Jak z wody i CO 2 zrobid cukier? Istota autotroficznego odżywiania roślin
|
|
|
- Maria Matusiak
- 8 lat temu
- Przeglądów:
Transkrypt


1 Wykład Fotosynteza. Barwniki fotosyntetyczne, przebieg fazy świetlnej i ciemniowej fotosyntezy Autor dr Agnieszka Gniazdowska-Piekarska W tym wykładzie dowiesz się co to jest fotosynteza i jak przebiega ten proces oraz jakie czynniki go regulują. Zapoznasz się również z właściwościami podstawowych barwników fotosyntetycznych występujących u roślin. Jak z wody i CO 2 zrobid cukier? Istota autotroficznego odżywiania roślin Fotosynteza to najważniejszy proces bioenergetyczny na Ziemi. Organizmy fotosyntetyczne wytwarzają związki organiczne (głównie cukry) z wody i CO 2 z wykorzystaniem energii słonecznej. 6H 2 O + 6CO 2 + hν (energia świetlna) C 6 H 12 O 6 + 6O 2 CO 2 H 2 O cykl Calvina O 2 asymilaty Uproszczony sumaryczny przebieg fotosyntezy z glukozą jako syntezowanym węglowodanem Fotosynteza (gr. światło,łączenie) proces anaboliczny polegający na redukcji dwutlenku węgla (CO 2 ) z wykorzystaniem promieniowania słonecznego w obecności barwników

2 asymilacyjnych i enzymów cyklu Calvina-Bensona. Fotosynteza prowadzi do powstania związków organicznych. Proces ten utrzymuje wysoki poziom tlenu w atmosferze oraz przyczynia się do wzrostu ilości węgla organicznego zwiększając masę materii organicznej, kosztem materii nieorganicznej. Zachodzi w dwóch etapach. Faza jasna (świetlna, fotochemiczna, bezpośrednio zależna od światła) to faza przemiany energii, w której światło jest absorbowane, a jego energia ulega zamianie na energię wiązań chemicznych. Jako produkt uboczny wydzielany jest tlen. Faza ciemniowa (faza biochemiczna, pośrednio zależna od światła), w której energia wiązań chemicznych, związków powstałych w fazie świetlnej, jest wykorzystywana do syntezy związków organicznych. Liśd jako podstawowy organ asymilacyjny Budowa anatomiczno - morfologiczna blaszki liścia jest determinowana przez podstawową funkcję asymilacyjną liścia. Liśd odpowiada za: -pochłanianie energii słonecznej, -pobieranie dwutlenku węgla (aparaty szparkowe), - produkcję asymilatów i ich odprowadzanie do niefotosyntetyzujących tkanek (np. korzeni). pomidora Liście asymilacyjne chrzanu pokrzywy truskawki Zdjęcia własne AG Funkcja liścia związana z wykorzystywaniem energii świetlnej w procesie fotosyntezy determinuje jego budowę. Liść pochłania energię świetlną, pobiera CO 2 i odprowadza asymilaty. Zwiększenie efektywności pochłaniania światła zapewnia zwiększenie stosunku powierzchni do objętości liścia. Przystosowaniem do funkcji asymilacyjnej jest zdolność liści do wykonywania ruchów tropicznych (fototropizm)- uzależnionych od kierunku działania bodźca (światła). Liście układając się optymalnie w stosunku do kierunku padania światła tworzą mozaikę liściową (każdy liść ma zapewniony dostęp do światła). Podobnie, ułożenie liści na łodydze pozwala na uniknięcia wzajemnego zacieniania się.

3 Lokalizacja procesu fotosyntezy Proces fotosyntezy zachodzi w komórkach miękiszu asymilacyjnego liści w wyspecjalizowanych organellach chloroplastach. Chloroplasty w komórkach glonu Elodea faculty.ksu.edu.sa/.../physiological%20photos Reakcje fotosyntezy zachodzą w komórkach miękiszowych liścia w wyspecjalizowanych organellach chloroplastach. Na zdjęciach z mikroskopu elektronowego chloroplasty mają kształt dyskoidalny, ich średnica wynosi 4-10 µm, grubość poniżej 2 µm. Zwykle w jednej komórce występuje kilkadziesiąt chloroplastów. Chloroplast ultrastruktura Opis schematu przedstawiającego ultrastrukturę chloroplastu: 1. zewnętrzna błona 2. przestrzeo międzybłonowa 3. wewnętrzna błona (1+2+3: otoczka) 4. stroma (roztwór koloidalny) 5. wnętrze tylakoidu (lumen) 6. błony tylakoidów 7. granum (stos tylakoidów) 8. tylakoidy (lamella) 9. skrobia 10. rybosomy 11. chloroplastowe DNA 12. plastoglobule (krople lipidów).

4 Analizując budowę wewnętrzną chloroplastu wyróżniamy kilka przedziałów: otoczkę (składającą się z dwóch błon), stromę oraz rozbudowany system spłaszczonych pęcherzyków nazywanych tylakoidami, z których część łączy się w stosy tzw. grana. Tylakoidy, które swobodnie przenikają stromę nazywamy tylakoidami stromy, część tylakoidów zasocjowanych ze sobą w formie stosów tylakoidami gran. Na granum składa się od 5-20 zespolonych błon tylakoidowych. Względny udział błon tylakoidowych gran i stromy jest dla większości roślin podobny, jeśli porównuje się rośliny rosnące w warunkach umiarkowanego natężenia promieniowania świetlnego. Tylakoidy gran stanowią około 80 % a tylakoidy stromy 20 % systemu tylakoidowego pojedynczego chloroplastu. W niektórych komórkach (komórki pochwy około wiązkowej liści roślin o fotosyntezie C4) występują tylko niezespolone tylakoidy stromy. W stromie chloroplastu poza strukturami błoniastymi występują obszary zawierające: DNA, rybosomy, ziarna skrobi, lipidy i białka. Replikacja DNA chloroplastowego zachodzi niezależnie od replikacji DNA jądrowego. Podziały chloroplastów są niezależne od podziałów komórkowych. Genom chloroplastowy uczestniczy w syntezie niewielkiej ilości białek zaangażowanych w procesie fotosyntezy, większość białek enzymów fotosyntetycznych jest kodowana przez genom jądrowy. Białko Rubisco (karboksylaza 1,5-bisfosforybulozy) kluczowego enzymu wiążącego CO 2 powstaje w wyniku współdziałania genomu jądrowego i plastydowego. Barwniki fotosyntetyczne występują w błonach chloroplastów Absorpcja światła w procesie fotosyntezy zachodzi dzięki obecności barwników fotosyntetycznych. Głównymi barwnikami fotosyntetycznymi są: chlorofil a i chlorofil b (u roślin wyższych) - barwniki zielone. Barwnikami pomocniczymi są karotenoidy: karoteny i ksantofile barwniki żółto-pomaraoczowe. U glonów i sinic występują fikobiliny barwniki niebiesko-filetowe. Zdjęcia własne AG
5 pl.wikipedia.org/wiki/chlorofil Spośród barwników fotosyntetycznych występujących w chloroplastach najliczniejszą grupę stanowią chlorofile. Ilościowo w błonach tylakoidów dominuje chlorofil a, jednak u wszystkich roślin wyższych stwierdza się także obecność chlorofilu b. Błony tylakoidowe glonów (brunatnic, okrzemek, złotowiciowców) zawierają także inne aktywne fotosyntetycznie formy chlorofilu np. chlorofile c 1 i c 2, u niektórych krasnorostów występuje ponadto chlorofil d. Chlorofile odgrywają kluczową role w fazie świetlnej fotosyntezy, dzięki zdolności do absorbowania promieniowania świetlnego w zakresie światła widzialnego, dlatego też chlorofile a występują w centrum reakcji fotochemicznych fotoukładu I (PSI) i fotoukładu II (PSII). Drugą grupą barwników fotosyntetycznych są żółte karotenoidy, na które przypada nie więcej niż 5 % suchej masy błon tylakoidów. Zawartość karotenoidów jest zwykle 4-7 razy niższa niż zawartość chlorofili. Karotenoidy wraz z chlorofilami wchodzą w skład anten fotosyntetycznych (kompleksów barwnikowo- białkowych) zbierających energię promieniowania i przekazujących ją na chlorofile a umieszczone w centrum reakcji fotochemicznej. W skład anten wchodzi ponad 99 % populacji cząsteczek chlorofilu a i 100% populacji cząsteczek chlorofilu b. Chlorofil a i chlorofil b podstawowe barwniki asymilacyjne roślin wyższych Wzór strukturalny cząsteczki chlorofilu a Wzór strukturalny cząsteczki chlorofilu b

6 pl.wikipedia.org/wiki/chlorofil U roślin wyższych chlorofil występuje w dwóch odmianach: niebieskozielony chlorofil a, absorbujący głównie światło fioletowe i czerwone, oraz żółtozielony chlorofil b, absorbujący głównie światło niebieskie i pomarańczowe. Chlorofil b różni się od chlorofilu a jednym podstawnikiem. W drugim pierścieniu pirolowym chlorofilu a występuje grupa metylowa CH 3, a w chlorofilu b grupa aldehydowa CHO. Stosunek ilościowy chlorofilu a do chlorofilu b wynosi zwykle 3:1, ale zależy on między innymi od warunków siedliskowych, rośliny cieniolubne mają więcej chlorofilu b, światłolubne chlorofilu a. Budowa cząsteczki chlorofilu feoporfiryna Wiązania sprzężone reszta fitolu Wzór strukturalny cząsteczki chlorofilu a Pod względem chemicznym chlorofil jest pochodną porfiryny. Cząsteczka chlorofilu składa się z polarnej głowy (feoporfiryny) i niepolarnego ogona fitolu (20 węglowy alkohol). Feoporfiryna to cztery pierścienie pirolowe i piąty pierścień niepirolowy. Wiązania pomiędzy atomami tworzącymi pierścienie to następujące po sobie wiązania pojedyncze i podwójne (-C=C-C) składające się na układ wiązań sprzężonych. Centralne miejsce w układzie porfiryny zajmuje atom magnezu (Mg) łączący się z atomami azotu każdego z pierścieni. Porfiryna tworząca kompleks z magnezem posiada zdolność do absorpcji promieniowania elektromagnetycznego w zakresie widzialnym dzięki obecności wiązań sprzężonych, które
7 zwierają wyjątkowo labilne elektrony, łatwo przechodzące z podstawowego stanu energetycznego na orbitale wzbudzone. Obecność magnezu w centrum układu porfirynowego umożliwia agregację cząsteczek chlorofilu, co ułatwia przekazywanie energii wzbudzenia pomiędzy cząsteczkami. Hydrofobowy (nie rozpuszczalny w wodzie) łańcuch fitolu pozwala na zakotwiczenie cząsteczki chlorofilu w błonie tylakoidu i zapewnia jej odpowiednią orientację. Chlorofil absorbuje światło widzialne w zakresie nm Widmo absorpcyjne cząsteczki chlorofilu a (zielona linia), chlorofilu b (czerwona linia) Widmo absorpcji roztworu chlorofilu w rozpuszczalnikach organicznych ma dwa wierzchołki (maksima). Maksima absorpcji chlorofilu przypadają na promieniowanie niebiesko-fioletowe i pomarańczowo-czerwone: Chlorofil a: 430 i 662 nm Chlorofil b: 453 i 642 nm. Chlorofil nie absorbuje światła zielonego, dlatego nadaje liściom barwę zieloną. W chloroplastach, chlorofile wchodzą w skład kompleksów barwnikowo-białkowych: fotosystemów oraz układów antenowych. Pojedynczy łańcuch polipeptydowy specyficznego białka wiąże od kilku do kilkunastu cząsteczek chlorofilu i dodatkowo kilka cząsteczek karotenoidów. Dlatego też, maksima widm absorpcji chlorofilu w nienaruszonych kompleksach barwnikowo-białkowych nie pokrywają się z widmami cząsteczek chlorofilu in vitro, lecz są przesunięte w kierunku fal dłuższych.
 ulega wzbudzeniu.")
8 Co się dzieje z energią fotonu zaabsorbowanego przez cząsteczkę chlorofilu? Cząsteczka chlorofilu po zaabsorbowaniu kwantu światła (fotonu) ulega wzbudzeniu. Pochłonięcie kwantu światła czerwonego (o niższej energii) wiąże się z przejściem do pierwszego stanu wzbudzenia, pochłonięcie kwantu światła niebieskiego (o wyższej energii) prowadzi do osiągnięcia drugiego stanu wzbudzenia. Wzbudzony elektron może powrócić do stanu podstawowego przez kilka różnych poziomów. Najprostszy sposób to relaksacja i emisja ciepła. Inny sposób to fluorescencja i emisja światła. Wreszcie, wzbudzona cząsteczka może utracić elektron przekazując go cząsteczce akceptorowej, następuje separacja ładunków i redukcja cząsteczki akceptorowej. Ten proces jest preferowany w cząsteczkach barwników. Stan wzbudzenia przekazywany jest przez kolejne cząsteczki chlorofilu do centrum reakcji cząsteczki chlorofilu a. Z chlorofilu stanowiącego centrum reakcji elektron jest wybijany, a następnie przekazywany na kolejne przekaźniki w fotosyntetycznym łańcuchu transportu elektronów. Zapamiętaj! Energia świetlna docierająca do blaszki liścia jest absorbowana przez barwniki fotosyntetyczne znajdujące się w kompleksach zbierających światło (antenach energetycznych). Z kompleksów antenowych jest ona przekazywana do centrów reakcji fotosyntetycznych PSI i PSII i wykorzystywana w reakcjach fotochemicznych.
9 Trzeba jednak pamiętać ze część energii wzbudzenia nim osiągnie centrum reakcji jest tracona w formie ciepła lub fluorescencji. Fluorescencję chlorofilu a obserwujemy jako emisję światła czerwonego Światło np. UV Roztwór zawierający cząsteczki chlorofilu Roztwór chlorofilu po oświetleniu światłem o dużej energii (światło krótkofalowe). Fluorescencja chlorofilu a jest to reemisja energii świetlnej zaabsorbowanej przez cząsteczkę chlorofilu a. Utrata energii w postaci fluorescencji jest nieznaczna, stanowi około 5 %. Fluorescencja chlorofilu a w fotosyntezie jest miernikiem energii zaabsorbowanych kwantów światła, która nie została wykorzystana w procesie fotosyntezy, ani nie została wyemitowana w postaci ciepła. Fluorescencja chlorofilu następuje zawsze z niższego stanu wzbudzenia (odpowiadającego stanowi wzbudzenia, który jest osiągany po oświetleniu chlorofilu światłem czerwonym), niezależnie od długości fali światła padającego. Maksimum fluorescencji dla cząsteczek chlorofilu a w roztworze eteru naftowego wynosi 668 nm, przy maksimum absorpcji 663 nm. Przy sprawnym przebiegu reakcji fotosyntetycznych intensywność fluorescencji chlorofilu jest niewielka. Zakłócenia w procesie fotosyntezy (głównie jej fazy fotochemicznej) wywołują wzrost fluorescencji chlorofilu a. Wartość wielu współczynników opisujących fluorescencję chlorofilu a może być mierzona przy użyciu specyficznych urządzeń - fluorymetrów. Uzyskane w wyniku pomiaru wskaźniki fluorescencji informują o przebiegu faz reakcji fotochemicznych.
 maskowanych do tej pory przez zielony barwnik.")
10 Inne barwniki fotosyntetyczne Karotenoidy (karoteny i ksantofile) Karoteny - barwniki pomaraoczowoczerwone, Ksantofile - barwniki żółtopomaraoczowe. Zdjęcia własne AG Pożółkły liśd róży, zanik chlorofilu uwidacznia obecnośd karotenoidów (karotenów i ksantofili) maskowanych do tej pory przez zielony barwnik. To sprawia, że liście jesienią są kolorowe. Inne barwniki fotosyntetyczne, takie jak karotenoidy (karoteny i ksantofile) pełnią pomocniczą rolę w procesie fotosyntezy ponieważ absorbują światło niebieskie i fioletowe. Spełniają one dwie zasadnicze funkcje: (1) uczestniczą w procesie absorpcji światła i transferze energii wzbudzenia elektronowego (przekazują energię stanu wzbudzenia na cząsteczkę chlorofilu, wchodzą w skład anten) (2) zabezpieczają aparat fotosyntetyczny przed skutkami ekspozycji na światło o zbyt wysokim natężeniu (pełnią funkcję ochronną przed procesami fotooksydacjii, na które narażone są głównie nienasycone kwasy tłuszczowe lipidów chloroplastowych). W liściach, żółta barwa karotenoidów jest maskowana przez zieloną barwę barwników chlorofilowych, uwidocznia się natomiast jesienią, kiedy chlorofile są degradowane. Zapamiętaj! Karoteny maja barwę pomarańczowo-czerwoną, ksantofile żółto-pomarańczową.
11 Karotenoidy (karoteny i ksantofile) budowa cząsteczki 1) β-karoten 2) luteina 3) wiolaksantyna Ksantofile zawierają atomy tlenu w cząsteczkach Karotenoidy zbudowane są z jednostek izoprenowych, zawierających pięć atomów węgla, należą do 40-węglowych terpenoidów, tetraterpenów. Karotenoidowy szkielet węglowy C40 budowany jest przez kolejne dodawanie jednostek C5. Z chemicznego punktu widzenia, charakterystyczną cechą karotenoidów jest występowanie dwóch pierścieni jononowych połączonych długim łańcuchem węglowodorowym, w którym występuje układ szeregu sprzężonych wiązań podwójnych. Obecność tych wiązań umożliwia absorpcję światła. Karotenoidy dzielimy na dwie grupy: karoteny (będące węglowodorami), oraz ksantofile (pochodne karotenów zawierające tlen przyłączony do pierścieni jononu w postaci grup: hydroksylowych, epoksydowych, ketonowych, karboksylowych). W błonach tylakoidów roślin wyższych spośród karotenów dominujący jest β-karoten, natomiast spośród ksantofili najczęściej występują: luteina, wiolaksantyna, neoksantyna, zeaksantyna, anteraksantyna i laktukaksantyna. Bardzo istotna fizjologicznie rola ksantofili związana jest z ochroną aparatu fotosyntetycznego przed nadmiernym natężeniem światła. Oparta jest ona na funkcjonowaniu tzw. cyklu ksantofilowego, prowadzącego do rozpraszania nadmiaru energii świetlnej w postaci ciepła (dokładne informacje na ten temat znajdziesz w wykładzie dotyczącym fotosyntezy C3, C4 i CAM).
12 Fikobiliny barwniki występujące u sinic i krasnorostów fikocyjanina fikoerytryna Ostatnią grupą barwników fotosyntetycznych są fikobiliny, występujące u dwóch jednostek systematycznych glonów kryptomonad i krasnorostów oraz cyjanobakterii. Są to pochodne biliwerdyny, produktu oksydacyjnej degradacji hemu. Najczęściej spotykanymi fikobilinami są: fikocyjanobilina i fikoerytrobilina. Fikobiliny jako jedyne barwniki fotosyntetyczne powiązane są z białkami rozpuszczalnymi w wodzie (fikobiliproteinami). Podobnie jak inne barwniki przekazują energię pochłoniętych fotonów na cząsteczki chlorofili. Fikobiliny szczególnie wydajnie absorbują światło czerwone, pomarańczowe, żółte i zielone, czyli w zakresie długości fali częściowo nieabsorbowanym przez chlorofile. Organizmy żyjące w wodach płytkich posiadają zazwyczaj fikobiliny absorbujące światło żółte i czerwone, natomiast żyjące w wodach głębszych światło zielone.
 oraz widma czynnościowego fotosyntezy roślin wyższych (linia ciągła niebieska).")
13 Wiesz już jakie są podstawowe barwniki fotosyntetyczne. Zwróć teraz uwagę na przebieg widm absorpcyjnych chlorofilu a i β-karotenu (linie przerywane zielona i pomarańczowa) oraz widma czynnościowego fotosyntezy roślin wyższych (linia ciągła niebieska). Zapamiętaj, że pokrywają się one w zakresie promieniowania nm. Podobnie, przeanalizuj przebieg widma absorpcyjnego fikoerytrobiliny (linia różowa przerywana) i widma czynnościowego fotosyntezy krasnorostu (linia ciągła czerwona). Zwróć uwagę, że fikoerytrobilina absorbuje światło o długości fali nie absorbowanej przez chlorofile i karotenoidy. Barwa światła wpływa na intensywnośd fotosyntezy. Widma absorpcyjne barwników fotosyntetycznych pokrywają się z widmem czynnościowym fotosyntezy. Widmo czynnościowe fotosyntezy roślin i krasnorostu oraz absorpcja światła dla głównych barwników fotosyntetycznych (chlorofilu a, - karotenu ) oraz fikoerytrobiliny.
14 Z tej części wykładu dowiesz się jak przebiegają reakcje świetlne fotosyntezy i jakie jest ich znaczenie fizjologiczne. Istotą procesu fotosyntezy jest zamiana energii świetlnej w energię wiązao chemicznych (ATP i NADPH), która jest wykorzystana do asymilacji CO 2 Promieniowanie świetlne w zakresie PAR Reakcje świetlne (błony tylakoidów) ATP NADPH Reakcje ciemniowe (stroma chloroplastów) H 2 O O 2 CO 2 CH 2 O PAR (lub PhAR) fotosyntetycznie aktywna radiacja ( nm), zakres światła absorbowany przez barwniki fotosyntetyczne W literaturze występuje skrót PAR lub PhAR z ang. Photosynthetic Active Radiation. Oznacza on światło absorbowane przez barwniki fotosyntetyczne, czyli mieszczące się w zakresie promieniowania nm. Zaabsorbowana przez barwniki fotosyntetycznie aktywna radiacja jest wykorzystywana w chloroplastach do syntezy ATP i NADPH. Oba te związki wytworzone w fazie jasnej (świetlnej, fotochemicznej) fotosyntezy są następnie zużywane do syntezy cukrów z CO 2 i H 2 O, która zachodzi w fazie ciemniowej (biochemicznej) fotosyntezy. Reakcje świetlne fotosyntezy związane są z funkcjonowaniem szeregu przenośników elektronów tworzących łańcuch transportu elektronów. Łańcuchy transportu elektronów funkcjonują na zasadzie przekazywania energii wzbudzenia elektronowego pomiędzy cząsteczkami zgodnie ze wzrastającym potencjałem oksydoredukcyjnym tych cząsteczek, np. w mitochondriach (porównaj informacje z wykładu Oddychanie ). Jak jest w łańcuchu fotosyntetycznym?
15 Produkcja ATP i NADPH w chloroplastach zależy od funkcjonowania łaocucha transportu elektronów Transport elektronów w chloroplastach zgodnie, nie zaś wbrew wzrostowi potencjału redoks przenośników elektronów odbywa się dzięki energii SŁOŃCA. Wzbudzenie centrów aktywnych PSII (P680) i PSI (P700) pozwala na wytworzenie cząsteczek wzbudzonych zdolnych przekazać elektrony dlatego, że ich potencjał redoks zmienia się, odpowiednio: Potencjał oksydoredukcyjny (V) - dla P680* z ok. 1V do ok. - 0,5V dla P700* z ok. 0,5 V do ok. -1,5 V +Potencjał oksydoredukcyjny (V) hν Chlorofil w stanie wzbudzonym * Chlorofil w stanie podstawowym -1,0 silny reduktant -0,5 reduktant 0 0,5 utleniacz NADP + 1,0 Silny utleniacz Fotosystem I (PSI) hν Fotosystem II (PSII) hν W wyniku zaabsorbowania energii świetlnej przez barwniki fotosyntetyczne następuje przeniesienie elektronów z niskiego poziomu energetycznego (potencjał oksydoredukcyjny dodatni: 0,82 V w H 2 O) na wysoki poziom energetyczny (potencjał oksydoredukcyjny ujemny: - 0,42 V w ferredoksynie).
16 Potencjał oksydoredukcyjny utleniający redukujący Niecykliczny transport elektronów w chloroplastach warunkuje powstawanie ATP i NADPH P700* P680* Fd NADP + + H + Phe PQ Kompleks cytochromu bf Reduktaza NADP NADPH P680 LHC II hν ADP + Pi Kompleks Mn 2H 2 O H+ ATP PC P700 LHC I hν Uproszczony schemat transportu niecyklicznego w chloroplastach Energia kwantów światła przekazana do centrum reakcji fotoukładu II (P680) powoduje wybicie elektronu. Elektron jest przekazywany przez cząsteczkę feofityny (Phe) na wolny plastochinon (PQ). Powstały wskutek redukcji plastochinonu, plastochinol przemieszcza się w błonie tylakoidu do kompleksu cytochromowego b 6 f. W obrębie tego kompleksu zachodzi cykl Q w wyniku którego dodatkowe protony (H + ) przemieszczane są ze stromy chloroplastów do wnętrza tylakoidów. Kompleks cytochromowy b 6 f przekazuje elektron na kolejny przenośnik plastocyjaninę (PC). Odbiorcą elektronów od plastocyjaniny jest fotoukład I (P700), z którego centrum reakcji uprzednio zostały wybite elektrony. Elektron wybity z centrum reakcji fotoukładu I przekazywany jest na cząsteczkę NADP +, co prowadzi do wytworzenia formy zredukowanej NADPH. W przekazaniu elektronu na cząsteczkę NADP + bierze udział ferredoksyna (Fd). Miejsce po elektronie oderwanym z centrum reakcji fotoukładu II (P680) zapełniane jest przez elektron powstający w wyniku fotolizy wody. Reakcja ta jest przeprowadzana przez kompleks rozkładający wodę centrum manganowe. Zawiera ono jon wapnia i cztery jony manganu. Centrum manganowe w formie zredukowanej utlenia dwie cząsteczki wody, aby utworzyć jedna cząsteczkę tlenu, po oderwaniu 4 elektronów następuje rozszczepienie 2 cząsteczek wody na 4 protony i cząsteczkę tlenu.
17 Potencjał oksydoredukcyjny Cykliczny transport elektronów dostarcza tylko ATP utleniający redukujący P700* Fd PQ Kompleks cytochromu bf ADP + Pi ATP PC P700 LHC I hν W sytuacji zwiększonego zapotrzebowania chloroplastu na ATP w stosunku do NADPH uruchomiony zostaje cykliczny transport elektronów, w którym uczestniczy tylko PS I (P700). Cyklicznemu przepływowi elektronów towarzyszy przenoszenie protonów przez błonę na kompleksie cyt b 6 f. Powstający gradient stężenia protonów jest siłą napędową fosforylacji cyklicznej. Podczas cyklicznego transportu elektronów w łańcuchu chloroplastowym wytwarzane jest ATP, natomiast nie powstaje NADPH.
18 Skąd się bierze gradient stężenia protonów warunkujący wytwarzanie ATP przez syntezę ATP? Transportowi elektronów z cząsteczki wody na NADP + towarzyszy tworzenie gradientu stężenia protonów w poprzek błony tylakoidu. Dochodzi do alkalizacji stromy i zakwaszenia wnętrza pęcherzyka tylakoidy. Protony gromadzące się wewnątrz pęcherzyka tylakoidu pochodzą z reakcji fotolizy wody oraz są uwalniane w procesie utleniania plastochinolu przez kompleks cytochromowy b 6 f. Spadek stężenia protonów w stromie jest wynikiem protonacji zredukowanych cząsteczek plastochinonu w miejscu Q b, aktywności cyklu Q oraz tworzenia cząsteczek NADPH. Gradient stężenia protonów w poprzek błony tylakoidów odpowiada za syntezę cząsteczek ATP w procesie fosforylacji fotosyntetycznej Stroma ph alkaliczne Światło tylakoidu ph kwaśne błona tylakoidu H + H + H + H + H + ADP + Pi H + ATP Syntaza ATP

19 Fosforylacja fotosyntetyczna prowadzi do syntezy ATP z ADP i fosforanu nieorganicznego Pi przy udziale kompleksu syntazy ATP Dittrich i wsp Biophysical Journal 87: Enzymem odpowiadającym za syntezę ATP w procesie fosforylacji fotosyntetycznej jest synteza ATP. Syntaza ATP nazywana jest też czynnikiem sprzęgającym transport elektronów z fosforylacją. Część CF O syntazy ATP, przez którą przepływa strumień protonów, zlokalizowana jest w wewnętrznej błonie chloroplastu. Część CF 1, syntezująca ATP z ADP i nieorganicznego fosforanu (Pi) znajduje się na zewnątrz błony w stromie. Protony (H + ) przepływające kanałem utworzonym przez białka części F O powodują, iż podjednostki CF 1 obracają się względem siebie. Prowadzi to do zmian konformacyjnych niektórych podjednostek CF 1 i umożliwia wiązanie ADP i Pi oraz uwalnianie powstającej cząsteczki ATP. Zapamiętaj! W wyniku: uwalniania protonów z rozkładu wody wewnątrz tylakoidu (lumen), pobierania protonów podczas redukcji NADP + w stromie chloroplastu transportu protonów w cyklu Q, ze stromy do wnętrza tylakoidu, powstaje gradient protonowy różnica stężeń protonów na zewnątrz i wewnątrz tylakoidu. Gradient protonowy jest wykorzystywany przez kompleks syntazy ATP do wytwarzania drugiego produktu fazy jasnej fotosyntezy ATP.
20 Pseudocykliczny transport elektronów- reakcja Mehlera Transport elektronów na O 2 powoduje tworzenie ATP (bo generuje gradient H + w poprzek błony tonoplastu) i prowadzi do powstawania anionorodnika ponadtlenkowego (O 2 - ) należącego do jednej z reaktywnych form tlenu (ROS) (o tym dowiesz się więcej na wykładzie o stresie oksydacyjnym) PSII 2H 2 O 2H +, 4e -, O 2 4e -, 4H + O 2 PSI 2O 2-2H + SOD APOX H 2 O 2 O 2 Przy znacznym natężeniu światła w chloroplastach zużywany jest nie tylko CO 2, ale także O 2. Tlen może być redukowany do nadtlenku wodoru H 2 O 2 z udziałem fotoukładu I (PSI). Przeniesienie elektronów na O 2 zamiast na NADP + nosi nazwę reakcji Mehlera. Tworzenie reaktywnych form tlenu O - 2 oraz H 2 O 2 ma miejsce przede wszystkim w chloroplastach ze zbyt małą ilością NADP +. Zbyt małe stężenie NADP + występuje głównie podczas oświetlania roślin światłem o wysokim natężeniu, kiedy fotosyntetyczny transport elektronów przebiega bardzo wydajnie i niemal cała pula NADP + obecna w chloroplastach pozostaje w stanie zredukowanym. W takiej sytuacji elektrony mogą być przekazywane na tlen, co prowadzi do powstawania gradientu protonów i syntezy ATP, zapobiegając jednocześnie uszkodzeniu fotoukładów. Ze względu na syntezę ATP przy braku syntezy NADPH taki transport elektronów jest określany jako pseudocykliczny transport elektronów. Badania wykazały, że poza ochroną fotoukładów transport elektronów na O 2 reguluje interakcje pomiędzy cyklicznym, a niecyklicznym transportem elektronów. Chociaż stężenia tlenu nie wymienia się zwykle jako czynnika wpływającego na natężenie fotosyntezy, to w sytuacji intensywnego oświetlenia w wyniku fotolizy wody stężenie tlenu w komórkach jest na tyle duże, że reakcja Mehlera ma znaczący udział w wydajności całego procesu fotosyntezy.

21 Wiesz już jakie reakcje prowadzą do tworzenia ATP i NADPH w fazie świetlnej fotosyntezy. Spróbuj jeszcze raz przeanalizować schemat transportu elektronów w łańcuchu chloroplastowym, zwracając uwagę na powstający w poprzek błony tylakoidu gradient stężenia protonów. Zapamiętaj że, poszczególne kompleksy łańcucha transportu elektronów nie działają jako pompy protonowe. Gradient stężenia protonów będący warunkiem powstania ATP w reakcji zależnej od syntazy ATP jest wynikiem procesu fotolizy wody oraz funkcjonowania cyklu plastochinonu. Schemat fazy jasnej fotosyntezy. Użyte skróty: P-680 centrum reakcji fotoukładu II; P-700 centrum reakcji fotoukładu I; Phe feofityna; K Mn kompleks rozkładający wodę; Q A Q B plastochinon połączony z białkiem; PQ wolny plastochinon; b 6w wysokopotencjałowy chem cytochromu b 6 ; b 6n niskopotencjałowy hem cytochromu b 6 ; FeS centrum żelazowo-siarkowe białka Rieskego; PC plastocyjanina; A cząsteczka chlorofilu; A 1 witamina K 1 ; F x centra żelazowosiarkowe; F D ferredoksyna; FNR reduktaza ferredoksyna-nadp Zapamiętaj: Faza świetlna fotosyntezy ma na celu wytworzenie ATP i NADPH ATP w chloroplastach jest syntetyzowane w procesie: fosforylacji niecyklicznej - podczas przepływu elektronów z PSII na PSI fosforylacji cyklicznej podczas przepływu elektronów ze zredukowanej ferredoksyny przez układ cytochromów z powrotem na PSI. (Reakcja ta zachodzi przy wysokim zapotrzebowaniu na ATP w stosunku do NADPH ponieważ nie powstaje wówczas NADPH) fosforylacji pseudocykliczna- Pierwotnym produktem tej reakcji jest anionorodnik ponadtlenkowy ulegający dysmutacji do H 2 O 2. Transport pseudocykliczny elektronów dostarcza ATP i usuwa O 2 z chloroplastu.
22 Poszczególne kompleksy białkowe wchodzące w skład łańcucha transportu elektronów są nierównomiernie rozmieszczone w błonach tylakoidów gran i stromy. Heterogenność błon tylakoidów zapewnia równowagę miedzy optymalnym wykorzystaniem energii absorbowanych fotonów, a koniecznością jej rozproszenia w sytuacji nadmiaru światła. Kompleksy białkowe (fotosystemy, syntaza ATP i cyt b 6 f) są nierównomiernie rozmieszczone w błonach tylakoidów gran i stromy Błony tylakoidowe gran są bogate w PSII, błony tylakoidowe stromy w PSI. 85 % PSII znajduje się w błonach zespolonych gran, a 15 % w tylakoidach stromy. Około 65 % cząsteczek PSI znajduje się w błonach tylakoidów stromy, pozostałe 35 % w brzegach gran. Większość kompleksów cytochromu b6f znajduje się w błonach zespolonych gran. Sądzi się, że nierównomierne rozmieszczenie kompleksów w obrębie błon jest związane z realizacją niecyklicznego transportu elektronów wyłącznie w obrębie gran, a transportu cyklicznego, wyłącznie w obrębie tylakoidów stromy.
23 Dla zainteresowanych. Ciekawostka. Selektywne działanie herbicydów (środki chwastobójcze) jest związane m.in. ze strukturą cząsteczki. W przypadku wielu powszechnie stosowanych herbicydów docelowym miejscem działania są komponenty chloroplastowego łańcucha transportu elektronów. Niektóre herbicydy hamują przebieg świetlnej fazy fotosyntezy herbicyd Pierwotne miejsce działania Hamowany szlak metaboliczny DCMU Atazyna Bromoksynil Parakwat (metylwiologen) Miejsce wiązania Qb w białku D1 kompleksu PS II Miejsce wiązania Qb w białku D1 kompleksu PS II Miejsce wiązania Qb w białku D1 kompleksu PS II Miejsce akceptorowe Fd w kompleksie PSI Trsansport elektronów i wydzielanie O2 w PS II Transport elektronów w PSII Transport elektronów w PSII Redukcja NADP i transport elektronów w PSI fosfinotrycyna Syntaza glutaminy Biosynteza glutaminy, asymilacja CO2
24 Wykorzystanie produktów fazy fotochemicznej fotosyntezy w metabolizmie azotu i siarki W chloroplastach oprócz reakcji asymilacji CO 2 zlokalizowanych jest wiele innych reakcji zużywających produkty fazy fotochemicznej fotosyntezy. Należą do nich przede wszystkim reakcje związane z asymilacja azotu i siarki. Chloroplasty są jedynym miejscem w komórce, gdzie zachodzi redukcja azotanów i siarczanów, a zatem w chloroplastach wytwarzane są związki organiczne zawierające azot i siarkę. Siarka jest pobierana przez rośliny głównie jako jony SO Redukcja tych anionów odbywa się głównie w chloroplastach z wykorzystaniem siły redukcyjnej powstającej w fazie świetlnej fotosyntezy, konkurując o elektrony z NADP+ i z reduktazą azotynową, uczestniczącą w redukcji azotynów. Redukcja anionu SO 4 2- pobranego z podłoża, jest procesem bardzo energochłonnym. Energia wydatkowana podczas redukcji jony SO 4 2- do S 2 (zmiana stopnia utlenienia z +6 do -2) i syntezy cysteiny wynosi 732 kj-mol. W etapie aktywacji, powstaje z udziałem ATP adenozyno-5-fosfosiarczan (APS ) i pirofosforan (PPi), rozpadający się w następnej reakcji na fosforan: S0 4 + ATP APS + PPi + H 2 0 Zaktywowany siarczan jest przenoszony na nośnik z grupą tiolową (RSH), którym może być glutation. Powstaje wówczas tiosulfonian, redukowany do tiosiarczku, a jednocześnie wydziela się AMP. W reakcji tej donorem elektronów jest ferredoksyna (Fd). Następnie zachodzi synteza cysteiny, po przeniesieniu grupy tiolowej na O-acetyloserynę. W syntezie sulfolipidów uniwersalnym donorem reszt SO 4 jest 5'-fosfosiarczan 3'-fosfoadenozyny. Redukcja azotanów (NO 3 - ) do azotynów (NO 2 - ) zachodzi na terenie cytozolu. Redukcja azotynów do jonów amonowych (NH 4 + ) w stromie chloroplastów. Donorem elektronów w tej reakcji jest zredukowana ferredoksyna. Powstający amoniak jest natychmiast wbudowany w strukturę amidową glutaminy w reakcji katalizowanej przez syntetazę glutaminową jeden z enzymów cyklu GS-GOGAT (sytetaza glutaminowa - syntaza glutaminianowa). Reakcje te zachodzą na terenie stromy chloroplastów i wymagają obecności NADPH lub zredukowanej ferredoksyny.

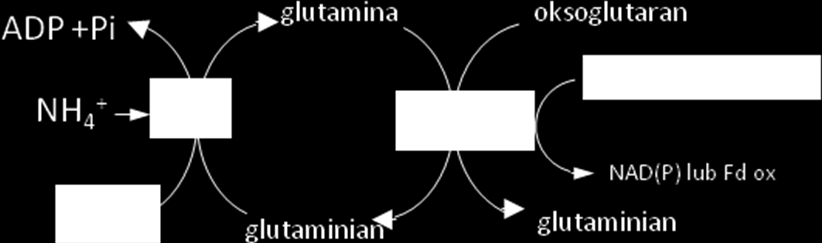
25
 zachodzącej w stromie chloroplastów.")
26 Wiesz już jakie procesy przebiegają w czasie fazy świetlnej w tylakoidach chloroplastów. Wiesz, że produktem tego etapu fotosyntezy jest siła redukcyjna w postaci ATP i NADPH. Teraz dowiesz się do czego produkty te są zużywane w fazie ciemniowej fotosyntezy (niezależnej od światła) zachodzącej w stromie chloroplastów. Przemiany fazy ciemniowej od nazwisk ich odkrywców nazywamy reakcjami cyklu Calvina- Bensona.

27 Przebieg fazy biochemicznej fotosyntezy Cykl Calvina-Bensona commons.wikimedia.org/wiki/file:cykl_calvina.svg W cyklu Calvina- Bensona produkty reakcji świetlnych fotosyntezy, ATP i NADPH zostają zużyte do wbudowania CO 2 w związki organiczne. W cyklu tym pierwotnym akceptorem CO 2 jest 5 węglowy związek bisfosforybuloza, a pierwszym trwałym produktem 3 węglowy kwas 3-fosfoglicerynowy. Ten typ fotosyntetycznego włączania CO 2 nazywamy C3. W cyklu Calvina-Bensona wyróżnia się trzy fazy: Faza karboksylacyjna, w której CO 2 wiązany jest do rybulozo-1,5-bisfosforanu. Ta reakcja jest katalizowana przez kluczowy enzym metabolizmu fotosyntetycznego karboksylazę/oksygenazę rybulozo 1,5 bisfosforybulozy (Rubisco -najbardziej rozpowszechnione białko biosfery). W efekcie tej reakcji powstaje nietrwały związek 6 węglowy, a następnie dwie cząsteczki 3-fosfoglicerynianu. Faza redukcyjna, w której 3-fosfoglicerynian ulega fosforylacji do 1,3-fosfoglicerynianu ze zużyciem ATP powstałego w fazie świetlnej fotosyntezy. Następnie 1,3-fosfoglicerynian jest przekształcany do fosfotriozy: aldehydu 3-fosfoglicerynowego. Na tym etapie zużywane jest NADPH powstałe w fazie świetlnej. Aldehyd 3-fosfoglicerynowy służy do syntezy heksoz. Faza regeneracyjna, w której z cząsteczek aldehydu 3-fosfoglicerunowego zostaje odtworzony akceptor CO 2 rybulozo-1,5-bisfosforan. Również na tym etapie zużywane jest ATP.

28 Za kluczową reakcję wiązania CO 2 w cyklu Calvina -Bensona odpowiada karboksylaza rybulozo-1,5-bisfosforanu (Rubisco). Szybkość reakcji cyklu Calvina-Bensona jest między innymi regulowana przez: Jony Mg - są aktywatorami Rubisco. Nieorganiczny fosforan (Pi) stały eksport fosfotrioz z chloroplastów musi być równoważony napływem fosforanu nieorganicznego do chloroplastów. Stężenie metabolitów pośrednich cyklu Calvina-Bensona warunkuje fazę regeneracji rybulozo-1,5-bisfosforanu. Światło - utrzymuje w stanie zredukowanym grupy sulfhydrylowe niektórych enzymów cyklu Calvina- Bensona stymulując ich aktywność.
29 Fotooddychanie, fotorespiracja to proces biochemiczny zachodzący na świetle w fotosyntetyzujących tkankach roślinnych, objawiający się pobieraniem O 2 i wydzielaniem CO 2. Proces ten nie dostarcza energii metabolicznej, a zużywa ją. Fotooddychanie wiąże się z oksygenacyjną aktywnością Rubisco. CO 2 i O 2 wiążą się do tego samego miejsca katalitycznego enzymu. Aktywnośd karboksylacyjna Rubisco 3-fosfoglicerynian 3-fosfoglicerynian rybulozo-1,5- bisfosforan Nietrwały sześciowęglowy produkt Aktywnośd oksygenacyjna Rubisco 3-fosfoglicerynian 2-fosfoglikolan

30 Fotooddychanie wymaga współdziałania trzech kompartmentów komórkowych: chloroplastów, peroksysomów i mitochondriów Schemat obrazujący przemiany zachodzące podczas fotooddychania. Rubisco karboksylaza/oksygenaza rybulozo-1,5-bisfosforanu, Phos fosfataza, OG - oksydaza glikolanowa, ATaminotransferaza, GDC+SHMT-dekarboksylaza glicyny i hydroksymetylotransferaza seryny, PyrR-reduktaza pirogronianowa, GK-kinaza glicerynianowa, GOGAT syntaza glutaminianowa (NADPH), GS-syntaza glutaminowa Proces fotooddychania związany jest karboksylacyjną i oksygenacyjną aktywnością Rubisco, enzymu odpowiedzialnego zarówno za przyłączenie do rybulozo-1,5-bisfoforanu (RuBP) cząsteczki CO 2, jak i cząsteczki O 2. CO 2 i O 2 konkurują o miejsce katalityczne Rubisco. W wyniku przyłączenia O 2 do RuBP powstaje jedna cząsteczka 3-fosfoglicerynianu (jak w fazie ciemnej fotosyntezy) oraz jedna cząsteczka 2-fosfoglikolanu, pierwszego (dwuwęglowego; C2) produktu fotooddychania. Te reakcje zachodzą w chloroplastach. Dalsze etapy przemian zachodzą w peroksysomach i mitochondriach, a następnie ponownie w chloroplastach. Chloroplasty, peroksysomy i mitochondria w komórkach liścia rośliny C3, zlokalizowane są blisko siebie, stykają się błonami, co ułatwia transport metabolitów w procesie fotoodychania. Powstający w chloroplastach 3-fosfoglikolan ulega defosforylacji i przenoszony jest do peroksysomów. Przy udziale oksydazy glikolanowej przekształcany jest do glioksalanu. Glioksalan ulega transaminacji w dwóch reakcjach przeprowadzanych przez aminotransferazę glutaminianową i aminotransferazę serynową, w wyniku których powstaje glicyna. Glicyna transportowana jest do mitochondriów i przy udziale kompleksu enzymatycznego dekarboksylazy glicyny (GDC) oraz hydroksymetylotrasferazy seryny (SHMT) przekształcana do seryny z wydzieleniem cząsteczki CO 2, NH 3, oraz NADH. Powstała w mitochondriach seryna transportowana jest do peroksysomów i przekształcana przy udziale aminotransferazy serynowej do kwasu hydroksypirogronowego. Kwas ten ulega redukcji do kwasu glicerynowego przy udziale reduktazy hydroksypirogronianowej. Produkt
31 reakcji przenoszony jest do chloroplastów i może służyć do odtworzenia cząsteczki rybulozo- 1-5-bisfosforanu. NADH produkowany przy dekarboksylacji glicyny może być transportowany do cytozolu lub utleniany w mitochondriach. Fotooddychanie jest charakterystyczne dla roślin o fotosyntezie typu C3. Prowadzi do uwolnienia wcześniej związanego CO 2, zmniejsza zatem wydajność fotosyntezy i przyrost biomasy tych roślin. Fizjologiczny sens fotooddychania nie jest wyjaśniony. Proces ten może chronić aparat fotosyntetyczny przed uszkodzeniem w warunkach gdy reakcje świetlne zachodzą z dużą wydajnością, a szybkość reakcji ciemniowych jest limitowana przez niską zawartość CO 2 w powietrzu. Proces ten zużywa nadmiar nagromadzonego NADPH. Wyniki badań prowadzone na mutantach nie zawierających któregoś z enzymów cyklu fotoodechowego wykazały, że funkcjonowanie tego cyklu jest konieczne do przeżycia w mieszaninie gazów, jaką mamy obecnie w atmosferze ziemskiej. Regulacja fotooddychania i znaczenie tego procesu dla roślin Wpływ temperatury na fotosyntezę netto i fotooddychanie liści fasoli Temperatura [ C] Fotosynteza (PN) [µmol CO 2 m -2 s- -1 ] Fotooddychanie (PR) [µmol CO 2 m -2 s- -1 ] PR/PN (%) , ,9 5, ,0 8, ,5 62 Intensywność fotoodychania rośnie przy podwyższaniu temperatury, z uwagi na zmianę właściwości kinetycznych Rubisco rośnie aktywność oksygenacyjna Rubisco. Przy wzroście temperatury rośnie stosunek intensywności fotooddychania do intensywności fotosyntezy (PR/PN). W temperaturze optymalnej dla procesu fotosyntezy wynosi on około 25 %.
32 Intensywność fotoodychania zależy także od stosunku stężenia CO 2 i O 2. W atmosferze o normalnym składzie, zwierającej 0,03 % CO 2 i 21 % O 2 karboksylacja 2-3 krotnie przewyższa proces utleniania. Zrównanie stężenia obu gazów prowadziłoby do znacznego wzrostu szybkości karboksylacji. W naturze w warunkach intensywnej fotosyntezy może dochodzić do miejscowego spadku stężenia CO 2 i znacznej stymulacji fotoodychania. Wiesz już jakie reakcje zachodzą w fazie ciemniowej fotosyntezy. Wiesz też jakie znaczenie dla metabolizmu roślin ma aktywność karboksylacyjna lub oksygenacyjna Rubisco. Chloroplasty dzięki reakcjom fotosyntezy dostarczają roślinie cukrów, które eksportowane są z chloroplastów w postaci fosforanów trioz (fosforan dihydroksyacetonu, aldehyd-3-fosfoglicerynowy i 3-fosfoglicerynian). W transporcie tych związków ze stromy chloroplastów do cytozolu uczestniczy przenośnik fosforanowy. Fosforany trioz są transportowane z chloroplastu na zasadzie antysportu z fosforanem nieorganicznym, co pozwala na stałe uzupełnianie ubytku fosforanu w stromie chloroplastów. Zapamiętaj, że produktami fotosyntezy w cyklu Calvina-Bensona są przede wszystkim fosforany trioz. W wyniku fotosyntetycznego włączania CO 2 w chloroplastach na terenie cytozolu syntetyzowana jest sacharoza. Synteza sacharozy zachodzi w cytozolu z fosforanów heksoz powstających z fosfotrioz eksportowanych z chloroplastu przez przenośnik fosforanowy. Regulacja syntezy skrobi i sacharozy w liściach chloroplast cytosol Cykl Calvina Fosforany trioz Pi Fosforany trioz Fosforany heksoz skrobia sacharoza Uproszczony schemat syntezy skrobi i sacharozy w liściach. Kolorem niebieskim zaznaczono przenośnik fosforanowy.
33 Gdy szybkość włączania CO 2 w chloroplastach przekracza szybkość z jaką fosfortiozy są przekształcane w sacharozę w cytoplazmie rozpoczyna się synteza skrobi asymilacyjnej. Skrobia asymilacyjna powstaje w chloroplastach z nadmiaru fosfotrioz powstających w fazie ciemniowej fotosyntezy. Skrobia asymilacyjna jest więc gromadzona w chloroplastach na świetle. Skrobie tę wykorzystuje roślina w czasie na potrzeby energetyczne. Skrobia nagromadzona w chloroplastach jest rozkładana do cukrów lub fosforanów cukrów i zużywana w okresie ciemności lub przy ograniczonej fotosyntezie. Regulacja syntezy skrobi i sacharozy w liściach chloroplast cytosol Cykl Calvina Fosforany trioz Pi Fosforany trioz skrobia sacharoza Uproszczony schemat syntezy skrobi i sacharozy w liściach. Kolorem niebieskim zaznaczono przenośnik fosforanowy. Synteza skrobi i sacharozy są procesami konkurencyjnymi, gdy aktywowany jest szlak syntezy sacharozy zahamowany jest szlak biosyntezy skrobi i odwrotnie. Ważną rolę w regulacji obu szlaków odgrywa stężenie fosforanu nieorganicznego Pi i fosfotrioz w chloroplastach i cytozolu. Podsumowanie wykładu Znasz podstawowe barwniki fotosyntetyczne i wiesz gdzie one występują oraz jaką pełnią funkcję. Wiesz jak zachodzi faza świetlna i ciemniowa fotosyntezy. Jakie produkty powstają w wyniku fazy świetlnej i ciemniowej fotosyntezy, oraz jakie jest ich wykorzystanie w komórce roślinnej. Omówiony tu metabolizm fotosyntetyczny jest charakterystyczny dla
34 roślin o fotosyntezie typu C3. W kolejnym wykładzie ( Fotosynteza typu C3, C4 i CAM ) dowiesz się jakie są modyfikacje procesu fotosyntezy i jakie są ich skutki. Literatura podstawowa: Fizjologia roślin, red. J. Kopcewicz, S. Lewak, rozdz. Fotosynteza i chemosynteza. Wydawnictwo Naukowe PWN Warszawa 2002 Fizjologia roślin wprowadzenie red. J. Kopcewicz, S. Lewak, Rozdz. 2.3 Fotosynteza i anabolizm. Wydawnictwo Naukowe PWN Warszawa 2009 Fizjologia roślin red. M. Kozłowska, rozdz. Fotosynteza i aktywność fotosyntetyczna roślin. PWRiL, Poznań 2007 Fizjologia roślin, A. Szwejkowska, rozdz. Fotosynteza Wydawnictwo Naukowe UAM, Poznań 1997 Fizjologia plonowania roślin. R.J. Górecki, S. Grzesiuk, rozdz. Fotosynteza. Wydawnictwo UWM, Olsztyn 2002 Literatura uzupełniająca Fotosynteza, Hall D.O., Rao K.K. Wyd. Naukowo-Techniczne, Warszawa Kalaji M.H., Łoboda T Fluorescencja chlorofilu w badaniach stanu fizjologicznego roślin. Wydawnictwo SGGW, Warszawa Podstawy biologii komórki roślinnej, red. A. Woźny, J. Michejda, L. Ratajczak. Rozdział Plastydy. Wydawnictwo naukowe UAM, Poznań Bioenergetyka 2. Nicholls D.G., Ferguson S.J. Wyd Naukowe PWN, Warszawa Video Na stronie Virtual Cell Animation Collection znajdziesz animowane filmy tłumaczące zjawisko fotosyntezy (niestety po angielsku), Polecam filmy zatytułowane: Photosynthesis, Photosystem II Na tej stronie znajdziesz animacje obrazujące reakcje zachodzące w cykl Calvina Ciekawostki
35 Na tej stronie znajdziesz przystępnie omówione zagadnienia związane z fotosyntezą i przepisy na proste doświadczenia opisujące właściwości barwników fotosyntetycznych Sztuczna fotosynteza Zadanie rachunkowe 1. Przy użyciu analizatora CO 2 w podczerwieni zmierzono fotosyntezę liścia fasoli. Powierzchnia liścia umieszczona w kamerze wynosiła 8 cm 2, objętość molowa układu do pomiaru fotosyntezy wynosiła 0,0015 mola powietrza, pomiar wykonywano w zakresie atmosferycznego stężenia CO 2, czas w którym doszło do zmiany stężenia CO 2 o 25 ppm wynosił 1 min i 5 s. Oblicz intensywność fotosyntezy liścia fasoli wyrażając ją w mol CO 2 m -2 s -1. Pytania problemowe 1. Zastanów się jakie czynniki środowiskowe sprzyjają intensywnemu fotoddychaniu i dlaczego? 2. Jaka jest zależność pomiędzy widmem absorpcyjnym barwników fotosyntetycznych a widmem działania fotosyntezy? Czy starzenie się liści ma wpływ na intensywność fazy fotochemicznej fotosyntezy? 3. Dlaczego niecykliczny transport elektronów w łańcuchu chloroplastowym przedstawia się jako tzw. schemat Z? Z czego wynika jego zygzakowaty kształt? 4. W słoneczny dzień, można niekiedy dostrzec pęcherzyki gazu uwalniane przez rośliny wodne. O czym świadczy ich obecność i jaki zawierają gaz?
Proplastydy. Plastydy. Chloroplasty biogeneza. Plastydy
 Plastydy Proplastydy rodzina organelli powstających w toku ontogenezy rośliny drogą różnicowania form prekursorowych proplastydów w tkankach merystematycznych sferyczne; 0.5-2 μm otoczka (2 błony) stroma
Plastydy Proplastydy rodzina organelli powstających w toku ontogenezy rośliny drogą różnicowania form prekursorowych proplastydów w tkankach merystematycznych sferyczne; 0.5-2 μm otoczka (2 błony) stroma
wielkość, kształt, typy
 Mitochondria 0,5-1µm wielkość, kształt, typy 1-7µm (10µm) Filmowanie poklatkowe (w mikroskopie fluorescencyjnym) sieci mitochondrialnej w komórkach droŝdŝy (krok czasowy 3 min) Mitochondria liczebność,
Mitochondria 0,5-1µm wielkość, kształt, typy 1-7µm (10µm) Filmowanie poklatkowe (w mikroskopie fluorescencyjnym) sieci mitochondrialnej w komórkach droŝdŝy (krok czasowy 3 min) Mitochondria liczebność,
Bliskie spotkania z biologią FOTOSYNTEZA. dr inż. Magdalena Kulczyk-Skrzeszewska Katedra Mykologii i Mykoryzy Instytut Biologii Środowiska
 Bliskie spotkania z biologią FOTOSYNTEZA dr inż. Magdalena Kulczyk-Skrzeszewska Katedra Mykologii i Mykoryzy Instytut Biologii Środowiska FOTOSYNTEZA SENS BIOLOGICZNY Podstawowy proces zapewniający utrzymanie
Bliskie spotkania z biologią FOTOSYNTEZA dr inż. Magdalena Kulczyk-Skrzeszewska Katedra Mykologii i Mykoryzy Instytut Biologii Środowiska FOTOSYNTEZA SENS BIOLOGICZNY Podstawowy proces zapewniający utrzymanie
Copyrights LCE LOGOS Centrum Edukacyjne Fotosynteza
 Fotosynteza Fotosynteza jest procesem anabolicznym, czyli z prostych substancji pobranych z otoczenia pod wpływem energii syntetyzowane są złożone substancje organiczne (głównie cukry). Energią niezbędną
Fotosynteza Fotosynteza jest procesem anabolicznym, czyli z prostych substancji pobranych z otoczenia pod wpływem energii syntetyzowane są złożone substancje organiczne (głównie cukry). Energią niezbędną
Plastydy. Proplastydy
 Plastydy rodzina organelli powstających w toku ontogenezy rośliny drogą róŝnicowania form prekursorowych proplastydów w tkankach merystematycznych Proplastydy sferyczne; 0.5-2 µm otoczka (2 błony) ziarna
Plastydy rodzina organelli powstających w toku ontogenezy rośliny drogą róŝnicowania form prekursorowych proplastydów w tkankach merystematycznych Proplastydy sferyczne; 0.5-2 µm otoczka (2 błony) ziarna
Na początek przyjrzymy się więc, jak komórka rośliny produkuje ATP, korzystając z energii światła w fazie jasnej fotosyntezy.
 Fotosynteza jako forma biosyntezy Bogactwo molekuł biologicznych przedstawionych w poprzednim rozdziale to efekt ich wytwarzania w komórkach w wyniku różnorodnych powiązanych ze sobą procesów chemicznych.
Fotosynteza jako forma biosyntezy Bogactwo molekuł biologicznych przedstawionych w poprzednim rozdziale to efekt ich wytwarzania w komórkach w wyniku różnorodnych powiązanych ze sobą procesów chemicznych.
FOTOSYNTEZA I CHEMOSYNTEZA. Prof. dr hab. Barbara Kieliszewska-Rokicka Instytut Biologii Środowiska
 FOTOSYNTEZA I CHEMOSYNTEZA Prof. dr hab. Barbara Kieliszewska-Rokicka Instytut Biologii Środowiska Fotosynteza i chemosynteza są to procesy, w których organizmy żywe przekształcają CO 2 w związki organiczne
FOTOSYNTEZA I CHEMOSYNTEZA Prof. dr hab. Barbara Kieliszewska-Rokicka Instytut Biologii Środowiska Fotosynteza i chemosynteza są to procesy, w których organizmy żywe przekształcają CO 2 w związki organiczne
Plastydy. Proplastydy
 Plastydy rodzina organelli powstających w toku ontogenezy rośliny drogą różnicowania form prekursorowych proplastydów w tkankach merystematycznych Proplastydy sferyczne; 0.5-2 μm otoczka (2 błony) ziarna
Plastydy rodzina organelli powstających w toku ontogenezy rośliny drogą różnicowania form prekursorowych proplastydów w tkankach merystematycznych Proplastydy sferyczne; 0.5-2 μm otoczka (2 błony) ziarna
Mitochondria - siłownie komórki
 Transformatory energii (mitochondria i chloroplasty) ewolucja eukariontów endosymbioza prakomórki eukariotycznej z prabakterią purpurową lub pracyjanobakterią Pochodzenie mitochondriów i chloroplastów
Transformatory energii (mitochondria i chloroplasty) ewolucja eukariontów endosymbioza prakomórki eukariotycznej z prabakterią purpurową lub pracyjanobakterią Pochodzenie mitochondriów i chloroplastów
Transformatory energii (mitochondria i chloroplasty) Pochodzenie mitochondriów i chloroplastów
 Pochodzenie mitochondriów i chloroplastów") Transformatory energii (mitochondria i chloroplasty) ewolucja eukariontów endosymbioza prakomórki eukariotycznej z prabakterią purpurową lub pracyjanobakterią Pochodzenie mitochondriów i chloroplastów
Transformatory energii (mitochondria i chloroplasty) ewolucja eukariontów endosymbioza prakomórki eukariotycznej z prabakterią purpurową lub pracyjanobakterią Pochodzenie mitochondriów i chloroplastów
FOTOSYNTEZA. Czynniki wpływające na intensywnośd fotosyntezy: 1)Wewnętrzne:
Wewnętrzne:") FOTOSYNTEZA Fotosynteza (photosynthesis )-proces biologiczny polegający na pochłanianiu energii światła słonecznego i przekształcaniu jej w energię chemiczną cząstek organicznych ( np.węglowodanów) syntezowanych
FOTOSYNTEZA Fotosynteza (photosynthesis )-proces biologiczny polegający na pochłanianiu energii światła słonecznego i przekształcaniu jej w energię chemiczną cząstek organicznych ( np.węglowodanów) syntezowanych
METABOLIZM. Zadanie 1. (3 pkt). Uzupełnij tabelę, wpisując w wolne kratki odpowiednio produkt oddychania tlenowego i produkty fermentacji alkoholowej.
. Uzupełnij tabelę, wpisując w wolne kratki odpowiednio produkt oddychania tlenowego i produkty fermentacji alkoholowej.") Zadanie 1. (3 pkt). Uzupełnij tabelę, wpisując w wolne kratki odpowiednio produkt oddychania tlenowego i produkty fermentacji alkoholowej. Zadanie 3. (3 pkt). Schemat mechanizmu otwierania aparatu szparkowego.
Zadanie 1. (3 pkt). Uzupełnij tabelę, wpisując w wolne kratki odpowiednio produkt oddychania tlenowego i produkty fermentacji alkoholowej. Zadanie 3. (3 pkt). Schemat mechanizmu otwierania aparatu szparkowego.
Komórka organizmy beztkankowe
 Grupa a Komórka organizmy beztkankowe Poniższy test składa się z 12 zadań. Przy każdym poleceniu podano liczbę punktów możliwą do uzyskania za prawidłową odpowiedź. Za rozwiązanie całego testu możesz otrzymać
Grupa a Komórka organizmy beztkankowe Poniższy test składa się z 12 zadań. Przy każdym poleceniu podano liczbę punktów możliwą do uzyskania za prawidłową odpowiedź. Za rozwiązanie całego testu możesz otrzymać
Metabolizm komórkowy i sposoby uzyskiwania energii
 Metabolizm komórkowy i sposoby uzyskiwania energii Metabolizm całokształt reakcji chemicznych i związanych z nimi przemian energii zachodzący w komórkach. Podstawa wszelakich zjawisk biologicznych. Metabolizm
Metabolizm komórkowy i sposoby uzyskiwania energii Metabolizm całokształt reakcji chemicznych i związanych z nimi przemian energii zachodzący w komórkach. Podstawa wszelakich zjawisk biologicznych. Metabolizm
Fotosynteza. Pozyskiwanie i przetwarzanie energii w komórkach roślinnych. Chloroplasty 2014-04-01. Życie na Ziemi zależy od dopływu energii od słońca
 Życie na Ziemi zależy od dopływu energii od słońca Pozyskiwanie i przetwarzanie energii w komórkach roślinnych. Chloroplasty Fotosynteza Zdjęcie z mikroskopu świetlnego Zdjęcia z mikroskopu elektronowego
Życie na Ziemi zależy od dopływu energii od słońca Pozyskiwanie i przetwarzanie energii w komórkach roślinnych. Chloroplasty Fotosynteza Zdjęcie z mikroskopu świetlnego Zdjęcia z mikroskopu elektronowego
Fotosynteza. Celem ćwiczenia jest obserwacja zjawiska oddychania roślin w czasie dnia i nocy wraz z krótką analizą procesu fotosyntezy.
 Fotosynteza Program: Coach 6 Projekt: komputer G : C:\Program Files (x86)\cma\coach6\full.en\cma Coach Projects\PTSN Coach 6\Przyroda\Fotosynteza.cma Cel ćwiczenia Celem ćwiczenia jest obserwacja zjawiska
Fotosynteza Program: Coach 6 Projekt: komputer G : C:\Program Files (x86)\cma\coach6\full.en\cma Coach Projects\PTSN Coach 6\Przyroda\Fotosynteza.cma Cel ćwiczenia Celem ćwiczenia jest obserwacja zjawiska
Nukleotydy w układach biologicznych
 Nukleotydy w układach biologicznych Schemat 1. Dinukleotyd nikotynoamidoadeninowy Schemat 2. Dinukleotyd NADP + Dinukleotydy NAD +, NADP + i FAD uczestniczą w procesach biochemicznych, w trakcie których
Nukleotydy w układach biologicznych Schemat 1. Dinukleotyd nikotynoamidoadeninowy Schemat 2. Dinukleotyd NADP + Dinukleotydy NAD +, NADP + i FAD uczestniczą w procesach biochemicznych, w trakcie których
MATERIAŁY Z KURSU KWALIFIKACYJNEGO
 Wszystkie materiały tworzone i przekazywane przez Wykładowców NPDN PROTOTO są chronione prawem autorskim i przeznaczone wyłącznie do użytku prywatnego. MATERIAŁY Z KURSU KWALIFIKACYJNEGO PROCESY BIOLOGICZNE
Wszystkie materiały tworzone i przekazywane przez Wykładowców NPDN PROTOTO są chronione prawem autorskim i przeznaczone wyłącznie do użytku prywatnego. MATERIAŁY Z KURSU KWALIFIKACYJNEGO PROCESY BIOLOGICZNE
Przemiana materii i energii - Biologia.net.pl
 Ogół przemian biochemicznych, które zachodzą w komórce składają się na jej metabolizm. Wyróżnia się dwa antagonistyczne procesy metabolizmu: anabolizm i katabolizm. Szlak metaboliczny w komórce, to szereg
Ogół przemian biochemicznych, które zachodzą w komórce składają się na jej metabolizm. Wyróżnia się dwa antagonistyczne procesy metabolizmu: anabolizm i katabolizm. Szlak metaboliczny w komórce, to szereg
Jak pewnie zauważyłeś jest to odwrócenie procesu oddychania, dzięki któremu organizmy żywe pozyskują energię z rozkładu związków organicznych.
 Czy wiesz, że pomimo powszechnie obowiązującej edukacji wielu ludzi ciągle wierzy w popularny mit, który mówi o tym, że rośliny pobierają związki organiczne do budowy swoich tkanek z gleby, tzn. żywią
Czy wiesz, że pomimo powszechnie obowiązującej edukacji wielu ludzi ciągle wierzy w popularny mit, który mówi o tym, że rośliny pobierają związki organiczne do budowy swoich tkanek z gleby, tzn. żywią
Dlaczego warto zajmować się fotosyntezą?
 8 Dlaczego warto zajmować się fotosyntezą? Květoslava Burda Instytut Fizyki UJ Fotosynteza jest procesem odpowiedzialnym za wykorzystanie energii słonecznej do produkcji związków organicznych niezbędnych
8 Dlaczego warto zajmować się fotosyntezą? Květoslava Burda Instytut Fizyki UJ Fotosynteza jest procesem odpowiedzialnym za wykorzystanie energii słonecznej do produkcji związków organicznych niezbędnych
Kierunek: Elektrotechnika wersja z dn Promieniowanie optyczne Laboratorium
 Kierunek: Elektrotechnika wersja z dn. 04.05.2018 Promieniowanie optyczne Laboratorium Temat: OCENA WPŁYWU LAMP ELEKTRYCZNYCH NA SKUTECZNOŚĆ PROCESU FOTOSYNTEZY Opracowanie wykonano na podstawie: [1] DIN
Kierunek: Elektrotechnika wersja z dn. 04.05.2018 Promieniowanie optyczne Laboratorium Temat: OCENA WPŁYWU LAMP ELEKTRYCZNYCH NA SKUTECZNOŚĆ PROCESU FOTOSYNTEZY Opracowanie wykonano na podstawie: [1] DIN
Bliskie spotkania z biologią METABOLIZM. dr hab. Joanna Moraczewska, prof. UKW. Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki
 Bliskie spotkania z biologią METABOLIZM dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki Metabolizm całokształt przemian biochemicznych i towarzyszących
Bliskie spotkania z biologią METABOLIZM dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki Metabolizm całokształt przemian biochemicznych i towarzyszących
Reakcje zachodzące w komórkach
 Reakcje zachodzące w komórkach W każdej sekundzie we wszystkich organizmach żywych zachodzi niezliczona ilość reakcji metabolicznych. Metabolizm (gr. metabole - przemiana) to przemiany materii i energii
Reakcje zachodzące w komórkach W każdej sekundzie we wszystkich organizmach żywych zachodzi niezliczona ilość reakcji metabolicznych. Metabolizm (gr. metabole - przemiana) to przemiany materii i energii
Oddychanie komórkowe. Pozyskiwanie i przetwarzanie energii w komórkach roślinnych. Oddychanie zachodzi w mitochondriach Wykład 7.
 Wykład 7. Pozyskiwanie i przetwarzanie energii w komórkach roślinnych Literatura dodatkowa: Oddychanie to wielostopniowy proces utleniania substratów związany z wytwarzaniem w komórce metabolicznie użytecznej
Wykład 7. Pozyskiwanie i przetwarzanie energii w komórkach roślinnych Literatura dodatkowa: Oddychanie to wielostopniowy proces utleniania substratów związany z wytwarzaniem w komórce metabolicznie użytecznej
Bliskie spotkania z biologią. METABOLIZM część II. dr hab. Joanna Moraczewska, prof. UKW
 Bliskie spotkania z biologią METABOLIZM część II dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki METABOLIZM KATABOLIZM - rozkład związków chemicznych
Bliskie spotkania z biologią METABOLIZM część II dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki METABOLIZM KATABOLIZM - rozkład związków chemicznych
(węglowodanów i tłuszczów) Podstawowym produktem (nośnikiem energii) - ATP
 Podstawowym produktem (nośnikiem energii) - ATP") śycie - wymaga nakładu energii źródłem - promienie świetlne - wykorzystywane do fotosyntezy - magazynowanie energii w wiązaniach chemicznych Wszystkie organizmy (a zwierzęce wyłącznie) pozyskują energię
śycie - wymaga nakładu energii źródłem - promienie świetlne - wykorzystywane do fotosyntezy - magazynowanie energii w wiązaniach chemicznych Wszystkie organizmy (a zwierzęce wyłącznie) pozyskują energię
Spis treści. Fotosynteza. 1 Fotosynteza 1.1 WĘGLOWODANY 2 Cykl Krebsa 2.1 Acetylokoenzym A
 Spis treści 1 Fotosynteza 1.1 WĘGLOWODANY 2 Cykl Krebsa 2.1 Acetylokoenzym A Fotosynteza Jest to złożony, wieloetapowy proces redukcji dwutlenku węgla do substancji zawierających atomy węgla na niższych
Spis treści 1 Fotosynteza 1.1 WĘGLOWODANY 2 Cykl Krebsa 2.1 Acetylokoenzym A Fotosynteza Jest to złożony, wieloetapowy proces redukcji dwutlenku węgla do substancji zawierających atomy węgla na niższych
Spis treści. 1. Wiadomości wstępne Skład chemiczny i funkcje komórki Przedmowa do wydania czternastego... 13
 Przedmowa do wydania czternastego... 13 Częściej stosowane skróty... 15 1. Wiadomości wstępne... 19 1.1. Rys historyczny i pojęcia podstawowe... 19 1.2. Znaczenie biochemii w naukach rolniczych... 22 2.
Przedmowa do wydania czternastego... 13 Częściej stosowane skróty... 15 1. Wiadomości wstępne... 19 1.1. Rys historyczny i pojęcia podstawowe... 19 1.2. Znaczenie biochemii w naukach rolniczych... 22 2.
Izolacja chlorofilu z wybranych kultur roślinnych
 Wydział Chemiczny Politechniki Gdańskiej Katedra Technologii Leków i Biochemii Kultury tkankowe i komórkowe roślin i zwierząt Izolacja chlorofilu z wybranych kultur roślinnych Wiele waŝnych substancji
Wydział Chemiczny Politechniki Gdańskiej Katedra Technologii Leków i Biochemii Kultury tkankowe i komórkowe roślin i zwierząt Izolacja chlorofilu z wybranych kultur roślinnych Wiele waŝnych substancji
Transport przez błony
 Transport przez błony Transport bierny Nie wymaga nakładu energii Transport aktywny Wymaga nakładu energii Dyfuzja prosta Dyfuzja ułatwiona Przenośniki Kanały jonowe Transport przez pory w błonie jądrowej
Transport przez błony Transport bierny Nie wymaga nakładu energii Transport aktywny Wymaga nakładu energii Dyfuzja prosta Dyfuzja ułatwiona Przenośniki Kanały jonowe Transport przez pory w błonie jądrowej
Antyoksydanty pokarmowe a korzyści zdrowotne. dr hab. Agata Wawrzyniak, prof. SGGW Katedra Żywienia Człowieka SGGW
 Antyoksydanty pokarmowe a korzyści zdrowotne dr hab. Agata Wawrzyniak, prof. SGGW Katedra Żywienia Człowieka SGGW Warszawa, dn. 14.12.2016 wolne rodniki uszkodzone cząsteczki chemiczne w postaci wysoce
Antyoksydanty pokarmowe a korzyści zdrowotne dr hab. Agata Wawrzyniak, prof. SGGW Katedra Żywienia Człowieka SGGW Warszawa, dn. 14.12.2016 wolne rodniki uszkodzone cząsteczki chemiczne w postaci wysoce
Metody badańżywych organizmów Skład chemiczny organizmów żywych (zwłaszcza aktywnych organów) cały czas się zmienia. Również martwe tkanki przez
 cały czas się zmienia. Również martwe tkanki przez") Metody badańżywych organizmów Skład chemiczny organizmów żywych (zwłaszcza aktywnych organów) cały czas się zmienia. Również martwe tkanki przez jakiś czas ulegają procesom metabolicznym lub rozkładu.
Metody badańżywych organizmów Skład chemiczny organizmów żywych (zwłaszcza aktywnych organów) cały czas się zmienia. Również martwe tkanki przez jakiś czas ulegają procesom metabolicznym lub rozkładu.
TEORIA KOMÓRKI (dlaczego istnieją osobniki?)
") Wstęp do biologii 2. TEORIA KOMÓRKI (dlaczego istnieją osobniki?) Jerzy Dzik Instytut Paleobiologii PAN Instytut Zoologii UW 2015 WSPÓLNE WŁAŚCIWOŚCI dzisiejszych organizmów procesy życiowe katalizowane
Wstęp do biologii 2. TEORIA KOMÓRKI (dlaczego istnieją osobniki?) Jerzy Dzik Instytut Paleobiologii PAN Instytut Zoologii UW 2015 WSPÓLNE WŁAŚCIWOŚCI dzisiejszych organizmów procesy życiowe katalizowane
B) podział (aldolowy) na 2 triozy. 2) izomeryzacja do fruktozo-6-p (aldoza w ketozę, dla umoŝliwienia kolejnych przemian)
 podział (aldolowy) na 2 triozy. 2) izomeryzacja do fruktozo-6-p (aldoza w ketozę, dla umoŝliwienia kolejnych przemian)") Glikoliza (Przegląd kluczowych struktur i reakcji) A) przygotowanie heksozy do podziału na dwie triozy: 1)fosforylacja glukozy (czyli przekształcenie w formę metabolicznie aktywną) 2) izomeryzacja do fruktozo-6-p
Glikoliza (Przegląd kluczowych struktur i reakcji) A) przygotowanie heksozy do podziału na dwie triozy: 1)fosforylacja glukozy (czyli przekształcenie w formę metabolicznie aktywną) 2) izomeryzacja do fruktozo-6-p
TEORIA KOMÓRKI (dlaczego istnieją osobniki?)
") Wstęp do biologii 2. TEORIA KOMÓRKI (dlaczego istnieją osobniki?) Jerzy Dzik Instytut Paleobiologii PAN Instytut Zoologii UW 2017 WSPÓLNE WŁAŚCIWOŚCI dzisiejszych organizmów procesy życiowe katalizowane
Wstęp do biologii 2. TEORIA KOMÓRKI (dlaczego istnieją osobniki?) Jerzy Dzik Instytut Paleobiologii PAN Instytut Zoologii UW 2017 WSPÓLNE WŁAŚCIWOŚCI dzisiejszych organizmów procesy życiowe katalizowane
oksydacyjna ADP + Pi + (energia z utleniania zredukowanych nukleotydów ) ATP
 ATP") Życie - wymaga nakładu energii źródłem - promienie świetlne - wykorzystywane do fotosyntezy - magazynowanie energii w wiązaniach chemicznych Wszystkie organizmy (a zwierzęce wyłącznie) pozyskują energię
Życie - wymaga nakładu energii źródłem - promienie świetlne - wykorzystywane do fotosyntezy - magazynowanie energii w wiązaniach chemicznych Wszystkie organizmy (a zwierzęce wyłącznie) pozyskują energię
CZYNNIKI WPŁYWAJĄCE NA SZYBKOŚĆ REAKCJI CHEMICZNYCH. ILOŚCIOWE ZBADANIE SZYBKOŚCI ROZPADU NADTLENKU WODORU.
 CZYNNIKI WPŁYWAJĄCE NA SZYBKOŚĆ REAKCJI CHEMICZNYCH. ILOŚCIOWE ZBADANIE SZYBKOŚCI ROZPADU NADTLENKU WODORU. Projekt zrealizowany w ramach Mazowieckiego programu stypendialnego dla uczniów szczególnie uzdolnionych
CZYNNIKI WPŁYWAJĄCE NA SZYBKOŚĆ REAKCJI CHEMICZNYCH. ILOŚCIOWE ZBADANIE SZYBKOŚCI ROZPADU NADTLENKU WODORU. Projekt zrealizowany w ramach Mazowieckiego programu stypendialnego dla uczniów szczególnie uzdolnionych
Źródła energii dla mięśni. mgr. Joanna Misiorowska
 Źródła energii dla mięśni mgr. Joanna Misiorowska Skąd ta energia? Skurcz włókna mięśniowego wymaga nakładu energii w postaci ATP W zależności od czasu pracy mięśni, ATP może być uzyskiwany z różnych źródeł
Źródła energii dla mięśni mgr. Joanna Misiorowska Skąd ta energia? Skurcz włókna mięśniowego wymaga nakładu energii w postaci ATP W zależności od czasu pracy mięśni, ATP może być uzyskiwany z różnych źródeł
PODSTAWOWE PROCESY METABOLICZNE ORGANIZMÓW
 PODSTAWOWE PROCESY METABOLICZNE ORGANIZMÓW METABOLIZM (gr. metabole = przemiana) - przemiana materii - całość procesów biochemicznych zachodzących w żywych organizmach, warunkujących ich wzrost i funkcjonowanie.
PODSTAWOWE PROCESY METABOLICZNE ORGANIZMÓW METABOLIZM (gr. metabole = przemiana) - przemiana materii - całość procesów biochemicznych zachodzących w żywych organizmach, warunkujących ich wzrost i funkcjonowanie.
Mitochondria. siłownie komórki
 śycie - wymaga nakładu energii źródłem - promienie świetlne - wykorzystywane do fotosyntezy - magazynowanie energii w wiązaniach chemicznych Wszystkie organizmy ( a zwierzęce wyłącznie) pozyskują energię
śycie - wymaga nakładu energii źródłem - promienie świetlne - wykorzystywane do fotosyntezy - magazynowanie energii w wiązaniach chemicznych Wszystkie organizmy ( a zwierzęce wyłącznie) pozyskują energię
Repetytorium z wybranych zagadnień z chemii
 Repetytorium z wybranych zagadnień z chemii Mol jest to liczebność materii występująca, gdy liczba cząstek (elementów) układu jest równa liczbie atomów zawartych w masie 12 g węgla 12 C (równa liczbie
Repetytorium z wybranych zagadnień z chemii Mol jest to liczebność materii występująca, gdy liczba cząstek (elementów) układu jest równa liczbie atomów zawartych w masie 12 g węgla 12 C (równa liczbie
CORAZ BLIŻEJ ISTOTY ŻYCIA WERSJA A. imię i nazwisko :. klasa :.. ilość punktów :.
 CORAZ BLIŻEJ ISTOTY ŻYCIA WERSJA A imię i nazwisko :. klasa :.. ilość punktów :. Zadanie 1 Przeanalizuj schemat i wykonaj polecenia. a. Wymień cztery struktury występujące zarówno w komórce roślinnej,
CORAZ BLIŻEJ ISTOTY ŻYCIA WERSJA A imię i nazwisko :. klasa :.. ilość punktów :. Zadanie 1 Przeanalizuj schemat i wykonaj polecenia. a. Wymień cztery struktury występujące zarówno w komórce roślinnej,
Substancje o Znaczeniu Biologicznym
 Substancje o Znaczeniu Biologicznym Tłuszcze Jadalne są to tłuszcze, które może spożywać człowiek. Stanowią ważny, wysokoenergetyczny składnik diety. Z chemicznego punktu widzenia głównym składnikiem tłuszczów
Substancje o Znaczeniu Biologicznym Tłuszcze Jadalne są to tłuszcze, które może spożywać człowiek. Stanowią ważny, wysokoenergetyczny składnik diety. Z chemicznego punktu widzenia głównym składnikiem tłuszczów
Budowa anatomiczna liścia roślin okrytonasiennych.
 Organy wegetatywne roślin nasiennych: liście, pędy, korzenie. Budowa anatomiczna liścia roślin okrytonasiennych. Budowa morfologiczna liścia. Przekrój przez blaszkę liściową. Budowa anatomiczna liścia.
Organy wegetatywne roślin nasiennych: liście, pędy, korzenie. Budowa anatomiczna liścia roślin okrytonasiennych. Budowa morfologiczna liścia. Przekrój przez blaszkę liściową. Budowa anatomiczna liścia.
Spis treści. Katabolizm
 METABOLIZM Istnienie żywych organizmów jest uzależnione od energii potrzebnej do aktywności komórki w tym syntezy i transportu energii. Energia, która została zużyta przez organizm do wykonania pracy biologicznej
METABOLIZM Istnienie żywych organizmów jest uzależnione od energii potrzebnej do aktywności komórki w tym syntezy i transportu energii. Energia, która została zużyta przez organizm do wykonania pracy biologicznej
etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
 Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
CHARAKTERYSTYKI SPEKTRALNE UTLENIONEJ I ZREDUKOWANEJ FORMY CYTOCHROMU C
 Ćwiczenie 4 CHARAKTERYSTYKI SPEKTRALNE UTLENIONEJ I ZREDUKOWANEJ FORMY CYTOCHROMU C REAKTYWNE FORMY TLENU DEGRADACJA NUKLEOTYDÓW PURYNOWYCH TWORZENIE ANIONORODNIKA PONADTLENKOWEGO W REAKCJI KATALIZOWANEJ
Ćwiczenie 4 CHARAKTERYSTYKI SPEKTRALNE UTLENIONEJ I ZREDUKOWANEJ FORMY CYTOCHROMU C REAKTYWNE FORMY TLENU DEGRADACJA NUKLEOTYDÓW PURYNOWYCH TWORZENIE ANIONORODNIKA PONADTLENKOWEGO W REAKCJI KATALIZOWANEJ
MECHANIZM DZIAŁANIA HERBICYDÓW
 Uniwersytet Przyrodniczy w Poznaniu Grzegorz Skrzypczak MECHANIZM DZIAŁANIA HERBICYDÓW metabolizm herbicydów Nowe technologie uprawy wymagają aby herbicyd był: - skuteczny biologicznie i efektywny ekonomicznie
Uniwersytet Przyrodniczy w Poznaniu Grzegorz Skrzypczak MECHANIZM DZIAŁANIA HERBICYDÓW metabolizm herbicydów Nowe technologie uprawy wymagają aby herbicyd był: - skuteczny biologicznie i efektywny ekonomicznie
Plan działania opracowała Anna Gajos
 Plan działania 15.09-15.10 opracowała Anna Gajos Jakie zagadnienia trzeba opanować z następujących działów: 1. Budowa chemiczna organizmów. 2. Budowa i funkcjonowanie komórki 3. Cykl komórkowy 4. Metabolizm
Plan działania 15.09-15.10 opracowała Anna Gajos Jakie zagadnienia trzeba opanować z następujących działów: 1. Budowa chemiczna organizmów. 2. Budowa i funkcjonowanie komórki 3. Cykl komórkowy 4. Metabolizm
Fizjologia i Regulacja Metabolizmu. Jarosław Szczepanik. Instytut Biologii Eksperymentalnej i Biotechnologii Roślin
 FOTOSYNTEZA I PROCESY ODDECHOWE ROŚLIN Fizjologia i Regulacja Metabolizmu Jarosław Szczepanik Instytut Biologii Eksperymentalnej i Biotechnologii Roślin Plan prezentacji 1 Wstęp Rośliny jako fotoautotrofy
FOTOSYNTEZA I PROCESY ODDECHOWE ROŚLIN Fizjologia i Regulacja Metabolizmu Jarosław Szczepanik Instytut Biologii Eksperymentalnej i Biotechnologii Roślin Plan prezentacji 1 Wstęp Rośliny jako fotoautotrofy
Wykład Fotosynteza typu C3, C4 i CAM. Autor dr Agnieszka Gniazdowska-Piekarska
 Wykład Fotosynteza typu C3, C4 i CAM Autor dr Agnieszka Gniazdowska-Piekarska Z tego wykładu dowiesz się jakie czynniki regulują intensywność fotosyntezy. Jakie są różnice pomiędzy roślinami o fotosyntezie
Wykład Fotosynteza typu C3, C4 i CAM Autor dr Agnieszka Gniazdowska-Piekarska Z tego wykładu dowiesz się jakie czynniki regulują intensywność fotosyntezy. Jakie są różnice pomiędzy roślinami o fotosyntezie
Biochemia Oddychanie wewnątrzkomórkowe
 Państwowa Wyższa Szkoła Zawodowa w Krośnie Biochemia Oddychanie wewnątrzkomórkowe Dr n. biol. Henryk Różański Laboratorium Biologii Przemysłowej i Eksperymentalnej Oddychanie Glikoliza beztlenowy, wewnątrzkomórkowy
Państwowa Wyższa Szkoła Zawodowa w Krośnie Biochemia Oddychanie wewnątrzkomórkowe Dr n. biol. Henryk Różański Laboratorium Biologii Przemysłowej i Eksperymentalnej Oddychanie Glikoliza beztlenowy, wewnątrzkomórkowy
Bliskie spotkania z biologią METABOLIZM. dr hab. Joanna Moraczewska, prof. UKW. Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki
 Bliskie spotkania z biologią METABOLIZM dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki Metabolizm całokształt przemian biochemicznych i towarzyszących
Bliskie spotkania z biologią METABOLIZM dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki Metabolizm całokształt przemian biochemicznych i towarzyszących
TEST Z CYTOLOGII GRUPA II
 TEST Z CYTOLOGII GRUPA II Zad. 1 (4p.) Rysunek przedstawia schemat budowy pewnej struktury komórkowej. a/ podaj jej nazwę i określ funkcję w komórce, b/ nazwij elementy oznaczone cyframi 2 i 5 oraz określ
TEST Z CYTOLOGII GRUPA II Zad. 1 (4p.) Rysunek przedstawia schemat budowy pewnej struktury komórkowej. a/ podaj jej nazwę i określ funkcję w komórce, b/ nazwij elementy oznaczone cyframi 2 i 5 oraz określ
Zakład Biofizyki Molekularnej i Bioenergetyki Katedra Fizyki Medycznej i Biofizyki, WFiIS, AGH
 dr Alekandra Orzechowska Zakład Biofizyki Molekularnej i Bioenergetyki Katedra Fizyki Medycznej i Biofizyki, WFiIS, AGH D-11, I piętro, pok. 114 Aleksandra.Orzechowska@fis.agh.edu.pl konsultacje: środa,
dr Alekandra Orzechowska Zakład Biofizyki Molekularnej i Bioenergetyki Katedra Fizyki Medycznej i Biofizyki, WFiIS, AGH D-11, I piętro, pok. 114 Aleksandra.Orzechowska@fis.agh.edu.pl konsultacje: środa,
Opracowała: mgr inż. Ewelina Nowak
 Materiały dydaktyczne na zajęcia wyrównawcze z chemii dla studentów pierwszego roku kierunku zamawianego Inżynieria Środowiska w ramach projektu Era inżyniera pewna lokata na przyszłość Opracowała: mgr
Materiały dydaktyczne na zajęcia wyrównawcze z chemii dla studentów pierwszego roku kierunku zamawianego Inżynieria Środowiska w ramach projektu Era inżyniera pewna lokata na przyszłość Opracowała: mgr
OKSYDOREDUKTAZY WPROWADZENIE
 Ćwiczenie 6 OKSYDOREDUKTAZY Część doświadczalna obejmuje: wykrywanie aktywności katalazy, peroksydazy, oksydazy polifenolowej i oksydazy cytochromowej w ekstrakcie z bulwy ziemniaka WPROWADZENIE Oksydoreduktazy
Ćwiczenie 6 OKSYDOREDUKTAZY Część doświadczalna obejmuje: wykrywanie aktywności katalazy, peroksydazy, oksydazy polifenolowej i oksydazy cytochromowej w ekstrakcie z bulwy ziemniaka WPROWADZENIE Oksydoreduktazy
X / \ Y Y Y Z / \ W W ... imię i nazwisko,nazwa szkoły, miasto
 Zadanie 1. (3 pkt) Nadtlenek litu (Li 2 O 2 ) jest ciałem stałym, występującym w temperaturze pokojowej w postaci białych kryształów. Stosowany jest w oczyszczaczach powietrza, gdzie ważna jest waga użytego
Zadanie 1. (3 pkt) Nadtlenek litu (Li 2 O 2 ) jest ciałem stałym, występującym w temperaturze pokojowej w postaci białych kryształów. Stosowany jest w oczyszczaczach powietrza, gdzie ważna jest waga użytego
Metody fosforylacji. Schemat 1. Powstawanie trifosforanu nukleozydu
 Metody fosforylacji Fosforylacja jest procesem przenoszenia reszty fosforanowej do nukleofilowego atomu dowolnego związku chemicznego. Najczęściej fosforylację przeprowadza się na atomie tlenu grupy hydroksylowej
Metody fosforylacji Fosforylacja jest procesem przenoszenia reszty fosforanowej do nukleofilowego atomu dowolnego związku chemicznego. Najczęściej fosforylację przeprowadza się na atomie tlenu grupy hydroksylowej
Cel ćwiczenia: Zapoznanie się z metodą analizy jakościowej oraz własnościami fizykochemicznymi barwników fotosyntetycznych.
 Ćwiczenie nr 7 Ekstrakcja i rozdział barwników roślinnych-pomiar i analiza widm absorpcji UV-Vis Cel ćwiczenia: Zapoznanie się z metodą analizy jakościowej oraz własnościami fizykochemicznymi barwników
Ćwiczenie nr 7 Ekstrakcja i rozdział barwników roślinnych-pomiar i analiza widm absorpcji UV-Vis Cel ćwiczenia: Zapoznanie się z metodą analizy jakościowej oraz własnościami fizykochemicznymi barwników
Makrocząsteczki. Przykłady makrocząsteczek naturalnych: -Polisacharydy skrobia, celuloza -Białka -Kwasy nukleinowe
 Makrocząsteczki Przykłady makrocząsteczek naturalnych: -Polisacharydy skrobia, celuloza -Białka -Kwasy nukleinowe Syntetyczne: -Elastomery bardzo duża elastyczność charakterystyczna dla gumy -Włókna długie,
Makrocząsteczki Przykłady makrocząsteczek naturalnych: -Polisacharydy skrobia, celuloza -Białka -Kwasy nukleinowe Syntetyczne: -Elastomery bardzo duża elastyczność charakterystyczna dla gumy -Włókna długie,
Widmo promieniowania
 Widmo promieniowania Spektroskopia Każde ciało wysyła promieniowanie. Promieniowanie to jest składa się z wiązek o różnych długościach fal. Jeśli wiązka światła pada na pryzmat, ulega ono rozszczepieniu,
Widmo promieniowania Spektroskopia Każde ciało wysyła promieniowanie. Promieniowanie to jest składa się z wiązek o różnych długościach fal. Jeśli wiązka światła pada na pryzmat, ulega ono rozszczepieniu,
WŁASNOŚCI SPEKTRALNE NUKLEOTYDÓW PIRYDYNOWYCH (NAD +, NADP + ) OZNACZANIE AKTYWNOŚCI TRANSAMINAZY ALANINOWEJ
 OZNACZANIE AKTYWNOŚCI TRANSAMINAZY ALANINOWEJ") WŁASNOŚCI SPEKTRALNE NUKLEOTYDÓW PIRYDYNOWYCH (NAD +, NADP + ) OZNACZANIE AKTYWNOŚCI TRANSAMINAZY ALANINOWEJ WSTĘP Nukleotydy pirydynowe (NAD +, NADP + ) pełnią funkcję koenzymów dehydrogenaz przenosząc
WŁASNOŚCI SPEKTRALNE NUKLEOTYDÓW PIRYDYNOWYCH (NAD +, NADP + ) OZNACZANIE AKTYWNOŚCI TRANSAMINAZY ALANINOWEJ WSTĘP Nukleotydy pirydynowe (NAD +, NADP + ) pełnią funkcję koenzymów dehydrogenaz przenosząc
Spektroskopowe metody identyfikacji związków organicznych
 Spektroskopowe metody identyfikacji związków organicznych Wstęp Spektroskopia jest metodą analityczną zajmującą się analizą widm powstających w wyniku oddziaływania promieniowania elektromagnetycznego
Spektroskopowe metody identyfikacji związków organicznych Wstęp Spektroskopia jest metodą analityczną zajmującą się analizą widm powstających w wyniku oddziaływania promieniowania elektromagnetycznego
MODEL ODPOWIEDZI I SCHEMAT PUNKTOWANIA
 MODEL ODPOWIEDZI I SCHEMAT PUNKTOWANIA Zadanie Odpowiedzi Uwagi a) za uzupełnienie tabeli: Symbol pierwiastka Konfiguracja elektronowa w stanie podstawowym Liczba elektronów walencyjnych S b) za uzupełnienie
MODEL ODPOWIEDZI I SCHEMAT PUNKTOWANIA Zadanie Odpowiedzi Uwagi a) za uzupełnienie tabeli: Symbol pierwiastka Konfiguracja elektronowa w stanie podstawowym Liczba elektronów walencyjnych S b) za uzupełnienie
Zasada pomiaru intensywności fotosyntezy netto i oddychania techniką wymiany gazowej, analizatorem gazu w podczerwieni
 Zasada pomiaru intensywności fotosyntezy netto i oddychania techniką wymiany gazowej, analizatorem gazu w podczerwieni Pomiar wymiany gazowej do oceny intensywności fotosyntezy stosowany jest przez fizjologów
Zasada pomiaru intensywności fotosyntezy netto i oddychania techniką wymiany gazowej, analizatorem gazu w podczerwieni Pomiar wymiany gazowej do oceny intensywności fotosyntezy stosowany jest przez fizjologów
2. Podczas spalania 2 objętości pewnego gazu z 4 objętościami H 2 otrzymano 1 objętość N 2 i 4 objętości H 2O. Jaki gaz uległ spalaniu?
 1. Oblicz, ilu moli HCl należy użyć, aby poniższe związki przeprowadzić w sole: a) 0,2 mola KOH b) 3 mole NH 3 H 2O c) 0,2 mola Ca(OH) 2 d) 0,5 mola Al(OH) 3 2. Podczas spalania 2 objętości pewnego gazu
1. Oblicz, ilu moli HCl należy użyć, aby poniższe związki przeprowadzić w sole: a) 0,2 mola KOH b) 3 mole NH 3 H 2O c) 0,2 mola Ca(OH) 2 d) 0,5 mola Al(OH) 3 2. Podczas spalania 2 objętości pewnego gazu
1. Od czego i w jaki sposób zależy szybkość reakcji chemicznej?
 Tematy opisowe 1. Od czego i w jaki sposób zależy szybkość reakcji chemicznej? 2. Omów pomiar potencjału na granicy faz elektroda/roztwór elektrolitu. Podaj przykład, omów skale potencjału i elektrody
Tematy opisowe 1. Od czego i w jaki sposób zależy szybkość reakcji chemicznej? 2. Omów pomiar potencjału na granicy faz elektroda/roztwór elektrolitu. Podaj przykład, omów skale potencjału i elektrody
Integracja metabolizmu
 Integracja metabolizmu 1 Kluczowe związki w metabolizmie Glukozo- 6 -fosforan Pirogronian AcetyloCoA 2 Glukoza po wejściu do komórki ulega fosforylacji Metaboliczne przemiany glukozo- 6-fosforanu G-6-P
Integracja metabolizmu 1 Kluczowe związki w metabolizmie Glukozo- 6 -fosforan Pirogronian AcetyloCoA 2 Glukoza po wejściu do komórki ulega fosforylacji Metaboliczne przemiany glukozo- 6-fosforanu G-6-P
Beata Mendak fakultety z chemii II tura PYTANIA Z KLASY PIERWSZEJ
 Beata Mendak fakultety z chemii II tura Test rozwiązywany na zajęciach wymaga powtórzenia stężenia procentowego i rozpuszczalności. Podaję również pytania do naszej zaplanowanej wcześniej MEGA POWTÓRKI
Beata Mendak fakultety z chemii II tura Test rozwiązywany na zajęciach wymaga powtórzenia stężenia procentowego i rozpuszczalności. Podaję również pytania do naszej zaplanowanej wcześniej MEGA POWTÓRKI
Kryteria oceniania z chemii kl VII
 Kryteria oceniania z chemii kl VII Ocena dopuszczająca -stosuje zasady BHP w pracowni -nazywa sprzęt laboratoryjny i szkło oraz określa ich przeznaczenie -opisuje właściwości substancji używanych na co
Kryteria oceniania z chemii kl VII Ocena dopuszczająca -stosuje zasady BHP w pracowni -nazywa sprzęt laboratoryjny i szkło oraz określa ich przeznaczenie -opisuje właściwości substancji używanych na co
Ćwiczenie nr 5 - Reaktywne formy tlenu
 Ćwiczenie nr 5 - Reaktywne formy tlenu I. Oznaczenie ilościowe glutationu (GSH) metodą Ellmana II. Pomiar całkowitej zdolności antyoksydacyjnej substancji metodą redukcji rodnika DPPH Celem ćwiczeń jest:
Ćwiczenie nr 5 - Reaktywne formy tlenu I. Oznaczenie ilościowe glutationu (GSH) metodą Ellmana II. Pomiar całkowitej zdolności antyoksydacyjnej substancji metodą redukcji rodnika DPPH Celem ćwiczeń jest:
Jan Drzymała ANALIZA INSTRUMENTALNA SPEKTROSKOPIA W ŚWIETLE WIDZIALNYM I PODCZERWONYM
 Jan Drzymała ANALIZA INSTRUMENTALNA SPEKTROSKOPIA W ŚWIETLE WIDZIALNYM I PODCZERWONYM Światło słoneczne jest mieszaniną fal o różnej długości i różnego natężenia. Tylko część promieniowania elektromagnetycznego
Jan Drzymała ANALIZA INSTRUMENTALNA SPEKTROSKOPIA W ŚWIETLE WIDZIALNYM I PODCZERWONYM Światło słoneczne jest mieszaniną fal o różnej długości i różnego natężenia. Tylko część promieniowania elektromagnetycznego
Wymagania edukacyjne. Poziomy oczekiwanych osiągnięć ucznia
 Wymagania edukacyjne Zawierają szczegółowy wykaz wiadomości i umiejętności, które uczeń powinien opanować po omówieniu poszczególnych lekcji z podręcznika Biologia na czasie 2 zakres rozszerzony. Są niezastąpione
Wymagania edukacyjne Zawierają szczegółowy wykaz wiadomości i umiejętności, które uczeń powinien opanować po omówieniu poszczególnych lekcji z podręcznika Biologia na czasie 2 zakres rozszerzony. Są niezastąpione
WOJEWÓDZKI KONKURS PRZEDMIOTOWY Z CHEMII DLA UCZNIÓW GIMNAZJÓW - rok szkolny 2016/2017 eliminacje rejonowe
 kod ŁÓDZKIE CENTRUM DOSKONALENIA NAUCZYCIELI I KSZTAŁCENIA PRAKTYCZNEGO Uzyskane punkty.. WOJEWÓDZKI KONKURS PRZEDMIOTOWY Z CHEMII DLA UCZNIÓW GIMNAZJÓW - rok szkolny 2016/2017 eliminacje rejonowe Zadanie
kod ŁÓDZKIE CENTRUM DOSKONALENIA NAUCZYCIELI I KSZTAŁCENIA PRAKTYCZNEGO Uzyskane punkty.. WOJEWÓDZKI KONKURS PRZEDMIOTOWY Z CHEMII DLA UCZNIÓW GIMNAZJÓW - rok szkolny 2016/2017 eliminacje rejonowe Zadanie
Odwracalność przemiany chemicznej
 Odwracalność przemiany chemicznej Na ogół wszystkie reakcje chemiczne są odwracalne, tzn. z danych substratów tworzą się produkty, a jednocześnie produkty reakcji ulegają rozkładowi na substraty. Fakt
Odwracalność przemiany chemicznej Na ogół wszystkie reakcje chemiczne są odwracalne, tzn. z danych substratów tworzą się produkty, a jednocześnie produkty reakcji ulegają rozkładowi na substraty. Fakt
Temat: Komórka jako podstawowa jednostka strukturalna i funkcjonalna organizmu utrwalenie wiadomości.
 SCENARIUSZ LEKCJI BIOLOGII DLA KLASY I GIMNAZJUM Temat: Komórka jako podstawowa jednostka strukturalna i funkcjonalna organizmu utrwalenie wiadomości. Cele: Utrwalenie pojęć związanych z budową komórki;
SCENARIUSZ LEKCJI BIOLOGII DLA KLASY I GIMNAZJUM Temat: Komórka jako podstawowa jednostka strukturalna i funkcjonalna organizmu utrwalenie wiadomości. Cele: Utrwalenie pojęć związanych z budową komórki;
TYPY REAKCJI CHEMICZNYCH
 1 REAKCJA CHEMICZNA: TYPY REAKCJI CHEMICZNYCH REAKCJĄ CHEMICZNĄ NAZYWAMY PROCES, W WYNIKU KTÓREGO Z JEDNYCH SUBSTANCJI POWSTAJĄ NOWE (PRODUKTY) O INNYCH WŁAŚCIWOŚCIACH NIŻ SUBSTANCJE WYJŚCIOWE (SUBSTRATY)
1 REAKCJA CHEMICZNA: TYPY REAKCJI CHEMICZNYCH REAKCJĄ CHEMICZNĄ NAZYWAMY PROCES, W WYNIKU KTÓREGO Z JEDNYCH SUBSTANCJI POWSTAJĄ NOWE (PRODUKTY) O INNYCH WŁAŚCIWOŚCIACH NIŻ SUBSTANCJE WYJŚCIOWE (SUBSTRATY)
ZWIĄZKI WĘGLA Z WODOREM 1) Uzupełnij i uzgodnij równania reakcji spalania całkowitego alkanów, alkenów i alkinów.
 Uzupełnij i uzgodnij równania reakcji spalania całkowitego alkanów, alkenów i alkinów.") Nauczanie domowe WIEM, CO TRZEBA Klasa VIII Chemia od listopada do czerwca aktualizacja 05.10.2018 ZWIĄZKI WĘGLA Z WODOREM 1) Uzupełnij i uzgodnij równania reakcji spalania całkowitego alkanów, alkenów
Nauczanie domowe WIEM, CO TRZEBA Klasa VIII Chemia od listopada do czerwca aktualizacja 05.10.2018 ZWIĄZKI WĘGLA Z WODOREM 1) Uzupełnij i uzgodnij równania reakcji spalania całkowitego alkanów, alkenów
Chemia Środowiska. Ćw. Wpływ kwaśnych deszczy na roślinność. Zielone technologie i monitoring
 Chemia Środowiska Ćw. Wpływ kwaśnych deszczy na roślinność Zielone technologie i monitoring Barwniki asymilacyjne Barwnikami nazywamy związki chemiczne, które odbijają pewien zakres fal światła widzialnego,
Chemia Środowiska Ćw. Wpływ kwaśnych deszczy na roślinność Zielone technologie i monitoring Barwniki asymilacyjne Barwnikami nazywamy związki chemiczne, które odbijają pewien zakres fal światła widzialnego,
Budowa atomu Poziom: rozszerzony Zadanie 1. (2 pkt.)
") Budowa atomu Poziom: rozszerzony Zadanie 1. (2 pkt.) Zadanie 1 2 3 4 5 6 7 8 9 10 Punkty Okres połowiczego rozpadu pewnego radionuklidu wynosi 16 godzin. a) Określ, ile procent atomów tego izotopu rozpadnie
Budowa atomu Poziom: rozszerzony Zadanie 1. (2 pkt.) Zadanie 1 2 3 4 5 6 7 8 9 10 Punkty Okres połowiczego rozpadu pewnego radionuklidu wynosi 16 godzin. a) Określ, ile procent atomów tego izotopu rozpadnie
OZNACZANIE ŻELAZA METODĄ SPEKTROFOTOMETRII UV/VIS
 OZNACZANIE ŻELAZA METODĄ SPEKTROFOTOMETRII UV/VIS Zagadnienia teoretyczne. Spektrofotometria jest techniką instrumentalną, w której do celów analitycznych wykorzystuje się przejścia energetyczne zachodzące
OZNACZANIE ŻELAZA METODĄ SPEKTROFOTOMETRII UV/VIS Zagadnienia teoretyczne. Spektrofotometria jest techniką instrumentalną, w której do celów analitycznych wykorzystuje się przejścia energetyczne zachodzące
Materiały pochodzą z Platformy Edukacyjnej Portalu www.szkolnictwo.pl
 Materiały pochodzą z Platformy Edukacyjnej Portalu www.szkolnictwo.pl Wszelkie treści i zasoby edukacyjne publikowane na łamach Portalu www.szkolnictwo.pl mogą byd wykorzystywane przez jego Użytkowników
Materiały pochodzą z Platformy Edukacyjnej Portalu www.szkolnictwo.pl Wszelkie treści i zasoby edukacyjne publikowane na łamach Portalu www.szkolnictwo.pl mogą byd wykorzystywane przez jego Użytkowników
Cukry właściwości i funkcje
 Cukry właściwości i funkcje Miejsce cukrów wśród innych składników chemicznych Cukry Z cukrem mamy do czynienia bardzo często - kiedy sięgamy po białe kryształy z cukiernicy. Większość z nas nie uświadamia
Cukry właściwości i funkcje Miejsce cukrów wśród innych składników chemicznych Cukry Z cukrem mamy do czynienia bardzo często - kiedy sięgamy po białe kryształy z cukiernicy. Większość z nas nie uświadamia
KARTA KURSU. Fizjologia roślin Ochrona środowiska studia stacjonarne I stopnia. Kod Punktacja ECTS* 3. Dr hab. Andrzej Rzepka Prof.
 KARTA KURSU Nazwa Nazwa w j. ang. Fizjologia roślin Ochrona środowiska studia stacjonarne I stopnia Plant physiology Kod Punktacja ECTS* 3 Koordynator Dr hab. Andrzej Rzepka Prof. UP Zespół dydaktyczny
KARTA KURSU Nazwa Nazwa w j. ang. Fizjologia roślin Ochrona środowiska studia stacjonarne I stopnia Plant physiology Kod Punktacja ECTS* 3 Koordynator Dr hab. Andrzej Rzepka Prof. UP Zespół dydaktyczny
Metabolizm białek. Ogólny schemat metabolizmu bialek
 Metabolizm białek Ogólny schemat metabolizmu bialek Trawienie białek i absorpcja aminokwasów w przewodzie pokarmowym w żołądku (niskie ph ~2, rola HCl)- hydratacja, homogenizacja, denaturacja białek i
Metabolizm białek Ogólny schemat metabolizmu bialek Trawienie białek i absorpcja aminokwasów w przewodzie pokarmowym w żołądku (niskie ph ~2, rola HCl)- hydratacja, homogenizacja, denaturacja białek i
EKSTRAHOWANIE KWASÓW NUKLEINOWYCH JAK ZMIERZYĆ ILOŚĆ KWASÓW NUKLEINOWYCH PO IZOLACJI? JAK ZMIERZYĆ ILOŚĆ KWASÓW NUKLEINOWYCH PO IZOLACJI?
 EKSTRAHOWANIE KWASÓW NUKLEINOWYCH Wytrącanie etanolem Rozpuszczenie kwasu nukleinowego w fazie wodnej (met. fenol/chloroform) Wiązanie ze złożem krzemionkowym za pomocą substancji chaotropowych: jodek
EKSTRAHOWANIE KWASÓW NUKLEINOWYCH Wytrącanie etanolem Rozpuszczenie kwasu nukleinowego w fazie wodnej (met. fenol/chloroform) Wiązanie ze złożem krzemionkowym za pomocą substancji chaotropowych: jodek
Ćwiczenie nr 12 Lipidy - tłuszcze nasycone i nienasycone. Liczba jodowa, metoda Hanusa ilościowego oznaczania stopnia nienasycenia tłuszczu
 Ćwiczenie nr 12 Lipidy - tłuszcze nasycone i nienasycone. Liczba jodowa, metoda Hanusa ilościowego oznaczania stopnia nienasycenia tłuszczu Celem ćwiczenia jest: wykrywanie nienasyconych kwasów tłuszczowych
Ćwiczenie nr 12 Lipidy - tłuszcze nasycone i nienasycone. Liczba jodowa, metoda Hanusa ilościowego oznaczania stopnia nienasycenia tłuszczu Celem ćwiczenia jest: wykrywanie nienasyconych kwasów tłuszczowych
... Nazwisko, imię zawodnika; Klasa Liczba punktów. ... Nazwa szkoły, miejscowość. I Podkarpacki Konkurs Chemiczny 2008/09
 ......... Nazwisko, imię zawodnika; Klasa Liczba punktów KOPKCh... Nazwa szkoły, miejscowość I Podkarpacki Konkurs Chemiczny 2008/09 ETAP III 28.02.2009 r. Godz. 10.00-13.00 Zadanie 1 (10 pkt.) ( postaw
......... Nazwisko, imię zawodnika; Klasa Liczba punktów KOPKCh... Nazwa szkoły, miejscowość I Podkarpacki Konkurs Chemiczny 2008/09 ETAP III 28.02.2009 r. Godz. 10.00-13.00 Zadanie 1 (10 pkt.) ( postaw
WYKRYWANIE ZANIECZYSZCZEŃ WODY POWIERZA I GLEBY
 WYKRYWANIE ZANIECZYSZCZEŃ WODY POWIERZA I GLEBY Instrukcja przygotowana w Pracowni Dydaktyki Chemii Zakładu Fizykochemii Roztworów. 1. Zanieczyszczenie wody. Polska nie należy do krajów posiadających znaczne
WYKRYWANIE ZANIECZYSZCZEŃ WODY POWIERZA I GLEBY Instrukcja przygotowana w Pracowni Dydaktyki Chemii Zakładu Fizykochemii Roztworów. 1. Zanieczyszczenie wody. Polska nie należy do krajów posiadających znaczne
Obliczenia stechiometryczne, bilansowanie równań reakcji redoks
 Obliczenia stechiometryczne, bilansowanie równań reakcji redoks Materiały pomocnicze do zajęć wspomagających z chemii opracował: dr Błażej Gierczyk Wydział Chemii UAM Obliczenia stechiometryczne Podstawą
Obliczenia stechiometryczne, bilansowanie równań reakcji redoks Materiały pomocnicze do zajęć wspomagających z chemii opracował: dr Błażej Gierczyk Wydział Chemii UAM Obliczenia stechiometryczne Podstawą
Bliskie spotkania z biologią METABOLIZM. dr hab. Joanna Moraczewska, prof. UKW. Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki
 Bliskie spotkania z biologią METABOLIZM dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki Metabolizm całokształt przemian biochemicznych i towarzyszących
Bliskie spotkania z biologią METABOLIZM dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki Metabolizm całokształt przemian biochemicznych i towarzyszących
Kuratorium Oświaty w Lublinie
 Kuratorium Oświaty w Lublinie KOD UCZNIA ZESTAW ZADAŃ KONKURSOWYCH Z CHEMII DLA UCZNIÓW GIMNAZJÓW ROK SZKOLNY 2015/2016 ETAP WOJEWÓDZKI Instrukcja dla ucznia 1. Zestaw konkursowy zawiera 12 zadań. 2. Przed
Kuratorium Oświaty w Lublinie KOD UCZNIA ZESTAW ZADAŃ KONKURSOWYCH Z CHEMII DLA UCZNIÓW GIMNAZJÓW ROK SZKOLNY 2015/2016 ETAP WOJEWÓDZKI Instrukcja dla ucznia 1. Zestaw konkursowy zawiera 12 zadań. 2. Przed
Aminotransferazy. Dehydrogenaza glutaminianowa. Szczawiooctan. Argininobursztynian. Inne aminokwasy. asparaginian. fumaran. Arginina.
 Inne aminokwasy Szczawiooctan Aminotransferazy asparaginian Cytrulina Argininobursztynian Cykl mocznikowy Arginina fumaran Ornityna Aminotransferazy -ketoglutaran karbamoilofosforan Mocznik kwas glutaminowy
Inne aminokwasy Szczawiooctan Aminotransferazy asparaginian Cytrulina Argininobursztynian Cykl mocznikowy Arginina fumaran Ornityna Aminotransferazy -ketoglutaran karbamoilofosforan Mocznik kwas glutaminowy
KARTA KURSU. Fizjologia roślin I. Plant physiology I
 Biologia, I stopień, stacjonarne, 2017/2018, semestr IV KARTA KURSU Nazwa Nazwa w j. ang. Fizjologia roślin I Plant physiology I Koordynator Prof. dr hab. Andrzej Skoczowski Punktacja ECTS* 3 Zespół dydaktyczny
Biologia, I stopień, stacjonarne, 2017/2018, semestr IV KARTA KURSU Nazwa Nazwa w j. ang. Fizjologia roślin I Plant physiology I Koordynator Prof. dr hab. Andrzej Skoczowski Punktacja ECTS* 3 Zespół dydaktyczny
Reakcje chemiczne. Typ reakcji Schemat Przykłady Reakcja syntezy
 Reakcje chemiczne Literatura: L. Jones, P. Atkins Chemia ogólna. Cząsteczki, materia, reakcje. Lesław Huppenthal, Alicja Kościelecka, Zbigniew Wojtczak Chemia ogólna i analityczna dla studentów biologii.
Reakcje chemiczne Literatura: L. Jones, P. Atkins Chemia ogólna. Cząsteczki, materia, reakcje. Lesław Huppenthal, Alicja Kościelecka, Zbigniew Wojtczak Chemia ogólna i analityczna dla studentów biologii.
Badanie oddziaływania polihistydynowych cyklopeptydów z jonami Cu 2+ i Zn 2+ w aspekcie projektowania mimetyków SOD
 Wydział Farmaceutyczny z Oddziałem Analityki Medycznej Badanie oddziaływania polihistydynowych cyklopeptydów z jonami Cu 2+ i Zn 2+ w aspekcie projektowania mimetyków SOD Aleksandra Kotynia PRACA DOKTORSKA
Wydział Farmaceutyczny z Oddziałem Analityki Medycznej Badanie oddziaływania polihistydynowych cyklopeptydów z jonami Cu 2+ i Zn 2+ w aspekcie projektowania mimetyków SOD Aleksandra Kotynia PRACA DOKTORSKA
relacje ilościowe ( masowe,objętościowe i molowe ) dotyczące połączeń 1. pierwiastków w związkach chemicznych 2. związków chemicznych w reakcjach
 dotyczące połączeń 1. pierwiastków w związkach chemicznych 2. związków chemicznych w reakcjach") 1 STECHIOMETRIA INTERPRETACJA ILOŚCIOWA ZJAWISK CHEMICZNYCH relacje ilościowe ( masowe,objętościowe i molowe ) dotyczące połączeń 1. pierwiastków w związkach chemicznych 2. związków chemicznych w reakcjach
1 STECHIOMETRIA INTERPRETACJA ILOŚCIOWA ZJAWISK CHEMICZNYCH relacje ilościowe ( masowe,objętościowe i molowe ) dotyczące połączeń 1. pierwiastków w związkach chemicznych 2. związków chemicznych w reakcjach
Zagadnienia z chemii na egzamin wstępny kierunek Technik Farmaceutyczny Szkoła Policealna im. J. Romanowskiej
 Zagadnienia z chemii na egzamin wstępny kierunek Technik Farmaceutyczny Szkoła Policealna im. J. Romanowskiej 1) Podstawowe prawa i pojęcia chemiczne 2) Roztwory (zadania rachunkowe zbiór zadań Pazdro
Zagadnienia z chemii na egzamin wstępny kierunek Technik Farmaceutyczny Szkoła Policealna im. J. Romanowskiej 1) Podstawowe prawa i pojęcia chemiczne 2) Roztwory (zadania rachunkowe zbiór zadań Pazdro