(19)PL (12) OPIS PATENTOWY PL B1 (11) (13) B1 C12N 15/63
|
|
|
- Witold Kowalczyk
- 8 lat temu
- Przeglądów:
Transkrypt
1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19)PL (11) (13) B1 (21) Numer zgłoszenia: (51) IntCl6: C12N 15/63 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: (54) Konstrukty wektora ekspresyjnego do ekspresji antysensownego RNA (73) Uprawniony z patentu: BAYER AKTIENGESELLSCHAFT, Leverkusen, DE (43) Zgłoszenie ogłoszono: BUP 18/94 (45) O udzieleniu patentu ogłoszono: WUP 11/98 (72) Twórcy wynalazku: Axel Kretschmer, Bergisch Gladbach, DE Horst-Peter Antonicek, Bergisch Gladbach, DE Jörg Baumgarten, Wuppertal, DE Antonius Loebberding, Wuppertal, DE Burkhard Mielke, Leverkusen, DE Wolfgang Springer, Wuppertal, DE Udo Stropp, Wuppertal, DE Mark-Michael D. Struck, Bern, DE Lothar Biesert, Farnkfurt, DE Helga Rübsamen-Waigmann, Bad Soden, DE Hary Suhartono, Frankfurt, DE Thomas-Peter Hausner, Leverkusen, DE (74) Pełnomocnik: Kossowska Janina, PATPOL Spółka z 0.0. PL B1 (57) 1. Konstrukty wektora ekspresyjnego do ekspresji antysensownego RNA, znamienne tym, że zawierają a) sekwencje silnego konstytutywnego promotora wybranego spośród promotora metalotioneiny i promotora CMV albo sekwencje hybrydowego promotora obejmującego wymienione silne promotory lub jeden z wymienionych silnych promotorów i promotor HIV-1 LTR oraz b)wirusowy DNA w orientacji antysensownej. FIG. 1
 Twórcy wynalazku: Axel Kretschmer, Bergisch Gladbach, DE Horst-Peter Antonicek, Bergisch Gladbach, DE Jörg Baumgarten, Wuppertal, DE Antonius Loebberding, Wuppertal, DE Burkhard")
2 Konstrukty wektora ekspresyjnego do ekspresji antysensownego RNA Zastrzeżenia patentowe 1. Konstrukty wektora ekspresyjnego do ekspresji antysensownego RNA, znamienne tym, że zawierają a) sekwencje silnego konstytutywnego promotora wybranego spośród promotora metalotioneiny i promotora CMV albo sekwencje hybrydowego promotora obejmującego wymienione silne promotory lub jeden z wymienionych silnych promotorów i promotor HIV-1 LTR oraz b)wirusowy DNA w orientacji antysensownej. 2. Konstrukty według zastrz. 1, znamienne tym, że zawierają subgenomowe sekwencje genowe patogennego wirusa w orientacji 3' > 5' (orientacja antysensowna), pod kontrolą promotora ekspresji antysensownego RNA, którego sekwencja zasad nukleinowych jest komplementarna do mrna patogennego wirusa. 3. Konstrukty według zastrz. 1, znamienne tym, że zawierają prowirusowy, subgenomowy DNA w orientacji antysensownej z retrowirusów takich jak HIV-1 albo HTV-2, onkowirusów. takich jak HTLY-I, HTLV-II, albo onkogenów takich jak abl, erb A, erb B, fms, fos, myb, myc, ras, sis, src. 4. Konstrukty według zastrz. 1, znamienne tym, że zawierają sekwencje promotora lub promotora hybrydowego, oraz prowirusowy DNA HTV-1 w orientacji antysensownej nukleotydów , albo , albo , albo , albo , albo > 4648, albo > 1635, albo , albo , albo , albo , albo Konstrukty według zastrz. 1, znamienne tym, że zawierają subgenowe, prowirusowe sekwencje genowe albo sekwencje onkogenowe w orientacji 3' >5', dla ekspresji krótszych transkryptów antysensownego RNA. 6. Konstrukty według zastrz. 5, znam ienne tym, że zawierają prowirusowe sekwencje genowe z HIV-1, nukleotydów 605 > 155, albo 670 > 440, albo 825 > 535, albo , albo > 5020, albo , albo > 8300, albo > Konstrukty według zastrz. 1, znamienne tym, że zawierają dwie, trzy lub więcej sekwencji subgenomowych z jednego wirusa albo z kilku wirusów. * * * Przedmiotem wynalazku są konstrukty wektora ekspresyjnego do ekspresji antysensownego RNA. Wektory ekspresyjne według wynalazku służą do wytworzenia opornych na HI V komórek ludzkich do celów terapeutycznych. Wynalazek dotyczy celowanego blokowania informacji genetycznej i replikacji wirusa (na przykład HIV) w wirusowym informacyjnym kwasie rybonukleinowym (mrna) w transfekowanych komórkach krwiotwórczych, przy pomocy specjalnych systemów ekspresyjnych, które wywołują ekspresję komplementarnego "antysensownego" RNA. W szczególności wynalazek dotyczy wektorów ekspresyjnych, kodujących sekwencje antysensownego RNA, które w przeciwieństwie do stanu techniki, dają całkowite hamowanie replikacji HIV-1 i wyraźnie hamują replikację HIV-2 w ludzkich komórkach zmodyfikowanych przy pomocy inżynierii genetycznej. Opisane tu jednostki genetyczne wektorów ekspresyjnych są przeznaczone szczególnie do somatycznej terapii genowej schorzeń HIV/AIDS, ponieważ chronią one komórki krwiotwórcze przed indukowanymi HIV efektami cytopatycznymi. W ten sposób można uniknąć niewydolności immunologicznej u osób zakażonych HIV. Namnażanie się wirusa w zakażonej komórce jest sterowane przez geny genomu wirusowego. W tym procesie z wirusowych genów powstają poprzez transkrypcję i translację, produkty

3 regulatorowe i białka strukturalne do budowy zakaźnych cząstek wirusowych do namnażania się wirusa. Istotnym pośrednikiem w tym procesie jest informacyjny kwas rybonukleinowy (mrna), który przekazuje informację o budowie białek regulatorowych i strukturalnych. Blokowanie informacyjnego kwasu rybonukleinowego w sposób wysoce swoisty, który opiera się całkowicie na swoistości struktury molekularnej wirusowego mrna, a mianowicie na swoistej dla wirusa sekwencji zasad nukleinowych tej cząsteczki można uzyskać za pomocą kwasu rybonukleinowego o komplementarnej sekwencji zasad. Przez swoiste dla sekwencji parowanie zasad nukleinowych, jak to opisano dla kwasu dezoksyrybonukleinowego, pierwotnie jednoniciowy mrna zostaje skompleksowany w RNA dwuniciowy i informacja genetyczna zawarta w sekwencji zasad nukleinowych staje się częściowo niedostępna dla biosyntezy białka. Drugi etap ekspresji genu, translacja, jest tym samym zahamowany. Komplementarny kwas rybonukleinowy określa się jako RNA antysensowny. Mechanizm hamowania ekspresji genu przez wzajemne oddziaływanie antysensownego RNA z docelowym mrna jest złożony i w dużym stopniu nie wyjaśniony. Jednym wyjaśnieniem jest hamowanie cięcia i składania RNA i obróbki RNA w jądrze komórkowym, jednak dokładne badania na ten temat nie są znane. Opisana jest wymiana adenozyna - inozyna w dwuniciowym RNA i sugeruje się, że hamowanie translacji jest następstwem tej wymiany. Hamowanie transportu RNA przez błonę jądrową do miejsca biosyntezy białka oraz maskowanie wzajemnego oddziaływania rybosomów i RNA przez dwuniciowy RNA mogą być dalszą przyczyną hamowania ekspresji genów przez antysensowny RNA. Opisano hamowanie procesów metabolicznych przez antysensowny RNA w naturalnie występujących układach biologicznych, takich jak bakterie i niektóre eukariotyczne organizmy wielokomórkowe (K.M. Takayamai M. Inouye, Critical Reviews in Biochemistry and Molecular Biology 25,155 (1990), jak również w otrzymanych sztucznie przez transfer genów organizmach transgenicznych (J. Mol. i in. FEBS Letters 268 (1990)). Donoszono również o próbach wytworzenia opornych na wirus organizmów roślinnych (M.Cuozzo i in., Biotechnology 6,549 (1988) i zwierzęcych (T. Riiden i E. Golboa, J. Virol. 63, 677 (1989)), po wprowadzeniu genu kodującego antysensowny RNA. Jak podają sami autorzy tych publikacji, nie ma jednak przekonujących, naukowo stwierdzonych, zależności między ekspresją antysensownego RNA i opornością na wirus. W szczególności w przypadku plazmidów, które wprowadza się przez mikroiniekcję, warunkujących ekspresję antysensownego RNA z ekspresją przejściową, działanie przeciwwirusowe jest słabe albo nie daje się zdecydowanie ocenić (K. Rittner i in., Nucleic Acids Research 19, 1421 (1991)). Doniesiono także o zastosowaniu wektorów retrowirusowych, warunkujących ekspresję antysensownego RNA, do wytwarzania komórek opornych na wirus (R. Y-L., To i in. w: Gene Regulation: Biology of antisense RNA and DNA, wyd. R.P. Ericskon i J.G. Izant, Raben Press Ltd., New York 1992). Te retrowirusowe wektory niosą ryzyko wywołania złośliwego wzrostu nowotworowego, ponieważ zawierają sekwencje genów podobnych do onkogenów i istnieją liczne dowody na onkogenezę zależną od retrowirusów. W przeciwieństwie do publikowanych prac na temat antysensownego RNA w regulacji replikacji wirusa, w niniejszym wynalazku opisane są konstrukty ekspresyjne, które odznaczają się szczególnym rodzajem konstrukcji promotora w kombinacji ze specjalnie wybranymi regionami wirusowego DNA w orientacji antysensownej, i przez to wykazują działania przeciwwirusowe, które jakościowo i ilościowo są zaskakujące i daleko wykraczają poza dotychczas znany zakres. Kombinacja składników wektora ekspresyjnego: sekwencjaflankująca 5' DNA, promotor, wstawka/antysensowny DNA i 3' - sekwencje sygnałowe obróbki po raz pierwszy dała w zakażeniach retrowirusowych dającą się pewnie oznaczyć i wyraźnie wyrażoną oporność na wirus w transfekowanych komórkach wykazujących ekspresję antysensownego RNA. Natomiast według stanu techniki, z innymi systemami wektorowymi do endogennej ekspresji antysensownego RNA związane są słabe, tylko nieznaczne efekty przeciwwirusowe, pojawiające się przejściowo (na przykład T. Riiden i E. Gilboa, J. Virol., 63, 677 (1989)), opis patentowy EP , G. Sczakiel i in., Biochem. Biophys. Res. Com. 169, 643 (1990), O.i. Mirosknichenko i in., Gene 84, 83 (1989), T. Shimada i in., Antiviral Chemistry and Chemotherapy 2, (1991), G. Sczakiel i in. J. Virol. 66, 5576 (1992) albo są one obarczone wyżej

4 wspomnianym ryzykiem (transfer genu retrowirusowego), albo są niepraktyczne ponieważ wymagają mikroiniekcji. Wynalazek dotyczy konstruktów wektora ekspresyjnego do ekspresji antysensownego DNA, które zawierają a) sekwencje silnego konstytutywnego promotora wybranego spośród promotora metalotioneiny i promotora CMV albo sekwencje hybrydowego promotora obejmującego wymienione silne promotory lub jeden z wymienionych silnych promotorów i promotor HIV-1 LTR oraz b)wirusowy DNA w orientacji antysensownej. Są to korzystnie takie konstrukty wektora ekspresyjnego, które zawierają subgenomowe sekwencje genowe patogennego wirusa w orientacji 3' > 5' (orientacja antysensowna), pod kontrolą promotora ekspresji antysensownego RNA, którego sekwencja zasad nukleinowych jest komplementarna do mrna patogennego wirusa. Bardzo korzystne są takie konstrukty, które zawierają prowirusowy, subgenomowy DNA w orientacji antysensownej z retrowirusów takich jak HI V-1 albo HI V-2, onkowirusów, takich jak HTLV-I, HTLV-II, albo onkogenów takich jak abl, erb A, erb B, fms, fos, myb, myc, ras, sis, src. Korzystne są konstrukty wektora ekspresyjnego, które zawierają sekwencje promotora lub promotora hybrydowego, oraz prowirusowy DNA HIV-1 w orientacji antysensownej nukleotydów > 474, albo > 474, albo > 474, albo > 681, albo > 4158, albo > 4648, albo > 1635, albo > 3003, albo > 678, albo > 2099, albo >4157, albo > Korzystne są również konstrukty wektora ekspresyjnego, które zawierają subgenowe, prowirusowe sekwencje genowe albo sekwencje onkogenowe w orientacji 3' >5', dla ekspresji krótszych transkryptów antysensownego RNA. Korzystne są również takie konstrukty wektora ekspresyjnego, które zawierają prowirusowe sekwencje genowe z HIV-1, nukleotydów 605 -> 155, albo 670 -> 440, albo 825 -> 535, albo > 5800, albo > 5020, albo > 5040, albo > 8300, albo > Korzystne są konstrukty wektora ekspresyjnego, które zawierają dwie, trzy lub więcej sekwencji subgenomowych z jednego wirusa albo z kilku wirusów. Konstruktem według wynalazku można transfekować komórki. Konstrukty wektorów ekspresyjnych nadają się też do wytwarzania środków leczniczych, w których substancję czynną stanowi jeden lub więcej konstruktów według wynalazku. Stosując opisane tu konstrukty wektora ekspresyjnego w odpowiednich ludzkich komórkach gospodarza wykazano działanie przeciwwirusowe w stosunku do replikacji HIV-1, które zapewniało całkowitą ochronę przed replikacją HIV-1 w komórkach ludzkich przez ponad 60 dni, a prócz tego w sposób istotny hamowało replikację HIV-2. Konstrukty wektora ekspresyjnego według wynalazku wytwarzają w transfekowanych komórkach jako transkrypt informacyjny kwas rybonukleinowy, który jest komplementarny do sekwencji mrna niepożądanych genów wirusowych, na przykład genów GAG, POL, VIF, REV, TAT i VPU ludzkiego wirusa niedoboru odporności, HIV-1 (W.A. Haseltine i in., Scientific American USA, 1988, 34 do 42). Ponadto opisana sekwencja antysensownego RNA jest wystarczająco homologiczna wobec HIV-2 (na przykład izolatu HIV-2 D 194, w: Human retroviruses and AIDS, wyd. Myers i in., Los Alamos National Lab., 1992, oraz EMBL nr depozytu X52223) w zakresie genów GAG, POL i VIF, aby wywierać działanie przeciwwirusowe także na izolaty HIV-2. Konstrukty wektora ekspresyjnego według wynalazku wyróżniają się szczególnym rodzajem uszeregowania wybranych sekwencji promotora i sekwencji promotora hybrydowego (na przykład CMV-IE/metalotioneina I), z subgenowymi fragmentami DNA HIV-1 w orientacji antysensownej (3' > 5'), co daje w wyniku kilku transkryptów, antysensownych RNA, przez konstytutywne lub indukowane cięcie i łączenie RNA. Tak utworzone transkrypty antysensownego RNA wykazują nieoczekiwanie drastyczne albo całkowite hamowanie replikacji HIV-1 w komórkach ludzkich (HUT 78, U 937, na przykład). Stwierdzono także wyraźne hamowanie replikacji HIV-2. HIV-2 wykazuje w opisywanych tu konstruktach wektora ekspresyjnego wystarczającą homologię sekwencji RNA, tak że dochodzi do hamowania replikacji wirusa przez antysensowny RNA. Stosując zatem konstrukty według wynalazku hamuje się replikację retrowirusów, np. HIV-1 i HIV-2, chroniąc w ten sposób komórki krwiotwórcze - a więc ważne komórki ludzkiej komórkowej reakcji odpornościowej (np. limfocyty T,

5 monocyty) - przed szkodliwym i niszczącym komórki namnażaniem się wirusa, przez określony sposób ekspresji działającego przeciwwirusowo antysensownego RNA. Następująca procedura eksperymentalna została przeprowadzona w celu nadania komórkom układu odpornościowego oporności na uszkadzające działanie namnażania się retrowirusów, znane z zakażenia HIV-1 i HIV-2, za pomocą ekspresji przeciwwirusowego antysensownego RNA. Promotory: Skonstruowano wektory ekspresyjne, zawierające promotory, które w komórkach krwiotwórczych powinny zapewniać konstytutywną i wydajną ekspresję, jak na przykład promotor IE wirusa cytomegalii (CMV-EE). Ponadto, skonstruowano także przez fuzję sekwencji kwasów nukleinowych, takie promotory, które przy inwazji wirusa powinny dodatkowo indukować amplifikowaną ekspresję. W szczególności składnikiem tych promotorów hybrydowych jest fragment promotora CMV-IE-metalotioneiny oraz fragment kwasu nukleinowego HI V-1 LTR od nukleotydów 289 do 474 albo 289 do 532 (numeracja nukleotydów: UWGCG GENEMBL Data Bank, File: HIVHXB2CG. VIRAL 25 września 1987), który przyczynia się do amplifikowanej, ewentualnie dającej się transaktywować, dodatkowej aktywności promotora. Fragmenty antysensownego kwasu nukleinowego. Dodatkowo konstrukty wektorów zawierają, pod kontrolą opisanych promotorów, subgenomowe prowirusowe fragmenty DNA w orientacji antysensownej, to znaczy nić (+) DNA ulega transkrypcji w kierunku 3' > 5'. Ze względu na bezpieczeństwo nie stosuje się kompletnego prowirusowego genomowego fragmentu DNA retrowirusa. Stosuje się jednak raczej dłuższe, subgenomowe prowirusowe fragmenty DNA, a nie krótkie, subgenowe prowirusowe fragmenty DNA, głównie ponieważ większe jest prawdopodobieństwo cięcia i składania RNA w dłuższych transkryptach RNA i w ten sposób z prowirusowych fragmentów DNA można oczekiwać większej liczby transkryptów antysensownego RNA, zgodnie z wynalazkiem, co w wymienionych przykładach rzeczywiście stwierdzono. (Rodzaj i liczba transkryptów antysensownego mrna po całkowitej obróbce, których można oczekiwać dla zastosowanych sekwencji HIV w orientacji antysensownej jest nie do przewidzenia, ponieważ nie ma wystarczająco zgodnych sekwencji do składania mrna, które pozwoliłyby na zdecydowane przewidywanie). Wraz ze wzrastającą liczbą transkryptów antysensownego RNA, z przynależnymi im cechami, jak na przykład eksport z jądra albo stabilność w cytoplazmie, wzrasta również prawdopodobieństwo wytworzenia transkryptu antysensownego RNA o silnym działaniu przeciwwirusowym. Wynalazek dotyczy konstruktów wektora ekspresyjnego, które zawierają sfuzowany fragment genu ulegającego transkrypcji do antysensownego RNA pod kontrolą silnego promotora. Sfuzowany fragment genu składa się z subgenomowych fragmentów DNA z dwu lub kilku różnych patogennych wirusów, na przykład HIV-1 i HIV-2. Stosując takie sfuzowane fragmenty genowe, które kodują transkrypt RNA zawierający kolejno kilka regionów sekwencji antysensownego RNA komplementarnych do regionów sekwencji mrna z różnych wirusów, można skonstruować wektory ekspresyjne o szerszym działaniu przeciwwirusowym. Słuszność tej zasady potwierdzają przykłady I, IV, V, X i XII. Na przykład konstrukty wektora ekspresyjnego KTX 11, psxk1 i psxk2 zawierają fragmenty antysensowego DNA z HIV-1 LAV/BRU, które ze swej strony zawierają krótsze odcinki, które są także komplementarne do mrna z HIV-2 D194 (naprzykład nukleotydy albo , albo ). W ten sposób antysensowny RNA z KTX11, psxk1 i psxk2 jest przeciw wirusowy także wobec HIV-2. Można teraz kodującą antysensowny RNA sekwencję tych wektorów dodatkowo poddać fuzji z fragmentami prowirusowego DNA z innych wirusów w orientacji antysensownej (na przykład izolaty HIV-2, inne izolaty HIV-1) tak, że w wyniku segmentowo zoptymalizowanej komplementarności antysensownego RNA do sekwencji innych wirusów, można osiągnąć równocześnie lepsze lub całkowite hamowanie różnych wirusów jednym konstruktem wektora ekspresyjnego.

6 Terminację transkryptów antysensownego RNA uzyskuje się przy pomocy sygnału poliadenylacji (na przykład z SV 40 albo z bydlęcego hormonu wzrostu), a celu wygenerowania wysoce stabilnego mrna. Ponadto wektory zawierają pełny operon dla ekspresji odpowiedniego markera selekcyjnego. W przytoczonych przykładach wykorzystano oporność na antybiotyki TNR 5 neo (neomycyna, genetycyna, G 418, kanamycyna) w celu wyselekcjonowania komórek, które po transfekcji zawierają konstrukt genetyczny dla ekspresji antysensownego DNA. Ten marker selekcyjny okazał się przydatny dla ludzkich komórek krwiotwórczych. Można było zastosować także inne markery selekcyjne, jak na przykład puromycynę albo higromycynę albo metotreksat, w połączeniu z transfekcją genu dla reduktazy dihydrofolianowej. Szczególnie przydatne jako antysensowne sekwencje nukleotydowe są sekwencje z regionu POL/VIF genu HIV, między nukleotydami Zwłaszcza antysensowne sekwencje nukleotydów wykazały wyróżniającą się aktywność przeciwwirusową. Można również stosować inne fragmenty z tego regionu, na przykład > 4648 albo > 4157, albo > 3880, albo > 4158, albo > W celu wykazania działania przeciwwirusowego antysensownego RNA, na przykład przeciw HIV-1 linie komórek ludzkich, które mogą być zakażone HIV-1, transfekowano wyżej wspomnianymi konstruktami wektora ekspresyjnego. Celem było wykazanie na szeregu typów ludzkich komórek krwiotwórczych, że endogenna ekspresja antysensownego RNA prowadzi do zahamowania replikacji wirusa w komórkach, które biorą szczególny udział w komórkowej reakcji immunologicznej. To niszczenie komórek układu odpornościowego w końcowej fazie choroby AIDS pozostaje w związku przyczynowym ze zwiększoną częstością zakażeń oportunistycznych, które wtedy mają często przebieg śmiertelny. Dlatego celem prac z komórkami krwiotwórczymi a najpierw ciągłymi liniami komórkowymi, jak na przykład HUT 78, MOLT 4, U 937, Jurkat, CEM-CM3 (wszystkie według American Type Culture Collection ATCC, Bethesda, Maryland, USA) po transfekcji wektorem, klonowaniu i genetycznym scharakteryzowaniu, w doświadczeniach nad zakażeniem odpowiednimi izolatami HTV-1 i HIV-2 było zbadanie "oporności na wirus" tych transfekowanych linii komórkowych. Wykazana ekspresja działającego przecwiwirusowo antysensownego RNA w tych komórkach wskazuje na użyteczność konstruktów ekspresyjnych w komórkach krwiotwórczych i w końcu także na ich zastosowanie do transformacji komórek tkanki krwiotwórczej szpiku kostnego dla celów terapeutycznych. Włączenie genów do limfocytów krwi obwodowej i komórek szpiku w celach leczniczych zostało opisane w innym miejscu (K.W. Culver i in., Transplant Proc. 23, (1991) i w zasadzie powinno być możliwe do zastosowania w odniesieniu do wymienionych tu konstruktów do ekspresji antysensownego RNA. Przykład I. Konstrukt wektora ekspresyjnego KTX 11. (Jeśli nie podano inaczej, cała numeracja nukleotydów odnosi się poniżej do sekwencji wirusa ludzkiego niedoboru odporności typu 1 (HXB2), WUGCG, GENENBL Data Bank, file: HTVHXB2CG VIRAL, od 25 września 1987). W celu endogennej ekspresji działającego przeciwwirusowo antysensownego RNA po transfekcji komórek, które mogą ulec zakażeniu HI V-1, skonstruowano następujący wektor ekspresji. Mysi promotor metalotioneiny I (1600 par zasad EcoRI Bgl II fragment (N. Glanville i in., Nature 292, (1982)) zligowano w miejscu cięcia Sal I (nukleotydów 5786) regionem 3' prowirusowej sekwencji DNA HIV-1 po ligacji z adaptorowym RNF Bgl Ii-Xho I, uzyskując w efekcie modyfikację miejsc cięcia Bgl II, Xho I i Sal I. Region 5' prowirusowego DNA HIV-1 w miejscu cięcia Bgl II (nukleotyd 474) poddano ligacji z miejscem cięcia Bam HI, do którego przyłączona jest sekwencja sygnału poliadenylacji SV 40 (nukleotydy 2564 do 4713 z delecją sekwencji z UWGCG GenEMBL file SV 40XX). Tę samą sekwencję poliadenylacji z SV 40 zastosowano w kierunku 3' > 5' dla terminacji TRN5NEO (marker selekcyjny neomycyna (fosfotransacetylaza), nukleotydy TRN5NEO BACTERIA, UW GCG GenEMBL). Ekspresja genu oporności na neomycynę jest inicjowana przez wczesny promotor SV 40 (nukleotydy ), UWGCG GenEMBL SV 40XX). W innym przypadku wektor zawiera także pbr 322 DNA (nukleotydy , WUGCG, GenEMBL File PBR 322) pochodzenia bakteryjnego oraz gen oporności na ampi-

7 cylinę, aby można było go użyć jako wektor wahadłowy (Shuttle) dla E. coli i komórek zwierzęcych. Mapa genowa tego wektora KTX 11 jest przedstawiona na fig. 1. Przykład II. Konstrukt wektora ekspresyjnego KTX 12. Wektor ekspresyjny KTX 12 antysensownego RNA HIV-1 jest skonstruowany tak samo jak wektor KTX 11 (przykład I). Zawiera on jednak podlegający indukcji promotor hybrydowy, a nić matrycy DNA HI V-1 rozciąga się w orientacji 3' > 5' poza ten promotor hybrydowy, od nukleotydów 7514 > 474. Dla otrzymania promotora hybrydowego, fragment promotora LTR HIV-1, nukleotydy 289 do 532, poddano fuzji z nukleotydem 368 promotora metalotioneiny (UW GCG File MYSMETI.RO). Mapa restrykcyjna tego wektora KTX 12 jest przedstawiona na fig. 2. Przykład III. Konstrukt wektora ekspresyjnego KTX 15. Wektor ekspresyjny KTX 15 antysensownego RNA HIV-1 jest skonstruowany podobnie jak wektor KTX 12 (przykład II). Jednakże fragment DNA nici matrycy HIV-1 ( w orientacji 3' - 5') jest istotnie skrócony i rozciąga się od nukleotydu 2096 do nukleotydu 474, obejmując tym samym sekwencję GAG, sekwencję lidera, sekwencję U5 oraz część sekwencji R. Dalszą ważną cechą wektora KTX jest delecja sekwencji TAR, z delecją nukleotydów promotora hybrydowego HI V-LTR. W ten sposób powinna nadal być możliwa aktywacja promotora hybrydowego w przypadku zakażenia HIV, ale bez obecności białka TAT HIV-1 nie powinna zachodzić terminacja transkrypcji, kodowanej przez TAR (S.Y. Kao i in., Nature 330, 489 (1987)). Ten promotor hybrydowy odznacza się zatem szczególnie wysoką konstytutywną transkrypcją pod kontrolą sekwencji promotora LTR HI V-1. Mapa restrykcyjna wektora KTX 15 jest przedstawiona na fig. 3. Przykład IV. Wektor ekspresyjny PSXK1 antysensownego RN A HIV -1 przygotowano celem wykazania ekspresji antysensownego RNA pod kontrolą innego silnego promotora warunkującego wydajną i konstytutywną ekspresją, w tym przykładzie promotora wirusa cytomegalii. Fragment genu HIV-1, nukleotydy , wprowadzono w orientacji 3' >5', do miejsca restrykcji Hind III przy nukleotydzie 891 (wektor prc/cmv, numer katalogu V , Invitrogen, San Diego CA 92121, USA). Sygnał poliadenylacji pochodzi z genu bydlęcego hormonu wzrostu (BGH). W innym przypadku wektor jest skonstruowany jako wektor wahadłowy, w ten sposób jak wektor KTX 11 (przykład I) z neomycynowym markerem selekcji i fragmentem DNA pbr322 dla replikacji i selekcji w E.coli. Mapa restrykcyjna wektora psxk 1 jest przedstawiona na fig. 4. Przykład V. Konstrukt wektora ekspresyjnego psxk 2. Wektor ekspresyjny psxk 2 antysensownego RNA HIV-1 różni się od wektora psxk 1 (przykład IV) tym, że posiada sekwencje promotora zawierające nukleotydy promotora CMV (patrz przykład IV) sfuzowane z nukleotydami promotora metalotiotioneiny (UWGCG GenEMBL file MUSMETI.RO), a więc jest promotorem hybrydowym. Ponadto, prowirusową sekwencję DNA nukleotydów wstawiono w orientacji 3' >5' jako matrycę dla ekspresji antysensownego RNA, pod kontrolą promotora hybrydowego CMV/metalotioneina. Mapa restrykcyjna wektora psxk 2 jest przedstawiona na fig. 5. Przykład VI. Konstrukty wektorów ekspresyjnych phtt 1, 4, 6, 10, 12 i 15. Wektory ekspresyjne phtt 1,4, 6,9,10,12 i 15 antysensownego RNA HIV-1 przygotowano w sposób podobny jak wektor psxk 1 (przykład IV, fig. 4). Wszystkie fragmenty genowe HIV-1 wstawione do wektorów phtt klonowano w orientacji 3' > 5' do wektora prc/cmv (Invitrogen, San Diego, CA 92121, nr katalogu V , 1991), pod kontrolą promotora cytomegalowirusa warunkującego konstytutywną ekspresję. Wektory phtt są podane niżej: (a) phtt 1: fragment genu HI V-1 (nukleotydy ) wbudowano do miejsca cięcia Not I (nukleotyd 966) wektora prc/cmv. (b) phtt 4: fragment genu HIV-1 (nukleotydy ) wbudowano do miejsca cięcia Xba I (nukleotyd 985) wektora prc/cmv. (c) phtt 6: fragment genu HIV-1 (nukleotydy ) wbudowano do miejsca cięcia Xba I jak opisano pod (b). (d) phtt 9: fragment genu HIV-1 (nukleotydy ) wbudowano do miejsca cięcia Xba I jak opisano pod (b).
.")
8 (e) phtt 10: fragment genu HIV-1 (nukleotydy ) wbudowano do miejsca cięcia Xba I jak opisano pod (b). (f) phtt 12: fragment genu HIV-1 (nukleotydy ) wbudowano do miejsca cięcia Xba I jak opisano pod (b). (g) phtt 15: fragment genu HI V-1 (nukleotydy ) wbudowano do miejsca cięcia Xba I jak opisano pod (b). Ogólny wykres fragmentów genowych HIV-1, które są wprowadzone w orientacji antysensownej 3' >5' w wektorach z przykładów I do VI jest przedstawiony na fig. 6. Przykład VII. Transfekcja i klonowanie limfocytów ludzkich wektorami dla ekspresji antysensownego RNA. Jako przykłady transfekowanych linii ludzkich komórek krwiotwórczych, które wykazują oporność na wirus w wyniku endogennej ekspresji antysensownego RNA przeciw HIV-1, wybrano rosnącą w zawiesinie monocytopodobną linię komórkową U 937 oraz tak samo rosnącą linię T limfocytów HUT 78. Obie linie komórkowe otrzymano z American Type Culture Collection (ATCC), Parklawn Drive, Rockville, MD , USA. Linie komórkowe hodowano w pożywce zalecanej przez ATCC. Transfekcję wektorem przeprowadzono różnymi sposobami, przy czym najlepsze wyniki dała elektroporacja: 1. Metoda z fosforanem wapnia: Modyfikacja metody Wiglera i in., 1977, Cell 11, 223. Transfekcję przeprowadza się precypitatami wapniowymi DNA. Różne ilości DNA od 0,05 do 20 μg rozpuszczono w 0,25 ml w 250 mm CaCl2 i wolno wkroplono do 0,25 ml buforu HEPES X 2. Transfekcję otrzymano dodając całą mieszaninę precypitatu do 10 ml hodowli komórkowej. 15 godzin później usunięto podłoże przed odessanie. Komórki poddano szokowi DMSO. Na 10 ml hodowli dodano 3 ml 25% roztworu DMSO w jałowym PBS. Po 4 minutach odessano płyn a komórki przemyto 5 ml PBS. Następnie komórki zawieszono w 10 ml pożywki DMEM. 2. DEAE dekstran Dzień przed transfekcją sporządzono zawiesinę komórek o gęstości 3-4 x 105 na ml pożywki wzrostowej. 2-4 godziny przed transfekcją komórki odwirowano i zawieszono w świeżej porcji pożywki. 0,5 do 20 μg DNA zmieszano z 1070 μl PBD i dodano 120 μg roztworu DEAE (roztwór transfekcyjny). Przed transfekcją odwirowano 40 ml zawiesiny komórek, zawieszono je w 10 ml PBS i ponownie wirowano. Osad komórek zawieszono w roztworze pbs-deae dekstran i inkubowano 30 minut. Następnie komórki poddano szokowi przez dodanie 0,8 ml zimnego roztworu DMSO (25% TBS). Po 3 minutach dodano 10 ml TBS i wirowano. Komórki przemyto pożywką wzrostową RPMI i zawieszono w 40 ml tej pożywki 3. Fuzja (metoda Sanari-Goldin) Rosnące wykładniczo komórki (1 x 107/ml) odwirowano i raz przemyto pożywką RPMI. Do osadu komórek dodano 2 ml zawiesiny protoplastów E.coli (2 x 103/ml) w 10% sacharozie w 10 mm MgCl2. Protoplasty z transfekującym plazmidem wytworzono według Sanari-Goldin i in., Po 8 minutach inkubacji wirowano i ostrożnie w ciągu 2 minut dodano roztwór PEG (glikol polietylenowy 4000, 45% roztwór w RPMI). Po 1 minucie inkubacji w ciągu 7 minut dodano 10 ml pożywki RPMI bez FCS. Komórki odwirowano, przemyto pożywką RPMI i zawieszono w 10 ml. 4. Elektroporacja Elektroporację (EP) przeprowadzono w aparacie Gene Pulser and Capacitance Extender, firmy Bio Rad. Komórki w wykładniczej fazie wzrostu doprowadzono do gęstości 1 x 107/ml, raz przemyto w PBS i zawieszono w pożywce RPMI bez FCS z 10 mm dekstrozą i 0,1 mm ditiotreitolem. 0,4 ml zawiesiny komórek umieszczono w kuwecie do EP i dodano μg DNA. DNA rozpuszczono do stężenia 1 μg na 1 μl TE. Warunki elektroporacji wahały się, jak następuje: Pojemność: (μf; napięcie : V; odległość elektrod 0,2/0,4 cm; oporność: omów.

9 Przed i po elektroporacji komórki inkubowano 10 minut w temperaturze pokojowej, po czym zawieszono w 10 ml pożywki wzrostowej + PCS. Selekcja transfektantów: Selekcję transfektantów przeprowadzono w każdym przypadku na podstawie oporności przeciw G Selekcja na podłożu półpłynnym Transfekowane komórki najpierw inkubowano w pożywce wzrostowej zawierającej G 418 tak długo, aż nie stwierdzono dalszego wzrostu. Następnie wysiewano na podłoże półpłynne. Do zestalenia środowiska służył agar (Gibco) albo metyloceluloza. Wstępnie wyselekcjonowane komórki wysiewano na płytkę do hodowli tkanek (10 cm) i dodano 10 ml pożywki selekcyjnej (RPMI plus 1 mg/ml G 418) i 2 ml 2% upłynnionego roztworu agaru. Po ochłodzeniu agaru, inkubowano w cieplarce w 37 C. W wariancie tego doświadczenia agar zastąpiono metylocelulozą. 2. Selekcja i klonowanie w komorze do hodowli komórek Transwell TM, firmy Costar 48 godzin po transfekcji komórki wysiewano na 96-studzienkową płytkę mikrotitracyjną, przy gęstości 1 x 105 komórek na studzienkę 0,2 ml. Dodano G 418 do stężenia 0,7 mg/ml dla komórek U 937 i 1,0 mg/ml dla HUT 78. Gdy tylko pożywka została zużyta dodawano 1/2 objętości studzienki i pożywki; w przypadku U 937 dodawano tylko 0,5 mg/ml G 418. Inkubację prowadzono tak długo, aż w poszczególnych studzienkach narosły komórki oporne. Te klony mieszane uzupełniano do większej objętości i następnie klonowano. Stosowano do tego celu specjalne komory hodowlane firmy Costar (Transwell TM, nr katalogu 3425, 205, Broadway, Cambridge, MA 02139, USA). Komórki w zawiesinie o niewielkiej gęstości (50 do 200 komórek na 6 ml) wysiewano do miękkiego agaru zawierającego G 418 (patrz selekcja w środowisku półpłynnym), a 6 ml umieszczano w komorze centralnej (przedział górny). Komora jest oddzielona błoną od bardzo gęsto rosnącej płynnej hodowli zaopatrującej U 937 względnie HUT 78 ( komórek/ml) w dolnym przedziale. Płynną pożywkę w miarę zużycia częściowo wymieniano, aby umożliwić dalszy wzrost komórek. Gdy w miękkim agarze pojawiły się kolonie oporne na G 418, izolowano je pipetą Pasteurowską i namnażano. W ten sposób z użyciem wektorów KTX 11, KTX 12, KTX 15, psxk 1 i psxk 2 sklonowano po około 10 niezależnych, transfekowanych linii komórek U 937 i HUT 78, które zbadano dalej dla wykazania ekspresji antysensownego RNA i oporności na HIV-1. Ponadto, z użyciem wektorów phtt1, phtt4, phtt9 i phtt10 generowano linie komórek HUT 78 i sprawdzono je w próbie zakażenia HIV. Przykład VIII. Transformacja komórek szpiku wektorem ekspresji KTX 15. Jako przykład, podstawowe postępowanie do badania transformacji pierwotnych komórek szpiku na modelu myszy przebiegało następująco: Pierwotne komórki szpiku otrzymano z kości udowych myszy C57 w wieku od sześciu do dziesięciu tygodni. Komórki pobrano w pożywce McCoy (firmy Flow) i wysiano na szalki Petriego, po zmieszaniu z interleukiną 3 i GM-CSF (czynnik stymulujący kolonie granulocytów-monocytów - Granulocyte-monocyte colony stimulating factor, firmy Genzyme) w stężeniu 2,5 μg/ml, oraz z agarem (Bacto Agar firmy Difco, końcowe stężenie 0,3%), w stężeniu 1 x 105 komórek na ml, w obecności lub bez obecności toksycznego antybiotyku G 418 (neomycyna). Transfekcję przeprowadzono bezpośrednio po pobraniu komórek szpiku, przed wysianiem w agarze. Następnie komórki hodowano przez szereg dni w cieplarni z przepływem CO2 (7,5%) w 37 C. W różnych odstępach czasu liczbę transformowanych kolonii (co najmniej 50 komórek) lub skupisk komórek (clusters, 10 do 50 komórek) liczono w każdym przypadku pod mikroskopem. W podanych warunkach doświadczenia, większość kolonii były to kolonie mieszane granulocytów/monocytów albo kolonie granulocytów, rzadko obserwowane "wybuchy" komórek erytroidalnych, w jednym przypadku stwierdzono megakariocyty. Typy komórek w koloniach identyfikowano przez barwienie May-Griinewald. Warunki transfekcji zostały zoptymalizowane. Pomyślne transfekcje komórek pierwotnych szpiku otrzymano w następującym układzie: 800 μl zawiesiny komórek szpiku (5 x 106 komórek w płynie do elektroporacji, jak opisano w przykładzie VII), 5 μg KTX15, elektroporacja przy 300 V, 100 Ω, 250 μfd, 6 ms.
 albo metyloceluloza.")
10 Po elektroporacji (aparat Gene Pulser and Capacitance Extender firmy Bio Rad) komórki poddano selekcji w opisanej pożywce hodowlanej w obecności 500 μg/ml G 418 (neomycyna). Oporność na G-418 wskazywała na udaną transfekcję zdolnych do propagacji komórek pierwotnych szpiku. Przykład IX. Wykazanie integracji nienaruszonych konstruktów wektorów ekspresyjnych oraz wykazanie ekspresji antysensownego RNA. Transfekowane linie komórkowe U 937 i HUT 78 poddano dezintegracji i DNA i RNA z tych linii komórkowych izolowano standardowymi metodami stosując wirowanie w gradiencie gęstości chlorku cezu (L.G. Davis i in., Basic Methods in Molecular Biology, Elsevier, Nowy Jork 1986). Dowód na integrację nienaruszonych konstruktów wektora ekspresyjnego antysensownego RNA przeprowadzono przez reakcję łańcuchową polimerazy (PCR Technology Principles and Application of DNA Amplification, wyd. Henry A. Erlich, M. Stockton Press, Nowy Jork 1989), stosując primery oligonukleotydowe, które amplifikowały zachodzące na siebie fragmenty DNA sekwencji promotora, sekwencji insertu i sekwencji terminacji. Następnie, amplifikowany DNA identyfikowano metodą analizy Southern Blot (Davis i in., patrz wyżej) używając sond genowych dla wstawek DNA. Przykłady wyników analizy są przedstawione na fig. 7, 8 i 9. Dla wektorów KTX 11, KTX 12 i KTX 15, na przykład, zastosowano flankujące primery 5'-CAA ACC CTT TGC GCC CG-3' albo 5'-ACT CGT CCA ACG ACT AT-3' z sekwencji promotora i 5'-TTT TTT CAC TGC ATT CTA CTT-3' z sekwencji poliadenylacji. W przypadku wektorów ekspresyjnych z wektorem prc-cmv na przykład, zastosowano flankujący primer 5'CTT TCC AAA ATG TCG TAA CAA CTCC-3' dla sekwencji promotora i 5'-ATT TAG GTG ACA CTA TAG AAT-3' dla sekwencji terminacji. We wstawce (sekwencja HIV) wybrano sekwencje primera, które zależnie od wybranej pary primerów dawały produkty PCR o około 500 do 2100 par zasad. Ekspresję antysensownego RNA wykazano przeprowadzając analizę Northern Blot (Davis i in., patrz wyżej) RNA z klonowanych, transfekowanych linii komórek. Przykład wyniku analizy wykazujący antysensowny RNA HIV-1 w transfekowanej linii komórek U 937 przedstawia fig. 10. Przykład X. Wykazanie oporności na wirus transfekowanych klonowanych linii komórkowych krwiotwórczych. Klonowane linie komórkowe, w których zachodzi ekspresja antysensownego RNA, otrzymane i scharakteryzowane według przykładów VII i IX, badano w porównawczych próbach zakażenia HIV-1 (L. Ratner i in., Nature 313, 277, 1985) albo HIV (H. Kühnei i in., Nucleic Acids Research 1990,18,6142) razem z wyjściowymi liniami komórek HUT 78 i U 937 jako kontrolnymi, na ekspresję oporności przeciwwirusowej. 1. Zakażenie HUT 78 i U 937 wirusem HIV-1 (kontrola) Wykonanie Porcje po 10 ml zawiesiny komórek traktowano przez 60 minut różnymi rozcieńczeniami płynu z hodowli HUT 78 (HUT-1)około 104 PFU/ml w hodowlach PBL, aktywność RT zamrożonego roztworu macierzystego cmp/ml). Następnie komórki odwirowano i trzykrotnie przemyto, w celu usunięcia niezwiązanych cząstek wirusa u białek wirusowych. Na koniec w każdym przypadku komórki zawieszano w 10 ml pożywki i hodowano w cieplarce. Co dni badano 200 μl wolnego od komórek płynu z hodowli na obecność antygenów HIV. Stosowano test wychwytywania antygenu firmy Organon Technika. Dla podtrzymania komórek za każdym razem usuwano 5 ml zawiesiny komórek i zastępowano 5 ml pożywki. 2. Zakażenie HI V-1 komórek HUT 78 i U 937 wykazujących ekspresję antysensownego RNA. Wykonanie W każdym przypadku 10 ml zawiesiny komórek (około 4 x 105 komórek na ml) traktowano przez 60 minut rozcieńczonym płynem z hodowli HUT 78/HIV-1 (około 10 PFU/ml w hodowlach PBL, aktywność RT zamrożonego roztworu macierzystego cpm/ml); końcowe rozcieńczenie roztworu macierzystego wirusa wynosiło 1:1000 dla komórek HUT 78 i 1:50 dla komórek U 937. Następnie komórki odwirowano i czterokrotnie ostrożnie przemyto w celu usunięcia niezwiązanych cząstek wirusa i białek wirusowych. Na koniec, komórki w

11 każdym przypadku zawieszano w 10 ml pożywki (RPMI % FCS) i hodowano przez 60 dni w cieplarce. Co 3-4 dni 160 μl wolnego od komórek płynu hodowlanego badano na obecność antygenów HIV, stosując test wychwytywania antygenu, firmy Organon Technika z odpowiednim czytnikiem płytek mikrotitracyjnych. Dla podtrzymania komórek w każdym przypadku co 3-4 dni usuwano 5 ml zawiesiny komórek i zastępowano 5 ml świeżej pożywki. Jako wynik reprezentatywny dla innych linii komórkowych wykazujących ekspresję antysensownego RNA można wymienić dwie serie doświadczeń z trzema transfektantami HUT 78 (HUT K14-KTX11, SSHUTTK16/2-SXK1, SSHUTK1/1-SXK2, patrz fig. 11) i z trzema transfektantami U 937 (ssuk 20/4-KTX1 la, ssuk 3/1-SXK2 i ssuk 20/15-KTX1 1b, patrz fig. 12). Po zakażeniu w ciągu inkubacji do 60 dni, zarówno w pomiarach aktywności odwrotnej transkryptazy (RT), jak również w oznaczeniach antygenu VIF te hodowle komórek wykazujące ekspresję antysensownego RNA i zawierające wektory ekspresji psck1 i psxk2, nie wykazały żadnych oznak namnażania się HI V. Natomiast w kontrolnej linii komórek namnażanie się HIV można było wyraźnie oznaczyć. Ponadto stwierdzono opóźniony wzrost replikacji wirusa w transfektantach o słabszej ekspresji wektora KTX11. Wynik jest przedstawiony na fig. 11 i 12. Dla sprawdzenia, że zakażenie HIV-1 nastąpiło w liniach komórek SSU K3/1-SXK2, SSHUT K16/2-SXK1 i SSHUT K1/1-SXK2, które po inkubacji przez 60 dni nie wykazywały antygenu HIV-1 ani aktywności RT, dodatnie zakażenie wykazało na przykład przez reakcję PCR sekwencji genu HI V-1 TAT i sekwencji genów otoczki w inkubowanych przez 60 dni liniach komórek SSHUT K16/2-SXK1 i SSHUT K1/1-SXK2 (fig. 13). Zastosowano w tym celu primer PCR 5'-ATG GAG CCA GTA GAT CCT-3' i 5'-TCT ACC ATG TCAT-3' do generowania fragmentu PCR o długości 690 par zasad. Wektory ekspresyjne psxk1 i psxk2 nie zawierają tych sekwencji i dlatego nie mogą dać fałszywie dodatniego produktu PCR. Przykład XI. Doświadczenia kontrolne. Transfekcja wektorów z przykładów I do VI, bez wstawek prowirusowego DNA HIV-1 w orientacji antysensownej, nie prowadziła w liniach do powstania komórek U 937 i HUT 78 do linii komórkowych, w których replikacja wirusa była zahamowana. Jednak doświadczenia kontrolne z wektorami z przykładów I do VI, w których są wstawki prowirusowego fragmentu DNA HIV-1 w orientacji sensownej, nie zostały przeprowadzone, ponieważ odpowiednie sensowne transkrypty mogą wiązać białka regulatorowe replikacji HIV, jak np. TAT i REV i dlatego mogą konkurencyjnie hamować replikację wirusa (G.J. Graham i in. PNAS 87, 5817, 1990) i dlatego okazują się nieprzydatne w doświadczeniach kontrolnych. Przykład XII. Zakażenie HIV-2 linii komórek HUT 78 i U 937 wykazujących ekspresję antysensownego RNA. Próby zakażenia przeprowadzono z izolatem HIV-2 D194 /H. Kϋhnle i in., Nucleic Acids Res. 18, 6142,1990/. Zawiesinę wirusa o aktywności RT 2,14 x 105 cpm/ml i stężeniu antygenu (w rozcieńczeniu 1:1000) 0,249 OD450 nm (próbka 160 μl) przechowywano w -80 C. Z tej zawiesiny wirusa do zakażenia 2 x 106 komórek HUT 78 pobrano 50 μl, dodano do 4 ml podłoża i inkubowano przez 3 godziny. Komórki U 937 zakażono (2 x 106) 100 μl zawiesiny wirusa w 4 ml podłoża i inkubowano przez 24 godziny. Następnie niezwiązane cząstki wirusa usunięto w hodowli przez przemywanie. W odstępach 3-4 dni hodowlę komórek podtrzymywano i pobierano supernatant do ilościowego oznaczania antygenu HIV, używając testu wychwytywania antygenu HIV firmy Organon Technika w połączeniu w odpowiednim fotometrem płytek mikrotitracyjnych. Przebieg replikacji HIV-2 D 194 w kontrolnych liniach komórek HUT 78 i U 937, które nie zawierają wektorów ekspresji antysensownego RNA jest porównany na fig. 14 i 15 z transfekowanymi liniami komórek, które przez ekspresję antysensownego RNA wykazują opóźnione namnażanie się HIV-2. Wyniki Stosując wektory ekspresji KTX 11, KTX 12, KTX 15, psxk 1, psxk 2, phtt 1, phtt 4, phtt 6, phtt 9, phtt 10, phtt 12 i phtt 15 (przykłady I do VI), w ludzkich liniach komórek krwi po transfekcji genu i klonowaniu transfekowanych linii komórek (przykład VII) stwierdzo-

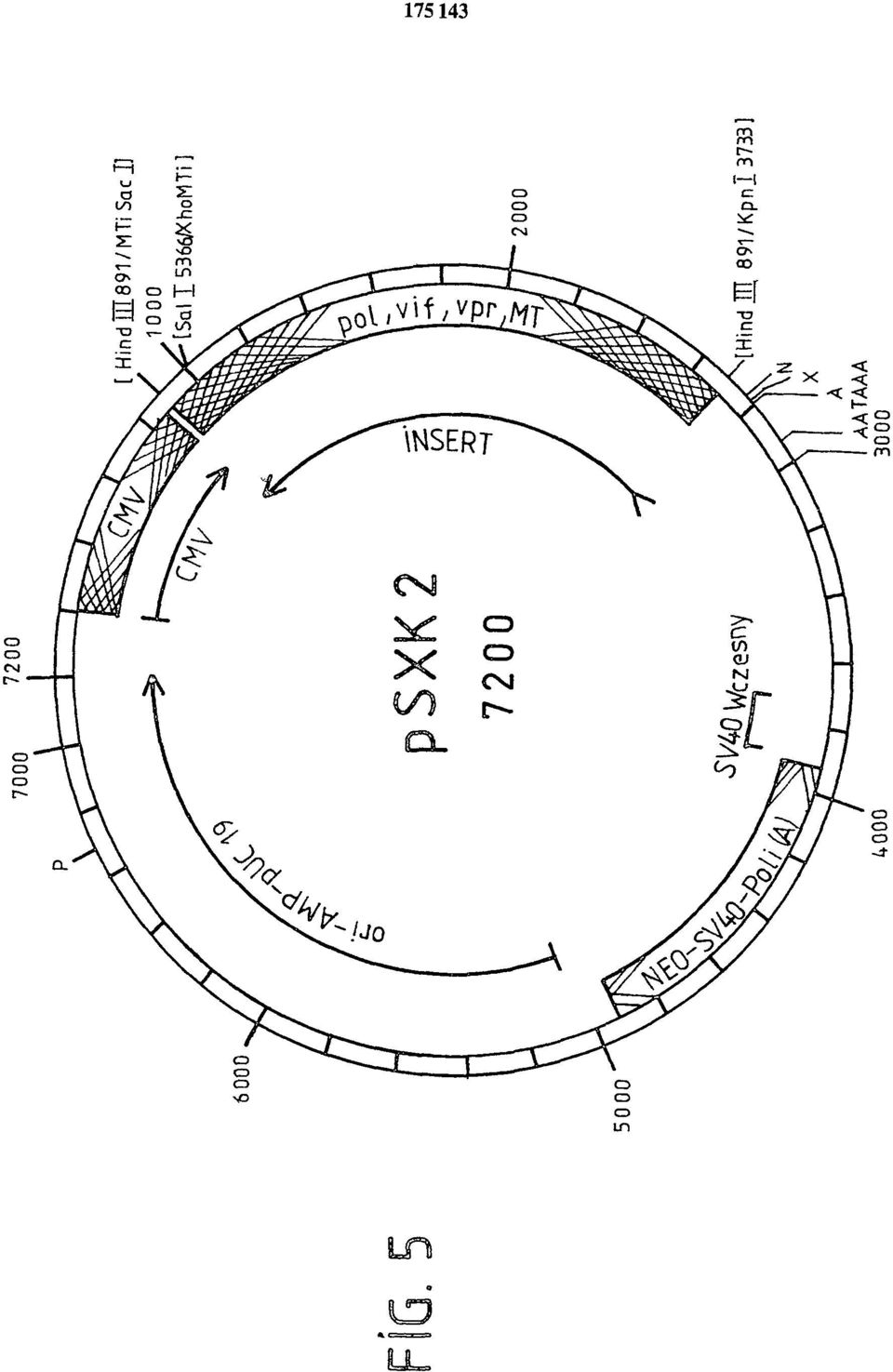
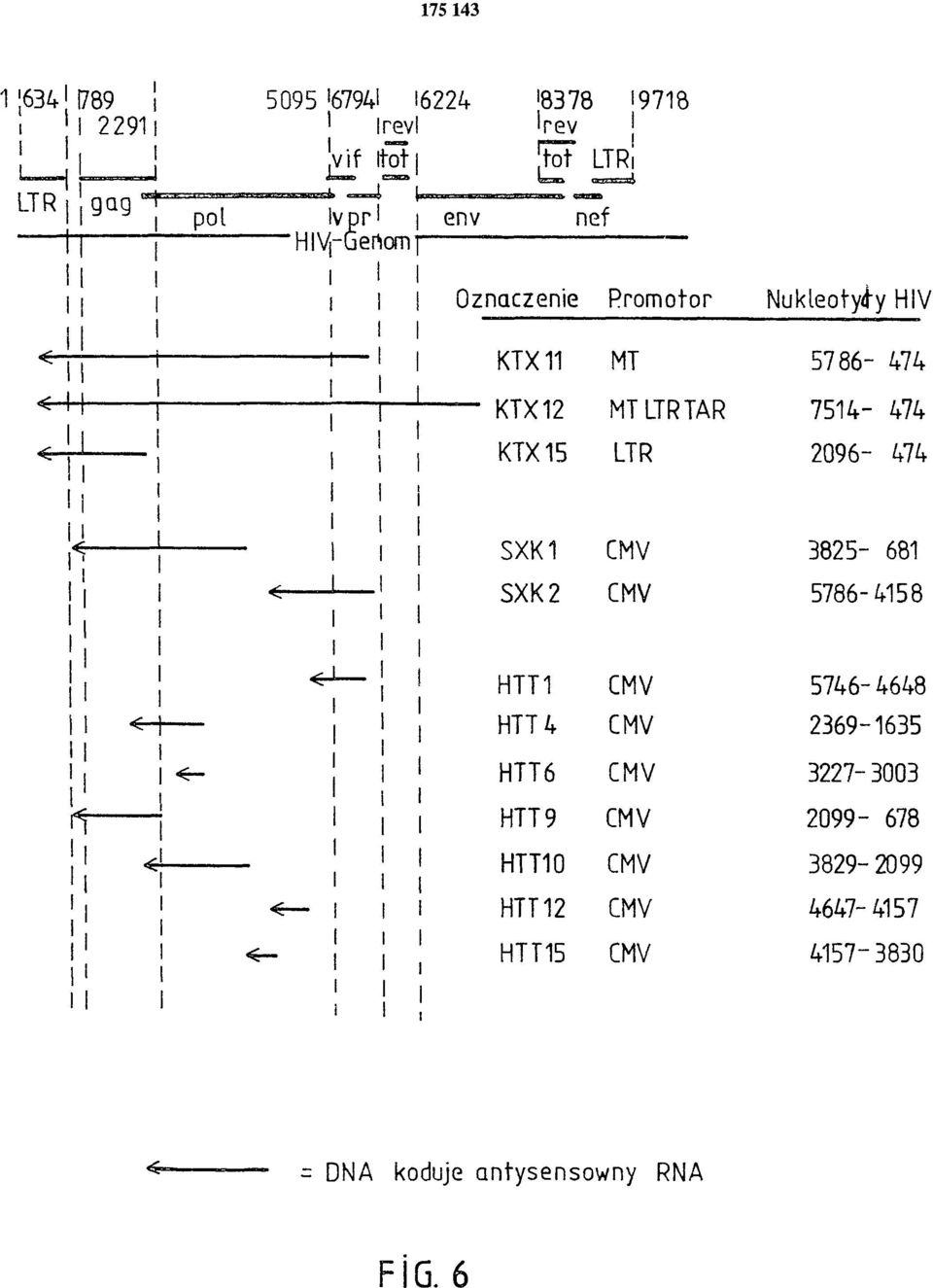
12 no endogenne wytwarzanie antysensownego RNA przeciw HIV-1 (przykład IX) i wykazano jego działanie przeciwwirusowe (przykłady X, XII). Nieoczekiwanie stwierdzono wyraźne hamowanie replikacji retrowirusa HIV-1 w transfekowanych liniach komórek. Podobnie przy użyciu HIV-1 specjalnie adaptowanego do transfekowanych komórek, można było wykazać oporność na wirus komórek z ekspresją antysensownego RNA (fig. 11, 12). Obserwowano również wyraźne cechy przeciwwirusowe antysensownego RNA przeciw HIV-2 (fig. 14,15). Wyniki te są przykładem, że odpowiednie promotory ekspresji cząsteczek antysensownego RNA i oczywiście rodzaj sekwencji antysensownego RNA, prowadzą w genetycznie zmienionych komórkach do wytworzenia oporności na zakażenie retrowirusem. W przykładzie IX wykazano, że w związku z dotychczas nieznanym procesem cięcia i składania RNA HI V-1, transkryptów nici /+/ prowirusowego DNA w orientacji 3' > 5', powstało kilka cząsteczek mrna, które prowadzą do efektu przeciwwirusowego. Prace doświadczalne, które opisano w przykładach I do XII, wykazały, że stosując odpowiednie konstrukty wektorów ekspresyjnych z fragmentami DNA retrowirusa, kodującymi antysensowny RNA, można otrzymać w komórkach właściwości przeciwretrowirusowe, które chronią przed zakażeniem retrowirusowym. Z fig. 11 do 13 widać, że linie komórek SSHUT K16/2-SXK1 i SSHUT K1/1-SXK2 po 60 dniach od zakażenia HIV są dodatnie w analizie PCR prowirusowego DNA HI V, chociaż oznaczanie antygenu HI V jest ujemne. Wynik ten pozwala wnosić, że po zakażeniu komórek HIV-1, wprawdzie w pierwszym etapie cyklu replikacyjnego HIV powstaje jeszcze prowirusowy DNA w transfekowanych liniach komórek HUT 78, ale antysensowny RNA zapobiega transkrypcji/translacji prowirusowego DNA i nie wytwarza się białko HIV do replikacji wirusa w dających się oznaczyć ilościach. W DNA linii komórkowej SSU K3/1-SXK2 w 60 dni po zakażeniu HIV-1, we wszystkich próbach testowych nie można było wykryć wytwarzania białek HIV względnie prowirusowego DNA HIV ani testem na antygeny ani analizą PCR. Ten wynik może oznaczać, że w linii komórek SSU K3/1-SXK2 wytworzony antysensowny RNA nie tylko zapobiega drugiemu etapowi cyklu replikacyjnego HIV (transkrypcja i translacja prowirusowego DNA) ale także hamuje już pierwszy etap cyklu replikacyjnego. Antysensowny RNA może najwyraźniej zapobiegać tworzeniu się prowirusowego DNA z genomowej nici /+/ RNA przez proces odwrotnej translacji. To działanie antysensownego RNA przyjęto spekulatywnie, jednak dotychczas nie zostało ono wykazane (R. Y.L. To i P.E. Neiman, Gene Regulation, Biology of antisense RNA and DNA, wyd. R.P. Erickson i J.G. Izant, Raven Press Ltd., Nowy Jork 1992), ponieważ dotychczas nie znaleziono odpowiednio wydajnych konstruktów wektorów ekspresyjnych. Opisy figur Figura 1 Wektor ekspresyjny KTX 11, do ekspresji antysensownego RNA HIV-1. Promotor metalotioneiny (MTI) uruchamia transkrypcję antysensownego RNA z prowirusowego fragmentu DNA HIV-1, obejmującego nukleotydy 5786 > 474 (numeracja: UWGCG GENEMBL DA- TA BANK File; HIV HXB2CG.VIRAL). Terminacja transkryptów antysensownego RNA następuje przez sekwencję poliadenylacji SV 40. Transkrypcja genu oporności na neomycynę TRN5neo jest zainicjowana przez wczesny promotor SV 40. Wektor wahadłowy zawiera także początek pbr322 oraz gen ampicyliny (pbr 322-amp). Dalsze szczegóły opisano w przykładzie I. Figura 2 Wektor ekspresyjny KTX 12, do ekspresji antysensownego RNA HIV-1 pod kontrolą promotora hybrydowego metalotioneiny LTR HIV-1. Szczegóły budowy wektora są opisane w przykładzie II. Figura 3 Wektor ekspresyjny KTX 15, do ekspresji antysensownego RNA HIV-1 pod kontrolą promotora hybrydowego metalotioneiny LTR HIV-1, w którym jest delecja sekwencji TAR
.")
13 HIV-1 dla osiągnięcia maksymalnej ekspresji bez obecności białka transaktywatora TAT. Szczegóły budowy wektora opisano w przykładzie III Figura 4 Wektor ekspresyjny psxk 1, do ekspresji antysensownego RNA HIV-1 pod kontrolą promotora wirusa cytomegalii (CMV). Szczegóły budowy wektora są opisane w przykładzie IV. Figura 5 Wektor ekspresyjny psxk 2, do ekspresji antysensownego RNA HIV-1 pod kontrolą promotora cytomegalowirusa (CMV). Prowirusowy fragment DNA HI V-1 ze środkowego regionu genomu HIV obejmuje w tym przykładzie nie sekwencje z regionu 5' jak w przykładach I do IV, ale sekwencję z regionu POL/VIF. Szczegóły budowy wektora opisano w przykładzie V. Figura 6 Orientacyjny digram dla przygotowania fragmentów genowych HIV-1, które w antysensownej orientacji 3' > 5' (kierunek strzałki) są zawarte w wektorach ekspresyjnych antysensownego RNA, z oznaczeniami podanymi w kolumnie 1. Promotor odpowiedniego wektora ekspresyjnego jest podany w kolumnie 2, w kolumnie 3 numeracja nukleotydów sekwencji HI V według UWGCG GENEMBL DATA BANK file: HIVHXB2CG.VIRAL. Figura 7 Analiza Southern Blot, po amplifikacji PCR, chromosomowego DNA z klonowanego transfektanta U 937, który zawiera wektor ekspresyjny psxk1 zintegrowany w chromosomowym DNA. Wielkość fragmentu produktów PCR, oraz hybrydyzacja z sondą genową HIV, która nie zawiera sekwencji primera produktów PCR, wskazują na nienaruszony charakter sekwencji promotor/insert. 1. Wektor próby kontrolnej, połączenie 5', fragment z 887 par zasad 2. Wektor próby kontrolnej, połączenie 3', fragment z 775 par zasad 3. Chromosomowy DNA, połączenie 5', fragment z 887 par zasad 4. Chromosomowy DNA, połączenie 3', fragment z 775 par zasad. Dalsze szczegóły podano w przykładzie IX. Figura 8 Wykrywanie fragmentu genu HIV-1 w chromosomowym DNA z klonowanych transfektantów HUT 78, przy pomocy amplifikacji PCR. Reakcję PCR rozdzielono na żelu agaroza- TAE i po barwieniu bromkiem etydyny fotografowano w świetle UV. Produkt PCR obejmuje 561 par zasad i odpowiada fragmentowi genu HIV-1(nukleotydy ), który jest również składnikiem wektora psxk1, Produkt PCR o długości 561 bp pokazuje strzałka. M = standard długości DNA, pokazane są 516 bp i = kontrola ujemna, DNA nie dodano do reakcji PCR 2 = chromosomowy DNA z komórek U 937 (typ dziki) 3 = chromosomowy DNA z komórek HUT 78 (typ dziki) 4 = kontrola ujemna, DNA nie dodano do reakcji PCR 5 = kontrola dodatnia 6 = chromosomowy DNA z transformantów HUT 78 (SSHUT 6/ /1) 7 = chromosomowy DNA z transformantów HUT 78 (SSHUT 6/ /1) 8 = chromosomowy DNA z transformantów HUT 78 (SSHUT 6/ /1) 9 = chromosomowy DNA z transformantów HUT 78 (SSHUT 6/ /1) Figura 9 Wykrywanie fragmentu genu HIV-1 (nukleotydy ) w chromosomowym DNA w monocytów U 937 i limfocytów T HUT 78, przy pomocy amplifikacji PCR. Komórki uprzednio transfekowano wektorem psxk2. Reakcję PCR rozdzielono na żelu agaroza TAE i po barwieniu bromkiem etydyny fotografowano w świetle UV. Produkt PCR obejmuje cały fragment genu HIV-1 w psxk2 (nukleotydy ) i ma długość 1811 bp. Strzałka pokazuje produkt PCR o długości 1811 bp. M = standard długości DNA, 1635 bp i 2036 bp są pokazane 1 = kontrola ujemna, DNA nie dodano reakcji PCR

14 = chromosomowy DNA z komórek U 937 (typ dziki) 3 = chromosomowy DNA z komórek HUT 78 (typ dziki) 4 = chromosomowy DNA z transformantów HUT 78 (SSUK 3/ /1) 5 = chromosomowy DNA z transformantów U 937 (SSUK 3/ /1) Figura 10 Analiza Northern Blot dla oznaczenia szeregu transkryptów antysensownego RNA w transfekowanej, klonowanej linii komórek U 937, która zawiera wektor ekspresji KTX 11 w linii komórek SSUK 20/15-KTX1 1b. Przy pomocy sondy genowej, która odpowiada wstawce DNA wektora KTX 11, w RNA linii komórek opornych na wirus (pasmo 1) można stwierdzić co najmiej trzy transkrypty antysensownego RNA, o długości 800,1200 i 3200 bp. Pasmo 2 zawiera RNA z nie transfekowanej linii komórek U 937 (kontrola ujemna). Pasmo 3 zawiera standardy ciężaru cząsteczkowego. Figura 11 Przebieg wytwarzania antygenu HIV-1 w ciągu 60 dni po zakażeniu (w dniu zero) linii komórek HUT 78 izolatem HIV-1 LAV/BRU. Na osi rzędnych podano gęstości optyczne przy 450 nm (OD450nm) wyników testu ELISA HIV z płynem z hodowli komórek. Przy gęstości optycznej > 2,0 płyn z hodowli rozcieńczano i podaną wartość wyliczano z rozcieńczenia. Wartość odciętej przedstawia granicę wykrywalności oznaczenia antygenu HIV. W liniach komórek SSHUT K 16/2-SXK1 i SSHUT K1/1-SXK2, wykazujących ekspresję antysensownego RNA, replikacja HIV jest całkowicie zahamowana w okresie 60 dni. W linii komórek HUT K14-KTX11 początek replikacji HIV jest opóźniony w porównaniu z nietransfekowaną linią komórek typu dzikiego HUT 78. Figura 12 Przebieg wytwarzania antygenu HIV-1 w okresie 60 dni po zakażeniu (w dniu zero) linii komórek U 937 izolatem HIV-1 LAV/BRU. Na osi rzędnych podano gęstość optyczną przy 450 nm (OD450nm) wyników testu ELISA HIV z płynem z hodowli komórek. Przy gęstości optycznej > 2,0 płyn z hodowli rozcieńczano i podaną wartość wyliczano z rozcieńczenia. Wartość odciętej przedstawia granicę wykrywalności oznaczenia antygenu HIV. W linii komórek SSUK 3/1-SXK2, wykazującej ekspresję antysensownego RNA, replikacja HIV jest w zakresie 60 dni całkowicie zahamowana, pozostałe transfekowane linie komórek wykazują opóźnienie replikacji HIV w porównaniu z linią komórek typu dzikiego (U 937). Figura 13 Analiza PCR celem oznaczenia DNA HIV-1 w liniach komórek, które zawierają wektory ekspresyjne antysensownego RNA i były badane 60 dni po zakażeniu HIV. Barwiona bromkiem etydyny elektroforeza w żelu agarozowym pokazuje produkty PCR chromosomowego DNA z podanych niżej linii komórek. Przy pomocy pary primerów 5'-ATG GAG CCA GTA GAT CCT-3' i 5'-TCT GT TCT ACC ATG TCAT, w komórkach zakażonych HIV poddano amplifikacji fragment PCR długości 702 par zasad z regionu genu TAT-ENV. Dla porównania z wynikiem PCR, w nawiasie podano wynik testu na antygen HIV odpowiednich próbek hodowli komórek. Pasmo M : standard ciężaru cząsteczkowego, zaznaczono 600 bp Pasmo 1 : HUT K14-KTX11, 702 bp dodatni (test antygenowy dodatni) Pasmo 2 : SSHUT K16/2-SXK1, 702 dodatni (test antygenowy ujemny) Pasmo 3 : Produkt PCR z równoległego doświadczenia z zakażeniem jak w paśmie 2 Pasmo 4 : Produkt PCR z równoległego doświadczenia z zakażeniem jak w paśmie 2 Pasmo 5 : SSHUT K1/1-SXK2, 702 dodatni (test antygenowy ujemny) Pasmo 6 : Produkt PCR z równoległego doświadczenia z zakażeniem jak w paśmie 5 Pasmo 7 : Równoległe doświadczenie z zakażeniem jak w paśmie 5 Pasmo 8 : HUT 78, kontrola dodatnia Pasma 9-11: Hodowle SSUK3/1-SXK2 - produkt PCR 702 bp niewykrywalny (test antygenowy ujemny)
 można stwierdzić co najmiej trzy transkrypty antysensownego RNA, o długości")
15 Figura 14 Przebieg wytwarzania antygenu HIV w ciągu 30 dni po zakażeniu (w dniu zero) linii komórek HUT 78 izolatem D 194 HIV-2. Na osi rzędnych podano gęstość optyczną przy 450 nm (OD450nm) wyników testu ELIS A HIV z płynem z hodowli komórek. Przy gęstości optycznej > 2,0 płyn z hodowli rozcieńczano i podaną wartość OD wyliczano z rozcieńczenia. Wartość odciętej przedstawia granicę wykrywalności oznaczenia antygenu HI V. W linii komórek SSHUT K1/1-SXK2, wykazującej ekspresję antysensownego RNA replikacja HIV jest w okresie 14 dni całkowicie zahamowana, pozostałe transfekowane linie komórek wykazują opóźnienie replikacji HIV w porównaniu z linią komórek typu dzikiego (HUT 78). Figura 15 Przebieg wytwarzania antygenu HIV w ciągu 30 dni po zakażeniu (w dniu zero) linii komórek U 937 izolatem D 194 HIV-2. Na osi rzędnych podano gęstość optyczną przy 450 nm (OD450nm) wyników testu ELIS A HIV z płynem z hodowli komórek. Przy gęstości optycznej > 2,0 płyn z hodowli rozcieńczano i podaną wartość OD wyliczano z rozcieńczenia. Wartość odciętej przedstawia granicę wykrywalności oznaczenia antygenu HI V. W linii komórek SSHUT K3/1-SXK2, wykazującej ekspresję antysensownego RNA, replikacja HIV jest w okresie 30 dni całkowicie zahamowana, pozostała transfekowana linia komórek wykazuj ą opóźnienie replikacji HIV w porównaniu z linią komórek typu dzikiego (U 937).

16 FIG. 2

17 FIG. 3

18 FIG

19 FIG. 5
20 FIG. 6
Klonowanie molekularne Kurs doskonalący. Zakład Geriatrii i Gerontologii CMKP
 Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Zawartość. Wstęp 1. Historia wirusologii. 2. Klasyfikacja wirusów
 Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
Zawartość 139585 Wstęp 1. Historia wirusologii 2. Klasyfikacja wirusów 3. Struktura cząstek wirusowych 3.1. Metody określania struktury cząstek wirusowych 3.2. Budowa cząstek wirusowych o strukturze helikalnej
Inżynieria Genetyczna ćw. 3
 Materiały do ćwiczeń z przedmiotu Genetyka z inżynierią genetyczną D - blok Inżynieria Genetyczna ćw. 3 Instytut Genetyki i Biotechnologii, Wydział Biologii, Uniwersytet Warszawski, rok akad. 2018/2019
Materiały do ćwiczeń z przedmiotu Genetyka z inżynierią genetyczną D - blok Inżynieria Genetyczna ćw. 3 Instytut Genetyki i Biotechnologii, Wydział Biologii, Uniwersytet Warszawski, rok akad. 2018/2019
KLONOWANIE DNA REKOMBINACJA DNA WEKTORY
 KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA
 Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa IZOLACJA DNA Z HODOWLI KOMÓRKOWEJ.
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa IZOLACJA DNA Z HODOWLI KOMÓRKOWEJ.
Ekspresja białek w komórkach ssaczych
 Ekspresja białek w komórkach ssaczych Ekspresja białek w komórkach ssaczych Używane powszechnie w laboratoriach ssacze linie komórkowe są zmodyfikowane w celu pełnienia roli gospodarza ekspresji białek
Ekspresja białek w komórkach ssaczych Ekspresja białek w komórkach ssaczych Używane powszechnie w laboratoriach ssacze linie komórkowe są zmodyfikowane w celu pełnienia roli gospodarza ekspresji białek
Inżynieria genetyczna- 6 ECTS. Inżynieria genetyczna. Podstawowe pojęcia Część II Klonowanie ekspresyjne Od genu do białka
 Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Nowoczesne systemy ekspresji genów
 Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Ćwiczenie 3. Amplifikacja genu ccr5 Homo sapiens wykrywanie delecji Δ32pz warunkującej oporność na wirusa HIV
 Ćwiczenie 3. Amplifikacja genu ccr5 Homo sapiens wykrywanie delecji Δ32pz warunkującej oporność na wirusa HIV Cel ćwiczenia Określenie podatności na zakażenie wirusem HIV poprzez detekcję homo lub heterozygotyczności
Ćwiczenie 3. Amplifikacja genu ccr5 Homo sapiens wykrywanie delecji Δ32pz warunkującej oporność na wirusa HIV Cel ćwiczenia Określenie podatności na zakażenie wirusem HIV poprzez detekcję homo lub heterozygotyczności
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA
 Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa tel. 22 572 0735, 606448502
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa tel. 22 572 0735, 606448502
E.coli Transformer Kit
 E.coli Transformer Kit zestaw do przygotowywania i transformacji komórek kompetentnych Escherichia coli. Metoda chemiczna. wersja 1117 6 x 40 transformacji Nr kat. 4020-240 Zestaw zawiera komplet odczynników
E.coli Transformer Kit zestaw do przygotowywania i transformacji komórek kompetentnych Escherichia coli. Metoda chemiczna. wersja 1117 6 x 40 transformacji Nr kat. 4020-240 Zestaw zawiera komplet odczynników
Ćwiczenia 1 Wirtualne Klonowanie Prowadzący: mgr inż. Joanna Tymeck-Mulik i mgr Lidia Gaffke. Część teoretyczna:
 Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
mikrosatelitarne, minisatelitarne i polimorfizm liczby kopii
 Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
Zawartość 139371 1. Wstęp zarys historii genetyki, czyli od genetyki klasycznej do genomiki 2. Chromosomy i podziały jądra komórkowego 2.1. Budowa chromosomu 2.2. Barwienie prążkowe chromosomów 2.3. Mitoza
Saccharomyces Transformer Kit zestaw do przygotowywania i transformacji komórek kompetentnych Saccharomyces cerevisiae. Metoda chemiczna.
 Saccharomyces Transformer Kit zestaw do przygotowywania i transformacji komórek kompetentnych Saccharomyces cerevisiae. Metoda chemiczna. wersja 0916 6 x 20 transformacji Nr kat. 4010-120 Zestaw zawiera
Saccharomyces Transformer Kit zestaw do przygotowywania i transformacji komórek kompetentnych Saccharomyces cerevisiae. Metoda chemiczna. wersja 0916 6 x 20 transformacji Nr kat. 4010-120 Zestaw zawiera
E.coli Transformer Zestaw do przygotowywania i transformacji komórek kompetentnych Escherichia coli
 E.coli Transformer Zestaw do przygotowywania i transformacji komórek kompetentnych Escherichia coli Wersja 0211 6x40 transformacji Nr kat. 4020-240 Zestaw zawiera komplet odczynników do przygotowania sześciu
E.coli Transformer Zestaw do przygotowywania i transformacji komórek kompetentnych Escherichia coli Wersja 0211 6x40 transformacji Nr kat. 4020-240 Zestaw zawiera komplet odczynników do przygotowania sześciu
Zestawy do izolacji DNA i RNA
 Syngen kolumienki.pl Gotowe zestawy do izolacji i oczyszczania kwasów nukleinowych Zestawy do izolacji DNA i RNA Katalog produktów 2012-13 kolumienki.pl Syngen Biotech Sp. z o.o., 54-116 Wrocław, ul. Ostródzka
Syngen kolumienki.pl Gotowe zestawy do izolacji i oczyszczania kwasów nukleinowych Zestawy do izolacji DNA i RNA Katalog produktów 2012-13 kolumienki.pl Syngen Biotech Sp. z o.o., 54-116 Wrocław, ul. Ostródzka
Polimeraza Taq (1U/ l) 1-2 U 1 polimeraza Taq jako ostatni składniki mieszaniny końcowa objętość
 1-2 U 1 polimeraza Taq jako ostatni składniki mieszaniny końcowa objętość") Ćwiczenie 6 Technika PCR Celem ćwiczenia jest zastosowanie techniki PCR do amplifikacji fragmentu DNA z bakterii R. leguminosarum bv. trifolii TA1 (RtTA1). Studenci przygotowują reakcję PCR wykorzystując
Ćwiczenie 6 Technika PCR Celem ćwiczenia jest zastosowanie techniki PCR do amplifikacji fragmentu DNA z bakterii R. leguminosarum bv. trifolii TA1 (RtTA1). Studenci przygotowują reakcję PCR wykorzystując
Inżynieria genetyczna
 Inżynieria genetyczna i technologia rekombinowanego DNA Dr n. biol. Urszula Wasik Zakład Biologii Medycznej Inżynieria genetyczna świadoma, celowa, kontrolowana ingerencja w materiał genetyczny organizmów
Inżynieria genetyczna i technologia rekombinowanego DNA Dr n. biol. Urszula Wasik Zakład Biologii Medycznej Inżynieria genetyczna świadoma, celowa, kontrolowana ingerencja w materiał genetyczny organizmów
Zestaw do wykrywania Chlamydia trachomatis w moczu lub w kulturach komórkowych
 Nr kat. PK15 Wersja zestawu: 1.2016 Zestaw do wykrywania w moczu lub w kulturach komórkowych na 50 reakcji PCR (50µl), włączając w to kontrole Detekcja oparta jest na amplifikacji fragmentu genu crp (cysteine
Nr kat. PK15 Wersja zestawu: 1.2016 Zestaw do wykrywania w moczu lub w kulturach komórkowych na 50 reakcji PCR (50µl), włączając w to kontrole Detekcja oparta jest na amplifikacji fragmentu genu crp (cysteine
Zestaw do wykrywania Anaplasma phagocytophilum w kleszczach, krwi i hodowlach komórkowych
 Nr kat. PK24N Wersja zestawu: 1.2016 Zestaw do wykrywania phagocytophilum w kleszczach, krwi i hodowlach komórkowych dwie oddzielne reakcje PCR 2x50 reakcji PCR (50 µl), włączając w to kontrole Detekcja
Nr kat. PK24N Wersja zestawu: 1.2016 Zestaw do wykrywania phagocytophilum w kleszczach, krwi i hodowlach komórkowych dwie oddzielne reakcje PCR 2x50 reakcji PCR (50 µl), włączając w to kontrole Detekcja
Dr. habil. Anna Salek International Bio-Consulting 1 Germany
 1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
ĆWICZENIA Z MECHANIZMÓW DZIAŁANIA WYBRANYCH GRUP LEKÓW
 UNIWERSYTET MARII CURIE-SKŁODOWSKIEJ WYDZIAŁ BIOLOGII I BIOTECHNOLOGII ZAKŁAD BIOLOGII MOLEKULARNEJ ĆWICZENIA Z MECHANIZMÓW DZIAŁANIA WYBRANYCH GRUP LEKÓW dla studentów I roku II 0 biotechnologii medycznej
UNIWERSYTET MARII CURIE-SKŁODOWSKIEJ WYDZIAŁ BIOLOGII I BIOTECHNOLOGII ZAKŁAD BIOLOGII MOLEKULARNEJ ĆWICZENIA Z MECHANIZMÓW DZIAŁANIA WYBRANYCH GRUP LEKÓW dla studentów I roku II 0 biotechnologii medycznej
TECHNIKI ANALIZY RNA TECHNIKI ANALIZY RNA TECHNIKI ANALIZY RNA
 DNA 28SRNA 18/16S RNA 5SRNA mrna Ilościowa analiza mrna aktywność genów w zależności od wybranych czynników: o rodzaju tkanki o rodzaju czynnika zewnętrznego o rodzaju upośledzenia szlaku metabolicznego
DNA 28SRNA 18/16S RNA 5SRNA mrna Ilościowa analiza mrna aktywność genów w zależności od wybranych czynników: o rodzaju tkanki o rodzaju czynnika zewnętrznego o rodzaju upośledzenia szlaku metabolicznego
Ćwiczenie 5 Klonowanie DNA w wektorach plazmidowych
 Ćwiczenie 5 Klonowanie DNA w wektorach plazmidowych Celem ćwiczenia jest sklonowanie fragmentu DNA (powielonego techniką PCR i wyizolowanego z żelu agarozowego na poprzednich zajęciach) do wektora plazmidowego
Ćwiczenie 5 Klonowanie DNA w wektorach plazmidowych Celem ćwiczenia jest sklonowanie fragmentu DNA (powielonego techniką PCR i wyizolowanego z żelu agarozowego na poprzednich zajęciach) do wektora plazmidowego
Powodzenie reakcji PCR wymaga właściwego doboru szeregu parametrów:
 Powodzenie reakcji PCR wymaga właściwego doboru szeregu parametrów: dobór warunków samej reakcji PCR (temperatury, czas trwania cykli, ilości cykli itp.) dobór odpowiednich starterów do reakcji amplifikacji
Powodzenie reakcji PCR wymaga właściwego doboru szeregu parametrów: dobór warunków samej reakcji PCR (temperatury, czas trwania cykli, ilości cykli itp.) dobór odpowiednich starterów do reakcji amplifikacji
Zestaw do wykrywania Babesia spp. i Theileria spp. w kleszczach, krwi i hodowlach komórkowych
 Nr kat. PK25N Wersja zestawu: 1.2012 Zestaw do wykrywania spp. i Theileria spp. w kleszczach, krwi i hodowlach komórkowych dwie oddzielne reakcje PCR 2x50 reakcji PCR (50 µl), włączając w to kontrole Zestaw
Nr kat. PK25N Wersja zestawu: 1.2012 Zestaw do wykrywania spp. i Theileria spp. w kleszczach, krwi i hodowlach komórkowych dwie oddzielne reakcje PCR 2x50 reakcji PCR (50 µl), włączając w to kontrole Zestaw
2. Enzymy pozwalające na manipulację DNA a. Polimerazy DNA b. Nukleazy c. Ligazy
 Metody analizy DNA 1. Budowa DNA. 2. Enzymy pozwalające na manipulację DNA a. Polimerazy DNA b. Nukleazy c. Ligazy 3. Klonowanie in vivo a. w bakteriach, wektory plazmidowe b. w fagach, kosmidy c. w drożdżach,
Metody analizy DNA 1. Budowa DNA. 2. Enzymy pozwalające na manipulację DNA a. Polimerazy DNA b. Nukleazy c. Ligazy 3. Klonowanie in vivo a. w bakteriach, wektory plazmidowe b. w fagach, kosmidy c. w drożdżach,
Instrukcje do ćwiczeń oraz zakres materiału realizowanego na wykładach z przedmiotu Inżynieria bioprocesowa na kierunku biotechnologia
 1 Zakład Mikrobiologii UJK Instrukcje do ćwiczeń oraz zakres materiału realizowanego na wykładach z przedmiotu Inżynieria bioprocesowa na kierunku biotechnologia 2 Zakład Mikrobiologii UJK Zakres materiału
1 Zakład Mikrobiologii UJK Instrukcje do ćwiczeń oraz zakres materiału realizowanego na wykładach z przedmiotu Inżynieria bioprocesowa na kierunku biotechnologia 2 Zakład Mikrobiologii UJK Zakres materiału
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE. Ewa Waszkowska ekspert UPRP
 WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
Dane mikromacierzowe. Mateusz Markowicz Marta Stańska
 Dane mikromacierzowe Mateusz Markowicz Marta Stańska Mikromacierz Mikromacierz DNA (ang. DNA microarray) to szklana lub plastikowa płytka (o maksymalnych wymiarach 2,5 cm x 7,5 cm) z naniesionymi w regularnych
Dane mikromacierzowe Mateusz Markowicz Marta Stańska Mikromacierz Mikromacierz DNA (ang. DNA microarray) to szklana lub plastikowa płytka (o maksymalnych wymiarach 2,5 cm x 7,5 cm) z naniesionymi w regularnych
PL B1. UNIWERSYTET PRZYRODNICZY W LUBLINIE, Lublin, PL BUP 26/11
 PL 214501 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 214501 (13) B1 (21) Numer zgłoszenia: 391458 (51) Int.Cl. C12Q 1/68 (2006.01) C12N 15/29 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej
PL 214501 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 214501 (13) B1 (21) Numer zgłoszenia: 391458 (51) Int.Cl. C12Q 1/68 (2006.01) C12N 15/29 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej
Ćwiczenie 2. Identyfikacja płci z wykorzystaniem genu amelogeniny (AMGXY)
") Ćwiczenie 2. Identyfikacja płci z wykorzystaniem genu amelogeniny (AMGXY) Cel ćwiczenia Amplifikacja fragmentu genu amelogeniny, znajdującego się na chromosomach X i Y, jako celu molekularnego przydatnego
Ćwiczenie 2. Identyfikacja płci z wykorzystaniem genu amelogeniny (AMGXY) Cel ćwiczenia Amplifikacja fragmentu genu amelogeniny, znajdującego się na chromosomach X i Y, jako celu molekularnego przydatnego
Wykład 14 Biosynteza białek
 BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
Diagnostyka wirusologiczna w praktyce klinicznej
 Diagnostyka wirusologiczna w praktyce klinicznej Ponad 60% zakażeń w praktyce klinicznej jest wywołana przez wirusy. Rodzaj i jakość materiału diagnostycznego (transport!) oraz interpretacja wyników badań
Diagnostyka wirusologiczna w praktyce klinicznej Ponad 60% zakażeń w praktyce klinicznej jest wywołana przez wirusy. Rodzaj i jakość materiału diagnostycznego (transport!) oraz interpretacja wyników badań
Badanie funkcji genu
 Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Metody badania ekspresji genów
 Metody badania ekspresji genów dr Katarzyna Knapczyk-Stwora Warunki wstępne: Proszę zapoznać się z tematem Metody badania ekspresji genów zamieszczonym w skrypcie pod reakcją A. Lityńskiej i M. Lewandowskiego
Metody badania ekspresji genów dr Katarzyna Knapczyk-Stwora Warunki wstępne: Proszę zapoznać się z tematem Metody badania ekspresji genów zamieszczonym w skrypcie pod reakcją A. Lityńskiej i M. Lewandowskiego
WPROWADZENIE DO GENETYKI MOLEKULARNEJ
 WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
Badanie funkcji genu
 Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Ćwiczenie 7 Klonowanie DNA w wektorach plazmidowych
 Ćwiczenie 7 Klonowanie DNA w wektorach plazmidowych Celem ćwiczenia jest sklonowanie fragmentu DNA (powielonego techniką PCR i wyizolowanego z żelu agarozowego na poprzednich zajęciach) do wektora plazmidowego
Ćwiczenie 7 Klonowanie DNA w wektorach plazmidowych Celem ćwiczenia jest sklonowanie fragmentu DNA (powielonego techniką PCR i wyizolowanego z żelu agarozowego na poprzednich zajęciach) do wektora plazmidowego
Biologia medyczna, materiały dla studentów
 Zasada reakcji PCR Reakcja PCR (replikacja in vitro) obejmuje denaturację DNA, przyłączanie starterów (annealing) i syntezę nowych nici DNA (elongacja). 1. Denaturacja: rozplecenie nici DNA, temp. 94 o
Zasada reakcji PCR Reakcja PCR (replikacja in vitro) obejmuje denaturację DNA, przyłączanie starterów (annealing) i syntezę nowych nici DNA (elongacja). 1. Denaturacja: rozplecenie nici DNA, temp. 94 o
Pytania Egzamin magisterski
 Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
fix RNA Roztwór do przechowywania i ochrony przed degradacją próbek przeznaczonych do izolacji RNA kat. nr. E0280 Sierpień 2018
 Sierpień 2018 fix RNA Roztwór do przechowywania i ochrony przed degradacją próbek przeznaczonych do izolacji RNA kat. nr. E0280 EURx Ltd. 80-297 Gdansk Poland ul. Przyrodnikow 3, NIP 957-07-05-191 KRS
Sierpień 2018 fix RNA Roztwór do przechowywania i ochrony przed degradacją próbek przeznaczonych do izolacji RNA kat. nr. E0280 EURx Ltd. 80-297 Gdansk Poland ul. Przyrodnikow 3, NIP 957-07-05-191 KRS
SYLABUS. Wydział Biologiczno-Rolniczy. Katedra Biochemii i Biologii Komórki
 SYLABUS 1.1. PODSTAWOWE INFORMACJE O PRZEDMIOCIE/MODULE Nazwa przedmiotu/ modułu Biologia molekularna Kod przedmiotu/ modułu* Wydział (nazwa jednostki prowadzącej kierunek) Nazwa jednostki realizującej
SYLABUS 1.1. PODSTAWOWE INFORMACJE O PRZEDMIOCIE/MODULE Nazwa przedmiotu/ modułu Biologia molekularna Kod przedmiotu/ modułu* Wydział (nazwa jednostki prowadzącej kierunek) Nazwa jednostki realizującej
Ćwiczenia 1 Wirtualne Klonowanie. Prowadzący: mgr Anna Pawlik i mgr Maciej Dylewski. Część teoretyczna:
 Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią ===============================================================================================
Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią ===============================================================================================
Biologia molekularna wirusów. Materiały dydaktyczne współfinansowane ze środków Unii Europejskiej w ramach Europejskiego Funduszu Społecznego.
 Biologia molekularna wirusów Materiały dydaktyczne współfinansowane ze środków Unii Europejskiej w ramach Europejskiego Funduszu Społecznego. Co to jest wirus? Cząsteczka złożona z kwasu nukleinowego (DNA
Biologia molekularna wirusów Materiały dydaktyczne współfinansowane ze środków Unii Europejskiej w ramach Europejskiego Funduszu Społecznego. Co to jest wirus? Cząsteczka złożona z kwasu nukleinowego (DNA
47 Olimpiada Biologiczna
 47 Olimpiada Biologiczna Pracownia biochemiczna zasady oceniania rozwia zan zadan Część A. Identyfikacja zawartości trzech probówek zawierających cukry (0 21 pkt) Zadanie A.1 (0 13 pkt) Tabela 1 (0 7 pkt)
47 Olimpiada Biologiczna Pracownia biochemiczna zasady oceniania rozwia zan zadan Część A. Identyfikacja zawartości trzech probówek zawierających cukry (0 21 pkt) Zadanie A.1 (0 13 pkt) Tabela 1 (0 7 pkt)
Warszawa, dnia 3 sierpnia 2016 r. Poz. 1173
 Warszawa, dnia 3 sierpnia 2016 r. Poz. 1173 ROZPORZĄDZENIE MINISTRA ŚRODOWISKA 1) z dnia 18 lipca 2016 r. w sprawie określenia wzorów wniosków oraz zgłoszeń związanych z zamkniętym użyciem mikroorganizmów
Warszawa, dnia 3 sierpnia 2016 r. Poz. 1173 ROZPORZĄDZENIE MINISTRA ŚRODOWISKA 1) z dnia 18 lipca 2016 r. w sprawie określenia wzorów wniosków oraz zgłoszeń związanych z zamkniętym użyciem mikroorganizmów
Techniki molekularne w mikrobiologii SYLABUS A. Informacje ogólne
 Techniki molekularne w mikrobiologii A. Informacje ogólne Elementy sylabusu Nazwa jednostki prowadzącej kierunek Nazwa kierunku studiów Poziom kształcenia Profil studiów Forma studiów Rodzaj Rok studiów
Techniki molekularne w mikrobiologii A. Informacje ogólne Elementy sylabusu Nazwa jednostki prowadzącej kierunek Nazwa kierunku studiów Poziom kształcenia Profil studiów Forma studiów Rodzaj Rok studiów
Biologia Molekularna Podstawy
 Biologia Molekularna Podstawy Budowa DNA Budowa DNA Zasady: Purynowe: adenina i guanina Pirymidynowe: cytozyna i tymina 2 -deoksyryboza Grupy fosforanowe Budowa RNA Budowa RNA Zasady: purynowe: adenina
Biologia Molekularna Podstawy Budowa DNA Budowa DNA Zasady: Purynowe: adenina i guanina Pirymidynowe: cytozyna i tymina 2 -deoksyryboza Grupy fosforanowe Budowa RNA Budowa RNA Zasady: purynowe: adenina
Biologia Molekularna z Biotechnologią ===============================================================================================
 Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 5 Droga od genu do
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 5 Droga od genu do
PROJEKT WSPÓŁFINANSOWANY PRZEZ UNIĘ EUROPEJSKĄ Z EUROPEJSKIEGO FUNDUSZU ROZWOJU REGIONALNEGO 1 z 7
 Poznań, dnia 28.04.2014 r. BioVentures Institute Spółka z ograniczoną odpowiedzialnością ul. Promienista 83 60 141 Poznań Zapytanie ofertowe nr 01/2014 Projekt Nowa technologia wytwarzania szczepionek
Poznań, dnia 28.04.2014 r. BioVentures Institute Spółka z ograniczoną odpowiedzialnością ul. Promienista 83 60 141 Poznań Zapytanie ofertowe nr 01/2014 Projekt Nowa technologia wytwarzania szczepionek
października 2013: Elementarz biologii molekularnej. Wykład nr 2 BIOINFORMATYKA rok II
 10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
S YL AB US MODUŁ U ( PRZEDMIOTU) I nforma c j e ogólne
 I nforma c j e ogólne") Załącznik Nr 3 do Uchwały Senatu PUM 14/2012 S YL AB US MODUŁ U ( PRZEDMIOTU) I nforma c j e ogólne Kod modułu Rodzaj modułu Wydział PUM Kierunek studiów Specjalność Poziom studiów Nazwa modułu INŻYNIERIA
Załącznik Nr 3 do Uchwały Senatu PUM 14/2012 S YL AB US MODUŁ U ( PRZEDMIOTU) I nforma c j e ogólne Kod modułu Rodzaj modułu Wydział PUM Kierunek studiów Specjalność Poziom studiów Nazwa modułu INŻYNIERIA
DNA musi współdziałać z białkami!
 DNA musi współdziałać z białkami! Specyficzność oddziaływań między DNA a białkami wiążącymi DNA zależy od: zmian konformacyjnych wzdłuż cząsteczki DNA zróżnicowania struktury DNA wynikającego z sekwencji
DNA musi współdziałać z białkami! Specyficzność oddziaływań między DNA a białkami wiążącymi DNA zależy od: zmian konformacyjnych wzdłuż cząsteczki DNA zróżnicowania struktury DNA wynikającego z sekwencji
ĆWICZENIE I BADANIE WRAŻLIWOŚCI BAKTERII NA ANTYBIOTYKI I CHEMIOTERAPEUTYKI
 ĆWICZENIE I BADANIE WRAŻLIWOŚCI BAKTERII NA ANTYBIOTYKI I CHEMIOTERAPEUTYKI Autor i główny prowadzący dr Dorota Korsak Wstęp Jednym z najważniejszych etapów rutynowej diagnostyki mikrobiologicznej zakażeń
ĆWICZENIE I BADANIE WRAŻLIWOŚCI BAKTERII NA ANTYBIOTYKI I CHEMIOTERAPEUTYKI Autor i główny prowadzący dr Dorota Korsak Wstęp Jednym z najważniejszych etapów rutynowej diagnostyki mikrobiologicznej zakażeń
WPROWADZENIE DO GENETYKI MOLEKULARNEJ
 WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
Bezpośrednia embriogeneza somatyczna
 Bezpośrednia embriogeneza somatyczna Zarodki somatyczne formują się bezpośrednio tylko z tych komórek roślinnych, które są kompetentne już w momencie izolowania z rośliny macierzystej, czyli z proembriogenicznie
Bezpośrednia embriogeneza somatyczna Zarodki somatyczne formują się bezpośrednio tylko z tych komórek roślinnych, które są kompetentne już w momencie izolowania z rośliny macierzystej, czyli z proembriogenicznie
Hybrydyzacja kwasów nukleinowych
 Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
Making the impossible possible: the metamorphosis of Polish Biology Olympiad
 Making the impossible possible: the metamorphosis of Polish Biology Olympiad Takao Ishikawa Faculty of Biology, University of Warsaw, Poland Performance of Polish students at IBO Gold Silver Bronze Merit
Making the impossible possible: the metamorphosis of Polish Biology Olympiad Takao Ishikawa Faculty of Biology, University of Warsaw, Poland Performance of Polish students at IBO Gold Silver Bronze Merit
starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg
 STRESZCZENIE Przewlekła białaczka limfocytowa (PBL) jest najczęstszą białaczką ludzi starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg kliniczny, zróżnicowane rokowanie. Etiologia
STRESZCZENIE Przewlekła białaczka limfocytowa (PBL) jest najczęstszą białaczką ludzi starszych na półkuli zachodniej. Typową cechą choroby jest heterogenny przebieg kliniczny, zróżnicowane rokowanie. Etiologia
Klonowanie i transgeneza. dr n.med. Katarzyna Wicher
 Klonowanie i transgeneza dr n.med. Katarzyna Wicher Klonowanie proces tworzenia idealnej kopii z oryginału oznacza powstawanie lub otrzymywanie genetycznie identycznych: cząsteczek DNA komórek organizmów
Klonowanie i transgeneza dr n.med. Katarzyna Wicher Klonowanie proces tworzenia idealnej kopii z oryginału oznacza powstawanie lub otrzymywanie genetycznie identycznych: cząsteczek DNA komórek organizmów
SYLABUS. Wydział Biologiczno-Rolniczy. Katedra Biochemii i Biologii Komórki
 SYLABUS 1.1. PODSTAWOWE INFORMACJE O PRZEDMIOCIE/MODULE Nazwa przedmiotu/ modułu Biologia molekularna z elementami inżynierii genetycznej Kod przedmiotu/ modułu* Wydział (nazwa jednostki prowadzącej kierunek)
SYLABUS 1.1. PODSTAWOWE INFORMACJE O PRZEDMIOCIE/MODULE Nazwa przedmiotu/ modułu Biologia molekularna z elementami inżynierii genetycznej Kod przedmiotu/ modułu* Wydział (nazwa jednostki prowadzącej kierunek)
WYKRYWANIE OBECNOŚCI BAKTERII Z RODZAJU LISTERIA W ŻYWNOŚCI
 ĆWICZENIE IV Autor i główny prowadzący dr Dorota Korsak WYKRYWANIE OBECNOŚCI BAKTERII Z RODZAJU LISTERIA W ŻYWNOŚCI Wstęp Rodzaj Listeria należy do typu Firmicutes wspólnie z rodzajem Staphylococcus, Streptococcus,
ĆWICZENIE IV Autor i główny prowadzący dr Dorota Korsak WYKRYWANIE OBECNOŚCI BAKTERII Z RODZAJU LISTERIA W ŻYWNOŚCI Wstęp Rodzaj Listeria należy do typu Firmicutes wspólnie z rodzajem Staphylococcus, Streptococcus,
WNIOSEK O WYDANIE ZGODY NA ZAMKNIĘTE UŻYCIE GMO
 WNIOSEK O WYDANIE ZGODY NA ZAMKNIĘTE UŻYCIE GMO 1. Informacje o użytkowniku GMO i osobach odpowiedzialnych za realizację planowanego zamkniętego użycia GMO 1.1 (*) Nazwa i siedziba użytkownika lub imię,
WNIOSEK O WYDANIE ZGODY NA ZAMKNIĘTE UŻYCIE GMO 1. Informacje o użytkowniku GMO i osobach odpowiedzialnych za realizację planowanego zamkniętego użycia GMO 1.1 (*) Nazwa i siedziba użytkownika lub imię,
1276:1997 13368 (ATCC
 Działanie bakteriobójcze Środka biobójczego Clinell GAMA Healthcare Ltd. zbadane za pomocą Europejskiego Standardowego Testu według normy BS EN 1276:1997 wobec: Klebsiella pneumoniae NCTC 13368 (ATCC 700603).
Działanie bakteriobójcze Środka biobójczego Clinell GAMA Healthcare Ltd. zbadane za pomocą Europejskiego Standardowego Testu według normy BS EN 1276:1997 wobec: Klebsiella pneumoniae NCTC 13368 (ATCC 700603).
Technika hodowli komórek leukemicznych
 Fizjologiczne Techniki Badań Technika hodowli komórek leukemicznych Zasady prowadzenia hodowli komórek leukemicznych, pasażowania, mrożenia, rozmrażania i przechowywania komórek leukemicznych Warunki wstępne:
Fizjologiczne Techniki Badań Technika hodowli komórek leukemicznych Zasady prowadzenia hodowli komórek leukemicznych, pasażowania, mrożenia, rozmrażania i przechowywania komórek leukemicznych Warunki wstępne:
Zastosowanie metody RAPD do różnicowania szczepów bakteryjnych
 Zastosowanie metody RAPD do różnicowania szczepów bakteryjnych Wstęp teoretyczny Technika RAPD (ang. Random Amplification of Polymorphic DNA) opiera się na prostej reakcji PCR, przeprowadzanej na genomowym
Zastosowanie metody RAPD do różnicowania szczepów bakteryjnych Wstęp teoretyczny Technika RAPD (ang. Random Amplification of Polymorphic DNA) opiera się na prostej reakcji PCR, przeprowadzanej na genomowym
Konstrukcja wektora plazmidowego DNA do klonowania genów i/lub wektora plazmidowego do sekrecji w bakteriach mlekowych
 Konstrukcja wektora plazmidowego DNA do klonowania genów i/lub wektora plazmidowego do sekrecji w bakteriach mlekowych Łukasz Tranda Promotor: doc. dr hab. Jacek Bardowski, IBB Promotor: dr hab. Edward
Konstrukcja wektora plazmidowego DNA do klonowania genów i/lub wektora plazmidowego do sekrecji w bakteriach mlekowych Łukasz Tranda Promotor: doc. dr hab. Jacek Bardowski, IBB Promotor: dr hab. Edward
StayRNA bufor zabezpieczający RNA przed degradacją wersja 0215
 StayRNA bufor zabezpieczający RNA przed degradacją wersja 0215 100 ml, 250 ml, 500 ml Nr kat. 038-100, 038-250, 038-500 Nietosyczny roztwór wodny do przechowywania i zabezpieczania różnego rodzaju tkanek
StayRNA bufor zabezpieczający RNA przed degradacją wersja 0215 100 ml, 250 ml, 500 ml Nr kat. 038-100, 038-250, 038-500 Nietosyczny roztwór wodny do przechowywania i zabezpieczania różnego rodzaju tkanek
Sylabus Biologia molekularna
 Sylabus Biologia molekularna 1. Metryczka Nazwa Wydziału Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Program kształcenia Farmacja, jednolite studia magisterskie, forma studiów: stacjonarne
Sylabus Biologia molekularna 1. Metryczka Nazwa Wydziału Wydział Farmaceutyczny z Oddziałem Medycyny Laboratoryjnej Program kształcenia Farmacja, jednolite studia magisterskie, forma studiów: stacjonarne
TATA box. Enhancery. CGCG ekson intron ekson intron ekson CZĘŚĆ KODUJĄCA GENU TERMINATOR. Elementy regulatorowe
 Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Przeglądanie bibliotek
 Przeglądanie bibliotek Czyli jak złapać (i sklonować) ciekawy gen? Klonowanie genów w oparciu o identyczność lub podobieństwo ich sekwencji do znanego już DNA Sonda homologiczna (komplementarna w 100%)
Przeglądanie bibliotek Czyli jak złapać (i sklonować) ciekawy gen? Klonowanie genów w oparciu o identyczność lub podobieństwo ich sekwencji do znanego już DNA Sonda homologiczna (komplementarna w 100%)
PL 217144 B1. Sposób amplifikacji DNA w łańcuchowej reakcji polimerazy za pomocą starterów specyficznych dla genu receptora 2-adrenergicznego
 PL 217144 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 217144 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 391926 (22) Data zgłoszenia: 23.07.2010 (51) Int.Cl.
PL 217144 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 217144 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 391926 (22) Data zgłoszenia: 23.07.2010 (51) Int.Cl.
Oznaczenie polimorfizmu genetycznego cytochromu CYP2D6: wykrywanie liczby kopii genu
 Ćwiczenie 4 Oznaczenie polimorfizmu genetycznego cytochromu CYP2D6: wykrywanie liczby kopii genu Wstęp CYP2D6 kodowany przez gen występujący w co najmniej w 78 allelicznych formach związanych ze zmniejszoną
Ćwiczenie 4 Oznaczenie polimorfizmu genetycznego cytochromu CYP2D6: wykrywanie liczby kopii genu Wstęp CYP2D6 kodowany przez gen występujący w co najmniej w 78 allelicznych formach związanych ze zmniejszoną
Genomic Maxi AX zestaw do izolacji genomowego DNA wersja 0616
 Genomic Maxi AX zestaw do izolacji genomowego DNA wersja 0616 10 izolacji Nr kat. 995-10 Pojemność kolumny do oczyszczania DNA wynosi 500 µg 1 Skład zestawu Składnik Ilość Temp. Przechowywania Kolumny
Genomic Maxi AX zestaw do izolacji genomowego DNA wersja 0616 10 izolacji Nr kat. 995-10 Pojemność kolumny do oczyszczania DNA wynosi 500 µg 1 Skład zestawu Składnik Ilość Temp. Przechowywania Kolumny
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych
 Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
ĆWICZENIE 5 MECHANIZMY PROMUJĄCE WZROST ROŚLIN
 ĆWICZENIE 5 MECHANIZMY PROMUJĄCE WZROST ROŚLIN CZĘŚĆ TEORETYCZNA Mechanizmy promujące wzrost rośli (PGP) Metody badań PGP CZĘŚĆ PRAKTYCZNA 1. Mechanizmy promujące wzrost roślin. Odczyt. a) Wytwarzanie
ĆWICZENIE 5 MECHANIZMY PROMUJĄCE WZROST ROŚLIN CZĘŚĆ TEORETYCZNA Mechanizmy promujące wzrost rośli (PGP) Metody badań PGP CZĘŚĆ PRAKTYCZNA 1. Mechanizmy promujące wzrost roślin. Odczyt. a) Wytwarzanie
Raport z badania Działanie wirusobójcze środka dezynfekującego wobec Feline calicivirus. 25 października 2006
 Raport z badania Działanie wirusobójcze środka dezynfekującego wobec Feline calicivirus 25 października 2006 Dr Tobias J. Tuthill Wydział Nauk Biologicznych Uniwersytet Leeds Leeds LS2 9JT www.fbs.leeds.ac.uk
Raport z badania Działanie wirusobójcze środka dezynfekującego wobec Feline calicivirus 25 października 2006 Dr Tobias J. Tuthill Wydział Nauk Biologicznych Uniwersytet Leeds Leeds LS2 9JT www.fbs.leeds.ac.uk
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach
 FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach FOCUS Plus to dodatek dostępny dla standardowych pasz tuczowych BioMaru, dostosowany specjalnie do potrzeb ryb narażonych na trudne
FOCUS Plus - Silniejsza ryba radzi sobie lepiej w trudnych warunkach FOCUS Plus to dodatek dostępny dla standardowych pasz tuczowych BioMaru, dostosowany specjalnie do potrzeb ryb narażonych na trudne
Analizy wielkoskalowe w badaniach chromatyny
 Analizy wielkoskalowe w badaniach chromatyny Analizy wielkoskalowe wykorzystujące mikromacierze DNA Genotypowanie: zróżnicowane wewnątrz genów RNA Komórka eukariotyczna Ekspresja genów: Które geny? Poziom
Analizy wielkoskalowe w badaniach chromatyny Analizy wielkoskalowe wykorzystujące mikromacierze DNA Genotypowanie: zróżnicowane wewnątrz genów RNA Komórka eukariotyczna Ekspresja genów: Które geny? Poziom
Metody inżynierii genetycznej SYLABUS A. Informacje ogólne
 Metody inżynierii genetycznej A. Informacje ogólne Elementy sylabusu Nazwa jednostki prowadzącej kierunek Nazwa kierunku studiów Poziom kształcenia Profil studiów Forma studiów Kod Rodzaj Rok studiów /semestr
Metody inżynierii genetycznej A. Informacje ogólne Elementy sylabusu Nazwa jednostki prowadzącej kierunek Nazwa kierunku studiów Poziom kształcenia Profil studiów Forma studiów Kod Rodzaj Rok studiów /semestr
Rozmnażanie i wzrost komórek sąściśle kontrolowane. Genetyczne podłoże nowotworzenia
 Rozmnażanie i wzrost komórek sąściśle kontrolowane Genetyczne podłoże nowotworzenia Rozmnażanie i wzrost komórek sąściśle kontrolowane Rozmnażanie i wzrost komórek sąściśle kontrolowane Połączenia komórek
Rozmnażanie i wzrost komórek sąściśle kontrolowane Genetyczne podłoże nowotworzenia Rozmnażanie i wzrost komórek sąściśle kontrolowane Rozmnażanie i wzrost komórek sąściśle kontrolowane Połączenia komórek
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2764103 (96) Data i numer zgłoszenia patentu europejskiego: 12.12.2013 13824232.6 (13) (51) T3 Int.Cl. C12N 15/63 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2764103 (96) Data i numer zgłoszenia patentu europejskiego: 12.12.2013 13824232.6 (13) (51) T3 Int.Cl. C12N 15/63 (2006.01)
Rozkład materiału z biologii dla klasy III AD. 7 godz / tyg rok szkolny 2016/17
 Rozkład materiału z biologii dla klasy III AD zakres rozszerzony LO 7 godz / tyg rok szkolny 2016/17 Biologia na czasie 2 zakres rozszerzony nr dopuszczenia 564/2/2012 Biologia na czasie 3 zakres rozszerzony
Rozkład materiału z biologii dla klasy III AD zakres rozszerzony LO 7 godz / tyg rok szkolny 2016/17 Biologia na czasie 2 zakres rozszerzony nr dopuszczenia 564/2/2012 Biologia na czasie 3 zakres rozszerzony
RAPORT Z BADAŃ 164/Z/20110825/D/JOGA. Dostarczony materiał: próbki tworzyw sztucznych. Ilość próbek: 1. Rodzaj próbek: tworzywo
 Blirt S.A. 80-172 Gdańsk, ul. Trzy Lipy 3/1.38 RAPORT Z BADAŃ Dział DNA-Gdańsk Nr zlecenia 164/Z/20110825/D/JOGA NAZWA I ADRES KLIENTA GROUND-Therm spółka z o.o. ul. Stepowa 30 44-105 Gliwice Tytuł zlecenia:
Blirt S.A. 80-172 Gdańsk, ul. Trzy Lipy 3/1.38 RAPORT Z BADAŃ Dział DNA-Gdańsk Nr zlecenia 164/Z/20110825/D/JOGA NAZWA I ADRES KLIENTA GROUND-Therm spółka z o.o. ul. Stepowa 30 44-105 Gliwice Tytuł zlecenia:
Informacje dotyczące pracy kontrolnej
 Informacje dotyczące pracy kontrolnej Słuchacze, którzy z przyczyn usprawiedliwionych nie przystąpili do pracy kontrolnej lub otrzymali z niej ocenę negatywną zobowiązani są do dnia 06 grudnia 2015 r.
Informacje dotyczące pracy kontrolnej Słuchacze, którzy z przyczyn usprawiedliwionych nie przystąpili do pracy kontrolnej lub otrzymali z niej ocenę negatywną zobowiązani są do dnia 06 grudnia 2015 r.
(86) Data i numer zgłoszenia międzynarodowego: , PCT/US99/21960 (87) Data i numer publikacji zgłoszenia międzynarodowego:
 Data i numer zgłoszenia międzynarodowego: , PCT/US99/21960 (87) Data i numer publikacji zgłoszenia międzynarodowego:") RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 197408 (21) Numer zgłoszenia: 346772 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 21.09.1999 (86) Data i numer zgłoszenia
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 197408 (21) Numer zgłoszenia: 346772 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 21.09.1999 (86) Data i numer zgłoszenia
WYKŁAD: Klasyczny przepływ informacji ( Dogmat) Klasyczny przepływ informacji. Ekspresja genów realizacja informacji zawartej w genach
 Klasyczny przepływ informacji. Ekspresja genów realizacja informacji zawartej w genach") WYKŁAD: Ekspresja genów realizacja informacji zawartej w genach Prof. hab. n. med. Małgorzata Milkiewicz Zakład Biologii Medycznej Klasyczny przepływ informacji ( Dogmat) Białka Retrowirusy Białka Klasyczny
WYKŁAD: Ekspresja genów realizacja informacji zawartej w genach Prof. hab. n. med. Małgorzata Milkiewicz Zakład Biologii Medycznej Klasyczny przepływ informacji ( Dogmat) Białka Retrowirusy Białka Klasyczny
AmpliTest Babesia spp. (PCR)
") AmpliTest Babesia spp. (PCR) Zestaw do wykrywania sekwencji DNA specyficznych dla pierwotniaków z rodzaju Babesia techniką PCR Nr kat.: BAC21-100 Wielkość zestawu: 100 oznaczeń Objętość pojedynczej reakcji:
AmpliTest Babesia spp. (PCR) Zestaw do wykrywania sekwencji DNA specyficznych dla pierwotniaków z rodzaju Babesia techniką PCR Nr kat.: BAC21-100 Wielkość zestawu: 100 oznaczeń Objętość pojedynczej reakcji:
Definicje. Białka rekombinowane (ang. recombinant proteins, r-proteins) Ukierunkowana mutageneza (ang. site-directed/site-specific mutagenesis)
 Ukierunkowana mutageneza (ang. site-directed/site-specific mutagenesis)") Definicje Białka rekombinowane (ang. recombinant proteins, r-proteins) Białka, które powstały w żywych organizmach (lub liniach komórkowych) w wyniku ekspresji rekombinowanego DNA. Rekombinowany DNA jest
Definicje Białka rekombinowane (ang. recombinant proteins, r-proteins) Białka, które powstały w żywych organizmach (lub liniach komórkowych) w wyniku ekspresji rekombinowanego DNA. Rekombinowany DNA jest
PL B1. UNIWERSYTET PRZYRODNICZY W LUBLINIE, Lublin, PL BUP 04/14. SYLWIA OKOŃ, Dąbrowica, PL KRZYSZTOF KOWALCZYK, Motycz, PL
 PL 220315 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 220315 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 400252 (22) Data zgłoszenia: 06.08.2012 (51) Int.Cl.
PL 220315 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 220315 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 400252 (22) Data zgłoszenia: 06.08.2012 (51) Int.Cl.
Transformacja pośrednia składa się z trzech etapów:
 Transformacja pośrednia składa się z trzech etapów: 1. Otrzymanie pożądanego odcinka DNA z materiału genetycznego dawcy 2. Wprowadzenie obcego DNA do wektora 3. Wprowadzenie wektora, niosącego w sobie
Transformacja pośrednia składa się z trzech etapów: 1. Otrzymanie pożądanego odcinka DNA z materiału genetycznego dawcy 2. Wprowadzenie obcego DNA do wektora 3. Wprowadzenie wektora, niosącego w sobie
SPECYFIKACJA TECHNICZNA AGAROZ
 SPECYFIKACJA TECHNICZNA AGAROZ D1 LOW EEO i D1 MEDIUM EEO, D1 HIGH EEO D1 LOW EEO GQT; (Genetic Quality Tested) D2 HIGH GELLING TEMPERATURE D5 HIGH STRENGTH GEL Agaroza LM Agaroza LM GQT; (Genetic Quality
SPECYFIKACJA TECHNICZNA AGAROZ D1 LOW EEO i D1 MEDIUM EEO, D1 HIGH EEO D1 LOW EEO GQT; (Genetic Quality Tested) D2 HIGH GELLING TEMPERATURE D5 HIGH STRENGTH GEL Agaroza LM Agaroza LM GQT; (Genetic Quality
BIOLOGIA KOMÓRKI. Mikroskopia fluorescencyjna -2 Przyżyciowe barwienia organelli wewnątrzkomórkowych
 BIOLOGIA KOMÓRKI Mikroskopia fluorescencyjna -2 Przyżyciowe barwienia organelli Wstęp Komórki eukariotyczne, w odróżnieniu od komórek prokariotycznych (bakterie, archeony) posiadają wysoce skomplikowaną
BIOLOGIA KOMÓRKI Mikroskopia fluorescencyjna -2 Przyżyciowe barwienia organelli Wstęp Komórki eukariotyczne, w odróżnieniu od komórek prokariotycznych (bakterie, archeony) posiadają wysoce skomplikowaną
Co to jest transkryptom? A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH 2
 ALEKSANDRA ŚWIERCZ Co to jest transkryptom? A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH 2 Ekspresja genów http://genome.wellcome.ac.uk/doc_wtd020757.html A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH
ALEKSANDRA ŚWIERCZ Co to jest transkryptom? A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH 2 Ekspresja genów http://genome.wellcome.ac.uk/doc_wtd020757.html A. Świercz ANALIZA DANYCH WYSOKOPRZEPUSTOWYCH
ROZPORZĄDZENIE KOMISJI (UE) / z dnia r.
 / z dnia r.") KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
Hybrydyzacja kwasów nukleinowych
 Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja tego genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja tego genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
ĆWICZENIE 1 i 2 Modyfikacja geu wołowej beta-laktoglobuliny przy użyciu metody Overlap Extension PCR (wydłużania nakładających się odcinków)
") ĆWICZENIE 1 i 2 Modyfikacja geu wołowej beta-laktoglobuliny przy użyciu metody Overlap Extension PCR (wydłużania nakładających się odcinków) Celem ćwiczenia jest wprowadzenie mutacji punktowej do genu
ĆWICZENIE 1 i 2 Modyfikacja geu wołowej beta-laktoglobuliny przy użyciu metody Overlap Extension PCR (wydłużania nakładających się odcinków) Celem ćwiczenia jest wprowadzenie mutacji punktowej do genu