Uniwersytet Rolniczy im. H. Kołłątaja w Krakowie Wydział Technologii Żywności
|
|
|
- Marta Szymczak
- 6 lat temu
- Przeglądów:
Transkrypt
1 Uniwersytet Rolniczy im. H. Kołłątaja w Krakowie Wydział Technologii Żywności Instytut Chemii Zakład Chemii Biopolimerów dr Anna Konieczna-Molenda ZAŁĄCZNIK 2 AUTOREFERAT W JĘZYKU POLSKIM Nowe metody aktywacji fizykochemicznej enzymów przemysłowych do przetwórstwa polisacharydów 1. Posiadane dyplomy, stopnie naukowe Informacje o dotychczasowym zatrudnieniu Wskazanie osiągnięcia naukowego Pozostałe osiągnięcia naukowe Tabelaryczne zestawienie dorobku naukowego...55
2 1. POSIADANE DYPLOMY, STOPNIE NAUKOWE Z PODANIEM NAZWY, MIEJSCA I ROKU ICH UZYSKANIA ORAZ TYTUŁU ROZPRAWY DOKTORESKIEJ. Praca doktorska: 2003 r. Uniwersytet Jagielloński, Wydział Chemii pt. Kinetyka degradacji celulozy w zakresie podwyższonych temperatur Promotor: Prof. dr hab. Andrzej Barański Praca wyróżniona przez Radę Wydziału Chemii UJ Praca magisterska: 1998 r. Uniwersytet Jagielloński, Wydział Chemii, Zakład Technologii Chemicznej, pt. Superabsorbery polimerowe. Otrzymywanie nowych środków sieciujących i ich charakterystyka (wynik bardzo dobry) Promotor: Prof. dr hab. Edgar Bortel Studia podyplomowe: 2011 r. Studia podyplomowe Dydaktyka na uczelni wyższej Politechnika Krakowska im. T. Kościuszki w Krakowie 2
3 2. INFORMACJE O DOTYCHCZASOWYM ZATRUDNIENIU W JEDNOSTKACH NAUKOWYCH umowa o dzieło w ramach Wieloletniego Projektu Rządowego pt. Kwaśny papier finansowanego przez MEN, Wydział Chemii, Uniwersytet Jagielloński, asystent naukowo-dydaktyczny, Katedra Chemii, Wydział Rolniczo - Ekonomiczny, Uniwersytet Rolniczy (Akademia Rolnicza) im. H. Kołłątaja w Krakowie, adiunkt naukowo-dydaktyczny, Zakład Chemii Biopolimerów, Instytut Chemii, Wydział Rolniczo-Ekonomiczny, Uniwersytet Rolniczy (Akademia Rolnicza) im. H. Kołłątaja w Krakowie, obecnie adiunkt naukowo-dydaktyczny, Zakład Chemii Biopolimerów, Instytut Chemii, Wydział Technologii Żywności, Uniwersytet Rolniczy im. H. Kołłątaja w Krakowie. 3
4 3. WSKAZANIE OSIĄGNIĘCIA NAUKOWEGO. WSKAZANIE OSIĄGNIĘCIA WYNIKAJĄCEGO Z ART. 16 UST. 2 USTAWY Z DNIA 14 MARCA 2003 ROKU O STOPNIACH NAUKOWYCH I TYTULE NAUKOWYM ORAZ O STOPNIACH I TYTULE W ZAKRESIE SZTUKI (DZ. U R. POZ. 882 ZE ZM. W DZ. U. Z 2016 R. POZ ): A) TYTUŁ OSIĄGNIĘCIA NAUKOWEGO/ARTYSTYCZNEGO, Cykl dziesięciu monotematycznych publikacji pod wspólnym tytułem: Nowe metody aktywacji fizykochemicznej enzymów przemysłowych do przetwórstwa polisacharydów B) (AUTOR/AUTORZY, TYTUŁ/TYTUŁY PUBLIKACJI, ROK WYDANIA, NAZWA WYDAWNICTWA, RECENZENCI WYDAWNICZY) B1. A. Konieczna-Molenda; (2012) Effect of illumionation of cellulose with the visible linearly polarized light upon digestion of cellulose, Recent. Devel. Carbohydrate Res, 3, 1-10 B2. A. Tata, K. Sokołowska, J. Świder, A. Konieczna-Molenda, E. Proniewicz, E. Witek, (2015) Study of cellulolytic enzyme immobilization on copolymers of N-vinylformamide, Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy, 149, IF = 2,653 B3. A. Para and Anna Konieczna Molenda; (2010) Starch dialdehyde from potato starch illuminated with linearly polarized white light; Carbohydrate Polymers, 79, (2) IF = 3,463 4
5 B4. K. Sokołowska, A. Konieczna-Molenda, E. Witek, (2016) Hydroliza odpadów celulozowych katalizowana enzymami celulolitycznymi immobilizowanymi na nośniku polimerowym, Polimery, 9, IF = 0,778 B5. M. Fiedorowicz, G. Khachatryan, A. Konieczna- Molenda, P. Tomasik, (2009) Formation of cyclodextrin with cyclodextringlucosyltransferase stimulated with polarized light, Biotechnoogy Progress, 25, (1), IF = 2,398 B6. A. Konieczna- Molenda, V. M. F. Lai, M. Fiedorowicz, G. Khachatryan, P. Tomasik; (2008) Polarized Light-stimulated Enzymatic Hydrolysis of Xylan Biotechnology Progress, 24, IF = 2,108 B7. A. Konieczna-Molenda, M. Fiedorowicz, P. J. Tomasik, (2010) Stimulation of Glucose Oxidase with White Linearly Polarized Light, Biotechnology Progress, 26(1) IF = 2,178 B8. A. Konieczna-Molenda, M. Fiedorowicz, W. Zhong, P. Tomasik, (2008) Polarized light-stimulated enzymatic hydrolysis of chitin and chitosan; Carbohydr. Res., 343, IF = 1,960 B9. M. Fiedorowicz, A. Konieczna-Molenda, G. Khachatryan, P. Tomasik; Sposób stymulowania katalizatorów białkowych używanych w reakcjach biochemicznych, zwłaszcza enzymów stosowanych w reakcjach otrzymywania sacharydów (PL ) patent przyznany w 2010r. B10. A. Konieczna-Molenda, Sposób określenia szybkości degradacji polisacharydów w produktach leczniczych, zwłaszcza w syropach ziołoleczniczych (PL ) patent przyznany w 2011r. 5
6 Całkowity IF z roku opublikowania prac wchodzących w skład osiągnięcia naukowego: 15,538. Liczba punktów MNiSW za cykl publikacji: 285 pkt. Oświadczenia współautorów prac określające wkład w powstanie publikacji zamieszczono w załączniku 5. C) OMÓWIENIE CELU NAUKOWEGO WW. PRAC I OSIĄGNIĘTYCH WYNIKÓW WRAZ Z OMÓWIENIEM ICH EWENTUALNEGO WYKORZYSTANIA. 4. OMÓWIENIE POZOSTAŁYCH OSIĄGNIĘĆ NAUKOWO - BADAWCZYCH Wstęp W krajach wysokorozwiniętych coraz więcej produktów otrzymywanych jest w procesach technologicznych z udziałem biokatalizatorów. W produkcji i przetwórstwie żywność na coraz większą skalę stosuje się enzymy jako katalizatory reakcji. Posiadają one wiele cech decydujących o atrakcyjności ich zastosowania, takich jak łatwość kontrolowania i regulacji katalizowanej reakcji, specyficzność, łagodność warunków działania (co w konsekwencji ogranicza straty cennych składników odżywczych), a także bezpieczeństwo stosowania wynikające z naturalnego pochodzenia i działania w niskich stężeniach. Enzymy odpowiedzialne są za przebieg wszystkich reakcji w żywych organizmach, bez ich udziału przebiegałyby one zbyt wolno. Katalizatory te przyspieszają zachodzenie reakcji chemicznych, poprzez obniżenie energii aktywacji. Powodują one zmniejszenie różnicy energii między reagentami, a stanem przejściowym [1]. Analogicznie, jak inne katalizatory enzymy nie zużywają się podczas reakcji, ani też nie wpływają na jej równowagę. W sposób ilościowy kinetykę reakcji enzymatycznych opisuje prawo Michaelisa-Menten [2]. Enzymy charakteryzują się specyficznością tzn. katalizują wyłącznie jeden określony rodzaj reakcji, wykazują specyficzność wobec określonej grupy funkcyjnej, substratu lub wiązania chemicznego. Za działanie katalityczne odpowiadają tylko miejsca aktywne enzymu, z tego powodu ich aktywność katalityczna uzależniona jest od ich trójwymiarowej struktury [3]. Funkcje katalityczne enzymów, ich swoistość, selektywność, zdolność do działania w łagodnych warunkach spowodowały duże zainteresowanie ze strony przemysłu. Po wyizolowaniu i oczyszczeniu są one powszechnie stosowane jako alternatywa dla 6
7 tradycyjnych chemicznych technologii. Badania dotyczące światowej sprzedaży enzymów wskazują że 31% to enzymy do celów spożywczych (głównie przetwórstwo skrobi i piekarnictwo), 6% enzymy paszowe, a pozostałe to enzymy techniczne (produkcja detergentów, tekstyliów, biopaliw) [4]. Kontrolowane reakcje z zastosowaniem enzymów lub grupy enzymów stanowią podstawę wielu procesów przetwórczych (np. hydrolizaty skrobiowe, białkowe) oraz operacji technologicznych. Obecnie z udziałem enzymów występujących w różnych mikroorganizmach wytwarza się wiele produktów spożywczych (pieczywo, napoje alkoholowe, soki, sery, jogurty, słodycze i inne) [5]. Dzięki biotechnologii i zastosowaniu enzymów przemysł spożywczy wprowadza nowe produkty, obniża ich koszty i poprawia ich jakość, usprawnia już istniejące procesy produkcyjne i przetwórcze [6]. Wykorzystanie coraz to nowych enzymów przekłada się na zwiększenie funkcjonalności artykułów spożywczych, poprawę wartości odżywczych, własności sensorycznych, takich jak tekstura czy smak produktów. Pozwala także, na obniżenie kosztów i ulepszenie procesów przetwarzania żywności oraz utylizacji odpadów w przemyśle spożywczym. Zastąpienie tradycyjnych chemicznych technologii reakcjami enzymatycznymi przynosi korzyści w zakresie ochrony środowiska, ze względu na biodegradowalność i obniżenie zużycia energii [7]. Niestety, enzymy oprócz doskonałych właściwości, posiadają cechy które, nie są pożądane w przemyśle, takie jak: wysoka rozpuszczalność, brak stabilności i podatność na inhibicję, hamowanie przez substraty lub produkty reakcji, wysoka cena spowodowana skomplikowaną izolacją i oczyszczeniem. Wdrożenie enzymów, jako katalizatorów przemysłowych, jest zadaniem interdyscyplinarnym i wymaga wyselekcjonowania enzymów o odpowiednich właściwościach, ulepszenia ich za pomocą technik biomolekularnych i fizykochemicznych oraz dostosowania reaktorów przemysłowych do ich stosowania [8,9]. Enzymy używane w przemyśle pozyskiwane są z tkanek roślinnych, zwierzęcych lub z drobnoustrojów. Obecnie najczęściej, stosowane są enzymy pochodzenia mikrobiologicznego, otrzymywane z kultur bakterii, grzybów pleśniowych lub drożdży. 7
8 Tabela 1. Wykorzystanie przykładowych enzymów w przemyśle spożywczym [10]. Enzym Sposób działania Zastosowanie w przemyśle spożywczym α-amylaza hydroliza skrobi - fermentacyjnym, (wiązań α-1,4- - piwowarskim (słodowanie), glikozydowych) - piekarskim, - wytwarzanie cukrów β-amylaza hydroliza skrobi (wiązań α- - wytwarzanie syropu słodowego 1,6-glikozydowych) glukoamylaza hydroliza dekstryn - scukrzanie skrobi, skrobiowych do glukozy - produkcja piwa (wiązań α-1,4 oraz α-1,6- glikozydowych) celulaza hydroliza celulozy - upłynnianie owoców podczas produkcji soków - w piekarnictwie - w przemyśle gorzelniczym glukozyltransferaza hydroliza skrobi i produkcja - produkcja cyklodekstryn cyklodekstynowa cyklodekstryn chitynaza hydroliza chityny - otrzymywanie glukozaminy chitozanaza hydroliza chitozanu - otrzymywanie N-acetylo glukozaminy hemicelulaza i hydroliza hemiceluloz - hydroliza nie skrobiowych polisacharydów w mące, ksylanaza - poprawa jakości pieczywa - warzenie piwa β-galaktozydaza hydroliza laktozy - produkcja nabiału bez laktozy, (laktaza) - dosładzanie produktów mlecznych β-glukanaza hydroliza β-glukanów - produkcja piwa oksydaza glukozowa utlenianie glukozy do kwasu - usuwanie tlenu z opakowań spożywczych, glukonowego - eliminacja glukozy z białka jaj pektynaza hydroliza pektyn - klarowanie soków pullanaza hydroliza wiązań α-1,6- - scukrzanie skrobi, glikozydowych - zwiększenie wydajności hydrolizy skrobi asparaginaza hydroliza grupy amidowej w - redukcja zawartości akryloamidu w produktach łańcuchu bocznym spożywczych poddawanych obróbce cieplnej asparaginy lipaza hydroliza tłuszczu - przyspieszanie dojrzewania sera, - modyfikacja struktury tłuszczów mlecznych, - sery modyfikowane enzymatycznie pepsyna hydroliza kazeiny koagulacja mleka przy wytwarzaniu serów 8
9 trypsyna hydroliza białek - hydroliza białek spożywczych, - wytwarzanie hydrolizatu do przyprawiania żywności Bogate źródło różnorodnych produktów, stanowią najbardziej dostępne i rozpowszechnione w przyrodzie polisacharydy: skrobia, celuloza, ksylan oraz chityna. Budowa tych polisacharydów determinuje ich stosunkowo wysoką odporność na degradację, a ich hydroliza wymaga zastosowania środków chemicznych lub kompleksu enzymów. Skrobia będąca związkiem zapasowym roślin jest powszechnie występującym surowcem odnawialnym. Rośliny zawierające skrobię (takie jak pszenica, ryż, ziemniaki, kukurydza, tapioka, sago i inne) są na szeroką skalę wykorzystywane przez człowieka. Od ponad 40 lat do przetwarzania skrobi na skalę przemysłową wykorzystuje się enzymy. Enzymy działające na skrobię można zasadniczo podzielić na: 1) hydrolazy egzo- i endo-, które rozkładają wiązania α-1,4- i α-1,6-glikozydowe, 2) glukotransferazy działające na wiązania α-1,4- i α-1,6-glikozydowe, 3) oksydoreduktazy katalizujące utlenianie i redukcję poszczególnych grup [11]. Celuloza jest składnikiem budulcowym ściany komórkowej roślin wyższych oraz niektórych glonów, grzybów i bakterii. To nierozgałęziony biopolimer, zbudowany z cząsteczek glukozy połączonych wiązaniami β-1,4-glikozydowymi [12]. Hydroliza celulozy wymaga współdziałania kilku enzymów. W zależności od typu katalizowanej reakcji można wyróżnić pięć podstawowych rodzajów celulaz: 1) endo-(1,4) β-d-glukanazy (EC ) hydrolizują wewnętrzne wiązania, niszcząc strukturę krystaliczną celulozy i odsłaniają pojedyncze łańcuchy, 2) egzo-(1,4) β-d-glukanazy (EC ) katalizują odrywanie krótkich fragmentów (2-4 reszt glukozowych) od końców odsłoniętych łańcuchów, 3) celobiazy (β-glukozydaza; EC ) rozkładają do pojedynczych jednostek glukozowych produkty uwolnione przez endo- i egzocelulazy, 4) celulazy utleniające, degradują celulozę w reakcjach rodnikowych, 5) fosfataza celulozy, depolimeryzuje celulozę z nie wody lecz fosforanu. Ksylan to jeden z najobficiej występujących na ziemi polisacharydów, należy do hemiceluloz, które wraz z ligniną i celulozą wchodzą w skład kompleksu ligninocelulozowego w ścianach komórkowych roślin. Ksylan stanowi ważny surowiec do wykorzystania na drodze biokonwersji, bowiem powstające w wyniku jego hydrolizy enzymatycznej produkty stanowią punkt wyjścia do produkcji żywności, pasz, paliw i 9
10 chemikaliów. Ksylan zbudowany jest z reszt β-d-ksylopiranozy połączonych wiązaniami glikozydowymi między 1 i 4 atomami węgla. Całkowita hydroliza ksylanu wymaga zastosowania kilku enzymów: 1) endo-β-1,4- ksylanazy (EC ) hydrolizują liniowe wiązania β-1,4-glikozydowe. Efektem ich działania są podstawione lub nie oligomery, ksylobioza lub ksyloza, 2) egzo-β-1,4-dksylozydazy (ksylanohydrolaza β-1,4-d-ksylanu; EC ) odszczepiają reszty D-ksylozy z nieredukujących końców ksylooligosacharydów, hydrolizuje liniowe wiązania, 3) α-dglukuronidazy (EC 3.2.1) hydrolizują wiązania uwalniając kwas D-glukuronowy lub kwas 4- O-metylo-α-D-glukuronowy, 4) α-l-arabinofuranozydazy (EC ) z nieredukujących końców odszczepią reszty L-arabinofuranozowe, hydrolizują zarówno wiązania α-1,3- jak α- 1,5-arabinoruranozowe, 5) esterazy octanowej lub acetyloksylanoesterazy usuwają grupy acetylowe, co zwiększa podatność na endo-β-1,4-ksylanaz. Chityna jest liniowym biopolimeren naturalnym zbudowanym z reszt N-acetylo-Dglukozo-2-aminowych połączonych wiązaniami β-1,4-glikozydowymi. W przyrodzie występuje jako materiał budulcowy szkieletów zewnętrznych stawonogów, a także u ramienionogów, mszywiołów i mięczaków, a ponadto w ścianach komórkowych niektórych grzybów, wodorostów i bakterii. Głównym zastosowaniem chityny jest otrzymywanie glukozaminy w wyniku hydrolizy oraz chitozanu na drodze częściowej deacetylacji. W przeciwieństwie do chityny, chitozan pojawia się w przyrodzie bardzo rzadko. Najbardziej popularną metodą przetwarzania chityny i chitozanu jest ich hydroliza enzymatyczna z zastosowaniem odpowiednio chitynaz i chitozanaz. Wybór enzymów do przemysłowego przetwarzania polisacharydów ukierunkowany jest na uzyskanie jak najkorzystniejszych osiągnięć w ich zastosowaniu, a wymagania stawiane enzymom do celów przemysłowych są stosunkowo wysokie [13]. Głównymi problemami wykorzystywania enzymów w przemyśle jest ich wysoki koszt oraz często trudna dostępność katalizatorów o ściśle sprecyzowanych właściwościach. Enzymy działają tylko na niektóre substraty, w określonych warunkach (ph, temperatura, obecność inhibitorów), mogą posiadać niską aktywność, niską rozpoznawalność substratu (wysokie wartości K M ). W takiej sytuacji należy poszukiwać nowych katalizatorów lub już dostępne poddawać modyfikacjom. W celu otrzymania nowych katalizatorów stosuje się wiele technik pozyskiwania materiału, modyfikacje genetyczne, inżynierię białek (np. rekombinację DNA). Organizmy transgeniczne i inżynieria białek, mimo wykorzystywania na całym świecie podlegają ściśle określonym regułom prawnym i budzą wiele wątpliwości. Dodatkowo, strategia pozyskiwania 10
11 nowych katalizatorów enzymatycznych jest często bardzo żmudna i kosztowna. Stosunkowo prostsze i mniej kosztowne są modyfikacje fizykochemiczne enzymów już stosowanych w przemyśle. Zastosowanie takich modyfikowanych fizykochemicznie katalizatorów nie wymaga konieczności zmiany technologii, infrastruktury przemysłowej czy wprowadzania nowych procesów produkcyjnych. Obecnie w przemyśle coraz popularniejsze są enzymy w formie immobilizowanej, co z jednej strony najczęściej obniża ich aktywność, natomiast z drugiej wielokrotnie zwiększa stabilność oraz zapewnia ciągłość i stałą kontrolę procesu produkcyjnego. Immobilizacja enzymów, białek, mikroorganizmów polega na unieruchomieniu, fizycznym ograniczeniu, zatrzymaniu preparatu w reaktorze [14]. Celem immobilizacji jest przeniesienie zalet katalizy heterogenicznej na rozpuszczalne katalizatory enzymatyczne. Immobilizacja enzymów posiada wiele zalet takich jak zwiększenie stabilności w szerszym zakresie ph i temperatury, możliwość użycia w wielu cyklach reakcji lub w procesach ciągłych, łatwiejsze oddzielenie produktów, możliwość immobilizacji kilku enzymów jednocześnie, prostsza praca reaktora i duży ich wybór. Głównymi wadami są utrata bądź zmniejszenie aktywności, ograniczenie dyfuzji oraz dodatkowe koszty związane z produkcją i aktywacją nośnika oraz mechanizmem unieruchamiania [15]. Znanych jest wiele metod immobilizacji enzymów, w zależności od typu wiązania (kowalencyjne, adsorpcyjne, uwięzienie w matrycy, mikrokapsułkowanie, sieciowanie), odwracalne i nieodwracalne [16,17]. W literaturze opisano wiele przykładów nośników do unieruchamiania enzymów: naturalne, syntetyczne, organiczne, nieorganiczne [18-20]. Poszczególne immobilizacje posiadają swoje zalety i wady, dlatego dla każdego enzymu i każdego substratu należy dobrać odpowiedni nośnik i metodę immobilizacji. Korzystnym sposobem unieruchomienia enzymu na nośniku jest wiązania kowalencyjne. Pozwala na silne umocowanie enzymu, zapobiega jego odszczepieniu ułatwiając jednocześnie kontakt z substratem, gdyż enzym eksponowany jest na powierzchni [21]. Ponadto wiązanie kowalencyjne enzymu z nośnikiem często zwiększa odporność temperaturową takiego biokatalizatora. Grupami funkcyjnymi nośników najczęściej wykorzystywanymi w procesie tworzenia wiązania kowalencyjnego z białkiem są: hydroksylowa, karboksylowa, aminowa i oksiranowa [22]. Enzym jest polipeptydem ze środkową częścią bardziej zwartą, a zewnętrzną dość labilną [23]. Właśnie w części zewnętrznej mieści się aktywne centrum katalityczne oraz grupy funkcyjne zdolne do tworzenia wiązań kowalencyjnych. Grupa ε-aminowa pochodzi od 11
12 lizyny, merkaptanowa od cysteiny, grupy β- i γ- karboksylowe pochodzą odpowiednio od kwasów asparginowego i glutaminowego. Takich ugrupowań mieści się w zewnętrznej warstwie enzymu bardzo dużo. Silne związanie enzymu z nośnikiem może powodować zjawisko obniżenia, a w skrajnym przypadku utraty aktywności katalitycznej spowodowane blokowaniem dostęp substratu do centrum aktywnego. Aby temu zjawisku zapobiec stosuje się tzw. łącznik w postaci tzw. spacera, którym jest najczęściej aldehyd glutarowy. Jest on nietoksyczny, a dodatkowo bakterio- wiruso- i grzybobójczy. Nieoczekiwanie okazało się, że zastosowanie w reakcji enzymu naświetlanego światłem widzialnym liniowo spolaryzowanym ma wpływ na wydajność i selektywność hydrolizy polisacharydów [24]. Polaryzacja to obok barwy i natężenia jedna z cech światła. Występują trzy rodzaje polaryzacji światła: liniowa, kołowa i eliptyczna. Powszechnie wiadomo, że światło wpływa na procesy biologiczne. Znane są także biologiczne efekty oddziaływania światła spolaryzowanego na organizmy żywe [25,26]. Ujawniono korzystne oddziaływanie światła spolaryzowanego o określonej barwie na ludzki organizm, co znalazło zastosowanie w medycynie i kosmetyce [27], w leczeniu oparzeń [28,29], leczeniu ran [30,31]. Wykryto także wpływ promieniowania spolaryzowanego na hydrolizę skrobi, co związane było z aktywacją towarzyszących jej amylaz [32]. Cel i zakres badań Celem badań stanowiących podstawę do ubiegania się o stopień naukowy doktora habilitowanego, których wyniki przedstawiłam w cyklu monotematycznych prac B1-B10 było otrzymanie nowych enzymatycznych biokatalizatorów aktywowanych fizykochemicznie, aktywnych w reakcjach przemiany polisacharydów i mogących znaleźć zastosowanie w przemyśle spożywczym. W wyniku fizykochemicznej aktywacji handlowo dostępnych i powszechnie stosowanych w przemyśle enzymów: stymulacji światłem spolaryzowanym lub immobilizacji na nośnikach otrzymano biokatalizatory enzymatyczne o zmienionej selektywności, wyższej aktywności, stabilności, tolerancji temperaturowej i ph w porównaniu z enzymami natywnymi. Określono kluczowe parametry stymulacji światłem oraz immobilizacji wpływające na właściwości biokatalizatorów. Otrzymane biokatalizatory zostały zastosowane i przetestowane pod względem funkcjonalności w reakcjach przemiany sacharydów. Biokatalizatory takie mogą być użyte w 12
13 przemyśle (spożywczym, farmaceutycznym, papierniczym, rafineryjnym), do utylizacji odpadów, oczyszczania ścieków oraz jako biosensory. Zastosowanie biokatalizatorów o wysokiej aktywności katalitycznej i stabilności w postaci enzymów stymulowanych elektromagnetycznie lub immobilizowanych na nośnikach polimerowych pozwoli zmniejszyć koszty wielu operacji jednostkowych w przemyśle. Nowe biokatalizatory wpisują się w założenia Zielonej Chemii, a ich zastosowanie w procesach produkcji i przetwarzania żywności może dostarczyć alternatywnych, ekologicznych produktów. Metody badawcze Enzymy handlowo dostępne i posiadające zastosowanie w przemyśle poddawano fizykochemicznej aktywacji. Aktywacja fizyczna polegała na naświetlaniu enzymów światłem widzialnym liniowo spolaryzowanym. Aktywacja chemiczna odbywała się poprzez unieruchomienie enzymów na odpowiednio dobranym nośniku. Naświetlanie enzymów światłem widzialnym liniowo spolaryzowanym Enzym w postaci zawiesiny umieszczano w kwarcowych celach o szerokości 1cm i naświetlano z odległości 30 cm z naświetlacza szczelinowego KB 502 (Kabid, Chorzów, Polska) wyposażonego w łuk ksenonowy 150W (XBO 150 Oriel, Maidstone, UK). Stosowano filtr o polaryzacji liniowej HN 22 filtr (Polaroid, Waltham, MA). Źródło światła emitowało ciągłe promieniowanie w zakresie widzialnym nm. Jego strumień energii w pozycji próbki wynosił 4, 8, 16 lub 18 mw/cm 2 (w zależności od enzymu), jak sprawdzono przez Radiometr YSI (Yellow Springs, OH). Próbki były naświetlane w atmosferze azotu, w stałej temperaturze 25 C przez 30, 45, 60, 120 lub 300 min. Biokatalizatory otrzymane w wyniku naświetlania światłem widzialnym liniowo spolaryzowanym testowano w reakcjach przemiany polisacharydów. Działanie katalizatorów stymulowanych światłem widzialnym liniowo spolaryzowanym porównywano z działaniem enzymów naświetlanych światłem nie spolaryzowanym lub enzymów nie naświetlanych. 13
14 Rys. 1. Przykładowy schemat naświetlania próbki enzymu światłem widzialnym liniowo spolaryzowanym. Immobilizacja enzymów Dla każdego z enzymów immobilizowanych na danym nośniku opracowano indywidualną metodę immobilizacji z uwzględnieniem temperatury, ph, czasu, sekwencji i rodzaju reakcji. Enzymy unieruchamiano na powierzchni nośnika bezpośrednio lub za pomocą łącznika aldehydu glutarowego. Przeważająca ilość immobilizacji polegała tworzeniu kowalencyjnych, wiązań iminowych nośnik-enzym lub nośnik-łącznik-enzym, ale też enzymy były otoczkowane w hydrożelu. Badano wydajność immobilizacji, stabilność wiązania nośnik-enzym oraz zawartość enzymu na powierzchni nośników metodą Lowry ego [33]. W przeprowadzonych badaniach korzystano z wielu metod fizykochemicznych w celu charakterystyki nośników, biokatalizatorów i analizy rezultatów reakcji: 1. Spektroskopia UV-Vis analiza produktów reakcji enzymatycznych oraz badania nośników enzymów, 2. Chromatografia HPLC analiza produktów reakcji, 3. Spektroskopia FT-RAMAN badania nośników enzymów oraz sposobu wiązania nośnikłącznik-enzym, 4. Spektroskopia FT-IR badania nośników enzymów, 5. Analiza DSC wyznaczanie zawartości wody, 6. Wiskozymetria i metody reologiczne wyznaczanie stopnia polimeryzacji polisacharydów, 7. Analiza termograwimetryczna w połączeniu ze spektrometrią masową (MS- TG/DTG/SDTA) badania nośników enzymów, 8. Dyfrakcja Roentgena (X-ray diffractograms) określanie struktury krystalograficznej, 14
15 9. Pomiar bezwzględnych mas cząsteczkowych oraz promieni bezwładności metoda wysokociśnieniowej chromatografii żelowej z użyciem detektorów refraktometrycznego oraz wielokątowego rozpraszania światła laserowego (HPSEC-MALLS-RI), 10. Metoda BET oznaczanie powierzchni właściwej nośników, 11. Analiza elementarna badania nośników enzymów, 12. Wysokorozdzielcza mikroskopia skaningowa SEM zdjęcia nośników i substratów. Materiały Fizykochemicznej aktywacji poddano stosowane w przemyśle i ogólnodostępne enzymy: a) termostabilna α-amylaza EC : SPEZMER PRIME (Genencor International, USA) z genetycznie modyfikowanych Geobacillus stearothermophilus oraz α- amylaza z Bacillus licheniformis EC (Sigma-Aldrich), b) glukoamylaza z grzybów EC (Genencor International, USA), c) glukozyltransferaza cyklodekstynowa (CGTase) z Thermoanaerobacter species EC (Novozyme), d) oksydaza glukozowa z Aspergillus niger E.C (Sigma-Aldrich), e) ksylanaza Thermomyces lanuginosus EC (Sigma-Aldrich), f) chitozanaza z Streptomyces species EC (Sigma-Aldrich), g) chitynaza z Serratia marcescens EC (Sigma-Aldrich), h) celulazy EC : celulazy z Aspergillus species (Sigma-Aldrich), Novozym 476 (Sigma-Aldrich), Viscozyme L multienzymatyczny kompleks karbohydraz (m.in. celulaz EC , β-glukanaz EC , hemicelulaz EC , ksylanaz EC ) w postaci roztworu (Sigma Aldrich). Polimery służące jako nośniki do immobilizacji wykonano w ramach współpracy w Zespole Polimerów na Wydziale Chemii Uniwersytetu Jagiellońskiego. Stosowane do immobilizacji enzymów nośniki polimerowe charakteryzowały się zazwyczaj sferycznym kształtem i posiadały na powierzchni pierwszorzędowe grupy aminowe podstawione do łańcuchów polimerowych, które stanowią silnie reaktywne centra dające się łatwo modyfikować w kontrolowanych reakcjach chemicznych [34]. Ilość grup aminowych zdolnych do wiązania enzymu miała znaczący wpływ, czy enzym wiązany był jednopunktowo czy wielopunktowo i decydowała o jego właściwościach katalitycznych. 15
16 Sferyczny kształt ziaren nośnika jest natomiast szczególnie pożądany, gdy proces hydrolizy prowadzony jest w reaktorze kolumnowym. Omówienie wyników Reakcje przetwarzania skrobi W patencie (B9) ujawniono sposób i efekty stymulacji światłem widzialnym liniowo spolaryzowanym enzymu 1,4-α-D-glukan glukohydrolazy (glukoamylaza, amyloglukozydaza EC ) powszechnie stosowanej w przemyśle do hydrolizy skrobi. Zastosowanie enzymu krótko stymulowanego przez 60 minut w reakcji hydrolizy skrobi spowodowało wzrost wydajności reakcji zaledwie w początkowym czasie reakcji tj. po 15 i 30 minutach hydrolizy (rys. 2). enzym nienaswietlany enzym nawietlany swiatlem spolaryzowanym przez 1godz. stezenie glukozy c [mg/ml] czas reakcji enzymatycznej t [min] Rys. 2. Zależność stężenia glukozy od czasu reakcji hydrolizy skrobi sago z zastosowaniem glukoamylazy: - nie naświetlanej, - naświetlanej światłem widzialnym spolaryzowanym przez 60 min. (B9). Podjęto także próbę aktywacji glukamylazy w wyniku immobilizacji na nośniku polimeru hydrożelowego [35]. Uzyskano biokatalizator aktywny w reakcji hydrolizy skrobi sago w dwóch cyklach reakcji, ale o niższej aktywności katalitycznej niż enzym w stanie wolnym. Korzystną właściwością biokatalizatora była natomiast jego stabilność. 16
17 Biokatalizator przechowywany przez 30 dni (temp.4 o C) posiadał wyższą aktywność katalityczną niż zastosowany bezpośrednio po immobilizacji (rys. 3) Glukoza [mg/ml] glukoamylaza immobilizowana - cylk I immobillizowana - cykl II immobilizowana - po 30 dniach nie immobilizowana Czas [min] Rys. 3. Zależność stężenia glukozy od czasu reakcji hydrolizy skrobi sago z zastosowaniem glukoamylazy: - - nie immobilizowanej, biokatalizatora z immobilizowanym enzymem: - - I cykl reakcji,.; - - II cykl reakcji, - - zastosowany po 30 dniach przechowywania. Warto zaznaczyć, że niższa aktywność glukoamylaz immobilizowanych związana jest z utworzeniem niekowalencyjnego wiązania z nośnikiem hydrożelowym, polegającego na otoczeniu i zamknięciu w żelu stosunkowo niewielkich rozmiarów enzymu jakim jest glukoamylaza. Powolne uwalnianie enzymu z hydrożelu skutkuje wyższą aktywnością po 30 dniach przechowywania. Światłem widzialnym liniowo spolaryzowanym stymulowano oksydazę glukozową (GOD) typii-s z Aspergillus niger (B7). Oksydaza glukozowa (E.C ) jest oksydoreduktazą, która katalizuje utlenianie β-d-glukozy do kwasu D-glukonowego. Enzym ten posiada największe znaczenie w przemyśle spożywczym oraz w zastosowaniach analitycznych, gdzie kontrolowane jest stężenie glukozy. W przemyśle spożywczym enzym ten jest powszechnie używany do eliminacji tlenu i glukozy z produktów spożywczych oraz jako biosensor do oznaczania β-d-glukozy w żywności, substratach do produkcji żywności oraz w produktach pośrednich otrzymywanych w procesach technologicznych. 17
18 Oksydazę glukozową naświetlano przez 30, 60 i 120 minut światłem widzialnym liniowo spolaryzowanym oraz w celu porównania, także światłem widzialnym nie spolaryzowanym o takim samym strumieniu energii. Oksydaza glukozowa była podatna na stymulację światłem widzialnym liniowo spolaryzowanym. Najwyższą aktywność enzymu uzyskano w wyniku naświetlania światłem przez 60 minut, a stała szybkości reakcji utleniania glukozy była w tym przypadku blisko dwukrotnie wyższa niż dla enzymu nie stymulowanego (rys. 4). 10 D-gluconic acid [mg/ml] GOD illuminated polarized light 60 min GOD illuminated polarized light 120 min GOD illuminated polarized light 30 min GOD non-illuminated Reaction time [min] Rys. 4. Zależność stężenia kawasu D-glukonowego od czasu reakcji utleniania glukozy z zastosowaniem oksydazy glukozowej (GOD): - - nie naświetlanej, naświetlanej światłem widzialnym liniowo spolaryzowanym przez: min., min., min. (B7). W wyniku stymulacji światłem widzialnym liniowo spolaryzowanym przez 60 minut biokatalizator zyskał większą stabilność temperaturową i ph w porównaniu do enzymu natywnego (rys. 5 i rys. 6). 18
19 0.35 GOD illuminated polarized light 60 min GOD non-illuminated Activity [(mg/ml)/min] Temperature [ o C] Rys. 5. Zależność aktywności oksydoreduktazy glukozowej od temperatury reakcji dla enzymu: - - nie naświetlanego oraz - - naświetlanego światłem widzialnym liniowo spolaryzowanym (B7) GOD illuminated polarized light 60 min GOD non-illuminated Activity [(mg/ml)/min] ph Rys. 6. Zależność aktywności oksydoreduktazy glukozowej od ph reakcji dla enzymu: - - nie naświetlanego oraz - - naświetlanego światłem widzialnym liniowo spolaryzowanym (B7). 19
20 Dodatkowo oksydoreduktazę glukozową bezpośrednio po stymulacji światłem poddano procesowi liofilizacji. Liofilizowany biokatalizator wykazywał w reakcji zaledwie 30% spadek aktywności katalitycznej w porównaniu do enzymu nie liofilizowanego. W wyniku stosunkowo niedługiej (60min.) stymulacji światłem widzialnym liniowo spolaryzowanym oksydoreduktazy glukozowej uzyskano biokatalizator o znacznie wyższej aktywności oraz większej stabilności temperaturowej i ph. Stymulację światłem widzialnym liniowo spolaryzowanym można z sukcesem stosować także dla mieszaniny enzymów. W pracy B9 przedstawiono reakcje jednoczesnej hydrolizy i utleniania skrobi z zastosowaniem mieszaniny enzymów stymulowanych światłem. Sporządzono mieszaninę enzymów: glukoamylazy (EC ) oraz oksydazy glukozowej (E.C ), które naświetlano światłem widzialnym liniowo spolaryzowanym. Hydroliza skrobi z zastosowaniem katalizatora uzyskanego w wyniku naświetlania enzymów przebiega z większą szybkością niż w przypadku zastosowania mieszaniny enzymów nie naświetlanych. Stymulacji światłem widzialnym liniowo spolaryzowanym poddano glukozyltransferazę cyklodekstynową (CGTase) (EC ), która katalizuje otrzymywanie ze skrobi cyklicznych, liniowych α-(1-4) oligosacharydów α-, β-, γ-cyklodekstryn zawierających odpowiednio 6, 7 i 8 jednostek α-d-glukozowych (B9 i B5). Produktem reakcji z zastosowaniem CGTase jest mieszanina cyklodekstryn, wymagająca rozdziału na poszczególne cyklodekstryny. Na skalę przemysłową CGTase używana jest do produkcji cyklodekstryn oraz w celu upłynniania skrobi [36]. Budowa cyklodekstryn korzystnie wpływa na możliwość tworzenia kompleksów z różnymi innymi cząsteczkami. Z tego powodu cyklodekstryn znajdują bardzo szerokie zastosowanie w przemyśle spożywczym, ale także farmaceutycznym, kosmetycznym, chemicznym a także w rolnictwie [37,38]. W przemyśle spożywczym cyklodekstryny stosowane są do pochłaniania niekorzystnych zapachów i smaków, wiązania substancji lotnych czy też toksycznych powstających podczas przechowywania. Cyklodekstryny mają też za zadanie utrwalać pożądany smak, zapach i barwę wybranych produktów. W wyniku reakcji otrzymuje się zazwyczaj najwięcej α- lub β-cyklodekstryn, a bardzo niewielkie ilości γ-cykolodekstryn. Wykonano reakcje hydrolizy o niskich stężeniach skrobi i enzymu dla których, optymalny czas stymulacji światłem wynosił 60 minut. Stymulacja światłem zmieniła 20
21 selektywność reakcji, w której otrzymano przeważające ilości β-cyklodekstryn, podczas kiedy zastosowanie enzymu nie naświetlanego dawało najwięcej α-cyklodekstryn (rys. 7). 0,25 CD CD CD c s =0,01 g/ml c e =0,004 ml/ml t r =2 h T=37 o C CD g/1g sago 0,20 0,15 0,10 0,05 0, czas naświetlania enzymu [min] Rys. 7. Zależność stężenia α-, β-, γ-cyklodekstryn od czasu naświetlania enzymu zastosowanego w reakcji przez 2godz. (małe stężenie skrobi i enzymu) (B9). W reakcjach hydrolizy o wyższych stężeniach skrobi i enzymu optymalna stymulacja światłem wynosiła 2 godziny i powodowała zwiększenie całkowitej wydajności reakcji. Znacznie wzrosła także ilość otrzymanych β-cyklodekstryn i γ-cykolodekstryn (rys. 8). 21
22 0,6 0,5 CD CD CD c s =0,02 g/ml c e =0,008 ml/ml t r =2 h T=37 o C CD g/1g sago 0,4 0,3 0,2 0, czas naświetlania enzymu [h] Rys. 8. Zależność stężenia α-, β-, γ-cyklodekstryn od czasu naświetlania enzymu zastosowanego w reakcji przez 2godz. (duże stężenie skrobi i enzymu) (B9). Dodatkowo określono trwałość stymulacji glukozyltransferazy cyklodekstynowej światłem widzialnym liniowo spolaryzowanym. Stymulowany enzym przechowywano przez 90 dni w temperaturze 4 o C bez dostępu światła, a następnie zastosowano w reakcji hydrolizy i tworzenia cyklodekstryn. Uzyskano podobne rezultaty jak w przypadku zastosowania enzymu bezpośrednio po naświetlaniu (rys. 9). 22
23 Cyklodekstryny [g/1g sago] cyklodekstryna cykoldekstryna cyklodekstryna Czas naświetlania enzymu [min] Rys. 9. Zależność stężenia α-, β-, γ-cyklodekstryn od czasu naświetlania enzymu zastosowanego w reakcji przez 2godz. (małe stężenie skrobi i enzymu). Enzym po aktywacji światłem przechowywano przez 90dni (B9). W wyniku stymulacji światłem widzialnym liniowo spolaryzowanym glukozyltransferazy cyklodekstynowej używanej powszechnie w przemyśle uzyskano biokatalizator o wyżej aktywności oraz odmiennej selektywności w stosunku do otrzymywanych produktów. Podjęto także próbę aktywacji glukozyltransferazy cyklodekstynowej w wyniku immobilizacji na nośniku polimeru hydrożelowego [35]. Uzyskano biokatalizator aktywny w reakcji hydrolizy skrobi sago w dwóch cyklach reakcji, ale o niższej aktywności katalitycznej niż enzym w stanie wolnym. Przechowywanie biokatalizatora przez 30 dni (temp.4 o C) nie wpływało na zmianę jego aktywności (rys. 10). 23
24 Cyklodekstryny [mg/ml] cyclodekstryn glycosyltransferaza immobilizowana - cykl I immobilizowana - cykl Il immobilizowana - po 30 dniach nie immobilizowana Czas [min] Rys. 10. Zależność stężenia cyklodekstryn w przeliczeniu na β-cyklodekstryny od czasu reakcji z zastosowaniem enzymu: - - nie immobilizowanego, biokatalizatora z immobilizowanym enzymem: - - I cykl reakcji,.; - - II cykl reakcji, - - zastosowany po 30 dniach przechowywania. Opisano sposób otrzymywania i działania biokatalizatorów do reakcji hydrolizy skrobi opartych na sześciu poliwinyloamidowych i trzech poliwineloforamidowych polimerach hydrożelowych, na których immobilizowano termostabilną α-amylazę [50]. Enzym był kowalencyjnie wiązany z nośnikami przy użyciu aldehydu glutarowego jako łącznika. Procedura unieruchamiania została zoptymalizowana z uwzględnieniem temperatury, ph, czasu, sekwencji i rodzaju reakcji dla każdego z nośników. Określono także wydajność i trwałość immobilizacji. Wszystkie uzyskane biokatalizatory były aktywne w reakcjach hydrolizy skrobi. Enzymy unieruchomione na dwóch z zastosowanych nośników (nie hydrolizowany i poddany hydrolizie zasadowej) wykazywały wyższą aktywność niż enzym w stanie wolnym i były aktywne katalitycznie w dwóch cyklach reakcji. Hydrolizę skrobi prowadzono także z zastosowaniem α-amylazy z Geobacillus stearothermophilus naświetlanej światłem widzialnym liniowo spolaryzowanym oraz światłem nie spolaryzowanym. Stwierdzono wzrost aktywności katalitycznej biokatalizatora po ekspozycji na światło spolaryzowane. Efektu stymulacji nie było w przypadku naświetlania światłem nie spolaryzowanym (rys. 11). 24
25 1.0 Maltoza [mg/1mg sago] k 1 =3, k 1 =2, k 1 =4, k 2 =7, k 2 =11, k 2 =8, nonilluminated nonpolarized linearly polarized Czas reakcji [min] Rys. 11. Zależność stężenia maltozy od czasu reakcji hydrolizy skrobi sago z zastosowaniem α-amylazy: - - nie naświetlanej, - - naświetlanej światłem nie spolaryzowanym, - - naświetlanej światłem widzialnym liniowo spolaryzowanym. Dwie α-amylazy, ale różnego pochodzenia: termostabilną z Geobacillus stearothermophilus oraz α-amylaze z Bacillus licheniformis immobilizowano na nośniku polimeru hydrożelowego [35]. Biokatalizator otrzymany w wyniku unieruchomienia termostabilnej α-amylazy z Geobacillus stearothermophilus pracował w dwóch cyklach reakcji hydrolizy skrobi sago i posiadał wyższą aktywność niż enzym w stanie wolnym (rys. 12). 25
26 1.8 Maltoza [mg/ml] immobilizowany enzym cykl I immobilizowany enzym cykl II nie immobilizowany enzym Czas [min] Rys. 12. Zależność stężenia maltozy od czasu reakcji hydrolizy skrobi z zastosowaniem α-amylazy z Geobacillus stearothermophilus; - - enzym nie immobilizowany, biokatalizator z enzymem immobilizowanym: - - pierwszy cykl reakcji, - - drugi cykl reakcji [35]. Biokatalizator otrzymany w wyniku unieruchomienia α-amylazy z Bacillus licheniformis posiadał niższą aktywność katalityczną niż enzym nie immobilizowany. Przechowywanie biokatalizatora przez 60 dni (w temp.4 o C) spowodowało dodatkowe obniżenie jego aktywności (rys. 13). Immobilizacja tej α-amylazy wpłynęła na obniżenie aktywności katalitycznej oraz stabilności. 26
27 1.0 Maltoza [mg/ml] amylaza immobilizowana immobilizowana - po 60 dniach nie immobilizowana Czas reakcji [min] Rys. 13. Zależność stężenia maltozy od czasu reakcji hydrolizy skrobi z zastosowaniem α-amylazy z Bacillus licheniformis; - - enzym nie immobilizowany, biokatalizator z enzymem immobilizowanym: - - zastosowany bezpośrednio po immobilizacji, - - zastosowany po 60 dniach od immobilizacji [35]. Na podstawie przeprowadzonych reakcji hydrolizy stwierdzono odmienny wpływ danego nośnika na ten sam enzym, ale o różnym pochodzeniu. W pracy (B3) opisano przetwarzanie skrobi polegające na delikatnej hydrolizie jej granuli, a następnie poddaniu reakcji utleniania w celu otrzymania skrobi dialdehydowej. W warunkach naturalnych w granuli skrobi znajduje się niewielka ilość enzymów amylolitycznych. W wyniku naświetlania, aktywowano enzymy amylolityczne towarzyszące granuli skrobi, wywołując delikatne uszkodzenie granuli. Na tym etapie nie stosowano dodatkowych enzymów. Podobne efekty łagodnej degradacji skrobi w wyniku stymulacji światłem widzialnym opisano w literaturze [39-41]. Światło widzialne liniowo spolaryzowane uszkodziło granulkę skrobi, bez zmiany krystaliczności wnętrza granuli. Granule skrobi poddane wstępnej hydrolizie w wyniku aktywacji światłem, posiadały większą podatność na utlenianie (rys. 14). Naświetlone skrobie dialdehydowe hydrolizowały szybciej pod wpływem glukoamylazy. 27

 jest stwierdzenie, że stymulacja światłem widzialnym liniowo spolaryzowanym dotyczy nie tylko enzymów")

28 a) b) c) Rys. 14. Zdjęcia SEM granuli skrobi ziemniaczanej: strona lewa nie naświetlane, prawa naświetlane światłem widzialnym linio spolaryzowanym, a) nie utleniane, b) utleniane prze 30 minut, c) utleniane przez 60 minut (B3). Jednym z wniosków pracy (B3) jest stwierdzenie, że stymulacja światłem widzialnym liniowo spolaryzowanym dotyczy nie tylko enzymów wprowadzanych do reaktora podczas hydrolizy, ale także wpływa na enzymy naturalnie towarzyszące skrobi. Reakcje przetwarzania celulozy i produktów celulozowych Celulazy (EC ) z Aspergillus species naświetlano światłem widzialnym liniowo spolaryzowanym przez 30, 60, 120 i 180 minut (B1). Otrzymane katalizatory celulolityczne testowano w reakcjach hydrolizy celulozy mikrokrystalicznej i długołańcuchowej. Zarówno w 28
29 reakcji hydrolizy celulozy mikrokrystalicznej jak i długołańcuchowej zaobserwowano efekt aktywacji enzymu światłem w miarę wzrostu czasu naświetlania (rys.15 i rys. 16). Rys. 15. Zależność stopnia polimeryzacji od czasu hydrolizy celulozy mikrokrystalicznej z zastosowaniem celulazy: nie naświetlanej, naświetlanej przez: 30 minut, 60 minut, 120 minut, 180 minut (B1). 29
30 Rys. 16. Zależność stopnia polimeryzacji od czasu hydrolizy celulozy długołańcuchowej z zastosowaniem celulazy: nie naświetlanej, naświetlanej przez: 30 minut, 60 minut, 120 minut, 180 minut (B1). Wzrost aktywności katalitycznej enzymu korelował z wydłużaniem czasu naświetlania, ale tylko do 120 minut. Naświetlanie dłuższe niż 120 minut nie dawało dodatkowej stymulacji, co potwierdziły wyznaczone wartości stałych szybkości reakcji (B1). Stymulacja enzymu światłem wpłynęła także pozytywnie na obniżenie wartości LODP zarówno dla celulozy mikrokrystalicznej jak i długołańcuchowej. Wartość LODP (leveloff DP) określa stopień polimeryzacji trudno hydrolizowalnych (nie hydrolizowalnych) fragmentów celulozy [42]. Dla reakcji hydrolizy celulozy długołańcuchowej i mikrokrystalicznej, wyznaczono wartości stałych Michaelisa-Menten (K M ) z zastosowaniem biokatalizatorów stymulowanych światłem i nie naświetlanych enzymów. Naświetlanie od 30 do 120 minut spowodowało obniżenie wartości stałych Michaelisa-Menten (K M ), co świadczy o większym powinowactwie stymulowanych biokatalizatorów do obu substratów tj. celulozy długołańcuchowej i mikrokrystalicznej. W wyniku stymulacji celulazy z Aspergillus species światłem widzialnym liniowo spolaryzowanym uzyskano biokatalizatory o wyższej aktywności, większym powinowactwie 30
31 do celulozy mikrokrystalicznej i długołańcuchowej i wydajniej hydrolizujące oba te rodzaje celulozy (B1). Trzy komercyjne enzymy celulolityczne: Viscozyme L, Novozym 476 oraz celulazy z Aspergillus sp. immobilizowano na polimerowym nośniku otrzymanym w procesie rodnikowej kopolimeryzacji N-winyloformamidu (NVF) z dimetakrylanem glikolu etylenowego (EGDMA) w odwróconej suspensji (otrzymano w Zakładzie Polimerów, Wydział Chemii, Uniwersytet Jagielloński) (B4). Enzymy immobilizowano stosując aldehyd glutarowy (GA) jako łącznik między nośnikiem a celulazami. Otrzymane biokatalizatory nośnik-ga-enzym testowano w reakcjach hydrolizy papieru celulozowego P1 [43,44], a także ścieru drzewnego pochodzącego ze ściętych drzew iglastych (tzw. drewno miękkie). Biokatalizator uzyskany w wyniku immobilizacji koktajlu enzymatycznego Viskozyme L wykazywał dużo wyższą aktywność niż enzymy w stanie wolny. Z dużą szybkością hydrolizował zarówno papier celulozowy jak i ścier drewniany, był aktywny aż w trzech cyklach reakcji. W przypadku hydrolizy papieru we wszystkich trzech cyklach miał wyższą aktywność niż enzym nie immobiliowany, w przypadku drewna aktywność ta była wyższa w dwóch cyklach hydrolizy. Biokatalizator otrzymany w wyniku unieruchomienia na nośniku celulaz Novozym 476 posiadał wyższą aktywność katalityczną niż enzym w stanie wolnym w reakcji hydrolizy papieru celulozowego. Otrzymane biokatalizatory wykazywały większe powinowactwo do substratów, o czym świadczy niższa wartość stałej Michaelisa-Menten K M w porównaniu do wolnych enzymów. Celulaza z Aspergillus sp. obniżyła swoją aktywność w wyniku immobilizacji na nośniku, ale i tak była aktywna w trzech cyklach reakcji hydrolizy ścieru drzewnego i papieru celulozowego. W pracy (B4) zaprezentowano heterofazowe biokatalizatory w postaci celulaz immobilizowanych na nośniku polimerowym posiadającym silnie polarne grupy formamidowe. Biokatalizatory były aktywne w trzech cyklach hydrolizy ścieru drzewnego i papieru celulozowego. Część otrzymanych katalizatorów posiadała wyższą aktywność katalityczną niż enzymy w stanie wolnym. Biokatalizatory wykazywały odmienne powinowactwo do każdego z substratów, dlatego w celu uzyskania optymalnej wydajności reakcji hydrolizy biokatalizatory należy indywidualnie dobierać dla każdego z materiałów celulozowych. 31
32 Na nośniku poli(n-winyloformamidowym) hydrolizowanym w środowisku kwaśnym unieruchomiono celulazę z Aspergillus sp. W jednym przypadku zastosowano aldehyd glutarowy (GA) jako łącznik, w drugim enzym unieruchomiono bezpośrednio na nośniku. Biokatalizator typu nośnik-ga-enzym testowano w rekcji hydrolizy celulozy długołańcuchowej (rys. 17). Stezenie cukrow redukujacych [mg/ml] cellulaza z Aspergillus sp. nie immobilizowana immobilizowana z GA immobilizowana bez GA Czas reakcji [min] Rys. 17. Zależność stężenia cukrów redukujących od czasu reakcji hydrolizy celulozy długołańcuchowej z zastosowaniem celulazy z Aspergillus sp.; - - enzym nie immobilizowany, biokatalizator z enzymem immobilizowanym: - - z GA jako łącznikiem, - - bez łącznika. Oba biokatalizatory posiadały wyższą aktywność katalityczną niż enzym w stanie wolnym, jednak enzym unieruchomiony bezpośrednio na nośniku był najbardziej aktywny, co potwierdzają wysokie wartości stałych szybkości reakcji oraz wydajność reakcji hydrolizy (Tabela 2). 32
33 Tabela 2. Stałe szybkości i wydajność reakcji hydrolizy celulozy długołańcuchowej z zastosowaniem celulazy z Aspergillus sp. Celulaza z Aspergillus sp. Stała szybkości [mg ml -1 min -1 ] k 1 x10-3 k 2 x10-4 Wydajność hydrolizy po 24 h [mg/ml] Nie immobilizowana 2,9 11,8 0,46 Immobilizowana z GA 7,9 8,7 0,56 Immobilizowana bez GA 11,7 3,4 0,75 Natomiast w przypadku biokatalizatora nośnik-ga-enzym zaobserwowano wzrost stabilności na zmiany temperatury i ph reakcji w porównaniu do biokatalizatora nośnik-enzym oraz enzymu w stanie wolnym (rys. 18) i (rys.19). Stezenie cukrow redukujacych [mg/ml] cellulaza z Aspergillus sp. nie immobilizowana immobilizowana z GA immobilizowana bez GA Temperatura [ o C] Rys. 18. Zależność stężenia cukrów redukujących od temperatury w reakcji hydrolizy celulozy długołańcuchowej z zastosowaniem celulazy z Aspergillus sp.: - - enzym nie immobilizowany, biokatalizator z enzymem immobilizowanym: - - z GA jako łącznikiem, - - bez łącznika. 33
34 0.9 Stezenie cukrow redukujacych [mg/ml] cellulaza z Aspergillus sp. nie immobilizowana immobilizowana z GA immobilizowana bez GA ph Rys. 19. Zależność stężenia cukrów redukujących od ph w reakcji hydrolizy celulozy długołańcuchowej z zastosowaniem celulazy z Aspergillus sp.: - - enzym nie immobilizowany, biokatalizator z enzymem immobilizowanym: - - z GA jako łącznikiem, - - bez łącznika. Immobilizacja celulaz na nośniku z zastosowaniem łącznika GA, powoduje najprawdopodobniej wielopunktowe związanie enzymu z nośnikiem, co skutkuje obniżeniem aktywności katalitycznej [45,46]. W immobilizacji wielopunktowej tworzą się ograniczenia sferyczne wokół cząsteczek enzymów dzięki czemu są one usztywnione i bardziej odporne, co objawia się podwyższoną stabilnością enzymu na zmiany temperatury i ph [47]. Biokatalizator nośnik-enzym Aspergillus sp. bez łącznika GA zastosowano także w reakcji hydrolizy stosunkowo odpornej celulozy mikrokrystalicznej (rys. 20). 34
35 Stezenie cukrow redukujacych [mg/ml] celulaza z Aspergillus sp. nie immobilizowana immobilizowana - cykl I immobilizowana - cykl II immobilizowana - cykl III Czas reakcji [min] Rys. 20. Zależność stężenia cukrów redukujących od czasu reakcji hydrolizy celulozy mikrokrystalicznej z zastosowaniem celulazy z Aspergillus sp.; - - enzym nie immobilizowany, biokatalizator z enzymem immobilizowanym: - - I cykl reakcji, - - II cykl reakcji, - - III cykl reakcji. Biokatalizator posiadał bardzo wysoką aktywność katalityczną, w dwóch cyklach reakcji wyższą niż enzym w stanie wolnym (Tabela 3). Tabela 3. Stałe szybkości i wydajność reakcji hydrolizy celulozy mikrokrystalicznej z zastosowaniem celulazy z Aspergillus sp. Celulaza z Aspergillus sp. Stała szybkości [mg ml -1 min -1 ] k 1 x10-3 k 2 x10-4 Wydajność hydrolizy po 24 h [mg/ml] Nie immobilizowana 4,2 13,0 0,48 Immobilizowana - cykl I 24,3 10,1 0,83 Immobilizowana - cykl II 18,3 3,1 0,74 Immobilizowana cykl III 3,2 7,4 0,41 35
36 Biokatalizatory w postaci celulazy Novozym 476 unieruchomionej na trzech nośnikach opisano w pracy (B2). Do immobilizacji enzymu wybrano trzy nośniki o różnym stopniu usieciowania (otrzymane w Zakładzie Polimerów, Wydział Chemii, Uniwersytet Jagielloński). Celulazy unieruchamiano na nośnikach poprzez łącznik aldehyd glutarowy. Otrzymano stabilne biokatalizatory posiadające kowalencyjne wiązania imidowe nośnikłącznik-enzym. Wszystkie biokatalizatory wykazywały aktywność katalityczną w reakcji hydrolizy celulozy mikrokrystalicznej. Stwierdzono że, wyższą aktywność niż enzym nie immobilizowany miał biokatalizator otrzymany poprzez unieruchomienie celulazy na nośniku o wysokim stopniu usieciowania i sferycznym kształcie ziaren. Reakcje przetwarzania ksylanu Hydrolizę ksylanu przeprowadzono z zastosowaniem katalizatora otrzymanego w wyniku stymulacji światłem widzialnym liniowo spolaryzowanym endo-1,4-β ksylanazy (EC ) z Themomyces lanuginosus otrzymanej na drodze fermentacji genetycznie modyfikowanych mikroorganizmów Aspergillus oryzae (B6 i B9). Enzym stymulowano światłem widzialnym liniowo spolaryzowanym oraz światłem nie spolaryzowanym (o podobnym strumieniu energii) przez 60 i 120 minut. W celu porównania rezultatów, wykonano także reakcje hydrolizy z naświetlanym wyłącznie substratem (ksylan) oraz z naświetlaną mieszaniną reakcyjną (ksylan + ksylanaza w buforze fosforanowym). Rezultaty hydrolizy ksylanu przedstawiono na rys. 21 (B9). 36
37 Ksyloza [mg/mg xylan] Czas reakcji [min] Rys. 21. Zależność stężenia ksylozy od czasu reakcji hydrolizy ksylanu z zastosowaniem ksylanazy z Themomyces lanuginosus.; - - enzym nie naświetlany, - - enzym naświetlany 120 min., - - naświetlany substrat, - - naświetlana mieszanina reakcyjna do 300 min. Naświetlanie mieszaniny reakcyjnej podczas hydrolizy nie przyniosło żadnych zmian w przebiegu reakcji. Najwolniej przebiega hydroliza naświetlanego substratu (ksylanu) z nie naświetlanym enzymem (ksylanazą). Biokatalizator o najwyższej aktywności katalitycznej uzyskano w wyniku naświetlania enzymu przez 120 minut. Wykazano także, że stymulujący wpływ na ksylanazę miało naświetlanie światłem nie spolaryzowanym. Wyższą aktywność katalityczną obserwuje się jednak w przypadku enzymu naświetlanego światłem widzialnym liniowo spolaryzowanym rys. 22 (B9). 37
38 0.8 wzorzec 1godz. bez filtra polaryzacyjnego 1godz. 2godz. 0.7 Stężnie ksylozy [mg/ml] Czas reakcji enzymatycznej [min] Rys. 22. Zależność stężenia ksylozy od czasu reakcji enzymatycznej hydrolizy ksylanazy z Themomyces lanuginosus: nie naświetlanej, - naświetlanej światłem nie spolaryzowanym przez 1 godz., - naświetlanym światłem widzialnym liniowo spolaryzowanym przez 1 godz., - naświetlanym światłem widzialnym liniowo spolaryzowanym przez 2 godz. Reakcje przetwarzania chityny i chitozanu Hydrolizę chityny i chitozanu z zastosowaniem enzymów stymulowanych światłem opisano w pracy (B8). Zaobserwowano stymulący wpływ światła spolaryzowanego na wzrost aktywności katalitycznej chitynazy rys. 23 i chitozanazy rys
39 0.6 N-Acetyl-D-glucosamine [mg/ml] Czas reakcji [min] Rys. 23. Zależność stężenia N-acetylo-D glukozaminy od czasu reakcji hydrolizy z zastosowaniem chitynazy: - - nie naświetlanej, - - naświetlanej światłem nie spolaryzowanym, oraz naświetlanej światłem spolaryzowanym przez: min., min., - - naświetlany substrat przez 60 min. (B8). 0.6 Glukozamina [mg/ml] Czas reakcji [min] Rys. 24. Zależność stężenia glukozaminy od czasu reakcji hydrolizy z zastosowaniem chitozanazy: - - nie naświetlanej, naświetlanej światłem spolaryzowanym przez: min., min., min. (B8). 39
40 Efekt stymulacji światłem widzialnym liniowo spolaryzowanym korelował ze wzrostem czasu naświetlania. Nie stwierdzono efektu aktywującego w przypadku naświetlania światłem nie spolaryzowanym oraz naświetlania światłem spolaryzowanym mieszaniny reakcyjnej. Podjęto próbę aktywacji chitozanazy i chitynazy w wyniku immobilizacji na nośniku polimeru hydrożelowego [35]. Uzyskano biokatalizator aktywny w reakcji hydrolizy chityny w dwóch cyklach reakcji, ale o niższej aktywności katalitycznej niż enzym w stanie wolnym (rys. 25). N-acetylo-glucosamina [mg/ml] chitynaza nie immobilizowana immobilizowana - cykl I immobilizowana - cykl Il Czas reakcji [min] Rys. 25. Zależność stężenia N-acetylo-D glukozaminy od czasu reakcji hydrolizy z zastosowaniem chitynazy: - - nie immobilizowanej, oraz biokatalizatora z immobilizowanym enzymem: - - I cykl reakcji, - - II cykl reakcji [35]. Chitozanaza immobilizowana na nośniku posiadała bardzo niską aktywność katalityczną (rys. 26). 40
41 Glucosamina [mg/ml] chitozanaza immobilizowana nie immobilizowana Czas reakcji [min] Rys. 26. Zależność stężenia glukozaminy od czasu reakcji hydrolizy z zastosowaniem chitozanazy: - - nie immobilizowanej oraz - - biokatalizatora z enzymem immobilizowanym [35]. Przeprowadzone badania spektroskopowe (RAMAN i UV-Vis) wykazały, że reaktywne grupy formamidowe na powierzchni nośników nie zapewniają trwałego wiązania enzym-ganośnik, z tego powodu w przeprowadzonych testach katalitycznych badane biokatalizatory charakteryzowały się mniejszą aktywnością niż enzym w formie natywnej. Reakcje przemiany mieszaniny polisacharydów W patencie (B10) głównym celem było opracowanie metody badania szybkości degradacji polisacharydów w produktach leczniczych, zwłaszcza w syropach ziołoleczniczych, przeznaczonych do dłuższego przechowywania. Szybkość degradacji polisacharydów jest miarą rozpadu produktu leczniczego. Ustalenie czasu, po którym nastąpi krytyczny stopień rozkładu polisacharydu w produkcie leczniczym pozwala na określenie, po jakim czasie produkt leczniczy traci swoje właściwości lecznicze. Umożliwia to tym samym wyznaczenie okresu przydatności do spożycia lub okresu ważności produktów leczniczych. Z tego powodu konieczne było przeprowadzenie badań i określenie jakim reakcjom ulegają polisacharydy w produktach. Badaniom poddano syrop prawoślazowy wyprodukowany z Zakładzie Farmaceutycznym AMARA w Krakowie. Badano między innymi degradację polisacharydów na drodze kwaśnej hydrolizy, utleniania a także hydrolizy enzymatycznej, określono wpływ promieniowania UV i światła liniowo spolaryzowanego na stopień 41
42 polimeryzacji polisacharydów w syropie. Część z powyższych rezultatów została zaprezentowana w pracy magisterskiej Edyty Pięty pt. Opracowanie alternatywnych metod badania preparatów farmaceutycznych w procesie produkcyjnym, której habilitantka była promotorem [51]. Podczas badań enzymatycznej hydrolizy syropu prawoślazowego z zastosowaniem mieszaniny enzymów: glukoamylazy i ksylanazy wykazano wpływ naświetlania światłem widzialnym liniowo spolaryzowanym na przebieg degradacji polisacharydów w syropie. Hydroliza polisacharydów z zastosowaniem enzymów aktywowanych światłem przebiegała z większą szybkością, co potwierdzają wyznaczone stałe szybkości reakcji (rys. 27). 1.0 Stezenie cukrow redukujacych [mg/ml] k 1 = [mg ml -1 min -1 ] k 1 = [mg ml -1 min -1 ] k 2 = [mg ml -1 min -1 ] k 2 = [mg ml -1 min -1 ] Czas reakcji enzymatycznej [min] Rys. 27. Enzymatyczna degradacja polisacharydów w syropie prawoślazowy z zastosowaniem mieszaniny enzymów: glukoamylaza + ksylanaza: - - nie stymulowanej światłem, - - naświetlanej światłem widzialnym liniowo spolaryzowanym. Wnioski wyciągnięte na podstawie degradacji polisacharydów w syropie prawoślazowym i potwierdzenie wpływu światła widzialnego liniowo spolaryzowanego na degradację syropu były powodem, dla którego habilitantka zajęła się tą problematyką badań. 42
43 Wnioski Otrzymano biokatalizatory homo- i heterofazowe w postaci enzymów aktywowanych fizycznie poprzez naświetlanie światłem widzialnym liniowo spolaryzowanym oraz chemicznie w wyniku immobilizacji na nośnikach polimerowych. Uzyskane biokatalizatory efektywnie hydrolizują surowce polisacharydowe: skrobię, celulozę, ksylan, chitynę, chitozan. Fizykochemiczna aktywacja spowodowała, że otrzymane biokatalizatory mają wyższą aktywność, stabilność temperaturową i ph, odmienną selektywność w porównaniu do enzymu natywnego. Wykazano korzystną stymulację enzymów poprzez naświetlanie światłem liniowo spolaryzowanym o długości fali z zakresu światła widzialnego. Naświetlanie światłem liniowo spolaryzowanym indukuje zmiany konformacji enzymów, co wpływa na zmianę ich aktywności katalitycznej, selektywności, a w przypadku niektórych enzymów zmieniało stabilność na zmiany temperatury i ph. Stymulacja enzymów bezpośrednio przed wprowadzeniem do reakcji zapewniała największe korzyści. Jednak zmiany konformacji wywołanie naświetlaniem były na tyle trwałe, że niektóre ze stymulowanych enzymów można było przechowywać przez okres 1-3 miesięcy bez utraty ich właściwość. Należy zaznaczyć że, reakcję enzymatyczną z naświetlanymi biokatalizatorami wykonywano w sposób standardowy i ze standardowych substratów. Stwierdzono, że optymalna ilość energii dostarczonej do enzymu w wyniku naświetlenia (regulowana natężeniem i czasem naświetlania) była stała dla danego enzymu. Badania z zakresu immobilizacji enzymów na nośnikach wykazały, że ten sam nośnik na jeden enzym działał synergizująco, jeśli chodzi o aktywność katalityczną, na inny wywierał wpływ wręcz dezaktywujący, a jeszcze w innych przypadkach pozostawał mniej lub bardziej obojętny. Podobne zmiany zaobserwowano w przypadku stabilności temperaturowej i ph immobilizowanych enzymów. Z przeprowadzonych badań jak i ze światowej literatury [48,49] wynika, że brak jest ogólnych wytycznych, które pozwalałyby usystematyzować dobór najefektywniejszego nośnika dla określonego enzymu, sposobu immobilizacji, zastosowania łączników itp. dominuje zasada prób i błędów. Stwierdzono, że właściwości nośników w odniesieniu do immobilizacji enzymów determinowane są takimi cechami jak porowatość matrycy, wielkość i kształt porów, hydrofilowość czy hydrofobowość szkieletu, polarność spowodowana obecnością różnych grup funkcyjnych. Udowodniono, że polimery oparte na poli(n-winyloformamidzie) mogą być z sukcesem stosowane jako nośniki do immobilizacji wielu enzymów. 43
44 Unieruchomienie enzymów na nośnikach przeprowadzono wieloma sposobami, najkorzystniejsze jednak okazało się związanie enzymów z podłożem wiązaniem kowalencyjnym. Zastosowanie aldehydu glutarowego jako łącznika determinowało jednopunktowy sposób wiązania i było dla niektórych enzymów korzystne, a dla innych nie. Wykazano, że większość nośników z grupami aminowymi wymaga aktywacji aldehydem glutarowym, zanim zostaną użyte do immobilizacji z wytworzeniem wiązania kowalencyjnego enzym-nośnik. Jednak w przypadku zastosowania nośników hydrożelowych enzymy były otaczane i zamykane przez łańcuchy polimerowe. Na skutek wielopunktowych, niekowalencyjnych odziaływań cząsteczek białek z grupami nośnika, dochodziło do ich unieruchomienia dzięki stosunkowo słabym oddziaływaniom elektrostatycznym i hydrofobowym oraz wiązaniom wodorowym. W cyklu prac wykazano, że wiele parametrów decyduje o efekcie końcowym immobilizacji, czyli o ilości, aktywności, trwałości wiązania enzym-nośnik i właściwościach enzymatycznych związanego białka (aktywności katalitycznej, selektywności, stabilności temperaturowej itp.). Głównymi czynnikami, które opisano są: skład nośnika, powierzchnia dostępna dla enzymu, ilość zaktywowanych grup funkcyjnych nośnika, rodzaj czynnika wiążącego enzym z nośnikiem, dostępność grup funkcyjnych białka w danym zakresie ph, odległość związanego enzymu od powierzchni nośnika oraz orientacja przestrzenna centrum aktywnego, mono- lub wielopunktowe związanie enzymu z nośnikiem, powinowactwo chemiczne enzymu do materiału nośnika. Różne właściwości i zachowanie enzymów po immobilizacji na nośniku, wynikają z ich struktury, konformacji, stopnia polimeryzacji i rodzaju i sekwencji jednostek proteinowych. W dalszym cyklu prac badawczych należałoby dokładniej wyjaśnić odmienny wpływ danego nośnika na ten sam enzym, ale o różnym pochodzeniu. Zaobserwowano również odmienną podatność otrzymanych biokatalizatorów w stosunku do hydrolizowanych substratów. Dlatego wymiernym efektem byłoby w przyszłości opracowanie bazy biokatalizatorów o pożądanych właściwościach, celem wykorzystania w dalszych badaniach inwestycyjnych przez potencjalnych partnerów z przemysłu. W zależności od wymagań procesowych (wzrost aktywności, zmiany selektywności, stabilności temperaturowej i ph) należy dla każdej reakcji, substratu, enzymu indywidualnie na drodze eksperymentu dobierać odpowiedni sposób aktywacji fizykochemicznej. Takie postępowanie gwarantuje uzyskanie biokatalizatora o pożądanych właściwościach. 44
45 Przeprowadzone prace badawcze wskazują, iż zaproponowane sposoby aktywacji fizykochemicznej są stosunkowo proste i niskokosztowe, stanowią więc obiecującą metodę otrzymywania biokatalizatorów dla potrzeb procesowych i mogą znaleźć zastosowanie w wielu dziedzinach przemysłu. Dodatkowo opisane biokatalizatory otrzymywane są z powszechnie dostępnych i stosowanych w przemyśle enzymów, więc ich zastosowanie nie wymaga konieczności zmiany technologii, infrastruktury przemysłowej czy wprowadzania nowych procesów produkcyjnych. W przemyśle spożywczym wykorzystanie nowych biokatalizatorów może przyczynić się do zwiększenia funkcjonalności artykułów spożywczych, poprawy wartości odżywczych, własności sensorycznych. Pozwoli także na obniżenie kosztów i ulepszenie procesów przetwarzania żywności oraz utylizacji odpadów. Otrzymane biokatalizatory mogą przynosić korzyści w zakresie ochrony środowiska, ze względu na biodegradowalność i obniżenie zużycia energii, wpisują się więc w założenia zasad Zielonej Chemii. Literatura [1] A.D. Smith, S.P. Datta, G.H. Smith, Campbell P.N., Bentley R., McKenzie H.A. (eds) Oxford Dictionary of Biochemistry and Molecular Biology. Oxford University Press, Oxford, [2] L. Michaelis, M. Menten, Biochemische Zeitschrift 49, [3] R. Boyer, Concepts in Biochemistry, 2nd edn (wyd. ang.) John Wiley & Sons, Inc., New York, [4] R.M. Berka, J.R. Cherry, Enzyme biotechnology. In: Ratledge C, Kristiansen B, editors. Basic Biotechnology. 3rd edition. Cambridge, UK: Cambridge University Press, [5] J. Norus, (2006) European Planning Studies, 14 (5), [6] P.B. Poulsen, H. Klaus Buchholz, History of enzymology with emphasis on food production. In: Whitaker JR, Voragen AGJ, Wong DWS, Handbook of Food Enzymology, New York, NY, USA: Marcel Dekker; [7] P. Fernandes, Enzymes in sugar industries. In: Panesar P, Marwaha SS, Chopra HK, editors, Enzymes in Food Processing: Fundamentals and Potential Applications, New Delhi, India: I.K. International Publishing House; [8] D.E. Robertson, B.A. Steer, (2004) Curr. Opin. Chem. Biol. 8, [9] L. Betancor, A. Hidalgo, G. Fernández-Lorente, C. Mateo, R. Fernández-Lafuente, J.M. Guisán (2003) Biotechnol. Prog. 19, [10] BRENDA Enzyme Information Database, Brenda-enzymes.info/ [11] [12] Celuloza (CID: ) (ang.) w bazie PubChem, United States National Library of Medicine, [13] P.W. Goodenough, (1995) Molecular Biotechnology, 4,
46 [14] E. Katchalski-Katzir, (1993) TIB, 11, [15] B. Wang, F. Cheng, Y. Lu, W. Ge, M. Zhang, B. Yue, (2013) J. Mol. Catal. B. Enzym. 97, [16] A. Sassolas, L.J. Blum, B.D. Leca-Bouvier, (2012) Biotech. Adv. 30, [17] J.M. Guisan, Methods in biotechnology: immobilization of enzymes and cells, 2nd ed., Humana Press, Totowa, NJ, [18] S.A. Ansari, Q. Husain, (2011) Biotech. Adv. 30, [19] J. Kim, J.W. Grate, P. Wang, (2008) Trends Biotechnol. 26 (11) [20] R.A. Sheldon, (2007) Adv. Synth. Catal. 349, [21] C. Walsh, Enzymatic Reaction Mechanism, Freeman, San Francisco, 1979 [22] L. Cao, L. van Langeny, R.A. Sheldon, (2003) Current Opinion in Biotechnology, 14, [23] A.R. Fersht, Structure and Mechanism in Protein Science, Freeman, New York, [24] M. Fiedorowicz, A. Konieczna Molenda, G. Khachatryan, (2007) Starch: Progress in structural studies, modifications and applications. Eds: P.Tomasik, V.Yuryev, E.Bertoft, Polish Society of Food Technologist Małopolska Branch, ISBN [25] T. Kubasowa, M. Fenyo, Z. Somosy, L. Gazso, I. Kertesz, (1988) Photochemistry and Photobiology, 48, 4, [26] Biologic Effects of Light; Eds. M. F. Holik, E. G. Jung, Proceedings of a Symposium Basel, Switzerland, [27] Patent DE (Int.Cl. 5 A61N5/06; A61F7/00 M. Munz; Heat and light therapy procedure - radiating selection of colored polarized light for treatment of skin ailments and bodily functions. [28] H. Hoeksema, S. Monstrey, K. Van Landuyt. Ph. Blondeel, P. Tonnard, A. Verpaele, The use of polarised light in the treatment of severely burned patients. 10th Congress of the International Society for Burn Injuries, Jerusalem, Israel, [29] K. Depuydt, S. Monstrey, H. Hoeksma, The stimulating effects of polarized light on wound healing and Avoiding surgery in the treatment of deep dermal burn wounds using polarized light. 10 th Annual Meeting of the European Association of Plastic Surgeons (EURAPS), Madrid, [30] E. Bazso, Sz. Varju, P. Szego, K. Roza, P. Apai, Application of Incoherent Wide Band Polarised Light To Promote Healing of Wounds; Central Research Institute for Physics: Budapest, Hungary, [31] W. Vanscheidt, (2002) Eur. J. Plast. Surg., 24, [32] A.E. Nave, B.B. Rubenstein, (1928) J. Biol. Chem., 89, [33] O.H. Lowry, N.J. Rosebrough, A.L. Farr, R.J. Randall, (1951) J. Biol. Chem., 193, [34] K. Yamamoto, Y. Imamura, E. Nagatomo, T. Sevizawa, Y. Muraoko, M. Akashi, (2003) J. Appl. Polym. Sci., 89, [35] A. Konieczna-Molenda, A. Kochanowski, A. Grabiec, E. Bortel, P. Tomasik; (2008) Starch: Recent Progress in Biopolymer and Enzyme Technology, (Eds.: P. Tomasik, E. Bertoft, A. Blennow), PTTZ-Malopolska Branch, Krakow, 8, [36] S. Pedersen, L. Dijkhuizen, B.W. Dijkstra, B.F. Jensen, S.T. Jřrgensen; (1995) Chemtech., 25,
47 [37] J. Szejtli, (1990) Carbohydr Polym., 12, [38] A. Lejeune, K. Sakaguchi, K. Imanaka, (1989) Anal. Biochem. 181, [39] M. Fiedorowicz, P. Tomasik, P. C.Y. Lii, (2001) Carbohydr. Polym., 45, [40] M. Fiedorowicz, C.Y. Lii, P. Tomasik, (2002) Carbohydr. Polym., 50, [41] M. Fiedorowicz, G. Khachatryan, (2003) J. Sci. Food Agric., 84, [42] O.A. Battista, S. Coppiek, J.A. Hoesmon, F.F. Morehead, W.A. Sisson, (1956) Ind. Eng. Chem., 48, [43] A. Barański, A. Konieczna-Molenda, J.M. Łagan, L.M. Proniewicz; (2003) Restaurator, 24, [44] L.M. Proniewicz, C. Paluszkiewicz, A. Wesełucha-Birczyńska A. i in.: (2001) Journal of Molecular Structure, 596, [45] C. Mateo, J.M. Palomo, G. Fernandez-Lorente, J.M. Guisan, R. Fernandez-Lafuente, (2007) Enzyme Microb. Technol., 40, [46] F. Lopez-Gallego, L. Betancor, A. Hidalgo, N. Alonso, G. Fernandez-Lorente, J.M. Guisan, R. Fernandez-Lafuente, (2005) Enzyme Microb. Technol., 37, [47] F. Lopez-Gallego, L. Betancor, C. Mateo, A. Hidalgo, N. Alonso-Morales, G. Dellamora-Ortiz, J.M. Guisian, R. Fernıandez- Lafuente, (2005) J. Biotechnol., 119, [48] J. Bryjak; (2003) Biochemical Engineering Journal, 16, [49] I. Gancarz, J. Bryjak, M. Bryjak, G. Pozniak, W. Tylus, (2003) European Polymer Journal, 39, [50] A. Konieczna-Molenda, A. Kochanowski, A. Walaszek, E. Bortel and P. Tomasik; (2009) Chemical Engineering Journal, 146, 3(15) [51] Edyta Pięta (2010) Praca magisterska pt. Opracowanie alternatywnych metod badania preparatów farmaceutycznych w procesie produkcyjnym Wydział Chemii, Uniwersytet Jagielloński. 47
48 4. OMÓWIENIE POZOSTAŁYCH OSIĄGNIĘĆ NAUKOWO - BADAWCZYCH Mój dotychczasowy dorobek naukowy obejmuje łącznie 29 prac, w tym 15 publikacji w czasopismach indeksowanych w bazie JCR (łączny IF wg roku opublikowania wynosi 21,875, a ilość punktów według MNiSW wynosi 549), 10 rozdziałów w monografiach (w tym 5 w materiałach konferencyjnych indeksowanych w bazie Web of Science) oraz 2 pełnotekstowe prace w innych recenzowanych czasopismach. Jestem autorem 2 patentów (w tym jednego wdrożonego w przemyśle) oraz współautorką 58 doniesień opublikowanych w materiałach konferencyjnych. Publikacje, których jestem współautorem były cytowane 205 razy (198 - bez autocytowań) według bazy Web of Science, współczynnik h (Hirsch index) w wymienionej bazie wynosi odpowiednio 6. Szczegółowa analiza bibliometryczna oraz wykaz wszystkich prac i patentów zostały przedstawione w Załączniku 6. Główny nurt prowadzonych przeze mnie badań dotyczy reakcji przemian, hydrolizy polimerów naturalnych głównie polisacharydów. W roku 1998 w ramach mojej pracy doktorskiej rozpoczęłam badania dotyczące degradacji celulozy. Badania realizowane w Zespole Katalizy Heterogenicznej oraz w Zakładzie Technologii Chemicznej pod kierunkiem Prof. dr. hab. Andrzeja Barańskiego dotyczyły problematyki degradacji papieru. Opracowałam wówczas metodykę testów przyspieszonego starzenia celulozy w warunkach tlenowych i beztlenowych, w szerokim zakresie temperatur i wilgotności względnych. 1,2 Określiłam optymalne warunki przechowywania (temperatury, wilgotności, oświetlenia itp.) książek i dokumentów w bibliotekach i archiwach. 3,4 Opracowałam kinetyczny model degradacji celulozy w zakresie temperatur o C. 5,6 1 A. Barański, R. Dziembaj, A. Konieczna, A. Kowalski, J.M. Łagan, L. Proniewicz; (2000) Methodology of Kinetic Investigation of Cellulose Degradation; Methodology of kinetic investigation of cellulose degradation, Technologia Chemiczna na Przełomie Wieków, Wydawnictwo Stałego Komitetu Kongresów Technologii Chemicznej, Gliwice, F. Krok, P. Piątkowski, P. Czuba, A. Konieczna, A. Barański, M. Szymoński; (2000) Atomic Force Microscopy Imaging of the Paper Surface Subjected to Process of Artificial Ageing; Electron Technology 33(3) A. Barański, A. Konieczna Molenda, J.M. Łagan, L.M. Proniewicz; (2003) Catastrophic room temperature degradation of cellulose; Restaurator A. Barański, R. Dziembaj, A. Konieczna-Molenda, J.M. Łagan, S. Walas; (2003) Wpływ siarczanu glinu na kwaśną degradację celulozy; Przemysł Chemiczny 82/ L.M. Proniewicz, C. Paluszkiewicz, A. Wesełucha-Birczyńska, H. Majcherczyk, A. Barański, A Konieczna; (2001) FT-IR and FT-Raman study of hydrothermally degraded cellulose; Journal of Molecular Structure A. Barański, R. Dziembaj, A. Konieczna Molenda, J.M. Łagan, S. Walas; (2004) On the Applicability of Arrhenius Equation to Accelerated Ageing Tests. The Case of Alum-impregnated Cellulose; Polish Journal of Chemical Technology 6,
49 Określiłam wpływ zakwaszenia papieru na jego trwałość podczas przechowywania. 7 Wykonałam ocenę i opis dostępnych w świecie technologii odkwaszania papieru. Po doktoracie kontynuowałam badania dotyczące problematyki degradacji polisacharydów w różnych warunkach temperatury, ph, obecności związków utleniających, naświetlania itp. Opracowałam metodę pozwalającej określić degradację polisacharydów w produktach leczniczych i spożywczych. Sposób ten został opatentowany oraz wdrożony w Zakładzie Farmaceutycznym AMARA w Krakowie 8,9. Metoda, została zatwierdzona przez Urząd Rejestracji Leków, wykorzystywana jest obecnie w przemyśle farmaceutycznym w celu określania terminu przydatności do spożycia syropu prawoślazowego. W 2005 roku rozpoczęłam pracę w Katedrze Chemii Uniwersytetu Rolniczego w Krakowie pod kierownictwem Prof. dr. hab. Piotra Tomasika. Na początku mojej pracy zajmowałam się modyfikacjami i hydrolizą polisacharydów, w szczególności skrobi oraz badałam wpływu światła liniowo spolaryzowanego na właściwości polisacharydów. Byłam współautorką prac wyjaśniających wpływ światła widzialnego liniowo spolaryzowanego na strukturę krystaliczną ziaren skrobi o różnym pochodzeniu biologicznym. Określiłam wpływ tego promieniowania na podatność na hydrolizę, właściwości termiczne, masę cząsteczkową, lepkość, a więc właściwości funkcjonalne skrobi posiadające znaczenie w jej przetwórstwie przemysłowym. 10,11 Wykazałam większą podatność polisacharydów naświetlanych światłem widzialnym liniowo spolaryzowanym na hydrolizę enzymatyczną. 12,13 7 A. Barański, D. Dutka, R. Dziembaj, A. Konieczna-Molenda, J.M. Łagan; (2004) Effect of Relative Humidity on the Degradation Rate of Cellulose. Methodology Studies; Restaurator A. Konieczna-Molenda; (2011) Sposób określenia szybkości degradacji polisacharydów w produktach leczniczych, zwłaszcza w syropach ziołoleczniczych, polski patent PL L. Krzemińska-Fiedorowicz, K. Khachatryan, A. Konieczna-Molenda (2018) Evaluation of Polysaccharides Durability in Syrup of Marsh Mallow (2018) Procceedings of the 14th International Conference on Polysaccharides-Glycoscience. Prague, Czech Republic, Prague, Czech Republic, Fiedorowicz M., Khachatryan G., Konieczna - Molenda A., Yuryev V. P., Wasserman L. A. (2006) Illumination of sago starch with linearly polarized visible light, In: Starch: Recent Achievements in Understanding of Structure and Functionality, (Eds. Yuryev V., Tomasik P., Bertoft E.) Nova Science Publishers, NY, ISBN: M. Fiedorowicz, A. Konieczna Molenda, G. Khachatryan, H. Staroszczyk, P. Tomasik, (2007), Polarized light as a powerful tool in physical and enzymatic modifications of carbohydrates, In: Consumer Protection through Food Process Improvement & Innovation in the Real World, Hellenic Association of Food Technologists, North Greece Branch, Thessaloniki GREECE ISBN M. Fiedorowicz, A. Konieczna-Molenda, G. Khachatryan, (2008) The Effect of Illumination of Polarized Light on Polysaccharides and Polysaccharide Degrading Enzymes, Polysacharidy (2008) Chem. Listy 102, A. Konieczna-Molenda, M. Fiedorowicz and P. Tomasik; (2008) The Polarized Light-induced Enzymatic Formation and Degradation of Biopolymers; Macromol. Symp. 272,
50 W kolejnej pracy przedstawiłam, że naświetlanie światłem widzialnym liniowo spolaryzowanym ziaren skrobi uszkadza je, przez co staje się ona bardziej podatna na utlenianie i hydrolizę. 14 Wykazałam, także wpływ naświetlania światłem widzialnym liniowo spolaryzowanym na właściwości i strukturę celulozy. 15 Scharakteryzowałam naświetlaną celulozę pod względem zmian struktury, krystaliczności, stopnia polimeryzacji, zawartości wody związanej, podatności na hydrolizę kwaśną i enzymatyczną. Jednym z głównych kierunków moich badań było określenie wpływu światła widzialnego liniowo spolaryzowanego na właściwości enzymów takie jak: aktywność katalityczna, stabilność temperaturowa i ph oraz selektywność. Enzymy powszechnie stosowane w przemyśle symulowałam światłem i otrzymałam biokatalizatory o odmiennych właściwościach niż enzymy natywne. Otrzymane biokatalizatory posiadały wysoką aktywność w reakcji hydrolizy skrobi 16,17, ksylanu 18, celulozy 19, chityny i chitozanu 20 oraz utleniania glukozy 21. Transferaza cyklodekstrynowa pod wpływem naświetlania światłem widzialnym liniowo spolaryzowanym zmieniła selektywność w stosunku do otrzymywanych produktów. 22 Sposób stymulacji enzymów światłem widzialnym liniowo spolaryzowanym został opatentowany A. Para and Anna Konieczna Molenda; (2010) Starch dialdehyde from potato starch illuminated with linearly polarized white light; Carbohydrate Polymers, 79 (2), A. Konieczna-Molenda A., M. Molenda, M. Fiedorowicz, P. Tomasik; (2008) Illumination of cellulose with linearly polarized visible light; Macromol. Symp. 272, M. Fiedorowicz, A. Konieczna-Molenda, G. Khachatryan, (2008) The Effect of Illumination of Polarized Light on Polysaccharides and Polysaccharide Degrading Enzymes, Polysacharidy (2008) Chem. Listy 102, M. Fiedorowicz, A. Konieczna Molenda and G. Khachatryan; (2007) Light stimulated enzymatic processes on selected polysaccharides; Starch: Progress in structural studies, modifications and applications. Polish Society of Food Technologist Małopolska Branch, ISBN , A. Konieczna- Molenda, V. M. F. Lai, M. Fiedorowicz, G. Khachatryan, P. Tomasik; (2008) Polarized Lightstimulated Enzymatic Hydrolysis of Xylan Biotechnology Progress, 24, A. Konieczna-Molenda; (2012) Effect of illumination of cellulose with the visible linearly polarized light upon digestion of cellulose, Recent. Devel. Carbohydrate Res. 3, A. Konieczna-Molenda, M. Fiedorowicz, W. Zhong, P. Tomasik, (2008) Polarized light-stimulated enzymatic hydrolysis of chitin and chitosan; Carbohydr. Res. 343, A. Konieczna-Molenda, M. Fiedorowicz, P. J. Tomasik, (2010) Stimulation of Glucose Oxidase with White Linearly Polarized Light, Biotechnology Progress, 26(1) M. Fiedorowicz, G. Khachatryan, A. Konieczna-Molenda, P. Tomasik, (2009) Formation of cyclodextrin with cyclodextringlucosyltransferase stimulated with polarized light, Biotechnogy Progress, 25 (1), M. Fiedorowicz, A. Konieczna-Molenda, G. Khachatryan, P. Tomasik; (2010) Sposób stymulowania katalizatorów białkowych używanych w reakcjach biochemicznych, zwłaszcza enzymów stosowanych w reakcjach otrzymywania sacharydów, polski patent PL
51 W kolejnych moich badaniach zajęłam się immobilizacją enzymów na nośnikach polimerowych. Badania odbywały się we współpracy z Zakładem Technologii Chemicznej i Zespołem Polimerów, Wydziału Chemii Uniwersytetu Jagiellońskiego, gdzie wykonano syntezy polimerów. Powszechnie stosowane w przemyśle enzymy immobilizowałam różnymi metodami na nośnikach polimerowych poli(winyloamidowych) oraz poli(winyloforamidowych) aktywowanych lub nie aldehydem glutarowym 24. Otrzymałam biokatalizatory aktywne w reakcjach hydrolizy skrobi 24, ksylanu, chityny i chitozanu 25. Opracowałam metody immobilizacji celulaz na nośnikach poli(amidowych) i poli(formamidowych) 26,27. Zbadałam sposób wiązania enzymu z nośnikiem i określiłam aktywność immobilizowanych enzymów w reakcjach hydrolizy 28,29. Otrzymane biokatalizatory posiadały wysoką aktywność w reakcjach hydrolizy celulozy oraz produktów celulozowych. Ostatnim wątkiem moich zainteresowań badawczych było otrzymywanie i badanie biodegradowalnych kompleksów 30 i folii 31 do żywności oraz bionanokompozytów posiadających potencjalne zastosowanie jako sensory do określania świeżości żywności A. Konieczna-Molenda, A. Kochanowski, A. Walaszek, E. Bortel and P. Tomasik; (2009) Immobilization of α-amylase on poly(vinylamine) and poly(vinylformamide) supports and its performance, Chemical Engineering Journal, 146, 3(15) A. Konieczna-Molenda, A, Kochanowski, A. Grabiec, E. Bortel, P. Tomasik, (2008) The Role of Polymeric Enzyme Carriers Based on Crosslinked Poly(N-vinylformamide) Polymers in Enhancing and Reducing Enzymatic Activities; Starch: Recent Progress in Biopolymer and Enzyme Technology, PTTZ-Malopolska Branch, Krakow, 8, K. Sokołowska, A. Konieczna-Molenda, E. Witek, (2016) Hydroliza odpadów celulozowych katalizowana enzymami celulolitycznymi immobilizowanymi na nośniku polimerowym, Polimery, 9, K. Sokołowska, A. Konieczna-Molenda, E. Witek, (2015) Cellulase immobilized on copolymers based on N- vinylformamide: Influence of immobilization on the catalytic activity of the enzyme (Celulaza immobilizowana na kopolimerach N-winyloformamidu: Wpływ immobilizacji na aktywność katalityczną enzymu), Chemik 69, A. Tata, K. Sokołowska, J. Świder, A. Konieczna-Molenda, E. Proniewicz, E. Witek, (2015) Study of cellulolytic enzyme immobilization on copolymers of N-vinylformamide, Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy, L. Krzemińska-Fiedorowicz, G. Khachatryan, A. Konieczna-Molenda (2018) Studies of Cellulose Hydrolysis Kinetics Affected Immobilized Cellulase, Procceedings of the 14th International Conference on Polysaccharides- Glycoscience. Prague, Czech Republic, Prague, Czech Republic, E. Jamróz, A. Konieczna-Molenda, A. Para, (2018) Otrzymywanie i charakterystyka binarnych kompleksów furcelleranu z żelatyną i albuminą surowicy bydlęcej, Polimery, 6, E. Jamróz, A. Konieczna-Molenda, A. Para, (2017) Trójskładnikowa folia: skrobia ziemniaczana-furcellaranżelatyna jako nowa generacja biodegradowalnych folii, Polimery, 9, K. Khachatryan, G. Khachatryan, A. Konieczna-Molenda, M. Janik, (2018) Bionanocomposite of ZnS nanoparticles in carboxymethyl cellulose as sensor of food freshness, Procceedings of the 14th International Conference on Polysaccharides-Glycoscience. Prague, Czech Republic,
52 Udział w projektach: 1) r. N N Synteza i spektroskopowa charakterystyka oraz badania aktywności biologicznej enzymatycznych katalizatorów immobilizowanych na nowych nośnikach polimerowych, Kierownik: Prof. dr hab. Edyta Proniewicz. Instytucja realizująca: Uniwersytet Jagielloński, Wydział Chemii (wykonawca), 2) r. N R Nanosensory do monitorowania zawartości, stanu świeżości i zawilgocenia żywności w opakowaniach foliowych, Kierownik: Prof. dr hab. Maciej Fiedorowicz. Instytucja realizująca: Uniwersytet Rolniczy im. Hugona Kołłątaja w Krakowie, Wydział Rolniczo-Ekonomiczny (wykonawca), 3) r. projekt badawczy 3T00B pt. Kinetyka degradacji celulozy w zakresie podwyższonych temperatur. Instytucja realizująca: Uniwersytet Jagielloński, Wydział Chemii promotorski (główny wykonawca), 4) r. Wieloletni Projekt Rządowy pt. Kwaśny papier finansowany przez MEN, Kierownik: Prof. dr hab. Andrzej Barański. Instytucja realizująca: Uniwersytet Jagielloński, Wydział Chemii (wykonawca), 5) r. projekt badawczy 3T09B pt. Naukowe podstawy studiów nad degradacją papieru dla potrzeb masowego odkwaszania zagrożonych zasobów archiwalnych i bibliotecznych w Polsce, Kierownik: Prof. dr hab. Andrzej Barański. Instytucja realizująca: Uniwersytet Jagielloński, Wydział Chemii (wykonawca), 6) r. projekt pt. Wiedza, praktyka, kadry klucz do sukcesu w biznesie dla przedsiębiorstw, pracowników przedsiębiorstw i ośrodków naukowo-badawczo-rozwojowych Projekt Małopolskiej Agencji Rozwoju Regionalnego SA, współfinansowany przez Unię Europejską w ramach Europejskiego Funduszu Społecznego (lider), 7) r. projekt Wiedza i kompetencje z fizyki, chemii i informatyki na potrzeby gospodarki WIKING organizowanym przez Uniwersytet Jagielloński, Kapitał Ludzki Narodowa strategia Spójności, projekt współfinansowany przez Unię Europejską w ramach Europejskiego Funduszu Społecznego (opiekun stażysty), 52
53 8) r. - projekt Wiedza i kompetencje z fizyki, chemii i informatyki na potrzeby gospodarki WIKING organizowany przez Uniwersytet Jagielloński, Kapitał Ludzki Narodowa Strategia Spójności, projekt współfinansowany przez Unię Europejską w ramach Europejskiego Funduszu Społecznego (opiekun stażysty), 9) projekt pt. Wędka technologiczna organizowany przez Zespół Badawczy Integracja Wydział Nauk Ekonomicznych i Zarządzania, Uniwersytet Szczeciński, realizowany z Zakładem Farmaceutycznym AMARA w Krakowie (wykonawca), Odbyłam zagraniczny staż naukowy w 2000r. w TNO Centre for Paper and Board Research, Delft, Holandia (1 miesiąc). Pracowałam na stanowisku Kierownika Laboratorium Kontrolnego (1 rok), Kierownika Kontroli Jakości (7 miesięcy) oraz Specjalisty ds. Naukowo- Badawczych (ponad 8 lat) w Zakładzie Farmaceutycznym AMARA w Krakowie. Nagrody: stypendium z Własnego Funduszu Stypendialnego, Rektora Uniwersytetu Rolniczego w Krakowie, r. stypendium z Rektorskiego Funduszu Stypendialnego na realizację projektu badawczego pt. Immobilizacja enzymów stymulowanych światłem, 2003r. wyróżnienie rozprawy doktorskiej pt. Kinetyka degradacji celulozy w zakresie podwyższonych temperatur przyznane przez Radę Wydziału Chemii Uniwersytetu Jagiellońskiego. Brałam czynny udział w konferencjach krajowych i zagranicznych: 1) EUPOC Biobased Polymers and Related Biomaterials, 2011, 2) International Symposium on Bioelectrochemistry and Bioenergetics of the Bioelectrochemical Society, 2011, 3) International Starch Convention, 2010, 2008, 2006, 3) (Bio)Degradable Polymers from Renewable Resources, 2007, 5) 3rd Workshop of Young European Scientists YES, 2007, 53
54 6) Cultural Heritage Research: a Pan-european Challenge, 5th EC Conference, 2002, 7) European Materials Research Society Spring Meeting, 2001, 8) Zjazd PTChem, 2018, 2015, 2014, 2002, 9) Kongres Technologii Chemicznej, 2018, 2015, 2012, 2003, 2000, 10) Ogólnopolskie Kolokwium Katalityczne, 2015,
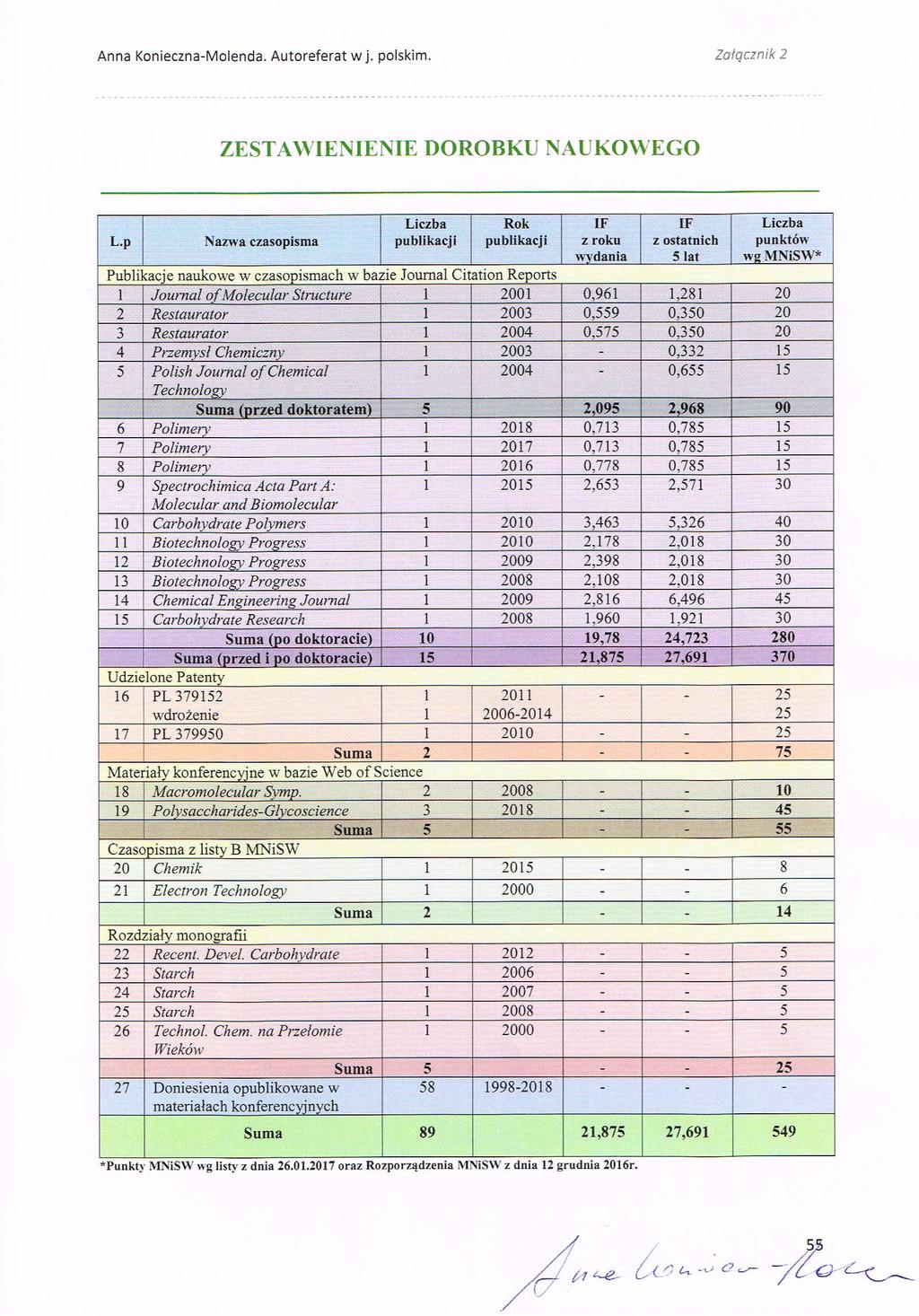
55
Sposób stymulowania katalizatorów białkowych używanych w reakcjach biochemicznych, zwłaszcza enzymów stosowanych w reakcjach otrzymywania sacharydów
 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 211549 (13) B1 (21) Numer zgłoszenia: 379950 (51) Int.Cl. C12Q 1/25 (2006.01) G01N 21/17 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22)
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 211549 (13) B1 (21) Numer zgłoszenia: 379950 (51) Int.Cl. C12Q 1/25 (2006.01) G01N 21/17 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22)
Projektowanie Procesów Biotechnologicznych
 Projektowanie Procesów Biotechnologicznych wykład 14 styczeń 2014 Kinetyka prostych reakcji enzymatycznych Kinetyka hamowania reakcji enzymatycznych 1 Enzymy - substancje białkowe katalizujące przemiany
Projektowanie Procesów Biotechnologicznych wykład 14 styczeń 2014 Kinetyka prostych reakcji enzymatycznych Kinetyka hamowania reakcji enzymatycznych 1 Enzymy - substancje białkowe katalizujące przemiany
Podstawy biogospodarki. Wykład 7
 Podstawy biogospodarki Wykład 7 Prowadzący: Krzysztof Makowski Kierunek Wyróżniony przez PKA Immobilizowane białka Kierunek Wyróżniony przez PKA Krzysztof Makowski Instytut Biochemii Technicznej Politechniki
Podstawy biogospodarki Wykład 7 Prowadzący: Krzysztof Makowski Kierunek Wyróżniony przez PKA Immobilizowane białka Kierunek Wyróżniony przez PKA Krzysztof Makowski Instytut Biochemii Technicznej Politechniki
Planowanie Projektów Odnawialnych Źródeł Energii Biomasa (odpady fermentowalne)
") Slajd 1 Lennart Tyrberg, Energy Agency of Southeast Sweden Planowanie Projektów Odnawialnych Źródeł Energii Biomasa (odpady fermentowalne) Prepared by: Mgr inż. Andrzej Michalski Verified by: Dr inż. Andrzej
Slajd 1 Lennart Tyrberg, Energy Agency of Southeast Sweden Planowanie Projektów Odnawialnych Źródeł Energii Biomasa (odpady fermentowalne) Prepared by: Mgr inż. Andrzej Michalski Verified by: Dr inż. Andrzej
Definicja immobilizacji
 Definicja immobilizacji Immobilizacja technika unieruchamiania biokatalizatorów / enzymów na nośnikach, stosowana powszechnie w badaniach naukowych i przemyśle (chemicznym, spożywczym) Problemy związane
Definicja immobilizacji Immobilizacja technika unieruchamiania biokatalizatorów / enzymów na nośnikach, stosowana powszechnie w badaniach naukowych i przemyśle (chemicznym, spożywczym) Problemy związane
BIOSYNTEZA ACYLAZY PENICYLINOWEJ. Ćwiczenia z Mikrobiologii Przemysłowej 2011
 BIOSYNTEZA ACYLAZY PENICYLINOWEJ Ćwiczenia z Mikrobiologii Przemysłowej 2011 Acylaza penicylinowa Enzym hydrolizuje wiązanie amidowe w penicylinach Reakcja przebiega wg schematu: acylaza Reszta: fenyloacetylowa
BIOSYNTEZA ACYLAZY PENICYLINOWEJ Ćwiczenia z Mikrobiologii Przemysłowej 2011 Acylaza penicylinowa Enzym hydrolizuje wiązanie amidowe w penicylinach Reakcja przebiega wg schematu: acylaza Reszta: fenyloacetylowa
Synteza nanocząstek magnetycznych pokrytych modyfikowaną skrobią dla zastosowań biomedycznych
 Synteza nanocząstek magnetycznych pokrytych modyfikowaną skrobią dla zastosowań biomedycznych mgr Katarzyna Węgrzynowska-Drzymalska Katedra Chemii i Fotochemii Polimerów Wydział Chemii Uniwersytet Mikołaja
Synteza nanocząstek magnetycznych pokrytych modyfikowaną skrobią dla zastosowań biomedycznych mgr Katarzyna Węgrzynowska-Drzymalska Katedra Chemii i Fotochemii Polimerów Wydział Chemii Uniwersytet Mikołaja
Cukry właściwości i funkcje
 Cukry właściwości i funkcje Miejsce cukrów wśród innych składników chemicznych Cukry Z cukrem mamy do czynienia bardzo często - kiedy sięgamy po białe kryształy z cukiernicy. Większość z nas nie uświadamia
Cukry właściwości i funkcje Miejsce cukrów wśród innych składników chemicznych Cukry Z cukrem mamy do czynienia bardzo często - kiedy sięgamy po białe kryształy z cukiernicy. Większość z nas nie uświadamia
SPIS TREŚCI OD AUTORÓW... 5
 SPIS TREŚCI OD AUTORÓW... 5 BIAŁKA 1. Wprowadzenie... 7 2. Aminokwasy jednostki strukturalne białek... 7 2.1. Klasyfikacja aminokwasów... 9 2.1.1. Aminokwasy białkowe i niebiałkowe... 9 2.1.2. Zdolność
SPIS TREŚCI OD AUTORÓW... 5 BIAŁKA 1. Wprowadzenie... 7 2. Aminokwasy jednostki strukturalne białek... 7 2.1. Klasyfikacja aminokwasów... 9 2.1.1. Aminokwasy białkowe i niebiałkowe... 9 2.1.2. Zdolność
1. Biotechnologia i inżynieria genetyczna zagadnienia wstępne 13
 Spis treści Przedmowa 11 1. Biotechnologia i inżynieria genetyczna zagadnienia wstępne 13 1.1. Wprowadzenie 13 1.2. Biotechnologia żywności znaczenie gospodarcze i społeczne 13 1.3. Produkty modyfikowane
Spis treści Przedmowa 11 1. Biotechnologia i inżynieria genetyczna zagadnienia wstępne 13 1.1. Wprowadzenie 13 1.2. Biotechnologia żywności znaczenie gospodarcze i społeczne 13 1.3. Produkty modyfikowane
Przemiana materii i energii - Biologia.net.pl
 Ogół przemian biochemicznych, które zachodzą w komórce składają się na jej metabolizm. Wyróżnia się dwa antagonistyczne procesy metabolizmu: anabolizm i katabolizm. Szlak metaboliczny w komórce, to szereg
Ogół przemian biochemicznych, które zachodzą w komórce składają się na jej metabolizm. Wyróżnia się dwa antagonistyczne procesy metabolizmu: anabolizm i katabolizm. Szlak metaboliczny w komórce, to szereg
Laboratorium 6. Immobilizacja enzymu metodą pułapkowania w matrycy hydrożelowej
 Laboratorium 6 Immobilizacja enzymu metodą pułapkowania w matrycy hydrożelowej Prowadzący: dr inż. Karolina Labus 1. CZĘŚĆ TEORETYCZNA Enzymy uważane są za niezwykle wydajne katalizatory różnorodnych reakcji,
Laboratorium 6 Immobilizacja enzymu metodą pułapkowania w matrycy hydrożelowej Prowadzący: dr inż. Karolina Labus 1. CZĘŚĆ TEORETYCZNA Enzymy uważane są za niezwykle wydajne katalizatory różnorodnych reakcji,
OFERTA TEMATÓW PROJEKTÓW DYPLOMOWYCH (MAGISTERSKICH) do zrealizowania w Katedrze INŻYNIERII CHEMICZNEJ I PROCESOWEJ
 do zrealizowania w Katedrze INŻYNIERII CHEMICZNEJ I PROCESOWEJ") OFERTA TEMATÓW PROJEKTÓW DYPLOMOWYCH (MAGISTERSKICH) do zrealizowania w Katedrze INŻYNIERII CHEMICZNEJ I PROCESOWEJ Badania kinetyki utleniania wybranych grup związków organicznych podczas procesów oczyszczania
OFERTA TEMATÓW PROJEKTÓW DYPLOMOWYCH (MAGISTERSKICH) do zrealizowania w Katedrze INŻYNIERII CHEMICZNEJ I PROCESOWEJ Badania kinetyki utleniania wybranych grup związków organicznych podczas procesów oczyszczania
Błonnik pokarmowy: właściwości, skład, występowanie w żywności
 Błonnik pokarmowy: właściwości, skład, występowanie w żywności Dr hab. Jarosława Rutkowska, prof. nadzwycz. SGGW Zakład Analiz Instrumentalnych Wydział Nauk o Żywieniu Człowieka i Konsumpcji, SGGW w Warszawie
Błonnik pokarmowy: właściwości, skład, występowanie w żywności Dr hab. Jarosława Rutkowska, prof. nadzwycz. SGGW Zakład Analiz Instrumentalnych Wydział Nauk o Żywieniu Człowieka i Konsumpcji, SGGW w Warszawie
Hydroliza skrobi przez unieruchomioną glukoamylazę
 Hydroliza skrobi przez unieruchomioną glukoamylazę Ćwiczenie ma na celu poznanie jednej z metod unieruchamiania enzymów oraz przeprowadzenie procesu ciągłej hydrolizy skrobi przez unieruchomioną glukoamylazę.
Hydroliza skrobi przez unieruchomioną glukoamylazę Ćwiczenie ma na celu poznanie jednej z metod unieruchamiania enzymów oraz przeprowadzenie procesu ciągłej hydrolizy skrobi przez unieruchomioną glukoamylazę.
Wydział Chemiczny Politechniki Gdańskiej Katedra Technologii Leków i Biochemii
 Wydział Chemiczny Politechniki Gdańskiej Katedra Technologii Leków i Biochemii Badanie wpływu temperatury, ph, aktywatorów i inhibitorów na aktywność α-amylazy Wstęp Szybkość reakcji enzymatycznej jest
Wydział Chemiczny Politechniki Gdańskiej Katedra Technologii Leków i Biochemii Badanie wpływu temperatury, ph, aktywatorów i inhibitorów na aktywność α-amylazy Wstęp Szybkość reakcji enzymatycznej jest
Mechanizmy działania i regulacji enzymów
 Mechanizmy działania i regulacji enzymów Enzymy: są katalizatorami, które zmieniają szybkość reakcji, same nie ulegając zmianie są wysoce specyficzne ich aktywność może być regulowana m.in. przez modyfikacje
Mechanizmy działania i regulacji enzymów Enzymy: są katalizatorami, które zmieniają szybkość reakcji, same nie ulegając zmianie są wysoce specyficzne ich aktywność może być regulowana m.in. przez modyfikacje
Enzymy katalizatory biologiczne
 Enzymy katalizatory biologiczne Kataliza zjawisko polegające na obniżeniu energii aktywacji reakcji i zwiększeniu szybkości reakcji chemicznej i/lub skierowaniu reakcji na jedną z termodynamicznie możliwych
Enzymy katalizatory biologiczne Kataliza zjawisko polegające na obniżeniu energii aktywacji reakcji i zwiększeniu szybkości reakcji chemicznej i/lub skierowaniu reakcji na jedną z termodynamicznie możliwych
Disacharydy. Chemia Medyczna dr inż.. Ewa Mironiuk-Puchalska, W CHem PW 1. disacharydy redukujace. disacharydy nieredukujace. atom anomeryczny.
 Disacharydy Disacharydy cukry hydrolizujące (pod wpływem roztworów kwasów mineralnych, maltazy wiązania -glikozydowe, emulsyny wiązania -glikozydowe) glikozydy (powstają z dwóch reszt monosacharydowych
Disacharydy Disacharydy cukry hydrolizujące (pod wpływem roztworów kwasów mineralnych, maltazy wiązania -glikozydowe, emulsyny wiązania -glikozydowe) glikozydy (powstają z dwóch reszt monosacharydowych
KINETYKA HYDROLIZY SACHAROZY
 Ćwiczenie nr 2 KINETYKA HYDROLIZY SACHAROZY I. Kinetyka hydrolizy sacharozy reakcja chemiczna Zasada: Sacharoza w środowisku kwaśnym ulega hydrolizie z wytworzeniem -D-glukozy i -D-fruktozy. Jest to reakcja
Ćwiczenie nr 2 KINETYKA HYDROLIZY SACHAROZY I. Kinetyka hydrolizy sacharozy reakcja chemiczna Zasada: Sacharoza w środowisku kwaśnym ulega hydrolizie z wytworzeniem -D-glukozy i -D-fruktozy. Jest to reakcja
SACHARYDY MONOSACHARYDY POLISACHARYDY OLIGOSACHARYDY
 SACHARYDY MONOSACHARYDY POLISACHARYDY OLIGOSACHARYDY C x H 2y O y y = 2-10 Oligosacharydy oligomery węglowodanowe, które zawierają od 2 do 10 monomerów, którymi są cukry proste (monosacharydy), np. glukoza,
SACHARYDY MONOSACHARYDY POLISACHARYDY OLIGOSACHARYDY C x H 2y O y y = 2-10 Oligosacharydy oligomery węglowodanowe, które zawierają od 2 do 10 monomerów, którymi są cukry proste (monosacharydy), np. glukoza,
Potencjał naukowo badawczy Wydziału Technologii Żywności, Uniwersytetu Rolniczego w Krakowie
 Potencjał naukowo badawczy Wydziału Technologii Żywności, Uniwersytetu Rolniczego w Krakowie 1 Wydział Technologii Żywności UR w Krakowie Kontakt: Ul. Balicka 122, 30-149 Kraków Strona internetowa: wtz.ur.krakow.pl
Potencjał naukowo badawczy Wydziału Technologii Żywności, Uniwersytetu Rolniczego w Krakowie 1 Wydział Technologii Żywności UR w Krakowie Kontakt: Ul. Balicka 122, 30-149 Kraków Strona internetowa: wtz.ur.krakow.pl
Szczegółowe plany specjalizacji na studiach niestacjonarnych II stopnia
 Szczegółowe plany specjalizacji na studiach niestacjonarnych II stopnia Kierunek studiów: Technologia żywności i żywienie człowieka Specjalność: Technologia żywności Specjalizacja : ANALIZY I OCENY JAKOŚCI
Szczegółowe plany specjalizacji na studiach niestacjonarnych II stopnia Kierunek studiów: Technologia żywności i żywienie człowieka Specjalność: Technologia żywności Specjalizacja : ANALIZY I OCENY JAKOŚCI
POLITECHNIKA GDAŃSKA WYDZIAŁ CHEMICZNY KATEDRA TECHNOLOGII POLIMERÓW
 POLITECHNIKA GDAŃSKA WYDZIAŁ CHEMICZNY KATEDRA TECHNOLOGII POLIMERÓW PRZETWÓRSTWO TWORZYW SZTUCZNYCH I GUMY Lab 8. Wyznaczanie optimum wulkanizacji mieszanek kauczukowych na reometrze Monsanto oraz analiza
POLITECHNIKA GDAŃSKA WYDZIAŁ CHEMICZNY KATEDRA TECHNOLOGII POLIMERÓW PRZETWÓRSTWO TWORZYW SZTUCZNYCH I GUMY Lab 8. Wyznaczanie optimum wulkanizacji mieszanek kauczukowych na reometrze Monsanto oraz analiza
Zanieczyszczenia organiczne takie jak WWA czy pestycydy są dużym zagrożeniem zarówno dla środowiska jak i zdrowia i życia człowieka.
 Projekt współfinansowany ze środków Narodowego Centrum Nauki (NCN) oraz Narodowego Centrum Badań i Rozwoju (NCBIR) w ramach projektu (TANGO1/266740/NCBR/2015) Mgr Dariusz Włóka Autor jest stypendystą programu
Projekt współfinansowany ze środków Narodowego Centrum Nauki (NCN) oraz Narodowego Centrum Badań i Rozwoju (NCBIR) w ramach projektu (TANGO1/266740/NCBR/2015) Mgr Dariusz Włóka Autor jest stypendystą programu
Zestaw pytań egzaminu inŝynierskiego przeprowadzanego w Katedrze Fizykochemii i Technologii Polimerów dla kierunku CHEMIA
 Zestaw pytań egzaminu inŝynierskiego przeprowadzanego w Katedrze Fizykochemii i Technologii Polimerów dla kierunku CHEMIA 1. Metody miareczkowania w analizie chemicznej, wyjaśnić działanie wskaźników 2.
Zestaw pytań egzaminu inŝynierskiego przeprowadzanego w Katedrze Fizykochemii i Technologii Polimerów dla kierunku CHEMIA 1. Metody miareczkowania w analizie chemicznej, wyjaśnić działanie wskaźników 2.
Hydroliza odpadów celulozowych katalizowana enzymami celulolitycznymi immobilizowanymi na nośniku polimerowym* )
") 2016, 61, nr 9 633 Hydroliza odpadów celulozowych katalizowana enzymami celulolitycznymi immobilizowanymi na nośniku polimerowym* ) Katarzyna Sokołowska 1), Anna Konieczna-Molenda 2), Ewa Witek 1), **
2016, 61, nr 9 633 Hydroliza odpadów celulozowych katalizowana enzymami celulolitycznymi immobilizowanymi na nośniku polimerowym* ) Katarzyna Sokołowska 1), Anna Konieczna-Molenda 2), Ewa Witek 1), **
Spis treści. asf;mfzjf. (Jan Fiedurek)
") asf;mfzjf Spis treści 1. Informacje wstępne 11 (Jan Fiedurek) 1.1. Biotechnologia w ujęciu historycznym i perspektywicznym... 12 1.2. Biotechnologia klasyczna i nowoczesna... 18 1.3. Rozwój biotechnologii:
asf;mfzjf Spis treści 1. Informacje wstępne 11 (Jan Fiedurek) 1.1. Biotechnologia w ujęciu historycznym i perspektywicznym... 12 1.2. Biotechnologia klasyczna i nowoczesna... 18 1.3. Rozwój biotechnologii:
BIOTECHNOLOGIA OGÓLNA
 BIOTECHNOLOGIA OGÓLNA 1. 2. 3. 4. 5. Ogólne podstawy biologicznych metod oczyszczania ścieków. Ścieki i ich rodzaje. Stosowane metody analityczne. Substancje biogenne w ściekach. Tlenowe procesy przemiany
BIOTECHNOLOGIA OGÓLNA 1. 2. 3. 4. 5. Ogólne podstawy biologicznych metod oczyszczania ścieków. Ścieki i ich rodzaje. Stosowane metody analityczne. Substancje biogenne w ściekach. Tlenowe procesy przemiany
Rys. 1 Mechanizm katalizy hydrolizy skrobi z udziałem glukoamylazy (A Glu179, B Glu400)
") Hydroliza skrobi przez unieruchomioną glukoamylazę Ćwiczenie ma na celu poznanie jednej z metod unieruchamiania enzymów oraz przeprowadzenie procesu ciągłej hydrolizy skrobi przez unieruchomiony enzym
Hydroliza skrobi przez unieruchomioną glukoamylazę Ćwiczenie ma na celu poznanie jednej z metod unieruchamiania enzymów oraz przeprowadzenie procesu ciągłej hydrolizy skrobi przez unieruchomiony enzym
KINETYKA HYDROLIZY SACHAROZY (REAKCJA ENZYMATYCZNA I CHEMICZNA)
") Ćwiczenie nr 2 KINETYKA HYDROLIZY SACHAROZY (REAKCJA ENZYMATYCZNA I CHEMICZNA) ĆWICZENIE PRAKTYCZNE I. Kinetyka hydrolizy sacharozy reakcja chemiczna Zasada: Sacharoza w środowisku kwaśnym ulega hydrolizie
Ćwiczenie nr 2 KINETYKA HYDROLIZY SACHAROZY (REAKCJA ENZYMATYCZNA I CHEMICZNA) ĆWICZENIE PRAKTYCZNE I. Kinetyka hydrolizy sacharozy reakcja chemiczna Zasada: Sacharoza w środowisku kwaśnym ulega hydrolizie
Niestandardowe wykorzystanie buraków cukrowych
 Niestandardowe wykorzystanie buraków cukrowych Stanisław Wawro, Radosław Gruska, Agnieszka Papiewska, Maciej Stanisz Instytut Chemicznej Technologii Żywności Skład chemiczny korzeni dojrzałych buraków
Niestandardowe wykorzystanie buraków cukrowych Stanisław Wawro, Radosław Gruska, Agnieszka Papiewska, Maciej Stanisz Instytut Chemicznej Technologii Żywności Skład chemiczny korzeni dojrzałych buraków
Laboratorium 4. Określenie aktywności katalitycznej enzymu. Wprowadzenie do metod analitycznych. 1. CZĘŚĆ TEORETYCZNA
 Laboratorium 4 Określenie aktywności katalitycznej enzymu. Wprowadzenie do metod analitycznych. Prowadzący: dr inż. Karolina Labus 1. CZĘŚĆ TEORETYCZNA Enzymy to wielkocząsteczkowe, w większości białkowe,
Laboratorium 4 Określenie aktywności katalitycznej enzymu. Wprowadzenie do metod analitycznych. Prowadzący: dr inż. Karolina Labus 1. CZĘŚĆ TEORETYCZNA Enzymy to wielkocząsteczkowe, w większości białkowe,
Laboratorium 5. Wpływ temperatury na aktywność enzymów. Inaktywacja termiczna
 Laboratorium 5 Wpływ temperatury na aktywność enzymów. Inaktywacja termiczna Prowadzący: dr inż. Karolina Labus 1. CZĘŚĆ TEORETYCZNA Szybkość reakcji enzymatycznej zależy przede wszystkim od stężenia substratu
Laboratorium 5 Wpływ temperatury na aktywność enzymów. Inaktywacja termiczna Prowadzący: dr inż. Karolina Labus 1. CZĘŚĆ TEORETYCZNA Szybkość reakcji enzymatycznej zależy przede wszystkim od stężenia substratu
PLAN STUDIÓW NR II PROFIL OGÓLNOAKADEMICKI POZIOM STUDIÓW: STUDIA DRUGIEGO STOPNIA (1,5-roczne magisterskie) FORMA STUDIÓW:
 FORMA STUDIÓW:") UNIWERSYTET TECHNOLOGICZNO-PRZYRODNICZY 1.TECHNOLOGIA PROCESÓW CHEMICZNYCH 2. BIOTECHNOLOGIA PRZEMYSŁOWA 3. ANALITYKA CHEMICZNA I SPOŻYWCZA 4. NOWOCZESNE TECHNOLOGIE MATERIAŁOWE godzin tygodniowo (semestr
UNIWERSYTET TECHNOLOGICZNO-PRZYRODNICZY 1.TECHNOLOGIA PROCESÓW CHEMICZNYCH 2. BIOTECHNOLOGIA PRZEMYSŁOWA 3. ANALITYKA CHEMICZNA I SPOŻYWCZA 4. NOWOCZESNE TECHNOLOGIE MATERIAŁOWE godzin tygodniowo (semestr
3b 2. przedstawione na poniższych schematach. Uzupełnij obserwacje i wnioski z nich wynikające oraz równanie zachodzącej reakcji.
 3b 2 PAWEŁ ZYCH IMIĘ I NAZWISKO: KLASA: GRUPA A 1. W celu zbadania właściwości sacharozy wykonano dwa doświadczenia, które zostały przedstawione na poniższych schematach. Uzupełnij obserwacje i wnioski
3b 2 PAWEŁ ZYCH IMIĘ I NAZWISKO: KLASA: GRUPA A 1. W celu zbadania właściwości sacharozy wykonano dwa doświadczenia, które zostały przedstawione na poniższych schematach. Uzupełnij obserwacje i wnioski
PL B1. Preparat o właściwościach przeciwutleniających oraz sposób otrzymywania tego preparatu. POLITECHNIKA ŁÓDZKA, Łódź, PL
 PL 217050 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 217050 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 388203 (22) Data zgłoszenia: 08.06.2009 (51) Int.Cl.
PL 217050 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 217050 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 388203 (22) Data zgłoszenia: 08.06.2009 (51) Int.Cl.
12. PRZYGOTOWANIE BIOMASY DO PROCESU FERMENTACJI WODOROWEJ
 12. PRZYGOTOWANIE BIOMASY DO PROCESU FERMENTACJI WODOROWEJ 12.1. Wstęp Wodór, ze względu na własności energetyczne i ekologiczne, jest uważany za najlepszy nośnik energii przyszłości. Spośród wielu sposobów
12. PRZYGOTOWANIE BIOMASY DO PROCESU FERMENTACJI WODOROWEJ 12.1. Wstęp Wodór, ze względu na własności energetyczne i ekologiczne, jest uważany za najlepszy nośnik energii przyszłości. Spośród wielu sposobów
Oznaczanie aktywności - i β- amylazy słodu metodą kolorymetryczną
 KATEDRA BIOCHEMII Wydział Biologii i Ochrony Środowiska Oznaczanie aktywności - i β- amylazy słodu metodą kolorymetryczną ĆWICZENIE 5 OZNACZANIE AKTYWNOŚCI -AMYLAZY SŁODU METODĄ KOLORYMETRYCZNĄ Enzymy
KATEDRA BIOCHEMII Wydział Biologii i Ochrony Środowiska Oznaczanie aktywności - i β- amylazy słodu metodą kolorymetryczną ĆWICZENIE 5 OZNACZANIE AKTYWNOŚCI -AMYLAZY SŁODU METODĄ KOLORYMETRYCZNĄ Enzymy
Chemiczne składniki komórek
 Chemiczne składniki komórek Pierwiastki chemiczne w komórkach: - makroelementy (pierwiastki biogenne) H, O, C, N, S, P Ca, Mg, K, Na, Cl >1% suchej masy - mikroelementy Fe, Cu, Mn, Mo, B, Zn, Co, J, F
Chemiczne składniki komórek Pierwiastki chemiczne w komórkach: - makroelementy (pierwiastki biogenne) H, O, C, N, S, P Ca, Mg, K, Na, Cl >1% suchej masy - mikroelementy Fe, Cu, Mn, Mo, B, Zn, Co, J, F
Badanie termostabilności oraz wpływu aktywatorów i inhibitorów na działanie α-amylazy [EC ]
![Badanie termostabilności oraz wpływu aktywatorów i inhibitorów na działanie α-amylazy [EC ]](/thumbs/66/55837946.jpg "Badanie termostabilności oraz wpływu aktywatorów i inhibitorów na działanie α-amylazy [EC ]") Badanie termostabilności oraz wpływu aktywatorów i inhibitorów na działanie α-amylazy [EC 3.2.1.1.] Termostabilność enzymów Dla większości enzymów zmiany denaturacyjne zachodzą bardzo intensywnie powyżej
Badanie termostabilności oraz wpływu aktywatorów i inhibitorów na działanie α-amylazy [EC 3.2.1.1.] Termostabilność enzymów Dla większości enzymów zmiany denaturacyjne zachodzą bardzo intensywnie powyżej
Kinetyka reakcji hydrolizy sacharozy katalizowanej przez inwertazę
 Kinetyka reakcji hydrolizy sacharozy katalizowanej przez inwertazę Prowadzący: dr hab. inż. Ilona WANDZIK mgr inż. Sebastian BUDNIOK mgr inż. Marta GREC mgr inż. Jadwiga PASZKOWSKA Miejsce ćwiczenia: sala
Kinetyka reakcji hydrolizy sacharozy katalizowanej przez inwertazę Prowadzący: dr hab. inż. Ilona WANDZIK mgr inż. Sebastian BUDNIOK mgr inż. Marta GREC mgr inż. Jadwiga PASZKOWSKA Miejsce ćwiczenia: sala
Co to jest FERMENTACJA?
 Co to jest FERMENTACJA? FERMENTACJA - rozkład niektórych monosacharydów, np. glukozy, pod wpływem enzymów wydzielanych przez drożdże lub bakterie. czyli tzw. biokatalizatorów. Enzymy (biokatalizatory)
Co to jest FERMENTACJA? FERMENTACJA - rozkład niektórych monosacharydów, np. glukozy, pod wpływem enzymów wydzielanych przez drożdże lub bakterie. czyli tzw. biokatalizatorów. Enzymy (biokatalizatory)
Wykład 3. Zielona chemia (część 2)
") Wykład 3 Zielona chemia (część 2) Glicerol jako zielony rozpuszczalnik Nietoksyczny, tani, łatwo dostępny, odnawialny, wysoka temp. wrzenia (nie jest klasyfikowany jako LZO/VOC), polarny, może być stosowany
Wykład 3 Zielona chemia (część 2) Glicerol jako zielony rozpuszczalnik Nietoksyczny, tani, łatwo dostępny, odnawialny, wysoka temp. wrzenia (nie jest klasyfikowany jako LZO/VOC), polarny, może być stosowany
Procesy biotransformacji
 Biohydrometalurgia jest to dział techniki zajmujący się otrzymywaniem metali przy użyciu mikroorganizmów i wody. Ma ona charakter interdyscyplinarny obejmujący wiedzę z zakresu biochemii, geomikrobiologii,
Biohydrometalurgia jest to dział techniki zajmujący się otrzymywaniem metali przy użyciu mikroorganizmów i wody. Ma ona charakter interdyscyplinarny obejmujący wiedzę z zakresu biochemii, geomikrobiologii,
PRZYKŁADOWE ZADANIA WĘGLOWODANY
 PRZYKŁADOWE ZADANIA WĘGLOWODANY Zadanie 1216 (2 pkt) Przeczytaj poniższy tekst i zapisz poniżej nazwy cukrów X i Y, o których mowa. Kwasy nukleinowe są długimi łańcuchami poliestrowymi, zbudowanymi z połączonych
PRZYKŁADOWE ZADANIA WĘGLOWODANY Zadanie 1216 (2 pkt) Przeczytaj poniższy tekst i zapisz poniżej nazwy cukrów X i Y, o których mowa. Kwasy nukleinowe są długimi łańcuchami poliestrowymi, zbudowanymi z połączonych
Badanie oddziaływania polihistydynowych cyklopeptydów z jonami Cu 2+ i Zn 2+ w aspekcie projektowania mimetyków SOD
 Wydział Farmaceutyczny z Oddziałem Analityki Medycznej Badanie oddziaływania polihistydynowych cyklopeptydów z jonami Cu 2+ i Zn 2+ w aspekcie projektowania mimetyków SOD Aleksandra Kotynia PRACA DOKTORSKA
Wydział Farmaceutyczny z Oddziałem Analityki Medycznej Badanie oddziaływania polihistydynowych cyklopeptydów z jonami Cu 2+ i Zn 2+ w aspekcie projektowania mimetyków SOD Aleksandra Kotynia PRACA DOKTORSKA
DETEKCJA I USUWANIE BIOFILMU, PRZY UŻYCIU METOD ENZYMATYCZNYCH
 DETEKCJA I USUWANIE BIOFILMU, PRZY UŻYCIU METOD ENZYMATYCZNYCH Prezentują: Mariusz Wlazło Wojciech Ćwikliński 1 Polski Kongres Napojowy Toruń 2016 Agenda 1 2 3 Firma Hypred; Współpraca z Realco Technologia
DETEKCJA I USUWANIE BIOFILMU, PRZY UŻYCIU METOD ENZYMATYCZNYCH Prezentują: Mariusz Wlazło Wojciech Ćwikliński 1 Polski Kongres Napojowy Toruń 2016 Agenda 1 2 3 Firma Hypred; Współpraca z Realco Technologia
BIOETANOL Z BIOMASY KONOPNEJ JAKO POLSKI DODATEK DO PALIW PŁYNNYCH
 Europejski Fundusz Rolny na rzecz Rozwoju Obszarów Wiejskich: Europa inwestująca w obszary wiejskie. INNOWACYJNE ROZWIĄZANIA DLA POLSKIEGO ROLNICTWA Polskie rośliny włókniste i zielarskie dla innowacyjnej
Europejski Fundusz Rolny na rzecz Rozwoju Obszarów Wiejskich: Europa inwestująca w obszary wiejskie. INNOWACYJNE ROZWIĄZANIA DLA POLSKIEGO ROLNICTWA Polskie rośliny włókniste i zielarskie dla innowacyjnej
Moduły kształcenia. Efekty kształcenia dla programu kształcenia (kierunku) MK_06 Krystalochemia. MK_01 Chemia fizyczna i jądrowa
 MK_06 Krystalochemia. MK_01 Chemia fizyczna i jądrowa") Matryca efektów kształcenia określa relacje między efektami kształcenia zdefiniowanymi dla programu kształcenia (efektami kierunkowymi) i efektami kształcenia zdefiniowanymi dla poszczególnych modułów
Matryca efektów kształcenia określa relacje między efektami kształcenia zdefiniowanymi dla programu kształcenia (efektami kierunkowymi) i efektami kształcenia zdefiniowanymi dla poszczególnych modułów
CZYNNIKI WPŁYWAJĄCE NA SZYBKOŚĆ REAKCJI CHEMICZNYCH. ILOŚCIOWE ZBADANIE SZYBKOŚCI ROZPADU NADTLENKU WODORU.
 CZYNNIKI WPŁYWAJĄCE NA SZYBKOŚĆ REAKCJI CHEMICZNYCH. ILOŚCIOWE ZBADANIE SZYBKOŚCI ROZPADU NADTLENKU WODORU. Projekt zrealizowany w ramach Mazowieckiego programu stypendialnego dla uczniów szczególnie uzdolnionych
CZYNNIKI WPŁYWAJĄCE NA SZYBKOŚĆ REAKCJI CHEMICZNYCH. ILOŚCIOWE ZBADANIE SZYBKOŚCI ROZPADU NADTLENKU WODORU. Projekt zrealizowany w ramach Mazowieckiego programu stypendialnego dla uczniów szczególnie uzdolnionych
BIOTECHNOLOGIA OGÓLNA
 BIOTECHNOLOGIA OGÓLNA 1. Wprowadzenie do biotechnologii. Rys historyczny. Zakres i znaczenie nowoczesnej biotechnologii. Opracowanie procesu biotechnologicznego. 7. Produkcja biomasy. Białko mikrobiologiczne.
BIOTECHNOLOGIA OGÓLNA 1. Wprowadzenie do biotechnologii. Rys historyczny. Zakres i znaczenie nowoczesnej biotechnologii. Opracowanie procesu biotechnologicznego. 7. Produkcja biomasy. Białko mikrobiologiczne.
Wykład 4. Fizykochemia biopolimerów- wykład 4. Anna Ptaszek. 5 listopada Katedra Inżynierii i Aparatury Przemysłu Spożywczego
 Wykład 4 - wykład 4 Katedra Inżynierii i Aparatury Przemysłu Spożywczego 5 listopada 2013 1/30 Czym są biopolimery? To polimery pochodzenia naturalnego. Należą do nich polisacharydy i białka. 2/30 Polisacharydy
Wykład 4 - wykład 4 Katedra Inżynierii i Aparatury Przemysłu Spożywczego 5 listopada 2013 1/30 Czym są biopolimery? To polimery pochodzenia naturalnego. Należą do nich polisacharydy i białka. 2/30 Polisacharydy
Zasady i cele stosowania dodatków kiszonkarskich
 .pl https://www..pl Zasady i cele stosowania dodatków kiszonkarskich Autor: dr hab. inż. Rafał Bodarski Data: 1 kwietnia 2016 Wykorzystanie na szeroką skalę kiszonek jako podstawowych gospodarskich pasz
.pl https://www..pl Zasady i cele stosowania dodatków kiszonkarskich Autor: dr hab. inż. Rafał Bodarski Data: 1 kwietnia 2016 Wykorzystanie na szeroką skalę kiszonek jako podstawowych gospodarskich pasz
3b Do dwóch probówek, w których znajdowały się olej słonecznikowy i stopione masło, dodano. 2. Zaznacz poprawną odpowiedź.
 3b 1 PAWEŁ ZYCH IMIĘ I NAZWISKO: KLASA: GRUPA A 1. Do dwóch probówek, w których znajdowały się olej słonecznikowy i stopione masło, dodano roztworu manganianu(vii) potasu. Napisz, jakich obserwacji można
3b 1 PAWEŁ ZYCH IMIĘ I NAZWISKO: KLASA: GRUPA A 1. Do dwóch probówek, w których znajdowały się olej słonecznikowy i stopione masło, dodano roztworu manganianu(vii) potasu. Napisz, jakich obserwacji można
WYMAGANIA EDUKACYJNE w klasie III
 WYMAGANIA EDUKACYJNE w klasie III Nr lekcji Temat lekcji Treści nauczania (pismem pogrubionym zostały zaznaczone treści Podstawy Programowej) Węgiel i jego związki z wodorem Wymagania i kryteria ocen Uczeń:
WYMAGANIA EDUKACYJNE w klasie III Nr lekcji Temat lekcji Treści nauczania (pismem pogrubionym zostały zaznaczone treści Podstawy Programowej) Węgiel i jego związki z wodorem Wymagania i kryteria ocen Uczeń:
Lek od pomysłu do wdrożenia
 Lek od pomysłu do wdrożenia Lek od pomysłu do wdrożenia KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU
Lek od pomysłu do wdrożenia Lek od pomysłu do wdrożenia KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU
BIOTECHNOLOGIA OGÓLNA
 BIOTECHNOLOGIA OGÓLNA 1. Wprowadzenie do biotechnologii. Rys historyczny. Zakres i znaczenie nowoczesnej biotechnologii. Opracowanie procesu biotechnologicznego. 7. Produkcja biomasy. Białko mikrobiologiczne.
BIOTECHNOLOGIA OGÓLNA 1. Wprowadzenie do biotechnologii. Rys historyczny. Zakres i znaczenie nowoczesnej biotechnologii. Opracowanie procesu biotechnologicznego. 7. Produkcja biomasy. Białko mikrobiologiczne.
CHEMIA klasa 3 Wymagania programowe na poszczególne oceny do Programu nauczania chemii w gimnazjum. Chemia Nowej Ery.
 CHEMIA klasa 3 Wymagania programowe na poszczególne oceny do Programu nauczania chemii w gimnazjum. Chemia Nowej Ery. Dział - Węgiel i jego związki. określa, czym zajmuje się chemia organiczna definiuje
CHEMIA klasa 3 Wymagania programowe na poszczególne oceny do Programu nauczania chemii w gimnazjum. Chemia Nowej Ery. Dział - Węgiel i jego związki. określa, czym zajmuje się chemia organiczna definiuje
PL 198188 B1. Instytut Chemii Przemysłowej im.prof.ignacego Mościckiego,Warszawa,PL 03.04.2006 BUP 07/06
 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 198188 (13) B1 (21) Numer zgłoszenia: 370289 (51) Int.Cl. C01B 33/00 (2006.01) C01B 33/18 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22)
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 198188 (13) B1 (21) Numer zgłoszenia: 370289 (51) Int.Cl. C01B 33/00 (2006.01) C01B 33/18 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22)
PLAN STUDIÓW NR VI. STUDIA PIERWSZEGO STOPNIA (3,5-letnie inżynierskie)
") Pozycja WYDZIAŁ TECHNOLOGII I INŻYNIERII CHEMICZNEJ TECHNOLOGIA PROCESÓW CHEMICZNYCH 2. BIOTECHNOLOGIA PRZEMYSŁOWA 3. ANALITYKA CHEMICZNA I SPOŻYWCZA 4. CHEMIA I TECHNOLOGIA KOSMETYKÓW w tym I II V godzin
Pozycja WYDZIAŁ TECHNOLOGII I INŻYNIERII CHEMICZNEJ TECHNOLOGIA PROCESÓW CHEMICZNYCH 2. BIOTECHNOLOGIA PRZEMYSŁOWA 3. ANALITYKA CHEMICZNA I SPOŻYWCZA 4. CHEMIA I TECHNOLOGIA KOSMETYKÓW w tym I II V godzin
BIOPOLIMERY. Rodzaj zajęć: Grupa: WIMiC I-III r. Termin: poniedziałek Sala: Prowadzący: KONSULTACJE. POK. 106a A3. seminarium 105 A3/A4
 BIOPOLIMERY Rodzaj zajęć: seminarium Grupa: WIMiC I-III r. Termin: poniedziałek 15.00-16.30 Sala: Prowadzący: 105 A3/A4 dr hab. inż. Jadwiga Laska KONSULTACJE CZWARTEK 11.00-12.00 POK. 106a A3 Kontakt
BIOPOLIMERY Rodzaj zajęć: seminarium Grupa: WIMiC I-III r. Termin: poniedziałek 15.00-16.30 Sala: Prowadzący: 105 A3/A4 dr hab. inż. Jadwiga Laska KONSULTACJE CZWARTEK 11.00-12.00 POK. 106a A3 Kontakt
CF 3. Praca ma charakter eksperymentalny, powstałe produkty będą analizowane głównie metodami NMR (1D, 2D).
.") Tematy prac magisterskich 2017/2018 Prof. dr hab. Henryk Koroniak Zakład Syntezy i Struktury Związków rganicznych Zespół Dydaktyczny Chemii rganicznej i Bioorganicznej 1. Synteza fosfonianowych pochodnych
Tematy prac magisterskich 2017/2018 Prof. dr hab. Henryk Koroniak Zakład Syntezy i Struktury Związków rganicznych Zespół Dydaktyczny Chemii rganicznej i Bioorganicznej 1. Synteza fosfonianowych pochodnych
Wykład 6. Anna Ptaszek. 8 września Katedra Inżynierii i Aparatury Przemysłu Spożywczego. Fizykochemia biopolimerów - wykład 6.
 Wykład 6 Katedra Inżynierii i Aparatury Przemysłu Spożywczego 8 września 2016 1 / 27 Konformacje łańcuchów Budowa amylozy i amylopektyny http://polysac3db.cermav.cnrs.fr/home.html 2 / 27 Konformacje łańcuchów
Wykład 6 Katedra Inżynierii i Aparatury Przemysłu Spożywczego 8 września 2016 1 / 27 Konformacje łańcuchów Budowa amylozy i amylopektyny http://polysac3db.cermav.cnrs.fr/home.html 2 / 27 Konformacje łańcuchów
Roman Marecik, Paweł Cyplik
 PROGRAM STRATEGICZNY ZAAWANSOWANE TECHNOLOGIE POZYSKIWANIA ENERGII ZADANIE NR 4 Opracowanie zintegrowanych technologii wytwarzania paliw i energii z biomasy, odpadów rolniczych i innych Roman Marecik,
PROGRAM STRATEGICZNY ZAAWANSOWANE TECHNOLOGIE POZYSKIWANIA ENERGII ZADANIE NR 4 Opracowanie zintegrowanych technologii wytwarzania paliw i energii z biomasy, odpadów rolniczych i innych Roman Marecik,
1. SACHARYDY W ŻYWNOŚCI - BUDOWA I PRZEKSZTAŁCENIA
 Chemia żywności : praca zbiorowa. T. 2, Sacharydy, lipidy i białka / pod red. Zdzisława E. Sikorskiego ; aut. Bronisław Drozdowski [et al.]. wyd. 6, dodr. 2. Warszawa, 2014 Spis treści PRZEDMOWA 11 1.
Chemia żywności : praca zbiorowa. T. 2, Sacharydy, lipidy i białka / pod red. Zdzisława E. Sikorskiego ; aut. Bronisław Drozdowski [et al.]. wyd. 6, dodr. 2. Warszawa, 2014 Spis treści PRZEDMOWA 11 1.
BIOCHEMICZNE ZAPOTRZEBOWANIE TLENU
 BIOCHEMICZNE ZAPOTRZEBOWANIE TLENU W procesach samooczyszczania wód zanieczyszczonych związkami organicznymi zachodzą procesy utleniania materii organicznej przy współudziale mikroorganizmów tlenowych.
BIOCHEMICZNE ZAPOTRZEBOWANIE TLENU W procesach samooczyszczania wód zanieczyszczonych związkami organicznymi zachodzą procesy utleniania materii organicznej przy współudziale mikroorganizmów tlenowych.
Wpływ ph i temperatury na aktywność enzymów na przykładzie α-amylazy [EC ]
![Wpływ ph i temperatury na aktywność enzymów na przykładzie α-amylazy [EC ]](/thumbs/66/56278117.jpg "Wpływ ph i temperatury na aktywność enzymów na przykładzie α-amylazy [EC ]") Wpływ ph i temperatury na aktywność enzymów na przykładzie α-amylazy [EC 3.2.1.1.] Szybkość katalizowanej przez enzym przemiany danego substratu w określony produkt jest ściśle uzależniona od stężenia
Wpływ ph i temperatury na aktywność enzymów na przykładzie α-amylazy [EC 3.2.1.1.] Szybkość katalizowanej przez enzym przemiany danego substratu w określony produkt jest ściśle uzależniona od stężenia
Polisacharydy skrobia i celuloza
 Polisacharydy skrobia i celuloza 1. Cele lekcji a) Wiadomości Uczeń zna: podział cukrów, właściwości fizyczne skrobi i celulozy, reakcję charakterystyczną służącą do identyfikacji skrobi. b) Umiejętności
Polisacharydy skrobia i celuloza 1. Cele lekcji a) Wiadomości Uczeń zna: podział cukrów, właściwości fizyczne skrobi i celulozy, reakcję charakterystyczną służącą do identyfikacji skrobi. b) Umiejętności
(Akty o charakterze nieustawodawczym) ROZPORZĄDZENIA
 ROZPORZĄDZENIA") 18.10.2017 PL L 267/1 II (Akty o charakterze nieustawodawczym) ROZPORZĄDZENIA ROZPORZĄDZENIE WYKONAWCZE KOMISJI (UE) 2017/1896 z dnia 17 października 2017 r. dotyczące na stosowanie preparatu endo-1,3(4)-beta-glukanazy
18.10.2017 PL L 267/1 II (Akty o charakterze nieustawodawczym) ROZPORZĄDZENIA ROZPORZĄDZENIE WYKONAWCZE KOMISJI (UE) 2017/1896 z dnia 17 października 2017 r. dotyczące na stosowanie preparatu endo-1,3(4)-beta-glukanazy
TWORZYWA BIODEGRADOWALNE
 TWORZYWA BIODEGRADOWALNE Opracowały: Joanna Grzegorzek kl. III a TE Katarzyna Kołdras kl. III a TE Tradycyjne tworzywa sztuczne to materiały składające się z polimerów syntetycznych. Większość z nich nie
TWORZYWA BIODEGRADOWALNE Opracowały: Joanna Grzegorzek kl. III a TE Katarzyna Kołdras kl. III a TE Tradycyjne tworzywa sztuczne to materiały składające się z polimerów syntetycznych. Większość z nich nie
Węglowodany (Cukry) Część 3. Związki wielofunkcyjne
 Część 3. Związki wielofunkcyjne") Węglowodany (Cukry) Część 3 Związki wielofunkcyjne Glikozydy Monosacharydy Ryboza, Deoksyryboza: - wzory - funkcje biologiczne, pochodne Disacharydy Sacharoza, Celobioza, Maltoza,Laktoza - wzór - właściwości
Węglowodany (Cukry) Część 3 Związki wielofunkcyjne Glikozydy Monosacharydy Ryboza, Deoksyryboza: - wzory - funkcje biologiczne, pochodne Disacharydy Sacharoza, Celobioza, Maltoza,Laktoza - wzór - właściwości
PLAN STUDIÓW NR IV PROFIL OGÓLNOAKADEMICKI POZIOM STUDIÓW: STUDIA DRUGIEGO STOPNIA (1,5-roczne magisterskie) FORMA STUDIÓW:
 FORMA STUDIÓW:") UNIERSYTET TECHNOLOGICZNO-PRZYRODNICZY 1.TECHNOLOGIA PROCESÓ CHEMICZNYCH. BIOTECHNOLOGIA PRZEMYSŁOA 3. ANALITYKA CHEMICZNA I SPOŻYCZA 4. NOOCZESNE TECHNOLOGIE MATERIAŁOE I godzin tygodniowo (semestr II
UNIERSYTET TECHNOLOGICZNO-PRZYRODNICZY 1.TECHNOLOGIA PROCESÓ CHEMICZNYCH. BIOTECHNOLOGIA PRZEMYSŁOA 3. ANALITYKA CHEMICZNA I SPOŻYCZA 4. NOOCZESNE TECHNOLOGIE MATERIAŁOE I godzin tygodniowo (semestr II
WĘGLOWODORY. Uczeń: Przykłady wymagań nadobowiązkowych Uczeń:
 WĘGLOWODORY Wymagania na ocenę dopuszczającą dostateczną dobrą bardzo dobrą pisze wzory sumaryczne, zna nazwy czterech początkowych węglowodorów nasyconych; zna pojęcie: szereg homologiczny; zna ogólny
WĘGLOWODORY Wymagania na ocenę dopuszczającą dostateczną dobrą bardzo dobrą pisze wzory sumaryczne, zna nazwy czterech początkowych węglowodorów nasyconych; zna pojęcie: szereg homologiczny; zna ogólny
Biotechnologia w produkcji piwa. Wykłady Samodzielna Katedra Biotechnologii i Biologii Molekularnej dr Sławomir Wierzba
 Biotechnologia w produkcji piwa Wykłady Samodzielna Katedra Biotechnologii i Biologii Molekularnej dr Sławomir Wierzba Literatura Wolfgang Kunze - Technologia Piwa i Słodu, Piwochmiel, 1999 r. Treść wykładów
Biotechnologia w produkcji piwa Wykłady Samodzielna Katedra Biotechnologii i Biologii Molekularnej dr Sławomir Wierzba Literatura Wolfgang Kunze - Technologia Piwa i Słodu, Piwochmiel, 1999 r. Treść wykładów
Cz. XXVIII - c Węglowodany - cukry - sacharydy: disacharydy i polisacharydy
 Cz. XXVIII - c Węglowodany - cukry - sacharydy: disacharydy i polisacharydy I. Budowa i właściwości disacharydów Wiązanie między monosacharydami powstaje z udziałem dwóch grup hydroksylowych pochodzących
Cz. XXVIII - c Węglowodany - cukry - sacharydy: disacharydy i polisacharydy I. Budowa i właściwości disacharydów Wiązanie między monosacharydami powstaje z udziałem dwóch grup hydroksylowych pochodzących
Dekstran i dekstranaza w przerobie buraków zdegradowanych
 Dekstran i dekstranaza w przerobie buraków zdegradowanych Aneta Antczak, Maciej Wojtczak Dekstran Zawartość dekstranu soku surowym zdrowe buraki
Dekstran i dekstranaza w przerobie buraków zdegradowanych Aneta Antczak, Maciej Wojtczak Dekstran Zawartość dekstranu soku surowym zdrowe buraki
KINETYKA INWERSJI SACHAROZY
 Dorota Warmińska, Maciej Śmiechowski Katedra Chemii Fizycznej, Wydział Chemiczny, Politechnika Gdańska KINETYKA INWERSJI SACHAROZY Wstęp teoretyczny Kataliza kwasowo-zasadowa Kataliza kwasowo-zasadowa
Dorota Warmińska, Maciej Śmiechowski Katedra Chemii Fizycznej, Wydział Chemiczny, Politechnika Gdańska KINETYKA INWERSJI SACHAROZY Wstęp teoretyczny Kataliza kwasowo-zasadowa Kataliza kwasowo-zasadowa
Nauczycielski plan dydaktyczny z chemii klasa: 1 LO, I ZS, 2 TA, 2 TŻ1, 2 TŻ2, 2 TŻR, 2 TI,2 TE1, 2 TE2, zakres podstawowy
 Nauczycielski plan dydaktyczny z chemii klasa: 1 LO, I ZS, 2 TA, 2 TŻ1, 2 TŻ2, 2 TŻR, 2 TI,2 TE1, 2 TE2, zakres podstawowy Temat lekcji Treści nauczania 1. Zapoznanie z przedmiotowym systemem oceniania
Nauczycielski plan dydaktyczny z chemii klasa: 1 LO, I ZS, 2 TA, 2 TŻ1, 2 TŻ2, 2 TŻR, 2 TI,2 TE1, 2 TE2, zakres podstawowy Temat lekcji Treści nauczania 1. Zapoznanie z przedmiotowym systemem oceniania
Adsorpcja wybranych jonów metali ciężkich na biowęglu pochodzącym z komunalnych osadów ściekowych
 Adsorpcja wybranych jonów metali ciężkich na biowęglu pochodzącym z komunalnych osadów ściekowych mgr Ewelina Ślęzak Opiekun pomocniczy: dr Joanna Poluszyńska Opiekun: prof. dr hab. inż. Piotr Wieczorek
Adsorpcja wybranych jonów metali ciężkich na biowęglu pochodzącym z komunalnych osadów ściekowych mgr Ewelina Ślęzak Opiekun pomocniczy: dr Joanna Poluszyńska Opiekun: prof. dr hab. inż. Piotr Wieczorek
Funkcjonalne nano- i mikrocząstki dla zastosowań w biologii, medycynie i analityce
 Funkcjonalne nano- i mikrocząstki dla zastosowań w biologii, medycynie i analityce dr Magdalena Oćwieja (ncocwiej@cyf-kr.edu.pl) Prace prowadzone w ramach projektu Funkcjonalne nano i mikrocząstki synteza
Funkcjonalne nano- i mikrocząstki dla zastosowań w biologii, medycynie i analityce dr Magdalena Oćwieja (ncocwiej@cyf-kr.edu.pl) Prace prowadzone w ramach projektu Funkcjonalne nano i mikrocząstki synteza
ZAGADNIENIA EGZAMINACYJNE (od roku ak. 2014/2015)
") (od roku ak. 2014/2015) A. Zagadnienia z zakresu Odpady biodegradowalne, przemysłowe i niebezpieczne: 1. Omówić podział niebezpiecznych odpadów szpitalnych (zakaźnych i specjalnych). 2. Omów wymagane warunki
(od roku ak. 2014/2015) A. Zagadnienia z zakresu Odpady biodegradowalne, przemysłowe i niebezpieczne: 1. Omówić podział niebezpiecznych odpadów szpitalnych (zakaźnych i specjalnych). 2. Omów wymagane warunki
Bliskie spotkania z biologią METABOLIZM. dr hab. Joanna Moraczewska, prof. UKW. Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki
 Bliskie spotkania z biologią METABOLIZM dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki Metabolizm całokształt przemian biochemicznych i towarzyszących
Bliskie spotkania z biologią METABOLIZM dr hab. Joanna Moraczewska, prof. UKW Instytut Biologii Eksperymetalnej, Zakład Biochemii i Biologii Komórki Metabolizm całokształt przemian biochemicznych i towarzyszących
Węgiel aktywny - Elbar Katowice - Oddział Carbon. Węgle aktywne ziarniste produkowane są z węgla drzewnego w procesie aktywacji parą wodną.
 Węgle aktywne - Węgle aktywne do uzdatniania wody i oczyszczania ściekãłw: - {jgbox linktext:=[węgiel aktywny ziarnisty 1-4,4-8 mm ]} Węgiel aktywny ziarnisty 1-4,4-8 mm Węgle aktywne ziarniste produkowane
Węgle aktywne - Węgle aktywne do uzdatniania wody i oczyszczania ściekãłw: - {jgbox linktext:=[węgiel aktywny ziarnisty 1-4,4-8 mm ]} Węgiel aktywny ziarnisty 1-4,4-8 mm Węgle aktywne ziarniste produkowane
Substancje o Znaczeniu Biologicznym
 Substancje o Znaczeniu Biologicznym Tłuszcze Jadalne są to tłuszcze, które może spożywać człowiek. Stanowią ważny, wysokoenergetyczny składnik diety. Z chemicznego punktu widzenia głównym składnikiem tłuszczów
Substancje o Znaczeniu Biologicznym Tłuszcze Jadalne są to tłuszcze, które może spożywać człowiek. Stanowią ważny, wysokoenergetyczny składnik diety. Z chemicznego punktu widzenia głównym składnikiem tłuszczów
Ocenę niedostateczną otrzymuje uczeń, który: Ocenę dopuszczającą otrzymuje uczeń, który: Ocenę dostateczną otrzymuje uczeń, który:
 Kryteria oceniania z chemii dla klasy 3A i 3B Gimnazjum w Borui Kościelnej Rok szkolny: 2015/2016 Semestr: pierwszy Opracowała: mgr Krystyna Milkowska, mgr inż. Malwina Beyga Ocenę niedostateczną otrzymuje
Kryteria oceniania z chemii dla klasy 3A i 3B Gimnazjum w Borui Kościelnej Rok szkolny: 2015/2016 Semestr: pierwszy Opracowała: mgr Krystyna Milkowska, mgr inż. Malwina Beyga Ocenę niedostateczną otrzymuje
Wykład 3. Termodynamika i kinetyka procesowa - wykład 2. Anna Ptaszek. 24 kwietnia Katedra Inżynierii i Aparatury Przemysłu Spożywczego
 Wykład 3 wykład 2 Katedra Inżynierii i Aparatury Przemysłu Spożywczego 24 kwietnia 2018 1 / 1 Konformacje łańcuchów Budowa amylozy i amylopektyny http://polysac3db.cermav.cnrs.fr/home.html 2 / 1 Konformacje
Wykład 3 wykład 2 Katedra Inżynierii i Aparatury Przemysłu Spożywczego 24 kwietnia 2018 1 / 1 Konformacje łańcuchów Budowa amylozy i amylopektyny http://polysac3db.cermav.cnrs.fr/home.html 2 / 1 Konformacje
Mikrokapsułki CS. Prof. dr hab. Stanisław Ignatowicz Konsultacje Entomologiczne Warszawa
 Mikrokapsułki CS Prof. dr hab. Stanisław Ignatowicz Konsultacje Entomologiczne Warszawa Kapsułkowanie 2 Kapsułkowanie jest techniką, za pomocą której jeden materiał lub mieszanina materiałów jest powlekana
Mikrokapsułki CS Prof. dr hab. Stanisław Ignatowicz Konsultacje Entomologiczne Warszawa Kapsułkowanie 2 Kapsułkowanie jest techniką, za pomocą której jeden materiał lub mieszanina materiałów jest powlekana
Politechnika. Łódzka
 Politechnika Łódzka Celuloza bakteryjna Alicja Krystynowicz, Wojciech Czaja, Stanisław Bielecki Instytut Biochemii Technicznej Politechnika Łódzka ul. Stefanowskiego 4/10 90-924 Łódź, stanb@ck-sg.p.lodz.pl
Politechnika Łódzka Celuloza bakteryjna Alicja Krystynowicz, Wojciech Czaja, Stanisław Bielecki Instytut Biochemii Technicznej Politechnika Łódzka ul. Stefanowskiego 4/10 90-924 Łódź, stanb@ck-sg.p.lodz.pl
Sposób wytwarzania żywic epoksydowych na bazie sacharydów i ich pochodnych acetalowych
 Sposób wytwarzania żywic epoksydowych na bazie sacharydów i ich pochodnych acetalowych Opis rozwiązania: Synteza opracowana jest dla związków zawierających ugrupowanie hydroksylowe, w szczególności dla
Sposób wytwarzania żywic epoksydowych na bazie sacharydów i ich pochodnych acetalowych Opis rozwiązania: Synteza opracowana jest dla związków zawierających ugrupowanie hydroksylowe, w szczególności dla
Cukry. C x H 2y O y lub C x (H 2 O) y
 y") Cukry Cukry organiczne związki chemiczne składające się z atomów węgla oraz wodoru i tlenu, zazwyczaj w stosunku H:O = 2:1. Zawierają liczne grupy hydroksylowe, karbonylowe a czasami mostki półacetalowe.
Cukry Cukry organiczne związki chemiczne składające się z atomów węgla oraz wodoru i tlenu, zazwyczaj w stosunku H:O = 2:1. Zawierają liczne grupy hydroksylowe, karbonylowe a czasami mostki półacetalowe.
Projektowanie Procesów Technologicznych
 V semestr, I stopień, kierunek: Biogospodarka Projektowanie Procesów Technologicznych Laboratorium 30 h Kierownik przedmiotu: dr inż. Paweł Ruśkowski Prowadzący: mgr inż. Monika Budnicka, mgr inż. Dominika
V semestr, I stopień, kierunek: Biogospodarka Projektowanie Procesów Technologicznych Laboratorium 30 h Kierownik przedmiotu: dr inż. Paweł Ruśkowski Prowadzący: mgr inż. Monika Budnicka, mgr inż. Dominika
Dział 9. Węglowodory. Wymagania na ocenę. dopuszczającą dostateczną dobrą bardzo dobrą. Przykłady wymagań nadobowiązkowych
 Dział 9. Węglowodory rozumie pojęcia: chemia nieorganiczna, chemia organiczna; wie, w jakich postaciach występuje węgiel w przyrodzie; pisze wzory sumaryczne, zna nazwy czterech początkowych węglowodorów
Dział 9. Węglowodory rozumie pojęcia: chemia nieorganiczna, chemia organiczna; wie, w jakich postaciach występuje węgiel w przyrodzie; pisze wzory sumaryczne, zna nazwy czterech początkowych węglowodorów
PLAN STUDIÓW NR V PROFIL OGÓLNOAKADEMICKI POZIOM STUDIÓW: STUDIA DRUGIEGO STOPNIA (1,5-roczne magisterskie) FORMA STUDIÓW:
 FORMA STUDIÓW:") UNIERSYTET TECHNOLOGICZNO-PRZYRODNICZY 1.TECHNOLOGIA PROCESÓ CHEMICZNYCH 2. BIOTECHNOLOGIA PRZEMYSŁOA 3. ANALITYKA CHEMICZNA I SPOŻYCZA 4. NOOCZESNE TECHNOLOGIE MATERIAŁOE I godzin tygodniowo (semestr
UNIERSYTET TECHNOLOGICZNO-PRZYRODNICZY 1.TECHNOLOGIA PROCESÓ CHEMICZNYCH 2. BIOTECHNOLOGIA PRZEMYSŁOA 3. ANALITYKA CHEMICZNA I SPOŻYCZA 4. NOOCZESNE TECHNOLOGIE MATERIAŁOE I godzin tygodniowo (semestr
Wydziału Biotechnologii i Nauk o Żywności
 Pracownie i laboratoria dydaktyczno-badawcze Wydziału Biotechnologii i Nauk o Żywności rozmieszczone są w czterech instytutach biorących udział w realizacji w/w zadań: Instytut Podstaw Chemii Żywności
Pracownie i laboratoria dydaktyczno-badawcze Wydziału Biotechnologii i Nauk o Żywności rozmieszczone są w czterech instytutach biorących udział w realizacji w/w zadań: Instytut Podstaw Chemii Żywności
Biotechnologia jest dyscypliną nauk technicznych, która wykorzystuje procesy biologiczne na skalę przemysłową. Inaczej są to wszelkie działania na
 Biotechnologia jest dyscypliną nauk technicznych, która wykorzystuje procesy biologiczne na skalę przemysłową. Inaczej są to wszelkie działania na żywych organizmach prowadzące do uzyskania konkretnych
Biotechnologia jest dyscypliną nauk technicznych, która wykorzystuje procesy biologiczne na skalę przemysłową. Inaczej są to wszelkie działania na żywych organizmach prowadzące do uzyskania konkretnych
Walory buraków cukrowych, jako surowca
 Walory buraków cukrowych, jako surowca przemysłowego Stanisław Wawro Radosław Gruska, Halina Kalinowska Politechnika Łódzka Wydział Biotechnologii i Nauk o Żywności Instytut Chemicznej Technologii Żywności
Walory buraków cukrowych, jako surowca przemysłowego Stanisław Wawro Radosław Gruska, Halina Kalinowska Politechnika Łódzka Wydział Biotechnologii i Nauk o Żywności Instytut Chemicznej Technologii Żywności
etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
 Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
Właściwości kinetyczne fosfatazy kwaśnej z ziemniaka
 Właściwości kinetyczne fosfatazy kwaśnej z ziemniaka Celem ćwiczenia jest zapoznanie się metodyką wyznaczania szybkości reakcji Vmax oraz stałej Michaelisa Menten dla fosfatazy kwaśnej z ziemniaka WPROWADZENIE
Właściwości kinetyczne fosfatazy kwaśnej z ziemniaka Celem ćwiczenia jest zapoznanie się metodyką wyznaczania szybkości reakcji Vmax oraz stałej Michaelisa Menten dla fosfatazy kwaśnej z ziemniaka WPROWADZENIE
(Tekst mający znaczenie dla EOG) (10) Przedłożono nowe dane wspierające wniosek. (11) Ocena pokazuje, że warunki ustanowione w art.
 (10) Przedłożono nowe dane wspierające wniosek. (11) Ocena pokazuje, że warunki ustanowione w art.") 3.3.2005 Dziennik Urzędowy Unii Europejskiej L 57/3 ROZPORZĄDZENIE KOMISJI (WE) NR 358/2005 z dnia 2 marca 2005 r. dotyczące bezterminowego zezwolenia na niektóre dodatki i zezwolenia na nowe wykorzystanie
3.3.2005 Dziennik Urzędowy Unii Europejskiej L 57/3 ROZPORZĄDZENIE KOMISJI (WE) NR 358/2005 z dnia 2 marca 2005 r. dotyczące bezterminowego zezwolenia na niektóre dodatki i zezwolenia na nowe wykorzystanie