.pl METODY HEURYSTYCZNE LABORATORIUM 4: Algorytmy ewolucyjne cz. 2 wpływ operatorów krzyżowania
Instytut Mechaniki i Inżynierii Obliczeniowej Wydział Mechaniczny Technologiczny, Politechnika Śląska www.imio.polsl.pl METODY HEURYSTYCZNE LABORATORIUM 4: Algorytmy ewolucyjne cz. 2 wpływ operatorów krzyżowania
Generowanie i optymalizacja harmonogramu za pomoca
 Generowanie i optymalizacja harmonogramu za pomoca na przykładzie generatora planu zajęć Matematyka Stosowana i Informatyka Stosowana Wydział Fizyki Technicznej i Matematyki Stosowanej Politechnika Gdańska
Generowanie i optymalizacja harmonogramu za pomoca na przykładzie generatora planu zajęć Matematyka Stosowana i Informatyka Stosowana Wydział Fizyki Technicznej i Matematyki Stosowanej Politechnika Gdańska
Algorytmy genetyczne
 9 listopada 2010 y ewolucyjne - zbiór metod optymalizacji inspirowanych analogiami biologicznymi (ewolucja naturalna). Pojęcia odwzorowujące naturalne zjawiska: Osobnik Populacja Genotyp Fenotyp Gen Chromosom
9 listopada 2010 y ewolucyjne - zbiór metod optymalizacji inspirowanych analogiami biologicznymi (ewolucja naturalna). Pojęcia odwzorowujące naturalne zjawiska: Osobnik Populacja Genotyp Fenotyp Gen Chromosom
Algorytmy ewolucyjne `
 Algorytmy ewolucyjne ` Wstęp Czym są algorytmy ewolucyjne? Rodzaje algorytmów ewolucyjnych Algorytmy genetyczne Strategie ewolucyjne Programowanie genetyczne Zarys historyczny Alan Turing, 1950 Nils Aall
Algorytmy ewolucyjne ` Wstęp Czym są algorytmy ewolucyjne? Rodzaje algorytmów ewolucyjnych Algorytmy genetyczne Strategie ewolucyjne Programowanie genetyczne Zarys historyczny Alan Turing, 1950 Nils Aall
Algorytmy ewolucyjne Część II
 Wydział Zarządzania AGH Katedra Informatyki Stosowanej Algorytmy ewolucyjne Część II Metaheurystyki Treść wykładu Zastosowania Praktyczne aspekty GA Reprezentacja Funkcja dopasowania Zróżnicowanie dopasowania
Wydział Zarządzania AGH Katedra Informatyki Stosowanej Algorytmy ewolucyjne Część II Metaheurystyki Treść wykładu Zastosowania Praktyczne aspekty GA Reprezentacja Funkcja dopasowania Zróżnicowanie dopasowania
ALGORYTMY EWOLUCYJNE. INTELIGENTNE TECHNIKI KOMPUTEROWE wykład 011. Napór selekcyjny (selektywny nacisk. Superosobniki: SELEKCJA
 INTELIGENTNE TECHNIKI KOMPUTEROWE wykład 0 ALGORYTMY EWOLUCYJNE 2 Dla danego problemu można określić wiele sposobów kodowania i zdefiniować szereg operatorów (np. zadanie komiwojażera). AE to rozwinięcie
INTELIGENTNE TECHNIKI KOMPUTEROWE wykład 0 ALGORYTMY EWOLUCYJNE 2 Dla danego problemu można określić wiele sposobów kodowania i zdefiniować szereg operatorów (np. zadanie komiwojażera). AE to rozwinięcie
Techniki ewolucyjne - algorytm genetyczny i nie tylko
 Reprezentacja binarna W reprezentacji binarnej wybór populacji początkowej tworzymy poprzez tablice genotypów (rys.1.), dla osobników o zadanej przez użytkownika wielkości i danej długości genotypów wypełniamy
Reprezentacja binarna W reprezentacji binarnej wybór populacji początkowej tworzymy poprzez tablice genotypów (rys.1.), dla osobników o zadanej przez użytkownika wielkości i danej długości genotypów wypełniamy
Techniki optymalizacji
 Techniki optymalizacji Algorytm kolonii mrówek Idea Smuga feromonowa 1 Sztuczne mrówki w TSP Sztuczna mrówka agent, który porusza się z miasta do miasta Mrówki preferują miasta połączone łukami z dużą
Techniki optymalizacji Algorytm kolonii mrówek Idea Smuga feromonowa 1 Sztuczne mrówki w TSP Sztuczna mrówka agent, który porusza się z miasta do miasta Mrówki preferują miasta połączone łukami z dużą
Standardowy algorytm genetyczny
 Standardowy algorytm genetyczny 1 Szybki przegląd 2 Opracowany w USA w latach 70. Wcześni badacze: John H. Holland. Autor monografii Adaptation in Natural and Artificial Systems, wydanej w 1975 r., (teoria
Standardowy algorytm genetyczny 1 Szybki przegląd 2 Opracowany w USA w latach 70. Wcześni badacze: John H. Holland. Autor monografii Adaptation in Natural and Artificial Systems, wydanej w 1975 r., (teoria
Algorytmy ewolucyjne (2)
") Algorytmy ewolucyjne (2) zajecia.jakubw.pl/nai/ ALGORYTM GEETYCZY Cel: znaleźć makimum unkcji. Założenie: unkcja ta jet dodatnia. 1. Tworzymy oobników loowych. 2. Stoujemy operacje mutacji i krzyżowania
Algorytmy ewolucyjne (2) zajecia.jakubw.pl/nai/ ALGORYTM GEETYCZY Cel: znaleźć makimum unkcji. Założenie: unkcja ta jet dodatnia. 1. Tworzymy oobników loowych. 2. Stoujemy operacje mutacji i krzyżowania
SZTUCZNA INTELIGENCJA
 ZTUCZNA INTELIGENCJA WYKŁAD 6. ALGORYTMY GENETYCZNE - CHEMATY, METODY ELEKCJI Częstochowa 204 Dr hab. inż. Grzegorz Dude Wydział Eletryczny Politechnia Częstochowsa CHEMATY chemat zbór chromosomów o wspólnych
ZTUCZNA INTELIGENCJA WYKŁAD 6. ALGORYTMY GENETYCZNE - CHEMATY, METODY ELEKCJI Częstochowa 204 Dr hab. inż. Grzegorz Dude Wydział Eletryczny Politechnia Częstochowsa CHEMATY chemat zbór chromosomów o wspólnych
Katedra Informatyki Stosowanej. Algorytmy ewolucyjne. Inteligencja obliczeniowa
 Wydział Zarządzania AGH Katedra Informatyki Stosowanej Algorytmy ewolucyjne Treść wykładu Wprowadzenie Zasada działania Podział EA Cechy EA Algorytm genetyczny 2 EA - wprowadzenie Algorytmy ewolucyjne
Wydział Zarządzania AGH Katedra Informatyki Stosowanej Algorytmy ewolucyjne Treść wykładu Wprowadzenie Zasada działania Podział EA Cechy EA Algorytm genetyczny 2 EA - wprowadzenie Algorytmy ewolucyjne
Teoria algorytmów ewolucyjnych
 Teoria algorytmów ewolucyjnych 1 2 Dlaczego teoria Wynik analiza teoretycznej może pokazać jakie warunki należy spełnić, aby osiągnąć zbieżność do minimum globalnego. Np. sukcesja elitarystyczna. Może
Teoria algorytmów ewolucyjnych 1 2 Dlaczego teoria Wynik analiza teoretycznej może pokazać jakie warunki należy spełnić, aby osiągnąć zbieżność do minimum globalnego. Np. sukcesja elitarystyczna. Może
ALHE Z11 Jarosław Arabas wykład 11
 ALHE Z11 Jarosław Arabas wykład 11 algorytm ewolucyjny inicjuj P 0 {x 1, x 2... x } t 0 while! stop for i 1: if a p c O t,i mutation crossover select P t, k else O t,i mutation select P t,1 P t 1 replacement
ALHE Z11 Jarosław Arabas wykład 11 algorytm ewolucyjny inicjuj P 0 {x 1, x 2... x } t 0 while! stop for i 1: if a p c O t,i mutation crossover select P t, k else O t,i mutation select P t,1 P t 1 replacement
ALGORYTMY GENETYCZNE
 ALGORYTMY GENETYCZNE Algorytmy Genetyczne I. Co to są algorytmy genetyczne? II. Podstawowe pojęcia algorytmów genetycznych III. Proste algorytmy genetyczne IV. Kodowanie osobników i operacje genetyczne.
ALGORYTMY GENETYCZNE Algorytmy Genetyczne I. Co to są algorytmy genetyczne? II. Podstawowe pojęcia algorytmów genetycznych III. Proste algorytmy genetyczne IV. Kodowanie osobników i operacje genetyczne.
Testy De Jonga. Problemy. 1 Optymalizacja dyskretna i ciągła
 Problemy 1 Optymalizacja dyskretna i ciągła Problemy 1 Optymalizacja dyskretna i ciągła 2 Środowisko pomiarowe De Jonga Problemy 1 Optymalizacja dyskretna i ciągła 2 Środowisko pomiarowe De Jonga 3 Ocena
Problemy 1 Optymalizacja dyskretna i ciągła Problemy 1 Optymalizacja dyskretna i ciągła 2 Środowisko pomiarowe De Jonga Problemy 1 Optymalizacja dyskretna i ciągła 2 Środowisko pomiarowe De Jonga 3 Ocena
PLAN WYKŁADU OPTYMALIZACJA GLOBALNA ZADANIE KOMIWOJAŻERA METODY ROZWIĄZYWANIA. Specyfika zadania komiwojażera Reprezentacje Operatory
 PLAN WYKŁADU Specyfika zadania komiwojażera Reprezentacje Operatory OPTYMALIZACJA GLOBALNA Wykład 5 dr inż. Agnieszka Bołtuć ZADANIE KOMIWOJAŻERA Koncepcja: komiwojażer musi odwiedzić każde miasto na swoim
PLAN WYKŁADU Specyfika zadania komiwojażera Reprezentacje Operatory OPTYMALIZACJA GLOBALNA Wykład 5 dr inż. Agnieszka Bołtuć ZADANIE KOMIWOJAŻERA Koncepcja: komiwojażer musi odwiedzić każde miasto na swoim
Strategie ewolucyjne (ang. evolution strategies)
") Strategie ewolucyjne (ang. evolution strategies) 1 2 Szybki przegląd Rozwijane w Niemczech w latach 60-70. Wcześni badacze: I. Rechenberg, H.-P. Schwefel (student Rechenberga). Typowe zastosowanie: Optymalizacja
Strategie ewolucyjne (ang. evolution strategies) 1 2 Szybki przegląd Rozwijane w Niemczech w latach 60-70. Wcześni badacze: I. Rechenberg, H.-P. Schwefel (student Rechenberga). Typowe zastosowanie: Optymalizacja
Przegląd budowy i funkcji białek
 Przegląd budowy i funkcji białek Co piszą o białkach? Wyraz wprowadzony przez Jönsa J. Berzeliusa w 1883 r. w celu podkreślenia znaczenia tej grupy związków. Termin pochodzi od greckiego słowa proteios,
Przegląd budowy i funkcji białek Co piszą o białkach? Wyraz wprowadzony przez Jönsa J. Berzeliusa w 1883 r. w celu podkreślenia znaczenia tej grupy związków. Termin pochodzi od greckiego słowa proteios,
WAE Jarosław Arabas Pełny schemat algorytmu ewolucyjnego
 WAE Jarosław Arabas Pełny schemat algorytmu ewolucyjnego Algorytm ewolucyjny algorytm ewolucyjny inicjuj P 0 {P 0 1, P 0 2... P 0 μ } t 0 H P 0 while! stop for (i 1: λ) if (a< p c ) O t i mutation(crossover
WAE Jarosław Arabas Pełny schemat algorytmu ewolucyjnego Algorytm ewolucyjny algorytm ewolucyjny inicjuj P 0 {P 0 1, P 0 2... P 0 μ } t 0 H P 0 while! stop for (i 1: λ) if (a< p c ) O t i mutation(crossover
Pamiętając o komplementarności zasad azotowych, dopisz sekwencję nukleotydów brakującej nici DNA. A C C G T G C C A A T C G A...
 1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
Konkurs szkolny Mistrz genetyki etap II
 onkurs szkolny istrz genetyki etap II 1.W D pewnego pierwotniaka tymina stanowi 28 % wszystkich zasad azotowych. blicz i zapisz, jaka jest zawartość procentowa każdej z pozostałych zasad w D tego pierwotniaka.
onkurs szkolny istrz genetyki etap II 1.W D pewnego pierwotniaka tymina stanowi 28 % wszystkich zasad azotowych. blicz i zapisz, jaka jest zawartość procentowa każdej z pozostałych zasad w D tego pierwotniaka.
Systemy Inteligentnego Przetwarzania wykład 4: algorytmy genetyczne, logika rozmyta
 Systemy Inteligentnego Przetwarzania wykład 4: algorytmy genetyczne, logika rozmyta Dr inż. Jacek Mazurkiewicz Katedra Informatyki Technicznej e-mail: Jacek.Mazurkiewicz@pwr.edu.pl Wprowadzenie Problemy
Systemy Inteligentnego Przetwarzania wykład 4: algorytmy genetyczne, logika rozmyta Dr inż. Jacek Mazurkiewicz Katedra Informatyki Technicznej e-mail: Jacek.Mazurkiewicz@pwr.edu.pl Wprowadzenie Problemy
Algorytmy memetyczne (hybrydowe algorytmy ewolucyjne)
") Algorytmy memetyczne (hybrydowe algorytmy ewolucyjne) 1 2 Wstęp Termin zaproponowany przez Pablo Moscato (1989). Kombinacja algorytmu ewolucyjnego z algorytmem poszukiwań lokalnych, tak że algorytm poszukiwań
Algorytmy memetyczne (hybrydowe algorytmy ewolucyjne) 1 2 Wstęp Termin zaproponowany przez Pablo Moscato (1989). Kombinacja algorytmu ewolucyjnego z algorytmem poszukiwań lokalnych, tak że algorytm poszukiwań
Przykład 1 W przypadku jednokrotnego rzutu kostką przestrzeń zdarzeń elementarnych
 Rozdział 1 Zmienne losowe, ich rozkłady i charakterystyki 1.1 Definicja zmiennej losowej Niech Ω będzie przestrzenią zdarzeń elementarnych. Definicja 1 Rodzinę S zdarzeń losowych (zbiór S podzbiorów zbioru
Rozdział 1 Zmienne losowe, ich rozkłady i charakterystyki 1.1 Definicja zmiennej losowej Niech Ω będzie przestrzenią zdarzeń elementarnych. Definicja 1 Rodzinę S zdarzeń losowych (zbiór S podzbiorów zbioru
Statystyka i opracowanie danych Podstawy wnioskowania statystycznego. Prawo wielkich liczb. Centralne twierdzenie graniczne. Estymacja i estymatory
 Statystyka i opracowanie danych Podstawy wnioskowania statystycznego. Prawo wielkich liczb. Centralne twierdzenie graniczne. Estymacja i estymatory Dr Anna ADRIAN Paw B5, pok 407 adrian@tempus.metal.agh.edu.pl
Statystyka i opracowanie danych Podstawy wnioskowania statystycznego. Prawo wielkich liczb. Centralne twierdzenie graniczne. Estymacja i estymatory Dr Anna ADRIAN Paw B5, pok 407 adrian@tempus.metal.agh.edu.pl
Imię i nazwisko...kl...
 Gimnazjum nr 4 im. Ojca Świętego Jana Pawła II we Wrocławiu SPRAWDZIAN GENETYKA GR. A Imię i nazwisko...kl.... 1. Nauka o regułach i mechanizmach dziedziczenia to: (0-1pkt) a) cytologia b) biochemia c)
Gimnazjum nr 4 im. Ojca Świętego Jana Pawła II we Wrocławiu SPRAWDZIAN GENETYKA GR. A Imię i nazwisko...kl.... 1. Nauka o regułach i mechanizmach dziedziczenia to: (0-1pkt) a) cytologia b) biochemia c)
Algorytmy genetyczne Michał Bereta Paweł Jarosz (część teoretyczna)
") 1 Zagadnienia Sztucznej Inteligencji laboratorium Wprowadzenie Algorytmy genetyczne Michał Bereta Paweł Jarosz (część teoretyczna) Dana jest funkcja f, jednej lub wielu zmiennych. Należy określić wartości
1 Zagadnienia Sztucznej Inteligencji laboratorium Wprowadzenie Algorytmy genetyczne Michał Bereta Paweł Jarosz (część teoretyczna) Dana jest funkcja f, jednej lub wielu zmiennych. Należy określić wartości
Automatyczny dobór parametrów algorytmu genetycznego
 Automatyczny dobór parametrów algorytmu genetycznego Remigiusz Modrzejewski 22 grudnia 2008 Plan prezentacji Wstęp Atrakcyjność Pułapki Klasyfikacja Wstęp Atrakcyjność Pułapki Klasyfikacja Konstrukcja
Automatyczny dobór parametrów algorytmu genetycznego Remigiusz Modrzejewski 22 grudnia 2008 Plan prezentacji Wstęp Atrakcyjność Pułapki Klasyfikacja Wstęp Atrakcyjność Pułapki Klasyfikacja Konstrukcja
SZTUCZNA INTELIGENCJA
 SZTUCZNA INTELIGENCJA WYKŁAD 17. ALGORYTMY EWOLUCYJNE Częstochowa 2014 Dr hab. inż. Grzegorz Dudek Wydział Elektryczny Politechnika Częstochowska KODOWANIE BINARNE Problem różnych struktur przestrzeni
SZTUCZNA INTELIGENCJA WYKŁAD 17. ALGORYTMY EWOLUCYJNE Częstochowa 2014 Dr hab. inż. Grzegorz Dudek Wydział Elektryczny Politechnika Częstochowska KODOWANIE BINARNE Problem różnych struktur przestrzeni
BIOCYBERNETYKA ALGORYTMY GENETYCZNE I METODY EWOLUCYJNE. Adrian Horzyk. Akademia Górniczo-Hutnicza
 BIOCYBERNETYKA ALGORYTMY GENETYCZNE I METODY EWOLUCYJNE Adrian Horzyk Akademia Górniczo-Hutnicza Wydział Elektrotechniki, Automatyki, Informatyki i Inżynierii Biomedycznej Katedra Automatyki i Inżynierii
BIOCYBERNETYKA ALGORYTMY GENETYCZNE I METODY EWOLUCYJNE Adrian Horzyk Akademia Górniczo-Hutnicza Wydział Elektrotechniki, Automatyki, Informatyki i Inżynierii Biomedycznej Katedra Automatyki i Inżynierii
METODY SZTUCZNEJ INTELIGENCJI algorytmy ewolucyjne
 METODY SZTUCZNEJ INTELIGENCJI algorytmy ewolucyjne dr hab. inż. Andrzej Obuchowicz, prof. UZ Instytut Sterowania i Systemów Informatycznych Uniwersytet Zielonogórski A. Obuchowicz: MSI - algorytmy ewolucyjne
METODY SZTUCZNEJ INTELIGENCJI algorytmy ewolucyjne dr hab. inż. Andrzej Obuchowicz, prof. UZ Instytut Sterowania i Systemów Informatycznych Uniwersytet Zielonogórski A. Obuchowicz: MSI - algorytmy ewolucyjne
Algorytmy stochastyczne laboratorium 03
 Algorytmy stochastyczne laboratorium 03 Jarosław Piersa 10 marca 2014 1 Projekty 1.1 Problem plecakowy (1p) Oznaczenia: dany zbiór przedmiotów x 1,.., x N, każdy przedmiot ma określoną wagę w(x i ) i wartość
Algorytmy stochastyczne laboratorium 03 Jarosław Piersa 10 marca 2014 1 Projekty 1.1 Problem plecakowy (1p) Oznaczenia: dany zbiór przedmiotów x 1,.., x N, każdy przedmiot ma określoną wagę w(x i ) i wartość
ALGORYTMY GENETYCZNE (wykład + ćwiczenia)
") ALGORYTMY GENETYCZNE (wykład + ćwiczenia) Prof. dr hab. Krzysztof Dems Treści programowe: 1. Metody rozwiązywania problemów matematycznych i informatycznych.. Elementarny algorytm genetyczny: definicja
ALGORYTMY GENETYCZNE (wykład + ćwiczenia) Prof. dr hab. Krzysztof Dems Treści programowe: 1. Metody rozwiązywania problemów matematycznych i informatycznych.. Elementarny algorytm genetyczny: definicja
OBLICZENIA EWOLUCYJNE
 1 OBLICZENIA EWOLUCYJNE FITNESS F. START COMPUTATION FITNESS F. COMPUTATION INITIAL SUBPOPULATION SENDING CHROM. TO COMPUTERS chromosome AND RECEIVING FITNESS F. VALUE fitness f. value wykład 2 communication
1 OBLICZENIA EWOLUCYJNE FITNESS F. START COMPUTATION FITNESS F. COMPUTATION INITIAL SUBPOPULATION SENDING CHROM. TO COMPUTERS chromosome AND RECEIVING FITNESS F. VALUE fitness f. value wykład 2 communication
LABORATORIUM 5: Wpływ reprodukcji na skuteczność poszukiwań AE. opracował: dr inż. Witold Beluch
 OBLICZENIA EWOLUCYJNE LABORATORIUM 5: Wpływ reprodukcji na skuteczność poszukiwań AE opracował: dr inż. Witold Beluch witold.beluch@polsl.pl Gliwice 2012 OBLICZENIA EWOLUCYJNE LABORATORIUM 5 2 Cel ćwiczenia
OBLICZENIA EWOLUCYJNE LABORATORIUM 5: Wpływ reprodukcji na skuteczność poszukiwań AE opracował: dr inż. Witold Beluch witold.beluch@polsl.pl Gliwice 2012 OBLICZENIA EWOLUCYJNE LABORATORIUM 5 2 Cel ćwiczenia
Metody przeszukiwania
 Metody przeszukiwania Co to jest przeszukiwanie Przeszukiwanie polega na odnajdywaniu rozwiązania w dyskretnej przestrzeni rozwiązao. Zwykle przeszukiwanie polega na znalezieniu określonego rozwiązania
Metody przeszukiwania Co to jest przeszukiwanie Przeszukiwanie polega na odnajdywaniu rozwiązania w dyskretnej przestrzeni rozwiązao. Zwykle przeszukiwanie polega na znalezieniu określonego rozwiązania
Algorytmy ewolucyjne 1
 Algorytmy ewolucyjne 1 2 Zasady zaliczenia przedmiotu Prowadzący (wykład i pracownie specjalistyczną): Wojciech Kwedlo, pokój 205. Konsultacje dla studentów studiów dziennych: poniedziałek,środa, godz
Algorytmy ewolucyjne 1 2 Zasady zaliczenia przedmiotu Prowadzący (wykład i pracownie specjalistyczną): Wojciech Kwedlo, pokój 205. Konsultacje dla studentów studiów dziennych: poniedziałek,środa, godz
1 Genetykapopulacyjna
 1 Genetykapopulacyjna Genetyka populacyjna zajmuje się badaniem częstości występowania poszczególnych alleli oraz genotypów w populacji. Bada także zmiany tych częstości spowodowane doborem naturalnym
1 Genetykapopulacyjna Genetyka populacyjna zajmuje się badaniem częstości występowania poszczególnych alleli oraz genotypów w populacji. Bada także zmiany tych częstości spowodowane doborem naturalnym
Zadanie 1. Liczba szkód N w ciągu roku z pewnego ryzyka ma rozkład geometryczny: k =
 Matematyka ubezpieczeń majątkowych 0.0.006 r. Zadanie. Liczba szkód N w ciągu roku z pewnego ryzyka ma rozkład geometryczny: k 5 Pr( N = k) =, k = 0,,,... 6 6 Wartości kolejnych szkód Y, Y,, są i.i.d.,
Matematyka ubezpieczeń majątkowych 0.0.006 r. Zadanie. Liczba szkód N w ciągu roku z pewnego ryzyka ma rozkład geometryczny: k 5 Pr( N = k) =, k = 0,,,... 6 6 Wartości kolejnych szkód Y, Y,, są i.i.d.,
OBLICZENIA EWOLUCYJNE
 METODY ANALITYCZNE kontra AG/AE OBLICZENIA EWOLUCYJNE FITNESS F. START COMPUTATION FITNESS F. COMPUTATION INITIAL SUBPOPULATION SENDING CHROM. TO COMPUTERS chromosome EVOLUTIONARY OPERATORS AND RECEIVING
METODY ANALITYCZNE kontra AG/AE OBLICZENIA EWOLUCYJNE FITNESS F. START COMPUTATION FITNESS F. COMPUTATION INITIAL SUBPOPULATION SENDING CHROM. TO COMPUTERS chromosome EVOLUTIONARY OPERATORS AND RECEIVING
przetworzonego sygnału
 Synteza falek ortogonalnych na podstawie oceny przetworzonego sygnału Instytut Informatyki Politechnika Łódzka 28 lutego 2012 Plan prezentacji 1 Sformułowanie problemu 2 3 4 Historia przekształcenia falkowego
Synteza falek ortogonalnych na podstawie oceny przetworzonego sygnału Instytut Informatyki Politechnika Łódzka 28 lutego 2012 Plan prezentacji 1 Sformułowanie problemu 2 3 4 Historia przekształcenia falkowego
LABORATORIUM 3: Wpływ operatorów krzyżowania na skuteczność poszukiwań AE
 Instytut Mechaniki i Inżynierii Obliczeniowej Wydział Mechaniczny Technologiczny, Politechnika Śląska www.imio.polsl.pl OBLICZENIA EWOLUCYJNE LABORATORIUM 3: Wpływ operatorów krzyżowania na skuteczność
Instytut Mechaniki i Inżynierii Obliczeniowej Wydział Mechaniczny Technologiczny, Politechnika Śląska www.imio.polsl.pl OBLICZENIA EWOLUCYJNE LABORATORIUM 3: Wpływ operatorów krzyżowania na skuteczność
Stochastyczna dynamika z opóźnieniem czasowym w grach ewolucyjnych oraz modelach ekspresji i regulacji genów
 Stochastyczna dynamika z opóźnieniem czasowym w grach ewolucyjnych oraz modelach ekspresji i regulacji genów Jacek Miękisz Instytut Matematyki Stosowanej i Mechaniki Uniwersytet Warszawski Warszawa 14
Stochastyczna dynamika z opóźnieniem czasowym w grach ewolucyjnych oraz modelach ekspresji i regulacji genów Jacek Miękisz Instytut Matematyki Stosowanej i Mechaniki Uniwersytet Warszawski Warszawa 14
W rachunku prawdopodobieństwa wyróżniamy dwie zasadnicze grupy rozkładów zmiennych losowych:
 W rachunku prawdopodobieństwa wyróżniamy dwie zasadnicze grupy rozkładów zmiennych losowych: Zmienne losowe skokowe (dyskretne) przyjmujące co najwyżej przeliczalnie wiele wartości Zmienne losowe ciągłe
W rachunku prawdopodobieństwa wyróżniamy dwie zasadnicze grupy rozkładów zmiennych losowych: Zmienne losowe skokowe (dyskretne) przyjmujące co najwyżej przeliczalnie wiele wartości Zmienne losowe ciągłe
SYSTEMY UCZĄCE SIĘ WYKŁAD 4. DRZEWA REGRESYJNE, INDUKCJA REGUŁ. Dr hab. inż. Grzegorz Dudek Wydział Elektryczny Politechnika Częstochowska
 SYSTEMY UCZĄCE SIĘ WYKŁAD 4. DRZEWA REGRESYJNE, INDUKCJA REGUŁ Częstochowa 2014 Dr hab. inż. Grzegorz Dudek Wydział Elektryczny Politechnika Częstochowska DRZEWO REGRESYJNE Sposób konstrukcji i przycinania
SYSTEMY UCZĄCE SIĘ WYKŁAD 4. DRZEWA REGRESYJNE, INDUKCJA REGUŁ Częstochowa 2014 Dr hab. inż. Grzegorz Dudek Wydział Elektryczny Politechnika Częstochowska DRZEWO REGRESYJNE Sposób konstrukcji i przycinania
SZTUCZNA INTELIGENCJA
 SZTUCZNA INTELIGENCJA WYKŁAD 15. ALGORYTMY GENETYCZNE Częstochowa 014 Dr hab. nż. Grzegorz Dudek Wydzał Elektryczny Poltechnka Częstochowska TERMINOLOGIA allele wartośc, waranty genów, chromosom - (naczej
SZTUCZNA INTELIGENCJA WYKŁAD 15. ALGORYTMY GENETYCZNE Częstochowa 014 Dr hab. nż. Grzegorz Dudek Wydzał Elektryczny Poltechnka Częstochowska TERMINOLOGIA allele wartośc, waranty genów, chromosom - (naczej
2. Tablice. Tablice jednowymiarowe - wektory. Algorytmy i Struktury Danych
 2. Tablice Tablica to struktura danych przechowująca elementy jednego typu (jednorodna). Dostęp do poszczególnych elementów składowych jest możliwy za pomocą indeksów. Rozróżniamy następujące typy tablic:
2. Tablice Tablica to struktura danych przechowująca elementy jednego typu (jednorodna). Dostęp do poszczególnych elementów składowych jest możliwy za pomocą indeksów. Rozróżniamy następujące typy tablic:
Aproksymacja funkcji a regresja symboliczna
 Aproksymacja funkcji a regresja symboliczna Problem aproksymacji funkcji polega na tym, że funkcję F(x), znaną lub określoną tablicą wartości, należy zastąpić inną funkcją, f(x), zwaną funkcją aproksymującą
Aproksymacja funkcji a regresja symboliczna Problem aproksymacji funkcji polega na tym, że funkcję F(x), znaną lub określoną tablicą wartości, należy zastąpić inną funkcją, f(x), zwaną funkcją aproksymującą
MACIERZE MUTACYJNE W ANALIZIE GENOMÓW czy możliwa jest rekonstrukcja filogenetyczna? Aleksandra Nowicka
 MAIERZE MUTAYJNE W ANALIZIE GENOMÓW czy możliwa jest rekonstrukcja filogenetyczna? Aleksandra Nowicka Zadaniem FILOGENETYKI jest : zrekonstruowanie ewolucyjnej historii wszystkich organizmów odkrycie przodka
MAIERZE MUTAYJNE W ANALIZIE GENOMÓW czy możliwa jest rekonstrukcja filogenetyczna? Aleksandra Nowicka Zadaniem FILOGENETYKI jest : zrekonstruowanie ewolucyjnej historii wszystkich organizmów odkrycie przodka
Metody numeryczne Technika obliczeniowa i symulacyjna Sem. 2, EiT, 2014/2015
 Metody numeryczne Technika obliczeniowa i symulacyjna Sem. 2, EiT, 2014/2015 1 Metody numeryczne Dział matematyki Metody rozwiązywania problemów matematycznych za pomocą operacji na liczbach. Otrzymywane
Metody numeryczne Technika obliczeniowa i symulacyjna Sem. 2, EiT, 2014/2015 1 Metody numeryczne Dział matematyki Metody rozwiązywania problemów matematycznych za pomocą operacji na liczbach. Otrzymywane
Algorytmy genetyczne służą głównie do tego, żeby rozwiązywać zadania optymalizacji
 Kolejna metoda informatyczna inspirowana przez Naturę - algorytmy genetyczne Struktura molekuły DNA nośnika informacji genetycznej w biologii Motto: Zamiast pracowicie poszukiwać najlepszego rozwiązania
Kolejna metoda informatyczna inspirowana przez Naturę - algorytmy genetyczne Struktura molekuły DNA nośnika informacji genetycznej w biologii Motto: Zamiast pracowicie poszukiwać najlepszego rozwiązania
Program "FLiNN-GA" wersja 2.10.β
 POLSKIE TOWARZYSTWO SIECI NEURONOWYCH POLITECHNIKA CZĘSTOCHOWSKA Zakład Elektroniki, Informatyki i Automatyki Maciej Piliński Robert Nowicki - GA Program "FLiNN-GA" wersja 2.10.β Podręcznik użytkownika
POLSKIE TOWARZYSTWO SIECI NEURONOWYCH POLITECHNIKA CZĘSTOCHOWSKA Zakład Elektroniki, Informatyki i Automatyki Maciej Piliński Robert Nowicki - GA Program "FLiNN-GA" wersja 2.10.β Podręcznik użytkownika
Teoria grafów - Teoria rewersali - Teoria śladów
 17 maja 2012 1 Planarność Wzór Eulera Kryterium Kuratowskiego Algorytmy testujące planarność 2 Genom i jego przekształcenia Grafy złamań Sortowanie przez odwrócenia Inne rodzaje sortowania Algorytmy sortujące
17 maja 2012 1 Planarność Wzór Eulera Kryterium Kuratowskiego Algorytmy testujące planarność 2 Genom i jego przekształcenia Grafy złamań Sortowanie przez odwrócenia Inne rodzaje sortowania Algorytmy sortujące
Algorytmy ewolucyjne (3)
") Algorytmy ewolucyjne (3) http://zajecia.jakubw.pl/nai KODOWANIE PERMUTACJI W pewnych zastosowaniach kodowanie binarne jest mniej naturalne, niż inne sposoby kodowania. Na przykład, w problemie komiwojażera
Algorytmy ewolucyjne (3) http://zajecia.jakubw.pl/nai KODOWANIE PERMUTACJI W pewnych zastosowaniach kodowanie binarne jest mniej naturalne, niż inne sposoby kodowania. Na przykład, w problemie komiwojażera
Mutacje jako źródło różnorodności wewnątrzgatunkowej
 Mutacje jako źródło różnorodności wewnątrzgatunkowej Zajęcia terenowe: Zajęcia w klasie: Poziom nauczania oraz odniesienie do podstawy programowej: Liceum IV etap edukacyjny zakres rozszerzony: Różnorodność
Mutacje jako źródło różnorodności wewnątrzgatunkowej Zajęcia terenowe: Zajęcia w klasie: Poziom nauczania oraz odniesienie do podstawy programowej: Liceum IV etap edukacyjny zakres rozszerzony: Różnorodność
Zadanie transportowe i problem komiwojażera. Tadeusz Trzaskalik
 Zadanie transportowe i problem komiwojażera Tadeusz Trzaskalik 3.. Wprowadzenie Słowa kluczowe Zbilansowane zadanie transportowe Rozwiązanie początkowe Metoda minimalnego elementu macierzy kosztów Metoda
Zadanie transportowe i problem komiwojażera Tadeusz Trzaskalik 3.. Wprowadzenie Słowa kluczowe Zbilansowane zadanie transportowe Rozwiązanie początkowe Metoda minimalnego elementu macierzy kosztów Metoda
spektroskopia elektronowa (UV-vis)
") spektroskopia elektronowa (UV-vis) rodzaje przejść elektronowych Energia σ* π* 3 n 3 π σ σ σ* daleki nadfiolet (λ max < 200 nm) π π* bliski nadfiolet jednostki energii atomowa jednostka energii = energia
spektroskopia elektronowa (UV-vis) rodzaje przejść elektronowych Energia σ* π* 3 n 3 π σ σ σ* daleki nadfiolet (λ max < 200 nm) π π* bliski nadfiolet jednostki energii atomowa jednostka energii = energia
Biostatystyka, # 3 /Weterynaria I/
 Biostatystyka, # 3 /Weterynaria I/ dr n. mat. Zdzisław Otachel Uniwersytet Przyrodniczy w Lublinie Katedra Zastosowań Matematyki i Informatyki ul. Głęboka 28, p. 221 bud. CIW, e-mail: zdzislaw.otachel@up.lublin.pl
Biostatystyka, # 3 /Weterynaria I/ dr n. mat. Zdzisław Otachel Uniwersytet Przyrodniczy w Lublinie Katedra Zastosowań Matematyki i Informatyki ul. Głęboka 28, p. 221 bud. CIW, e-mail: zdzislaw.otachel@up.lublin.pl
Rachunek Prawdopodobieństwa i Statystyka
 Rachunek Prawdopodobieństwa i Statystyka W 2. Probabilistyczne modele danych Zmienne losowe. Rozkład prawdopodobieństwa i dystrybuanta. Wartość oczekiwana i wariancja zmiennej losowej Dr Anna ADRIAN Zmienne
Rachunek Prawdopodobieństwa i Statystyka W 2. Probabilistyczne modele danych Zmienne losowe. Rozkład prawdopodobieństwa i dystrybuanta. Wartość oczekiwana i wariancja zmiennej losowej Dr Anna ADRIAN Zmienne
ZARZĄDZANIE POPULACJAMI ZWIERZĄT
 ZARZĄDZANIE POPULACJAMI ZWIERZĄT Ćwiczenia 1 mgr Magda Kaczmarek-Okrój magda_kaczmarek_okroj@sggw.pl 1 ZAGADNIENIA struktura genetyczna populacji obliczanie frekwencji genotypów obliczanie frekwencji alleli
ZARZĄDZANIE POPULACJAMI ZWIERZĄT Ćwiczenia 1 mgr Magda Kaczmarek-Okrój magda_kaczmarek_okroj@sggw.pl 1 ZAGADNIENIA struktura genetyczna populacji obliczanie frekwencji genotypów obliczanie frekwencji alleli
Programowanie genetyczne, gra SNAKE
 STUDENCKA PRACOWNIA ALGORYTMÓW EWOLUCYJNYCH Tomasz Kupczyk, Tomasz Urbański Programowanie genetyczne, gra SNAKE II UWr Wrocław 2009 Spis treści 1. Wstęp 3 1.1. Ogólny opis.....................................
STUDENCKA PRACOWNIA ALGORYTMÓW EWOLUCYJNYCH Tomasz Kupczyk, Tomasz Urbański Programowanie genetyczne, gra SNAKE II UWr Wrocław 2009 Spis treści 1. Wstęp 3 1.1. Ogólny opis.....................................
GENETYKA POPULACJI. Ćwiczenia 1 Biologia I MGR /
 GENETYKA POPULACJI Ćwiczenia 1 Biologia I MGR 1 ZAGADNIENIA struktura genetyczna populacji obliczanie frekwencji genotypów obliczanie frekwencji alleli przewidywanie struktury następnego pokolenia przy
GENETYKA POPULACJI Ćwiczenia 1 Biologia I MGR 1 ZAGADNIENIA struktura genetyczna populacji obliczanie frekwencji genotypów obliczanie frekwencji alleli przewidywanie struktury następnego pokolenia przy
N ma rozkład Poissona z wartością oczekiwaną równą 100 M, M M mają ten sam rozkład dwupunktowy o prawdopodobieństwach:
 Zadanie. O niezależnych zmiennych losowych N, M M, M 2, 3 wiemy, że: N ma rozkład Poissona z wartością oczekiwaną równą 00 M, M M mają ten sam rozkład dwupunktowy o prawdopodobieństwach: 2, 3 Pr( M = )
Zadanie. O niezależnych zmiennych losowych N, M M, M 2, 3 wiemy, że: N ma rozkład Poissona z wartością oczekiwaną równą 00 M, M M mają ten sam rozkład dwupunktowy o prawdopodobieństwach: 2, 3 Pr( M = )
Zadania laboratoryjne i projektowe - wersja β
 Zadania laboratoryjne i projektowe - wersja β 1 Laboratorium Dwa problemy do wyboru (jeden do realizacji). 1. Water Jug Problem, 2. Wieże Hanoi. Water Jug Problem Ograniczenia dla każdej z wersji: pojemniki
Zadania laboratoryjne i projektowe - wersja β 1 Laboratorium Dwa problemy do wyboru (jeden do realizacji). 1. Water Jug Problem, 2. Wieże Hanoi. Water Jug Problem Ograniczenia dla każdej z wersji: pojemniki
Zadania do wykonania. Rozwiązując poniższe zadania użyj pętlę for.
 Zadania do wykonania Rozwiązując poniższe zadania użyj pętlę for. 1. apisz program, który przesuwa w prawo o dwie pozycje zawartość tablicy 10-cio elementowej liczb całkowitych tzn. element t[i] dla i=2,..,9
Zadania do wykonania Rozwiązując poniższe zadania użyj pętlę for. 1. apisz program, który przesuwa w prawo o dwie pozycje zawartość tablicy 10-cio elementowej liczb całkowitych tzn. element t[i] dla i=2,..,9