PL B1. UNIWERSYTET JAGIELLOŃSKI, Kraków, PL BIOCENTRUM SPÓŁKA Z OGRANICZONĄ ODPOWIEDZIALNOŚCIĄ, Kraków, PL
|
|
|
- Małgorzata Kowal
- 6 lat temu
- Przeglądów:
Transkrypt
 Numer zgłoszenia, z którego nastąpiło wydzielenie: 382638 (13) B1 (51) Int.Cl. C12N 9/52 (2006.01) C07H 21/04 (2006.01) C40B 10/00 (2006.")
1 PL B1 RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) (21) Numer zgłoszenia: (22) Data zgłoszenia: (62) Numer zgłoszenia, z którego nastąpiło wydzielenie: (13) B1 (51) Int.Cl. C12N 9/52 ( ) C07H 21/04 ( ) C40B 10/00 ( ) (54) Proteinaza posiadająca aktywność proteinazy SpIB (43) Zgłoszenie ogłoszono: BUP 26/11 (45) O udzieleniu patentu ogłoszono: WUP 06/13 (73) Uprawniony z patentu: UNIWERSYTET JAGIELLOŃSKI, Kraków, PL BIOCENTRUM SPÓŁKA Z OGRANICZONĄ ODPOWIEDZIALNOŚCIĄ, Kraków, PL (72) Twórca(y) wynalazku: GRZEGORZ DUBIN, Kraków, PL JAN POTEMPA, Kraków, PL (74) Pełnomocnik: rzecz. pat. Anna Rożkowicz
2 2 PL B1 Opis wynalazku Wynalazek dotyczy proteinazy posiadającej aktywność proteinazy SplB. Ujawniono metodę otrzymywania proteinazy SplB, jej zastosowania do specyficznej hydrolizy łańcucha polipeptydowego, sekwencji aminokwasowych przez nią rozpoznawanych oraz ich zastosowań. Enzymy proteolityczne (proteinazy) o wysokiej specyficzności działania (rozpoznające i hydrolizujące jedynie wybrane wiązania peptydowe) są stosowane na szeroką skalę w laboratoriach i przemyśle biotechnologicznym do specyficznej hydrolizy polipeptydów (przede wszystkim następujące: enterokinaza, czynnik X, trombina, proteinaza TEV, proteinaza PreScission oraz o mniejszej specyficzności, ale też szeroko wykorzystywane jak V8 i trypsyna). W szczególności, lecz nie jed y- nie, stosuje się omawiane enzymy do usuwania tzw. metek fuzyjnych, fragmentów polipeptydów rekombinowanych użytecznych na pośrednich etapach analizy lub produkcji (służących np. do detekcji, oczyszczania) jednak niepożądanych w produkcie końcowym. W teorii wysoka specyficzność zastosowanego enzymu w połączeniu z wydajnie rozpoznawanym przez niego miejscem wprowadzonym pomiędzy metką" a częścią polipeptydu stanowiącą ostatecznie pożądany produkt pozwala na precyzyjne usunięcie metki ' bez ryzyka degradacji pożądanego polipeptydu. Niestety brak ca ł- kowicie specyficznych enzymów proteolitycznych powoduje, że w wielu przypadkach rezultatem ich działania jest nie tylko pożądane odcięcie "metki fuzyjnej, ale także niespecyficzna degradacja interesującego polipeptydu, jeśli zawiera on miejsca podobne do sekwencji specyficznie rozpozn a- wanej przez enzym. Ponadto najpopularniejsze ze stosowanych obecnie enzymów nie są dostępne w postaci białek zrekombinowanych lub z uwagi na trudności w produkcji są w tej formie znacznie droższe od białek natywnych izolowanych z osocza krwi (trombina, czynnik X) lub jelit (enterokin a- za). Zastosowanie białek natywnych stwarza jednak ryzyko zanieczyszczenia preparatów niepożądaną aktywnością innych enzymów lub patogenami. Wymienione czynniki stwarzają zapotrzebow a- nie na nowe enzymy, które będą mogły sprostać specyficznym zadaniom. Szczególnie pożądane jest uzyskanie proteinaz o wąskiej specyficzności substratowej, które mogłyby znaleźć zastosowanie jako precyzyjne narzędzie biotechnologiczne (przykładowe opisy patentowe: US , US , US , US ). Sekwencja aminokwasowa proteinazy SplB pochodzącej ze Staphylococcus aureus jest znana i została opisana w publikacji J. Mol. Biol. (2006) 358, W pracy tej ujawniono także mało wydajną, niewygodną, laboratoryjną metodę produkcji rekombinowanej proteinazy SplB poprzez ekspresję w E. coli oraz wykazano aktywność proteolityczną na kazeinie powyższego preparatu metodą zymografii. Nie była jednak poznana specyficzność substratowa tego enzymu ani w y- dajny sposób jego otrzymywania. Ponadto, znane jest wiele proteinaz ser ynowych chymotrypsynopodobnych (w przypadku podobieństw strukturalnych określenie chymotrypsynopodobny odnosi się do tej samej grupy proteinaz co określenie trypsynopodobny i są one tutaj stosowane zamiennie; jedynie w przypadku określania typów aktywności określenia te są rozdzielne jednak jako takie nie są używane w opisie), do których należy proteinaza SplB (MEROPS: S01.282). Podobieństwo sekwencji aminokwasowej do innych proteinaz z tej grupy pozwoliła zaliczyć proteinazę SplB do rodziny S1 wg ogólnie przyjętej klasyfikacji za bazą danych MEROPS ( Rawlings, N.D., Morton, F.R. & Barrett, A.J. (2006) MEROPS: the peptidase database. Nucleic Acids Res 34, D270-D272). Zgodnie z powszechnym przeświadczeniem wyrażonym m.in. w wiodącej referencji z dziedziny enzymów proteolitycznych, bazie danych MEROPS uznaje się, że: Wszystkie scharakteryzowane peptydazy należące do rodziny chymotrypsynopodobnych są endopeptydaz a- mi. Istnieją także liczne, nie będące peptydazami homologii, w których reszty katalityczne zostały zastąpione. Istnieją trzy główne typy aktywności: trypsynopodobny, w którym następuje odtrawienie substratu amidowego następującego po resztach Arg lub Lys w pozycji P1, chymotrypsynopodobny, w którym trawienie następuje za jednym z aminokwasów hydrofobowych w P1 i elastazopodobny, w którym trawienie następuje za resztą Ala w pozycji P1. Specyficzność substratowa rodziny S1 zależy jedynie od aminokwasu znajdującego się w pozycji P1. Większość peptydaz należących do tej rodziny podlega sekrecji i posiada N-końcowy sekrecyjny peptyd sygnałowy. Są one syntetyzowane w postaci prekursorów z dodatkową sekwencją na N-końcu, której usunięcie daje aktywną formę enzymu. Aktywacja nie zawsze wymaga usunięcia propeptydu. Jak pokazano w dalszej części niniejszego opisu ogólne wskazówki zawarte w stanie techniki mogą prowadzić jedynie do błę d- nych wniosków dotyczących specyficzności substratowej proteinazy SplB i uznania ją za enzym o nikłej przydatności przemysłowej.
3 PL B1 3 W świetle opisanego stanu techniki celem niniejszego wynalazku jest dostarczenie wysoce specyficznej proteinazy oraz sposobu jej otrzymywania oraz charakterystyki jej aktywności pozwalającej na jej przemysłowe wykorzystywanie. Nieoczekiwanie twórcy tego wynalazku ustalili, że proteinaza SplB posiada dużo węższą niż oczekiwana specyficzność substratową. Bazując na tym odkryciu zaproponowano nowe specyficzne substraty dla proteinazy SplB oraz sposoby hydrolizy i/lub otrzymywania białek wykorzystujące takie peptydy (fragmenty sekwencji) oraz nowe zastosowania proteinazy SplB. Uzyskanie tych wyników było możliwe dzięki opracowaniu wydajnej metody produkcji proteinazy SplB, którą ujawniono w niniejszym opisie. W opisie ujawniono polipeptyd wykazujący powinowactwo do centrum aktywnego proteinazy SplB posiadający sekwencję aminokwasową Xaa1-Xaa2-Xaa3-Xaa4-Xaa5, gdzie: Xaa1 jest aminokwasem wybranym spośród: Trp, Ala, Ile, Leu, Met, Phe, Tyr, Val, Ser, Thr lub Gly, Xaa2 jest aminokwasem wybranym spośród Glu, Gln, Asp, Asn, Val, Leu, He, Gly, Arg, Ser lub Thr, Xaa3 jest aminokwasem wybranym spośród Leu, Ile, Val, Thr, Ser lub Gly. Xaa4 jest aminokwasem wybranym spośród: Gln, Glu, Thr, Ser, Asp lub Asn, Xaa5 jest pominięty lub jest dowolnym aminokwasem. Ujawniony polipeptyd charakteryzuje się tym, że korzystnie posiada sekwencję wybraną spośród: Trp-Glu-Leu-Gln-Gly, Trp-Glu-Leu-Gln, Trp-Glu-Leu-Thr, Trp-Glu-Val-Gln, Val-Glu-Leu-Gln, Trp-Gln-Leu-Asp, Trp-Val-Leu-Gln, Phe-Glu-Val-Glu, Gly-Arg-Gly-Val-Gly, Gly-Arg-Gly-Val, Val-Glu-Ile-Asp. Kolejno w niniejszym opisie ujawniono białko rozpoznawane przez proteinazę SplB posiadające sekwencję aminokwasową zawierającą zdefiniowany powyżej polipeptyd. Następnie ujawniono sekwencję nukleotydową kodująca ujawniony polipeptyd zdefiniowany powyżej oraz sekwencję nukleotydową kodująca ujawnione białko zdefiniowane powyżej. Ujawniono również zastosowanie sekwencji polipeptydu zdefiniowanej powyżej lub jego pochodnej przy wytwarzaniu białka rozpoznawanego przez proteinazę SplB lub jej pochodną. Ujawniono kolejno sposób otrzymywania pożądanego białka charakteryzujący się tym, że: a) dostarcza się białko fuzyjne posiadające sekwencję Z1-Xaa1-Xaa2-Xaa3-Xaa4-Z2, gdzie: Xaa1 jest aminokwasem wybranym spośród: Trp, Ala, Ile, Leu, Met, Phe, Tyr, Val, Ser, Thr lub Gly, Xaa2 jest aminokwasem wybranym spośród Glu, Gln, Asp, Asn, Val, Leu, Ile, Gly, Arg, Ser lub Thr, Xaa3 jest aminokwasem wybranym spośród Leu, Ile, Val, Thr, Ser lub Gly. Xaa4 jest aminokwasem wybranym spośród: Gln, Glu, Thr, Ser, Asp lub Asn, Z1 i Z2 oznacza polipeptyd zawierający jeden lub więcej aminokwasów, przy czym jeden z nich oznacza polipeptyd zawierający pożądane białko a drugi polipeptyd zawierający polipeptyd znacznikowy, b) izoluje się białko fuzyjne, korzystnie techniką chromatograficzną stosując złoże posiadające powinowactwo do polipeptydu znacznikowego. c) prowadzi się reakcję hydrolizy białka fuzyjnego za pomocą proteinazy posiadającej aktywność enzymatyczną proteinazy SplB, przy czym korzystnie izoluje się pożądane białko z mieszaniny reakcyjnej. Dla celów niniejszego opisu jako polipeptyd zawierający polipeptyd znacznikowy, zwany też w niniejszym opisie metką lub polipeptydem znacznikowym, należy rozumieć sekwencję pozwalającą na izolowanie zawierającego ją polipeptydu, zwłaszcza techniką chromatografii powinowactwa.
4 4 PL B1 Specjalista będzie w stanie zaproponować opierając się na powszechnie dostępnej wiedzy szereg tego rodzaju sekwencji, które można wykorzystać do zaprojektowania układu do izolowania produkowanego białka w szczególności techniką chromatografii powinowactwa. Przykładowo, wprowadzenie sekwencji rozpoznawanej przez przeciwciało pozwala na izolowanie zawierającego ją białka za pomocą tego przeciwciała. Innym przykładem są sekwencje aminokwasowe posiadające powinowactwo do glutationu. Kolejnym przykładem są techniki opierające się na znanym zjawisku tworzenia kompleksów niektórych jonów metali z niektórymi resztami aminokwasowymi. Najbardziej znanym przykładem takiego układu jest kompleksowanie jonów niklu przez pierścienie imidazolowe histydyn wprowadzonych do izolowanego łańcucha polipeptydowego. Wszystkie tego typu układy składające się ze znacznikowej sekwencji aminokwasowej i substancji, do której taka sekwencja posiada odpowiednio silne powinowactwo, pozwalają zaprojektować system oczyszczania białka zawierającego sekwencję znacznikową. Zwykle będzie to technika chromatografii powinowactwa na złożu zawierającym wspomnianą substancję. W związku z powyższym, znacznikowa sekwencja aminokwasowa może zawierać sekwencję składającą się z sześciu kolejnych histydyn (His6). Pożądane białko wchodzące w skład ujawnionego białka fuzyjnego wspominanego powyżej może być dowolnym znanym białkiem, dla którego znana jest sekwencja aminokwasowa lub sekwencja kodująca. Przykładowo, może to być białko lecznicze, którego produkcja pożądana jest ze względu na jego właściwości terapeutyczne. W oparciu o instrukcje ujawnione w niniejszym opisie oraz powszechnie dostępną wiedzę fachowiec będzie w stanie opracować sekwencję kodującą bia ł- ko fuzyjne zawierającą sekwencję kodującą pożądane białko. Sekwencje aminokwasowe lub s e- kwencje kodujące znanych białek mogą być przykładowo pozyskane z bazy GenBank dostępnej w sieci internet pod adresem w której zgromadzono sekwencje znanych genów oraz sekwencje aminokwasowe znanych białek. Aby zwiększyć p o- ziom ekspresji białka fuzyjnego w układzie bakteryjnym można zastosować znane metody podn o- szenia poziomu ekspresji w komórkach bakteryjnych, które obejmują stosowanie silnych promot o- rów, stosowanie sekwencji wzmacniających transkrypcję lub stosowanie kodonów preferowanych przez wybraną komórkę bakteryjną. Ujawniony sposób charakteryzuje się tym, że białko fuzyjne posiada, korzystnie, sekwencję wybraną spośród: Z1 - Trp-Glu-Leu-Gln-Z2, Z1 - Trp-Glu-Leu-Thr-Z2, Z1 - Trp-Glu-Val-Gln-Z2, Z1 - Val-Glu-Leu-Gln-Z2, Z1 - Trp-Gln-Leu-Asp-Z2, Z1 - Trp-Val-Leu-Gln-Z2, Z1 - Phe-Glu-Val-Glu-Z2, Z1 - Gly-Arg-Gly-Val-Gly-Z2, Z1 - Gly-Arg-Gly-Val-Z2, Z1 - Val-Glu-Ile-Asp-Z2. W ujawnionym sposobie hydrolizę prowadzi się korzystnie w temperaturze od 0 C do 45 C, w ph od 6,0 do 9,0 lub w buforze fosforanowym, Bis-Tris, CAPS lub Tris o stężeniu od 1 do 250 mm lub w roztworze zawierającym od 0 do 500 mm NaCl. Kolejno ujawniono mutanta proteinazy SplB charakteryzującego się tym, że posiada sekwencję aminokwasową zawierającą przynajmniej jedną z następujących modyfikacji: - zamiana histydyny znajdującej się w pozycji 39 sekwencji SplB na inny aminokwas. - zamiana kwasu asparaginowego znajdującego się w pozycji 77 sekwencji SplB na inny aminokwas, - zamiana seryny znajdującej się w pozycji 157 sekwencji SplB na inny aminokwas, - przyłączenie wiązaniem peptydowym do aminokwasu znajdującego się na N-końcu dojrzałej formy proteinazy SplB polipeptydu zawierającego co najmniej jedną spośród następujących sekwencji: znaną sekwencję sekrecyjną, znaną bakteryjną sekwencję sekrecyjną, sekwencję polipeptydu wykazującego powinowactwo do centrum aktywnego wzmiankowanej proteinazy SplB, sekwencję znanego polipeptydu znacznikowego. Korzystnie ujawniony mutant proteinazy charakteryzuje się tym, że sekwencja sekrecyjna jest bakteryjną sekwencją sekrecyjną z Bacillus subtilis.
5 PL B1 5 Równie korzystnie ujawniony mutant proteinazy charakteryzuje się tym, posiada sekwencję wybraną spośród: SEQ ID NO.: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10 lub SEQ ID NO: 12. Ujawniono również sekwencję nukleotydową kodująca mutanta proteinazy zdefiniowanego powyżej. Korzystnie ta sekwencja nukleotydowa posiada sekwencję nukleotydową wybraną spośród: SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9 lub SEQ ID NO: 11. Kolejno ujawniono sposób otrzymywania proteinazy SplB lub jej mutanta charakteryzujący się tym, że: a) w komórkach gospodarza bakteryjnego lub innego prowadzi się ekspresję białka jak zdefiniowano powyżej, korzystnie kodowanego przez sekwencję nukleotydową zdefiniowaną powyżej, a następnie; b) izoluje się pożądany enzym lub zawierającą go frakcję. W korzystnej realizacji ujawnionego sposobu gospodarzem bakteryjnym jest szczep Bacillus subtilis ekspresjonujący białko kodowane przez sekwencję nukleotydową przedstawioną jako SEQ ID NO: 3. W równie korzystnej realizacji ujawnionego sposobu w etapie b) oddziela się brzeczkę fermentacyjną od masy bakteryjnej poprzez wirowanie, białka sekrecyjne znajdujące się w pozbawionej bakterii pożywce wysala się siarczanem amonu, oddziela się wysolone białka i rozpuszcza w niewielkiej ilości roztworu buforowego i dializuje się do buforu o ph około 5,5. W równie korzystnej realizacji ujawnionego sposobu w etapie b) dodatkowo oczyszcza się wyizolowane białko techniką chromatografii powinowactwa, chromatografii jonowymiennej i/lub sączenia molekularnego, a ostatecznie oczyszczony preparat zagęszcza się i ewentualnie poddaje krystalizacji. W jednej z korzystnych realizacji ujawnionego sposobu produkcji aktywnej proteinazy SplB można wykorzystać zdolności katalityczne samego enzymu. W metodzie tej produkuje się enzym z N-terminalną metką fuzyjną wybraną korzystnie z bogatej puli opisanych metek lub nowym peptydem o własnościach pożądanych dla metki. Metkę taką może stanowić przykładowo, lecz nie jedynie metka histydynowa (tzw. ang. His-tag). Pomiędzy metkę fuzyjną a sekwencję proteinazy SplB wstawia się dogodnie sekwencję rozpoznawaną i przecinaną przez proteinazę SplB. Po wyprodukowaniu opisanego białka fuzyjnego izoluje się je wykorzystując właściwości metki, a następnie odcina się metkę przy pomocy katalitycznych ilości proteinazy SplB. Uwolniona od metki proteinaza SplB zwiększa pulę aktywnego enzymu przyspieszając zakończenie procesu odcinania. Odcinanie metki można prowadzić bezpośrednio na złożu stosowanym do izolacji białka fuzyjnego lub też po elucji, przy czym pierwsza metoda pozwala na jednoczesne oczyszczenie proteinazy od metki fuzyjnej, natomiast w drugim przypadku konieczne jest wprowadzenie dodatkowego stopnia oczyszczania. W opisie ujawniono zastosowanie proteinazy SplB do specyficznej hydrolizy polipeptydu zawierającego sekwencję aminokwasową Xaa1-Xaa2-Xaa3-Xaa4, gdzie: Xaa1 jest aminokwasem wybranym spośród: Trp, Ala, Ile, Leu, Met, Phe, Tyr, Val, Ser, Thr lub Gly, Xaa2 jest aminokwasem wybranym spośród Glu, Gln, Asp, Asn, Val, Leu, Ile, Gly, Arg, Ser lub Thr, Xaa3 jest aminokwasem wybranym spośród Leu, Ile, Val, Thr, Ser lub Gly. Xaa4 jest aminokwasem wybranym spośród: Gln, Glu, Thr, Ser, Asp lub Asn. Hydrolizowany polipeptyd może posiadać sekwencję aminokwasową zawierającą sekwencję wybraną spośród: Trp-Glu-Leu-Gln-Gly, Trp-Glu-Leu-Gln, Trp-Glu-Leu-Thr, Trp-Glu-Val-Gln, Val-Glu-Leu-Gln, Trp-Gln-Leu-Asp, Trp-Val-Leu-Gln, Phe-Glu-Val-Glu, Gly-Arg-Gly-Val-Gly, Gly-Arg-Gly-Val, Val-Glu-Ile-Asp.
6 6 PL B1 Hydrolizę można prowadzić w temperaturze od 0 C do 45 C, w ph od 6,0 do 9,0. Hydrolizę prowadzi się również w buforze fosforanowym, Bis-Tris, CAPS lub Tris o stężeniu od 1 do 250 mm lub w roztworze zawierającym od 0 do 500 mm NaCl. Przedmiotem wynalazku jest proteinaza posiadająca aktywność proteinazy SplB posiadająca następujące elementy strukturalne: - w pozycji odpowiadającej Val28 w sekwencji SplB zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Val29 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Ile34 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Leu35 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Thr36 zawiera aminokwas wybrany spośród Ser, Thr; - w pozycji odpowiadającej His39 zawiera His; - w pozycji odpowiadającej Ile66 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Ile69 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Asp77 zawiera Asp; - w pozycji odpowiadającej Val78 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Val80 zawiera aminokwas wybrany spośród VaL Leu, Ile, Ala, Met; - w pozycji odpowiadającej Ile81 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala, Met; - w pozycji odpowiadającej Val118 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala, Ser, Thr; - w pozycji odpowiadającej Gly120 zawiera Gly; - w pozycji odpowiadającej Tyr121 zawiera aminokwas wybrany spośród Tyr, Phe, Trp; - w pozycji odpowiadającej Leu131 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala, Met; - w pozycji odpowiadającej Gly155 zawiera Gly; - w pozycji odpowiadającej Asn156 zawiera aminokwas wybrany spośród Asn, Gln, Asp, Glu; - w pozycji odpowiadającej Ser157 zawiera Ser; - w pozycji odpowiadającej Gly158 zawiera Gly; - w pozycji odpowiadającej Ser159 zawiera aminokwas wybrany spośród Val, Ala, Ser, Thr, Gly; - w pozycji odpowiadającej Pro 160 zawiera Pro; - w pozycji odpowiadającej Gly170 zawiera Gly; - w pozycji odpowiadającej Ile171 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Phe197 zawiera aminokwas wybrany spośród Tyr, Phe, Trp; - w pozycji odpowiadającej Ile198 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala. Uszczegóławiając w opisie ujawniono polipeptyd wykazujący powinowactwo do centrum aktywnego proteinazy SplB posiadający sekwencję aminokwasową Xaa1-Xaa2-Xaa3-Xaa4-Xaa5, gdzie Xaa1 jest aminokwasem z hydrofobowym łańcuchem bocznym lub aminokwasem z niewielkim łańcuchem bocznym, korzystnie wybranym spośród: Trp, Ala, Ile, Leu, Met, Phe, Tyr, Val, Ser, Thr lub Gly, Xaa2 to kwas glutaminowy, glutamina, kwas asparaginowy, asparagina lub aminokwas z niewielkim łańcuchem bocznym lub arginina, korzystnie Xaa2 jest aminokwasem wybranym spośród Glu, Gln, Asp, Asn, Val, Leu, Ile, Gly, Arg, Ser lub Thr. Xaa3 to aminokwas z niewielkim hydrofobowym łańcuchem bocznym albo aminokwas z niewielkim łańcuchem bocznym, korzystnie Xaa3 jest aminokwasem wybranym spośród Leu, Ile, Val, Thr, Ser lub Gly, Xaa4 jest aminokwasem wybranym spośród: Gln, Glu, Thr, Ser, Asp lub Asn, Xaa5 jest pominięty lub jest dowolnym aminokwasem. W sekwencjach odpowiednie symbole oznaczają: A, Ala, alanina; V, Val, walina; L, Leu, leucyna; I, Ile, izoleucyna; P, Pro, prolina; F, Phe, fenyloalanina; W, Trp, tryptofan; M, Met, metionina; G, Gly, glicyna; S, Ser, seryna; T, Thr, treonina; C, Cys, cysteina; Y, Tyr, tyrozyna; N, Asn, asparagina; Q Gln, glutamina; D, Asp, kwas asparaginowy; E, Glu, kwas glutaminowy; K. Lys, lizyna; R, Arg, arginina; H, His, histydyna; Xaa5 może być w przypadku proteinazy SplB pominięty lub być dowolnym aminokwasem gdyż nieoczekiwanie proteinaza SplB odróżnia się od innych proteinaz cechujących się wysoką specyficznością substratową, które wykazują zazwyczaj określoną specyficzność również wobec aminokwasu znajdującego się bezpośrednio za hydrolizowanym wiązaniem (na nowym N-końcu powstającym w wyniku hydrolizy wiązania peptydowego; tj. w miejscu P1 zgodnie z systemem
7 PL B1 7 numeracji przyjętym w tym zgłoszeniu, a zaproponowanym przez: Schechter, I., and Berger, A. (1967) Biochem. Biophys. Res. Commun. 27, ). W przypadku proteinazy SplB preferencji takiej nie obserwujemy. Cecha ta jest szczególnie korzystna, ponieważ pozwala na dowolne projektowanie N-końca uwalnianych produktów. Kolejny aspekt ujawnienia dotyczy białek posiadających ustaloną aktywność proteinazy SplB dzięki zachowaniu przez te białka struktury trzeciorzędowej proteinazy SplB przedstawionej w tabeli 1. Istnieje powszechnie stosowany parametr określający podobieństwo struktur trzeciorzędowych, który pozwala na zdefiniowanie grupy ujawnionych białek o pożądanych właściwościach. Parametr ten to RMS distance (deviation) (ang. root mean square distance (deviation)) określany także jako RMSD. Wartość parametru RMSD wylicza się porównując położenia odpowiadających sobie atomów po uprzednim nałożeniu na siebie porównywanych struktur w celu ich optymalnego dopasowania. Wartość parametru wyraża się w angstremach (A) i tak też przyjęto w dalszym tekście. Ogólnie, im niższa wartość parametru tym struktury bardziej podobne. Zatem niniejszy opis ujawnia białka o wzmiankowanej aktywności proteolitycznej, których struktura trzeciorzędowa jest dostatecznie podobna do struktury proteinazy SplB. Rzeczone podobieństwo mierzymy wartością parametru RMSD dla istotnych komponentów strukturalnych proteinazy SplB w stosunku do odpowiadających im komponentów strukturalnych porównywanego białka. W szczególności ujawniono enzym, którego: a) RMSD wszystkich atomów reszt triady katalitycznej (seryny, histydyny i kwasu asparaginowego) jest mniejsze lub równe 1,7Å, korzystnie mniejsze lub równe 1,5 Å, przy czym korzystnie dodatkowo spełnia ona co najmniej jedno z następujących kryteriów: b) RMSD odpowiadających strukturalnie węgli C łańcucha głównego w obrębie dobrze zdefiniowanych struktur drugorzędowych jest mniejsze lub równe 2,0 Å, korzystnie mniejsze lub równe 1,5 Å, c) tak jak w (b) z tym ze dobrze zdefiniowane strukturalnie elementy cząsteczki obejmują fragmenty łańcucha polipeptydowego wybrane korzystnie spośród następujących fragmentów określonych wg numeracji SplB oraz odpowiadających im strukturalnie fragmentów porównywanej cząsteczki: Val4 do Lys6, Thr16 do Ala20, Ala24 do Val29, Thr33 do Val40, Ile50 do Ala52, Ile63 do Asn71, Val78 do Glu84, Arg115 do Ile119, Leu131 do Val138, Ser145 do Tyr148, Thr152 do Leu162, Gly170 do Ser175, Ala185 do Tyr189, Lys196 do Ala199; d) RMSD atomów łańcucha głównego w obrębie dobrze zdefiniowanych struktur drugorzędowych jest mniejsze lub równe 2,2 Å, korzystnie mniejsze lub równe 1,8 Å; e) tak jak w (d) z tym ze dobrze zdefiniowane strukturalnie elementy cząsteczki obejmują fragmenty łańcucha polipeptydowego wybrane korzystnie spośród następujących fragmentów określonych wg numeracji SplB oraz odpowiadających im strukturalnie fragmentów porównywanej cząsteczki: Val4 do Lys6, Thr16 do Ala20, Ala24 do Val29, Thr33 do Val40, Ile50 do Ala52, Ile63 do Asn71, Val78 do Glu84, Arg115 do Ile119, Leu131 do Val138, Ser145 do Tyr148, Thr152 do Leu162, Gly170 do Ser175, Ala185 do Tyr189, Lys196 do Ala199; f) Fragmenty odpowiadające strukturalnie fragmentom białka SplB wybranym korzystnie spośród określonych poniżej tworzą β-harmonijkę: Val4 do Thr5, Val18 do Ala20, Thr25 do Val28, Thr33 do Thr36, Arg49 do Ala52, Ile63 do Asn71, Ser79 do Val83, Arg115 do Ile119, Tyr132 do Gly136, Ser145 do Tyr148, Val161 do Leu162, Gly170 do Ser175, Ala185 do Val188 g) Fragmenty odpowiadające strukturalnie fragmentom białka SplB wybranym korzystnie spośród określonych poniżej tworzą α-helisę: Lys38 do Ser41, Lys196 do Glu200. Analiza struktury trzeciorzędowej proteinazy SplB pozwoliła zlokalizować regiony oraz reszty szczególnie istotne w procesie rozpoznawania substratu i katalizy. Ujawniono zatem białka posiadające reszty odpowiadające następującym kluczowym resztom aminokwasowym w sekwencji prote i- nazy SplB: a.) reszty tzw. triady katalitycznej które w przypadku proteinazy SplB stanowią: S157, H39 i D77. Zamiana tych reszt skutkuje całkowitą utratą zdolności katalitycznych. Przykładowo, dla proteinazy SplB wykazano, że mutant S157 A jest całkowicie pozbawiony aktywności proteolitycznej. b.) reszty odpowiedzialne za rozpoznanie substratu: P1: przede wszystkim S175, H172, T152 do N156, A174
8 8 PL B1 P2: przede wszystkim F173, H39 i D77 P3: przede wszystkim S175 P4: przede wszystkim F173 i Y186, d.) reszta kwasu glutaminowego na N-końcu łańcucha polipeptydowego, która to reszta jest odpowiedzialna za stabilizację N-końca białka przez wiązania wodorowe a tym samym umożliwia wyrażanie pełnej aktywności proteolitycznej. Reszta ta może być ewentualnie zastąpiona resztą kwasu asparaginowego wykazującą podobne właściwości fizykochemiczne. Ujawnione w pracy J. Mol. Biol. (2006) 358, porównanie sekwencji aminokwasowych homologicznych białek gronkowcowych (proteinaz.) V8 oraz toksyn epidermolitycznych) i trypsyny wskazuje na ważne rejony w sekwencji białka niezbędne dla prawidłowego fałdowania i/lub zachowania funkcji (fig. 1): V28 do V40; D77 do I81; G120 do P122 oraz G155 do I171. Ponadto widać wyraźnie konserwację pojedynczych reszt: I50: S134; I146: V188 i I198. Jednak dopiero rozwiązanie struktury trzeciorzędowej proteinazy SplB pozwala na stworzenie porównania sekwencji na podstawie porównania struktur - a więc porównania sekwencji odpowiadających sobie elementów strukturalnych (najpierw porównuje się struktury, a następnie tam gdzie są podobne zestawia się tylko sekwencje, nawet jeśli nie są one homologiczne w klasycznym rozumieniu). Rozwiązanie takie niesie ze sobą dużo więcej informacji niż zwykłe porównanie sekwencji gdyż wskazuje elementy ważne dla zachowania funkcji białka. Porównanie takie zostało przedstawione na fig. 2. gdzie odpowiednie fragmenty są zgrupowane na podstawie podobieństw strukturalnych. Takie podejście pozwala na ewidentne wyróżnienie regionów konserwatywnych istotnych dla funkcji białka. Zatem białko według wynalazku powinno posiadać w miejscach odpowiadających następującym aminokwasom w sekwencji SplB następujące reszty: Val28 - optymalnie zawiera aminokwasy z niewielkim, hydrofobowym łańcuchem bocznym wybrane spośród Val, Leu, Ile, Ala: Val29 - optymalnie zawiera aminokwasy z niewielkim, hydrofobowym łańcuchem bocznym wybrane spośród Val, Leu, Ile, Ala; Ile34 - optymalnie zawiera aminokwasy z niewielkim, hydrofobowym łańcuchem bocznym wybrane spośród Val, Leu, Ile, Ala; Leu35 - optymalnie zawiera aminokwasy z niewielkim, hydrofobowym łańcuchem bocznym wybrane spośród Val, Leu, Ile, Ala; Thr36 - optymalnie zawiera aminokwasy wybrane spośród Ser, Thr; His39 - histydyna triady katalitycznej; Ile66 - optymalnie zawiera aminokwasy z niewielkim, hydrofobowym łańcuchem bocznym wybrane spośród Val, Leu, Ile, Ala; Ile69 - optymalnie zawiera aminokwasy z niewielkim, hydrofobowym łańcuchem bocznym wybrane spośród Val, Leu, Ile, Ala; Asp77 - kwas asparaginowy triady katalitycznej; Val78 - optymalnie zawiera aminokwasy z niewielkim, hydrofobowym łańcuchem bocznym wybrane spośród Val, Leu, Ile, Ala; Val80 - optymalnie zawiera aminokwasy wybrane spośród Val, Leu, Ile, Ala, Met; Ile81 - optymalnie zawiera aminokwasy wybrane spośród Val, Leu, Ile, Ala, Met; Val118 - optymalnie zawiera aminokwasy wybrane spośród Val, Leu, Ile, Ala, Ser, Thr: Gly120 - optymalnie Gly; Tyr121- optymalnie zawiera aminokwasy z dużym, hydrofobowym łańcuchem bocznym wybrane spośród Tyr, Phe, Trp; Leu131 - optymalnie zawiera aminokwasy wybrane spośród Val, Leu, Ile, Ala, Met Gly155 - optymalnie Gly; Asn156 - optymalnie zawiera aminokwasy wybrane spośród Asn, Gln, Asp, Glu; Ser157 - seryna triady katalitycznej, o której była mowa wcześniej, musi być Ser; Gly158 - optymalnie Gly; Ser159 - optymalnie zawiera aminokwasy wybrane spośród Val, Ala, Ser, Thr, Gly; Pro160 - optymalnie Pro; Gly170 - optymalnie Gly; Ile171 - optymalnie zawiera aminokwasy wybrane spośród Val, Leu, Ile, Ala;
9 PL B1 9 Phe197 - optymalnie zawiera aminokwasy z dużym, hydrofobowym łańcuchem bocznym wybrane spośród Tyr, Phe, Trp; Ile198 -optymalnie zawiera aminokwasy z niewielkim, hydrofobowym łańcuchem bocznym wybrane spośród Val, Leu, Ile, Ala. Szczególną realizacją jest białko posiadające strukturę proteinazy SplB określoną w tabeli 1. Zgodnie z ujawnieniem proteinaza SplB rozpoznaje specyficzną sekwencję aminokwasową i hydrolizuje łańcuch polipeptydowy zaraz za lub w obrębie rozpoznawanej sekwencji. Z uwagi na długość rozpoznawanej sekwencji (cztery kolejne aminokwasy) liczba identycznych sekwencji w proteomie człowieka wynosi zaledwie kilkanaście, a więc enzym ten nadaje się m. in. do odcinania metek fuzyjnych przy produkcji pozostałych kilkudziesięciu tysięcy białek ludzkich. Ujawniono przede wszystkim sekwencje aminokwasowe łańcucha polipeptydowego specyficznie rozpoznawane lub specyficznie rozpoznawane i hydrolizowane przez proteinazę SplB, sekwencje nukleotydowe kodujące rzeczone sekwencje aminokwasowe (a więc pozwalające na produkcję polipeptydów je zawierających przy pomocy technologii białek rekombinowanych) oraz metodę specyficznej hydrolizy polipeptydów zawierających rzeczone sekwencje aminokwasowe przy pomocy proteinazy SplB. Ujawniono także samą proteinazę SplB jako enzym rozpoznający lub rozpoznający i hydrolizujący wybrane sekwencje aminokwasowe oraz sposoby produkcji proteinazy SplB w systemie rekombinowanym. Ponadto ujawniono syntetyczne substraty oparte na sekwencjach specyficznie rozpoznawanych i hydrolizowanych przez proteinazę SplB. Podsumowując najważniejsze zalety prezentowanego wynalazku, należy uznać, że przedmiot wynalazku oraz szczególnie korzystne ujawnione aspekty mogą znaleźć zastosowanie w następujących procesach: a) rozpoznanie specyficznej sekwencji aminokwasowej łańcucha polipeptydowego (w szczególności sekwencji białka rekombinowanego) i jego specyficzna hydroliza w ściśle określonym miejscu w obrębie lub w niewielkiej odległości od rozpoznawanej sekwencji. b) wysoce wydajna produkcja proteinazy SplB. Przedmiot wynalazku został zdefiniowany w zastrzeżeniach patentowych. Dla lepszego zilustrowania istoty wynalazku i ujawnionych aspektów niniejszy opis wzbogacono o wykaz sekwencji i figury. Sekwencja nr 1 (SEQ ID NO 1) prezentuje sekwencję kodującą proteinazę SplB ze Staphylococcus aureus wraz z jej natywnym peptydem sygnalnym. Sekwencja nr 2 (SEQ ID NO 2) prezentuje sekwencję aminokwasową proteinazy SplB ze Staphylococcus aureus (dojrzałe białko: aminokwasy od 1 do 204) wraz z jej natywnym peptydem sygnalnym (aminokwasy od -36 do - 1). Sekwencja nr 3 (SEQ ID NO 3) prezentuje sekwencję kodującą wariant proteinazy SplB ze Staphylococcus aureus, w której sekwencję kodującą natywny peptyd sygnalny zastąpiono sekwencją kodującą peptyd sygnalny pochodzący z Bacillus subtilis. Sekwencja nr 4 (SEQ ID NO 4) prezentuje sekwencję aminokwasową wariantu proteinazy SplB ze Staphylococcus aureus, w której sekwencję natywnego peptydu sygnalnego zastąpiono sekwencją peptydu sygnalnego pochodzącego z Bacillus subtilis (aminokwasy od -29 do -1). Sekwencja nr 5 (SEQ ID NO 5) prezentuje sekwencję kodującą wariant proteinazy SplB z S. aureus z peptydem sygnalnym z B. subtilis zawierający substytucję S157A natomiast sekwencja nr 6 (SEQ ID NO 6) prezentuje sekwencję aminokwasową tego wariantu. Sekwencja nr 7 (SEQ ID NO 7) prezentuje sekwencję kodującą wariant proteinazy SplB z S. aureus z peptydem sygnalnym z B. subtilis zawierający substytucję EI39A natomiast sekwencja nr 8 (SEQ ID NO 8) prezentuje sekwencję aminokwasową tego wariantu. Sekwencja nr 9 (SEQ ID NO 9) prezentuje sekwencję kodującą wariant proteinazy SplB z S. aureus z peptydem sygnalnym z B. subtilis zawierający substytucję D77A natomiast sekwencja nr 10 (SEQ ID NO 10) prezentuje sekwencję aminokwasową tego wariantu. Sekwencja nr 11 (SEQ ID NO 11) prezentuje sekwencję kodującą białko fuzyjne zawierające sekwencję dojrzałej formy SplB z S. aureus, do której przyłączono metkę histydynową i sekwencję rozpoznawaną przez SplB natomiast sekwencja nr 12 (SEQ ID NO 12) prezentuje sekwencję aminokwasową tego białka.
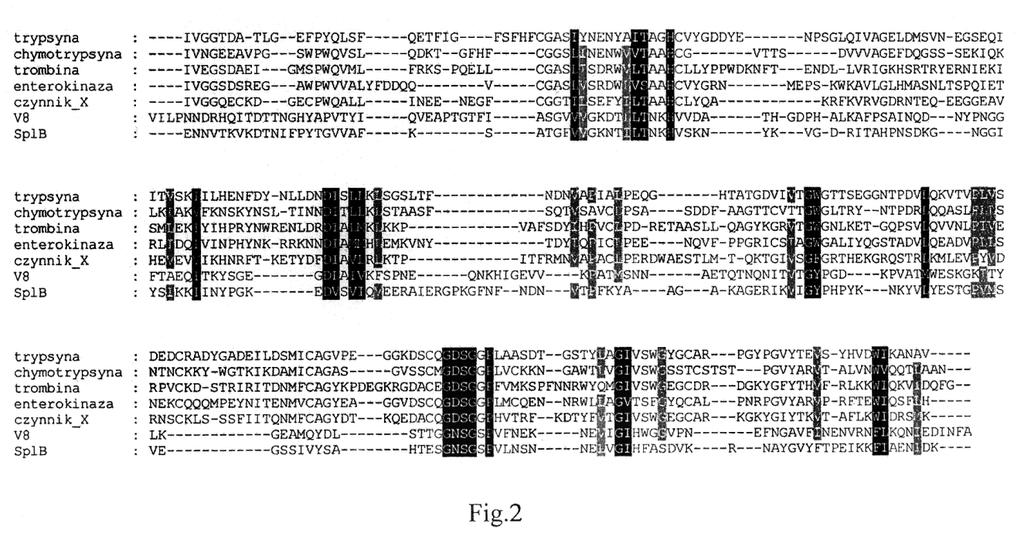
10 10 PL B1 Figura 1 zawiera porównanie sekwencji aminokwasowych blisko spokrewnionych proteinaz: proteinaza SplB, proteinaza SplC, V8 (proteinaza V8 ze Staphylococcus aureus zwana inaczej gluta-mylendopeptydazą), ETA - toksyna epidermolityczna A ze Staphylococcus aureus oraz daleko spokrewnionego enzymu - trypsyny (modelowy enzym dla grupy proteinaz trypsynopodobnych). Podobieństwa sekwencji oznaczono odcieniami szarości - im ciemniejsze tym większe podobieństwo. Regiony o wyraźnej homologii sekwencji oraz pojedyncze konserwatywne reszty zaznaczono ramkami. Figura 2 prezentuje porównanie sekwencji aminokwasowych blisko spokrewnionych proteinaz stworzone na podstawie znajomości ich struktur trzeciorzędowych oraz znajomości struktury trzeciorzędowej ustalonej dla proteinazy SplB; wskazano reszty szczególnie istotne dla zachowania struktury i aktywności proteinazy. Poniższe przykłady zostały umieszczone jedynie w celu lepszego wyjaśnienia poszczególnych aspektów wynalazku i nie powinny być utożsamiane z całym jego zakresem, który zdefiniowano w załączonych zastrzeżeniach. P r z y k ł a d 1. Wstępna charakterystyka proteinazy SplB Wyjściowym eksperymentem, umożliwiającym dalsze prace było wyznaczenie optimum ph i temperatur, dopuszczalnych stężeń soli i innych odczynników oraz stabilności enzymu. W tym celu należało opracować ilościową metodę oznaczania aktywności enzymu. Przeprowadzone liczne pr ó- by z syntetycznymi substratami niskocząsteczkowymi opisane w J. Mol. Biol. (2006) 358, oraz kontynuowane również po opublikowaniu tej pracy dla większej liczby substratów nie pozwoliły jednak uzyskać lepszej charakterystyki proteinazy SplB. Enzym wykorzystany w tych eksperymentach otrzymano techniką opisaną w J. Mol. Biol. (2006) Wśród znanych substratów proteinaz trypsynopodobnych nie zidentyfikowano substratu trawionego nawet w minimalnym stopniu przez proteinazę SplB. Po długich poszukiwaniach ustalono, że FTC kazeina (kazeina znakowana barwnikiem fluorescencyjnym, umożliwiająca badanie hydrolizy metodą spektrofluorescencji) jest mało wydajnym substratem białkowym trawionym przez proteinazę SplB. W przypadku trawienia tego substratu konieczne było stosowanie nadmiaru molowego enzymu w stosunku do substratu oraz długich (rzędu godzin) czasów inkubacji. Przy jego użyciu udało się jednak określić wstępną charakterystykę aktywności proteinazy SplB jako: optimum ph na 8,25 (+/- 1,5 jednostki w dół i 1 jednostkę w górę), brak wyraźnej zależności aktywności od stężeń popularnych soli do 0.5M, brak wpływu środków redukujących do kilku Mm, szeroką tolerancję temperaturową. Na tej podstawie ustalono podstawowe parametry reakcji hydrolizy zalecane dla reakcji prowadzonej z wykorzystaniem proteinazy SplB. W efekcie, wszystkie kolejne eksperymenty prowadzono w temperaturze od 20 C do 37 C, w 10 do 100 mm buforze fosforanowym lub Tris przy wartości ph od 7,0 do 8,5 i stężeniu NaCl od 0 do 150 mm. Ponadto ustalono, że enzym można przechowywać w stanie zamrożonym bez wyraźnej utraty aktywności, oraz kilkakrotnie zamrażać i rozmrażać a także liofilizować. Można go także przechowywać kilka miesięcy w temperaturze 4 C bez wyraźnej utraty aktywności. Wszystkie powyższe warunki stanowią dogodne formy przechowywania enzymu, co jest niezwykle istotne w codziennej praktyce. P r z y k ł a d 2. Wstępne próby ustalenia specyficzności substratowej proteinazy SplB Trawienie -kazeiny Standardowo dla oznaczenia specyficzności substratowej proteinazy kontaktuje się ją z r óżnymi białkami, oznacza się miejsca hydrolizy i na podstawie odpowiedniej ilości prób metodami analizy statystycznej ustala się najbardziej optymalne miejsce cięcia. Takie standardowe post ę- powanie zastosowane zostało w pierwszym podejściu także dla proteinazy SplB, jednak nie przyniosło ono spodziewanych wyników. Wykazano, że szereg testowanych białek (lizozym białka jaja kury, inhibitor trypsyny z nasion soi, transferyna osocza ludzkiego, albumina osocza wołowego, owalbumina jaja kury. -galaktozydaza E. coli, anhydraza węglanowa, alfa-2-makroglobulina osocza ludzkiego, cytochrom c, kozie przeciwciała IgG, RNAza, fibrynogen, mioglobina wieloryba, cała gama serpin ludzkich i mysich) nawet przy przedłużonej inkubacji z nadmiarem enzymu nie ulega proteolizie. Wykrywalną aktywność białka SplB wykazano jedynie metodą zymografii na -kazeinie (J. Mol. Biol. (2006) 358, ). Dalsze eksperymenty innymi metodami (kontaktowanie proteinazy i kazeiny w roztworze, analiza produktów proteolizy przy pomocy SDS-PAGE) potwierdziły ten fakt jednak
11 PL B1 11 wykazały też, że dla przeprowadzenia reakcji hydrolizy potrzeba zastosowania molowego nadmiaru enzymu i bardzo długich czasów inkubacji - kilkunastu godzin. Oznacza to, że enzym bardzo niechętnie" hydrolizuje -kazeinę (standardowo przy tego typu badaniach stosuje się katalityczne ilości enzymu - 100x i mniej niż substratu oraz krótkie czasy inkubacji - rzędu minut). Metodami spektrometrii masowej oraz chemicznego sekwencjonowania aminokwasowego udało się oznaczyć cztery miejsca cięcia w obrębie cząsteczki -kazeiny (spacja oznacza miejsce cięcia): EEQQQ TEDEL FAQTQ SLVYP FTESQ SLTLT LPLLQ SWMHQ Na podstawie tej niewielkiej (a więc mało reprezentatywnej) grupy można by założyć, że zgodnie z wiedzą, iż w tego typu proteinazach specyficzność determinuje reszta w pozycji P1 proteinaza SplB potrzebuje reszty glutaminy (Q) w miejscu P1 substratu (wyróżnione tłustym drukiem). Założenie takie nie tłumaczy zupełnie dlaczego trawieniu nie ulegają inne białka zawierające bardzo wiele reszt glutaminy (Q) w szczególności sama -kazeinia, która była trawiona jedynie na kilka fragmentów pomimo, że zawiera kilkanaście reszt Q. Ponadto założenie takie nie tłumaczy także, dlaczego proteinaza SplB jest tak mało wydajna. Warto także zauważyć, że w pozostałych pozycjach sekwencji trawionych przez SplB w kazeinie, poza P1', tj. P5 do P2 oraz P2' do P5' nie można ustalić żadnego wspólnego elementu czy charakterystycznego układu, który sugerowałby znaczenie tych pozycji dla specyficzności substratowej badanej proteinazy. Wydaje się jedynie, że w pozycji P1' preferowane są reszty S lub T. Trawienie substratów syntetycznych, denaturowanych białek i peptydów syntetycznych W obliczu niepowodzenia eksperymentów opisanych powyżej przyjęto robocze założenie, że proteinaza SplB może trawić jedynie w specjalnych, eksponowanych rejonach białek, a z uwagi na ukrycie innych rejonów zawierających reszty Q wewnątrz struktury cząsteczki białek stosowanych jako substraty nie są one rozpoznawane i trawione. Dlatego przeanalizowano ponownie wybrane białka po uprzedniej denaturacji (karboksymetylowany lizozym, karboksymetylowany BPTI, apomioglobina i apocytochrom c w formie zdenaturowanej, po usunięciu cząsteczki hemu), oraz syntetyczny peptyd (KEGLTETTFEEDGVATGNHEYCVEV) i fluorescencyjne substraty syntetyczne charakteryzujące się brakiem struktur drugorzędowych (Abz-Glu-Ala-Leu-Gly-Thr-Ser-Pro-Arg-Lys(Dnp)-Asp-OH i Abz-Gln- Gly-Ile-Gly-Thr-Ser-Arg-Pro-Lys(Dnp)-Asp-OH). Ponadto zsyntetyzowano chemicznie i testowano peptydy odpowiadające regionom flankującym miejsce cięcia zidentyfikowane w cząsteczce kazeiny, mianowicie: FTESQSLTLT oraz EEQQQTEDEL. We wszystkich przypadkach, pomimo stosowania także nadmiarów molowych proteinazy SplB oraz przedłużonych czasów inkubacji (do 72h) nie udało się wykazać hydrolizy badanych polipeptydów. Wynik ten jest szczególnie zaskakujący w przypadku dwóch ostatnich peptydów o sekwencji identycznej do oznaczonych wcześniej miejsc cięcia. Zatem, standardowa metoda oznaczania specyficzności substratowej opisana powyżej, w przypadku proteinazy SplB zupełnie zawiodła. Potwierdzenie, że białko SplB jest proteinazą W świetle powyższych wyników, w obliczu wykazanego trawienia cząsteczki β-kazeiny przy zastosowaniu nadmiaru enzymu i przedłużonego czasu inkubacji możliwym do zaakceptowania wytłumaczeniem było zanieczyszczenie badanego preparatu proteinazy SplB śladami innej aktywności proteolitycznej. Innymi słowy proteinaza SplB mogła być, tak jak blisko spokrewniona, homologiczna proteinaza SplC, białkiem bez aktywności proteolitycznej (w pracy J. Mol. Biol. (2006) 358, wykazano brak aktywności proteolitycznej białka SplC bardzo blisko spokrewnionego z proteinazą SplB), a przy stosowaniu jej nadmiaru drobne zanieczyszczenia stosowanego prepar a- tu mogły ujawniać swoją aktywność. Ewentualność taką wyeliminowano zamieniając katalityczną resztę seryny enzymu na resztę alaniny. W proteinazach trypsynopodobnych zamiana taka prowadzi zawsze do całkowitego zahamowania aktywności. Oczyszczony preparat muteiny proteinazy SplB (S157 A) nie wykazywał zdolności hydrolizy -kazeiny nawet przy trzykrotnym nadmiarze molowym i 72 godzinnej inkubacji. Eksperyment ten dowiódł roli reszty S157 w mechanizmie katalizy proteinazy SplB oraz potwierdził, że to właśnie ten enzym a nie zanieczyszczenia obecne w preparacie są odpowiedzialne za hydrol i- zę -kazeiny.
12 12 PL B1 P r z y k ł a d 3. Nowa metoda otrzymywania proteinazy SplB SEQ ID NO: 1 i 2 przedstawia odpowiednio sekwencję nukleotydową genu kodującego proteinazę SplB ze Staphylococcus aureus oraz odpowiadającą jej sekwencję aminokwasową. Numeracja nukleotydów rozpoczyna się od a(1) trójki startu translacji (atg) a kończy na a(723)" trójki stopu translacji (taa). Łańcuch polipeptydowy proteinazy powstaje w procesie translacji w połącz e- niu z peptydem sygnalnym (numeracja reszt aminokwasowych od M(-36) do A(-1)), który w procesie sekrecji jest odcinany przez proteinazę sygnalną. Powstaje wtedy aktywna zewnątrzkomórkowa forma proteinazy SplB, którą można wyizolować z pożywki hodowlanej (numeracja reszt aminokwasowych od E1 do K204). W dalszym opisie stosuje się numerację wprowadzoną na tych sekwencjach. Sekwencje kodujące dojrzałą formę proteinazy SplB (E1 do K204) sklonowano do odpowiedniego plazmidu ekspresyjnego otrzymując plazmid umożliwiający produkcję zewnątrzkomórkową dojrzałej formy proteinazy SplB w bakteriach gramdodatnich. Sekwencję białka fuzyjnego składającego się z sygnałowej sekwencji sekrecyjnej specyficznej dla B. subtilis oraz dojrzałej formy proteinazy SplB. a także sekwencję nukleotydową kodującą to białko przedstawiono odpowiednio jako SEQ ID No. 3 i SEQ ID No. 4. W celu uzyskania białka zrekombinowanego bakterie B. subtilis szczep WB800 transformowano plazmidem ekspresyjnym i prowadzono selekcję transformantów na płytkach zawierających kanamycynę (50 g/ml). Wyselekcjonowanymi klonami inokulowano niewielką ilość płynnej pożywki (TSB; Sigma) zawierającej antybiotyk selekcyjny i inkubowano w 37 C z intensywnym mieszaniem przez 8 do 10 h. Tak przygotowaną hodowlą startową inokulowano hodowlę właściwą (4-16L płynnej pożywki z antybiotykami) i inkubowano przy intensywnym mieszaniu w 37 C przez 13 do 16 godzin. Wszystkie dalsze etapy oczyszczania przeprowadzano w 4 C. Bakterie oddzielano od pożywki przez wirowanie przy przyspieszeniu 6000xg przez 30 min. Białka sekrecyjne znajdujące się w pozbawionej bakterii pożywce wysalano siarczanem amonu do 80% nasycenia (561 g/l w 4 C). Wysolone białka oddzielano od pożywki przez wirowanie (15000xg, 1h), rozpuszczano w niewielkiej ilości 50 mm buforu octanowego ph 5,5 i dializowano przez noc do dużego nadmiaru tego samego buforu. Przedializowaną próbkę poddawano chromatografii jonowymiennej na złożu SP Sepharose FF (GE Healthcare) i zbierano frakcje zawierające największy szczyt białkowy w y- mywający się przy prze-wodnictwie buforu wynoszącym ok. 30 ms/cm. W razie wątpliwości frakcje testowano na obecność aktywności proteolitycznej metodą zymografii lub na obecność białka o odpowiedniej masie cząste-czkowej przy pomocy elektroforezy SDS-PAGE albo w inny dogodny sposób. Preparat dializowano do 50 mm buforu octanowego ph 4,8 i poddawano chromatografii jonowymiennej na złożu SOURCE 15S (GE Healthcare). Zbierano frakcje zawierające główny szczyt białkowy i poddawano sączeniu molekularnemu na złożu Superdex S75 w buforze PBS. Tak przygotowany, oczyszczony preparat zagęszczano, porcjowano i przechowywano zamrożony w -20 C. P r z y k ł a d 4. Ustalenie struktury trzeciorzędowej i specyficzności substratowej proteinazy SplB. Metoda opisana w przykładzie 3 pozwoliła na wydajną produkcję badanego białka umożliwiając prowadzenie dalszej analizy jego struktury, a zwłaszcza uzyskanie krystalicznej postaci proteinazy SplB i ustalenie struktury trzeciorzędowej badanej proteinazy, co w efekcie przyczyniło się do określenia specyficzności substratowej proteinazy SplB. Analiza struktury trzeciorzędowej proteinazy SplB W celu wskazania na ewentualne determinanty strukturalne obserwowanej bardzo słabej kinetyki hydrolizy wiązań peptydowych oraz ewentualne wskazanie lepszych substratów, metodą krystalografii rentgenowskiej określono strukturę trzeciorzędową proteinazy SplB. Ustalone koordynaty poszczególnych atomów białka dojrzałej proteinazy SplB zgromadzono w tabeli 1. Analiza otrzymanego modelu strukturalnego wskazała, że proteinaza SplB wykazuje budowę charakterystyczną dla proteinaz rodziny S1 (trypsynopodobnych/chymotrypsynopodobnych) nie wykazując wyraźnych uwarunkowań w budowie triady katalitycznej dla obserwowanej słabej aktywności. Ponadto analiza wykazała dobrze wykształcone miejsce P1 zdolne do przyjęcia aminokwasów: D, E, Q lub N oraz charakterystyczną hydrofobową łatę" na powierzchni białka w okolicy miejsca P3/P4 wskazującą na możliwość, że proteinaza SplB rozpoznaje poza resztą P1 także dalsze reszty substratu, a brak takiego rozpoznania (tj. odpowiedniego miejsca w badanych substratach) może warunkować obserwowaną we wcześniejszych eksperymentach słabą aktywność proteolityczną.
13 PL B1 13 Kluczowe reszty aminokwasowe w sekwencji proteinazy SplB Ze stanu techniki wiadomo, że kluczowymi resztami dla aktywności trypsynopodobnych proteinaz serynowych są reszty tzw. triady katalitycznej. W przypadku proteinazy SplB, uzyskana struktura trzeciorzędowa potwierdza, że są to: S157, H39 i D77. Zamiana tych reszt skutkuje całkowitą utratą zdolności katalitycznych. Odpowiednie sekwencje nukleotydowe i aminokwasowe takich mutantów przedstawione zostały jako SEQ ID No: Stosując ujawnione sekwencje oraz metodę opisaną w przykładzie 3 można uzyskać białka odpowiednich mutantów i poddać je dalszej analizie. Przykładowo, potwierdzono eksperymentalnie, że mutant S157 A jest całkowicie pozbawiony aktywności proteolitycznej. Na podstawie ustalonej struktury trzeciorzędowej proteinazy SplB oraz modelowania sposobu blokowania substratu m. in. na podstawie znajomości struktur kompleksów homologicznych białek z ich substratami i inhibitorami wynika, że reszty odpowiedzialne za rozpoznanie substratu to: P1: przede wszystkim S175, H172, T152 do N156, A174 P2: przede wszystkim F173, H39 i D77 P3: przede wszystkim S175 P4: przede wszystkim F173 i Y186 Porównanie struktur trzeciorzędowych form w pełni aktywnej (identyczna z natywną) i sł a- boaktywnej (zawierająca dwa dodatkowe aminokwasy na N-końcu) proteinazy SplB wskazuje na rolę dokładnego umiejscowienia N-terminalu białka oraz początkowej reszty kwasu glutaminowego (E1). Porównanie sekwencji aminokwasowych oraz struktur trzeciorzędowych homologicznych bi a- łek gronkowcowych (proteinazy V8 oraz toksyn epidermolitycznych) i trypsyny wskazuje na ważne rejony w sekwencji białka niezbędne dla prawidłowego fałdowania i/lub zachowania funkcji (patrz fig. 1): V28 do V40; D77 do I81; G120 do P122 oraz G155 do I171. Ponadto widać wyraźnie konserwację pojedynczych reszt: I50; S134; I146; V188 i I198. Analiza bibliotek substratów syntetycznych Bazując na wynikach analizy krystalograficznej, w dalszym etapie poszukiwania optymalnych substratów dla proteinazy SplB wykorzystano kombinatoryczną bibliotekę substratów syntetycznych zawierającą różnych substratów. Biblioteka zawiera substraty, w których na pozycjach P4 do P1 znajdują się wszystkie możliwe permutacje 18 aminokwasów (poza metioniną i cysteiną) a pozycję P1' zajmuje 7-amido-4-fluorometylokumaryna. barwnik wykazujący fluorescencję po odcięciu przez proteinazę od części peptydowej co pozwala na detekcję preferowanych substratów (szczegółowy opis Biol. Chem. (2004). 385: ). W pierwszym etapie badań skupiono się na ustaleniu preferowanej reszty w pozycji P1. Przegląd biblioteki proteinazą SplB pozwolił na ustalenie, że proteinaza SplB w pozycji P1 na 18 testowanych peptydów toleruje jedynie następujące aminokwasy: Asp, Asn, Gln (wynik zgodny z wynikami trawienia -kazeiny oraz z przewidywaniami na podstawie analizy struktury proteinazy). Szybkość trawienia wyselekcjonowanych substratów była porównywalna z innymi proteinazami demonstrując, że proteinaza SplB wcale nie jest mało wydajna jak sugerowały wyniki wcześniejszych eksperymentów opisanych w stanie techniki i przykładach 1 i 2. Stosowana biblioteka nie umożliwia odczytania wyników selekcji w pozycjach P2 do P4. Rozważając jednak odpowiedź na pytanie dlaczego proteinaza SplB nie trawi innych poza β-kazeiną białek, w których znajduje się cały szereg reszt Asp, Asn i Gln, na tym etapie prac stało się oczywiste, że trypsynopodobna proteinaza SplB posiada znacznie większą specyficzność substratową w porównaniu z jej bliskimi (proteinaza V8) i dalekimi (trypsyna, chym otrypsyna i wiele innych) homologami. Analiza biblioteki kombinatorycznej CLiPS Wysoka specyficzność substratowa zmusza do przesiania znacznie większej ilości substratów dla znalezienia tego właściwego i stąd konieczność zastosowania bardziej zaawansowanej metody CLiPS (opisanej w publikacji PNAS, (2006). 130: ). Bardzo ogólnie, w uzyskanej tą techniką bibliotece jedno z białek zewnętrznej błony komórkowej bakterii jest tak skonstruowane synt e- tycznie, że zawiera wszystkie możliwe permutacje liniowej sekwencji kilku aminokwasów (każdy szczep bakterii należący do biblioteki zawiera białko o konkretnej sekwencji, ale inne niż pozostałe szczepy bakterie należące do biblioteki). Za sekwencją zmienną znajduje się sekwencja umożliwiająca detekcję fluorescencyjną. Pierwszy etap selekcji (cytometria przepływowa) wybiera komórki fluoryzujące (gdzie interesujące białko ulega ekspresji) następnie komórki te kontaktuje się z testo-
14 14 PL B1 waną proteinazą i selekcjonuje się te, które nie fluoryzują, a więc te, dla których proteinaza odcięła część fluoryzującą. Następnie dla wyselekcjonowanych w ten sposób szczepów określa się sekwencję interesującego białka, a więc i sekwencje cięcia. Zastosowanie tej metody pozwala na przesianie 64 milionów substratów i uzyskanie informacji co do aminokwasów występujących w pozycjach P5 do P1', a nie tylko P1 (jak w technice opisanej powyżej). Wykorzystując tą metodę wyselekcjonowano następujące sekwencje rozpoznawane i cięte przez proteinazę SplB: P 4 P 3 P 2 P 1 *P 1 ' Szybkość cięcia G W E L Q * S 0,81 S W E L Q * G 0,62 S W E L T * G 0,62 S W E L T * V 0,90 S W E V Q * E 0,84 V V E L Q * S 0,65 S W E L Q * V 0,61 S W E L Q * S 0,86 S W E L Q * E 0,82 S W E L Q * M 0,75 E W E L Q * S 0,58 S W E L Q * A 0,53 S W Q L D * A 0,57 S W V L Q * A 0,32 W E L Q * S e k w e n c j a k o n s e n s u s o w a Wytłuszczoną czcionką zaznaczono aminokwasy odpowiadające dokładnie wyselekcjonowanej sekwencji konsensusowej, podkreślono aminokwasy dobiegające od sekwencji konsensuowej, gwiazdką oznaczono miejsce cięcia. Liczba po prawej stronie sekwencji jest miarą szybkości tr a- wienia. W świetle wcześniejszych eksperymentów i stanu techniki uzyskany wynik jest co najmniej nieoczywisty. Dotychczasowa wiedza o biochemii dziesiątek proteinaz trypsynopodobnych wskazuje prawie wyłącznie na miejsce P1 jako determinujące specyficzność substratową w tego typu białkach. Również wysoce homologiczna do proteinazy SplB - proteinaza V8 także ze Staphylococcus aureus wykazuje specyficzność jedynie dla reszty P1. Dlatego pierwotnie oczekiwano, zgodnie z ogólnym stanem wiedzy, że tak samo będzie w przypadku proteinazy SplB. Ponieważ proteinazy specyficzne tylko dla P1 nie są szczególnie specyficzne mierząc miarą metody CLiPS, metoda ta nie jest zalecana do określania specyficzności takich enzymów. Dopiero wiele niepowodzeń przy próbach przyrównania proteinazy SplB do wiedzy wynikającej ze stanu techniki skłoniło twórców do postawienia i przetestowania innej, mniej prawdopodobniej hipotezy dotyczącej specyficzności badanej proteinazy, która to hipoteza nieoczekiwanie okazała się prawdziwa. P r z y k ł a d 5. Wykorzystanie proteinazy SplB do specyficznej hydrolizy białek zawierających sekwencje aminokwasowe według wynalazku Trafność wyboru sekwencji konsensusowej oraz przydatność proteinazy SplB zostały potwierdzone w kolejnych eksperymentach. Wykorzystano plazmid umożliwiający ekspresję stafost a- tyny A jako białka fuzyjnego z GST odcinalnym przy pomocy trombiny (opisany w Mol. Microbiol. (2003). 49: ; zawierający sekwencję kodującą białko stafostatyna A, którą sklonowano techniką PCR z matrycy genomowego DNA S. aureus do plazmidu pgex-5t w miejsca BamHI/XhoI uzyskując plazmid umożliwiający ekspresję białka fuzyjnego GST-miejsce cięcia trombinystafostatyna A). Inkubacja białka fuzyjnego GST-miejsce cięcia trombiny-stafostatyna A z proteinazą SplB, nawet przy przedłużonym czasie inkubacji z nadmiarem enzymu, nie prowadzi do widocznej w metodzie SDS-PAGE hydrolizy interesującego łańcucha polipeptydowego. Metodami inżynierii genetycznej (mutageneza punktowa) zamieniono w omówionym wyżej plazmidzie sekwencję nukl e- otydową kodującą miejsce cięcia dla trombiny (LVPR*G) na sekwencję konsensusową (W ELQ*G) uzyskując plazmid umożliwiający ekspresję białka fuzyjnego GST-miejsce cięcia proteinazy SplBstafostatyna A. Białko to wyprodukowano w bakteriach E. coli szczepu BL21 plyss i oczyszczono wykorzystując powinowactwo białka fuzyjnego GST do immobilizowanego glutationu analogicznie jak opisano Mol. Microbiol. (2003). 49: dla białka fuzyjnego GST-miejsce cięcia trombiny-
15 PL B1 15 stafostatyna A. Kontaktując tak przygotowane białko z proteinazą SplB wykazano bardzo szybką hydrolizę łańcucha polipeptydowego (w czasie rzędu kilkunastu minut przy stukrotnym nadmiarze molowym substratu nad proteinazą SplB). Oznacza to, że proteinaza SplB nie jest mało wydajnym katalitycznie enzymem jak sugerowały eksperymenty z trawieniem -kazeiny. Przeciwnie, dowodzi to, że jest ona enzymem bardzo wydajnym katalitycznie ale jedynie w stosunku do substratów o prawidłowej sekwencji, która jest nieoczekiwanie znacznie bardziej rozbudowana w porównaniu ze znanymi proteinazami trypsynopodobnymi. Ponadto wyizolowano stafostatynę A uwolnioną z białka fuzyjnego przez trawienie proteinazą SplB i oznaczono metodą degradacji Edmana jej N-końcową sekwencję wykazując, że proteinaza SplB tnie specyficznie i precyzyjnie w obrębie rozpoznawanej sekwencji jedynie w określonym *miejscu (WELQ*G). Podobny wynik powinien przynieść eksperyment, w którym sekwencję rozpoznawaną przez proteinazę SplB wg opisu, zwłaszcza sekwencję konsensusową WELQG, umieszcza się pomiędzy metką" histydynową (His-Tag), lub dowolną inną metką a dowolnym interesującym białkiem, lub między dowolnym interesującym białkiem a dowolną metką, by tak samo jak poprzednio uzyskać precyzyjne odcinanie metki od interesującego białka. P r z y k ł ad 6. Rola miejsca P1' w rozpoznaniu substratu Dla wielu specyficznych proteinaz dużą rolę W' rozpoznaniu substratu grają nie tylko miejsca P ale także P' a głównie P1' (np. dla trombiny musi to być mały aminokwas (zwykle Gly)). Jest to dość niewygodne gdyż nie pozwala na dowolne kształtowanie N-końca białka po odcięciu metki. Z analizy substratów wyselekcjonowanych w metodzie CLiPS wynika, że w przypadku proteinazy SplB reszta ta nie ma większego znaczenia (na miejscu PT znajdujemy S, G. V, A ale też aminokwasy o dużym łańcuchu bocznym jak E lub M). Podobny wynik powinien przynieść eksperyment, w którym w białku fuzyjnym GST-miejsce cięcia przez proteinazę SplB - stafostatyna A w pozycji P1' umieszcza się różne aminokwasy (np. zamiana sekwencji WELQ*G na WELQ*Q, WELQ*N), by w sposób analogiczny do opisanego w przykładzie 5, potwierdzić brak wpływu wprowadzonej zamiany na szybkość hydrolizy wiązania. W tym celu, do białka fuzyjnego GST-miejsce cięcia przez proteinazę SplB - stafostatyna A wprowadzono na pozycji P1' następujące aminokwasy: E, K, N, Q, L, F, M. W efekcie uzyskano konstrukty zawierające w białku fuzyjnym opisanym w przykładzie 5 w miejsce sekwencji WELQ GS następujące fragmenty sekwencji: WELQ ES WELQ KS WELQ NS WELQ QS WELQ LS WELQ FS WELQ MS Wszystkie białka zrekombinowane otrzymane z tych konstruktów były cięte równie wydajnie przez proteinazę SplB potwierdzając brak wpływu miejsca P1' na rozpoznawanie i hydrolizę substratu. Miejsce cięcia zaraz za rozpoznawaną sekwencją konsensusową (WELQ) potwierdzono w każdym przypadku przez sekwencjonowanie uwolnionego nowego N-końca białka metodą degradacji Edmana.

16 16 PL B1 T a b e l a 1 Koordynaty struktury trzeciorzędowej proteinazy SplB ze Staphylococcus aureus, oznaczenia: NA - numer porządkowy atomu, A - rodzaj atomu, AK- rodzaj aminokwasu, NAK - numer porządkowy aminokwasu w strukturze pierwszorzędowej, X, Y, Z - koordynaty atomu

17 PL B1 17

18 18 PL B1

19 PL B1 19

20 20 PL B1

21 PL B1 21

22 22 PL B1

23 PL B1 23

24 24 PL B1

25 PL B1 25
26 26 PL B1
27 PL B1 27
28 28 PL B1
29 PL B1 29
30 30 PL B1
31 PL B1 31
32 32 PL B1
33 PL B1 33
34 34 PL B1
35 PL B1 35
36 36 PL B1 Zastrzeżenia patentowe 1. Proteinaza posiadająca aktywność proteinazy SplB, znamienna tym, że posiada następujące elementy strukturalne: - w pozycji odpowiadającej Val28 w sekwencji SplB zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Val29 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Ile34 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Leu35 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Thr36 zawiera aminokwas wybrany spośród Ser, Thr; - w pozycji odpowiadającej His39 zawiera His; - w pozycji odpowiadającej Ile66 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Ile69 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Asp77 zawiera Asp; - w pozycji odpowiadającej Val78 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Val80 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala, Met; - w pozycji odpowiadającej Ile81 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala, Met; - w pozycji odpowiadającej Val118 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala, Ser, Thr; - w pozycji odpowiadającej Gly120 zawiera Gly; - w pozycji odpowiadającej Tyr121 zawiera aminokwas wybrany spośród Tyr, Phe, Trp; - w pozycji odpowiadającej Leu131 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala, Met; - w pozycji odpowiadającej Glyl55 zawiera Gly; - w pozycji odpowiadającej Asn156 zawiera aminokwas wybrany spośród Asn, Gln, Asp, Glu; - w pozycji odpowiadającej Ser157 zawiera Ser; - w pozycji odpowiadającej Gly158 zawiera Gly; - w pozycji odpowiadającej Ser159 zawiera aminokwas wybrany spośród Val, Ala, Ser, Thr, Gly; - w pozycji odpowiadającej Pro160 zawiera Pro; - w pozycji odpowiadającej Gly170 zawiera Gly; - w pozycji odpowiadającej Ile171 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala; - w pozycji odpowiadającej Phe197 zawiera aminokwas wybrany spośród Tyr, Phe, Trp; - w pozycji odpowiadającej Ile198 zawiera aminokwas wybrany spośród Val, Leu, Ile, Ala.

37 PL B1 37 Rysunki
Przegląd budowy i funkcji białek
 Przegląd budowy i funkcji białek Co piszą o białkach? Wyraz wprowadzony przez Jönsa J. Berzeliusa w 1883 r. w celu podkreślenia znaczenia tej grupy związków. Termin pochodzi od greckiego słowa proteios,
Przegląd budowy i funkcji białek Co piszą o białkach? Wyraz wprowadzony przez Jönsa J. Berzeliusa w 1883 r. w celu podkreślenia znaczenia tej grupy związków. Termin pochodzi od greckiego słowa proteios,
Informacje. W sprawach organizacyjnych Slajdy z wykładów
 Biochemia Informacje W sprawach organizacyjnych malgorzata.dutkiewicz@wum.edu.pl Slajdy z wykładów www.takao.pl W sprawach merytorycznych Takao Ishikawa (takao@biol.uw.edu.pl) Kiedy? Co? Kto? 24 lutego
Biochemia Informacje W sprawach organizacyjnych malgorzata.dutkiewicz@wum.edu.pl Slajdy z wykładów www.takao.pl W sprawach merytorycznych Takao Ishikawa (takao@biol.uw.edu.pl) Kiedy? Co? Kto? 24 lutego
46 i 47. Wstęp do chemii -aminokwasów
 46 i 47. Wstęp do chemii -aminokwasów Chemia rganiczna, dr hab. inż. Mariola Koszytkowska-Stawińska, WChem PW; 2017/2018 1 21.1. Budowa ogólna -aminokwasów i klasyfikacja peptydów H 2 H H 2 R H R R 1 H
46 i 47. Wstęp do chemii -aminokwasów Chemia rganiczna, dr hab. inż. Mariola Koszytkowska-Stawińska, WChem PW; 2017/2018 1 21.1. Budowa ogólna -aminokwasów i klasyfikacja peptydów H 2 H H 2 R H R R 1 H
21. Wstęp do chemii a-aminokwasów
 21. Wstęp do chemii a-aminokwasów Chemia rganiczna, dr hab. inż. Mariola Koszytkowska-Stawińska, WChem PW; 2016/2017 1 21.1. Budowa ogólna a-aminokwasów i klasyfikacja peptydów H 2 N H kwas 2-aminooctowy
21. Wstęp do chemii a-aminokwasów Chemia rganiczna, dr hab. inż. Mariola Koszytkowska-Stawińska, WChem PW; 2016/2017 1 21.1. Budowa ogólna a-aminokwasów i klasyfikacja peptydów H 2 N H kwas 2-aminooctowy
Chemiczne składniki komórek
 Chemiczne składniki komórek Pierwiastki chemiczne w komórkach: - makroelementy (pierwiastki biogenne) H, O, C, N, S, P Ca, Mg, K, Na, Cl >1% suchej masy - mikroelementy Fe, Cu, Mn, Mo, B, Zn, Co, J, F
Chemiczne składniki komórek Pierwiastki chemiczne w komórkach: - makroelementy (pierwiastki biogenne) H, O, C, N, S, P Ca, Mg, K, Na, Cl >1% suchej masy - mikroelementy Fe, Cu, Mn, Mo, B, Zn, Co, J, F
Oznaczanie aktywności proteolitycznej trypsyny Zajęcia 3-godzinne część A, zajęcia 4-godzinne część A i B
 znaczanie aktywności proteolitycznej trypsyny Zajęcia 3-godzinne część A, zajęcia 4-godzinne część A i B el ćwiczenia elem ćwiczenia jest zapoznanie się z metodą oznaczania aktywności endopeptydaz na przykładzie
znaczanie aktywności proteolitycznej trypsyny Zajęcia 3-godzinne część A, zajęcia 4-godzinne część A i B el ćwiczenia elem ćwiczenia jest zapoznanie się z metodą oznaczania aktywności endopeptydaz na przykładzie
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 4 Jak działają geny?
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Wykład 4 Jak działają geny?
spektroskopia elektronowa (UV-vis)
") spektroskopia elektronowa (UV-vis) rodzaje przejść elektronowych Energia σ* π* 3 n 3 π σ σ σ* daleki nadfiolet (λ max < 200 nm) π π* bliski nadfiolet jednostki energii atomowa jednostka energii = energia
spektroskopia elektronowa (UV-vis) rodzaje przejść elektronowych Energia σ* π* 3 n 3 π σ σ σ* daleki nadfiolet (λ max < 200 nm) π π* bliski nadfiolet jednostki energii atomowa jednostka energii = energia
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Budowa aminokwasów i białek
 Biofizyka Ćwiczenia 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas Budowa aminokwasów i białek E.Banachowicz 1 Ogólna budowa aminokwasów α w neutralnym p α N 2 COO N
Biofizyka Ćwiczenia 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas Budowa aminokwasów i białek E.Banachowicz 1 Ogólna budowa aminokwasów α w neutralnym p α N 2 COO N
etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
 Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
Temat: Białka Aminy Pochodne węglowodorów zawierające grupę NH 2 Wzór ogólny amin: R NH 2 Przykład: CH 3 -CH 2 -NH 2 etyloamina Aminy mają właściwości zasadowe i w roztworach kwaśnych tworzą jon alkinowy
WYKŁAD 4: MOLEKULARNE MECHANIZMY BIOSYNTEZY BIAŁEK. Prof. dr hab. n. med. Małgorzata Milkiewicz Zakład Biologii Medycznej.
 Pierwsza litera Trzecia litera 2018-10-26 WYKŁAD 4: MOLEKULARNE MECHANIZMY BIOSYNTEZY BIAŁEK Prof. dr hab. n. med. Małgorzata Milkiewicz Zakład Biologii Medycznej Druga litera 1 Enancjomery para nienakładalnych
Pierwsza litera Trzecia litera 2018-10-26 WYKŁAD 4: MOLEKULARNE MECHANIZMY BIOSYNTEZY BIAŁEK Prof. dr hab. n. med. Małgorzata Milkiewicz Zakład Biologii Medycznej Druga litera 1 Enancjomery para nienakładalnych
Slajd 1. Slajd 2. Proteiny. Peptydy i białka są polimerami aminokwasów połączonych wiązaniem amidowym (peptydowym) Kwas α-aminokarboksylowy aminokwas
 Kwas α-aminokarboksylowy aminokwas") Slajd 1 Proteiny Slajd 2 Peptydy i białka są polimerami aminokwasów połączonych wiązaniem amidowym (peptydowym) wiązanie amidowe Kwas α-aminokarboksylowy aminokwas Slajd 3 Aminokwasy z alifatycznym łańcuchem
Slajd 1 Proteiny Slajd 2 Peptydy i białka są polimerami aminokwasów połączonych wiązaniem amidowym (peptydowym) wiązanie amidowe Kwas α-aminokarboksylowy aminokwas Slajd 3 Aminokwasy z alifatycznym łańcuchem
Aminokwasy, peptydy i białka. Związki wielofunkcyjne
 Aminokwasy, peptydy i białka Związki wielofunkcyjne Aminokwasy, peptydy i białka Aminokwasy, peptydy i białka: - wiadomości ogólne Aminokwasy: - ogólna charakterystyka - budowa i nazewnictwo - właściwości
Aminokwasy, peptydy i białka Związki wielofunkcyjne Aminokwasy, peptydy i białka Aminokwasy, peptydy i białka: - wiadomości ogólne Aminokwasy: - ogólna charakterystyka - budowa i nazewnictwo - właściwości
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii
 Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych aspekty praktyczne dr Małgorzata Kozłowska Ekspert
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych aspekty praktyczne dr Małgorzata Kozłowska Ekspert
Klonowanie molekularne Kurs doskonalący. Zakład Geriatrii i Gerontologii CMKP
 Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Sposób otrzymywania białek o właściwościach immunoregulatorowych. Przedmiotem wynalazku jest sposób otrzymywania fragmentów witellogeniny.
 1 Sposób otrzymywania białek o właściwościach immunoregulatorowych Przedmiotem wynalazku jest sposób otrzymywania fragmentów witellogeniny. Wynalazek może znaleźć zastosowanie w przemyśle spożywczym i
1 Sposób otrzymywania białek o właściwościach immunoregulatorowych Przedmiotem wynalazku jest sposób otrzymywania fragmentów witellogeniny. Wynalazek może znaleźć zastosowanie w przemyśle spożywczym i
Test kwalifikacyjny Lifescience dla licealistów 2015
 Test kwalifikacyjny Lifescience dla licealistów 2015 Imię nazwisko (pseudonim): 1. Daltonizm (d) jest cechą recesywną sprzężoną z płcią. Rudy kolor włosów (r) jest cechą autosomalną i recesywną w stosunku
Test kwalifikacyjny Lifescience dla licealistów 2015 Imię nazwisko (pseudonim): 1. Daltonizm (d) jest cechą recesywną sprzężoną z płcią. Rudy kolor włosów (r) jest cechą autosomalną i recesywną w stosunku
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE. Ewa Waszkowska ekspert UPRP
 WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
Budowa i funkcje białek
 Budowa i funkcje białek Białka Wszystkie organizmy zawierają białko Każdy organizm wytwarza własne białka Podstawowe składniki białek - aminokwasy Roślinne mogą wytwarzać aminokwasy ze związków nieorganicznych
Budowa i funkcje białek Białka Wszystkie organizmy zawierają białko Każdy organizm wytwarza własne białka Podstawowe składniki białek - aminokwasy Roślinne mogą wytwarzać aminokwasy ze związków nieorganicznych
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych
 Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Bioinformatyka. z sylabusu... (wykład monograficzny) wykład 1. E. Banachowicz. Wykład monograficzny Bioinformatyka.
 wykład 1. E. Banachowicz. Wykład monograficzny Bioinformatyka.") Bioinformatyka (wykład monograficzny) wykład 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas z sylabusu... Wykład 1, 2006 1 Co to jest Bioinformatyka? Zastosowanie technologii
Bioinformatyka (wykład monograficzny) wykład 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas z sylabusu... Wykład 1, 2006 1 Co to jest Bioinformatyka? Zastosowanie technologii
STRESZCZENIE ROZPRAWY DOKTORSKIEJ. In search of substrates and inhibitors of human cathepsin C. Design, chemical synthesis and biological studies
 Monika Łęgowska Katedra Biochemii Nr indeksu: 2541 STRESZCZENIE ROZPRAWY DOKTORSKIEJ In search of substrates and inhibitors of human cathepsin C. Design, chemical synthesis and biological studies Katepsyna
Monika Łęgowska Katedra Biochemii Nr indeksu: 2541 STRESZCZENIE ROZPRAWY DOKTORSKIEJ In search of substrates and inhibitors of human cathepsin C. Design, chemical synthesis and biological studies Katepsyna
Materiały pochodzą z Platformy Edukacyjnej Portalu www.szkolnictwo.pl
 Materiały pochodzą z Platformy Edukacyjnej Portalu www.szkolnictwo.pl Wszelkie treści i zasoby edukacyjne publikowane na łamach Portalu www.szkolnictwo.pl mogą byd wykorzystywane przez jego Użytkowników
Materiały pochodzą z Platformy Edukacyjnej Portalu www.szkolnictwo.pl Wszelkie treści i zasoby edukacyjne publikowane na łamach Portalu www.szkolnictwo.pl mogą byd wykorzystywane przez jego Użytkowników
protos (gr.) pierwszy protein/proteins (ang.)
 pierwszy protein/proteins (ang.)") Białka 1 protos (gr.) pierwszy protein/proteins (ang.) cząsteczki życia materiał budulcowy materii ożywionej oraz wirusów wielkocząsteczkowe biopolimery o masie od kilku tysięcy do kilku milionów jednostek
Białka 1 protos (gr.) pierwszy protein/proteins (ang.) cząsteczki życia materiał budulcowy materii ożywionej oraz wirusów wielkocząsteczkowe biopolimery o masie od kilku tysięcy do kilku milionów jednostek
PL B1. SANOFI-AVENTIS DEUTSCHLAND GmbH, Frankfurt nad Menem,DE ,DE, BUP 26/
 RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 196626 (21) Numer zgłoszenia: 326936 (22) Data zgłoszenia: 20.06.1998 (13) B1 (51) Int.Cl. C12N 15/17 (2006.01)
RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 196626 (21) Numer zgłoszenia: 326936 (22) Data zgłoszenia: 20.06.1998 (13) B1 (51) Int.Cl. C12N 15/17 (2006.01)
Zastosowanie metody Lowry ego do oznaczenia białka w cukrze białym
 Zastosowanie metody Lowry ego do oznaczenia białka w cukrze białym Dr inż. Bożena Wnuk Mgr inż. Anna Wysocka Seminarium Aktualne zagadnienia dotyczące jakości w przemyśle cukrowniczym Łódź 10 11 czerwca
Zastosowanie metody Lowry ego do oznaczenia białka w cukrze białym Dr inż. Bożena Wnuk Mgr inż. Anna Wysocka Seminarium Aktualne zagadnienia dotyczące jakości w przemyśle cukrowniczym Łódź 10 11 czerwca
ROZPORZĄDZENIE KOMISJI (UE) / z dnia r.
 / z dnia r.") KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
KOMISJA EUROPEJSKA Bruksela, dnia 29.5.2018 C(2018) 3193 final ROZPORZĄDZENIE KOMISJI (UE) / z dnia 29.5.2018 r. zmieniające rozporządzenie (WE) nr 847/2000 w odniesieniu do definicji pojęcia podobnego
IZOMERIA Izomery - związki o takim samym składzie lecz różniące się budową
 IZMERIA Izomery - związki o takim samym składzie lecz różniące się budową TAK zy atomy są tak samo połączone? NIE izomery konstytucyjne stereoizomery zy odbicie lustrzane daje się nałożyć na cząsteczkę?
IZMERIA Izomery - związki o takim samym składzie lecz różniące się budową TAK zy atomy są tak samo połączone? NIE izomery konstytucyjne stereoizomery zy odbicie lustrzane daje się nałożyć na cząsteczkę?
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU KSZTAŁT BIAŁEK.
 SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU KSZTAŁT BIAŁEK. SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy. 1. Karta
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU KSZTAŁT BIAŁEK. SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy. 1. Karta
BIOSYNTEZA ACYLAZY PENICYLINOWEJ. Ćwiczenia z Mikrobiologii Przemysłowej 2011
 BIOSYNTEZA ACYLAZY PENICYLINOWEJ Ćwiczenia z Mikrobiologii Przemysłowej 2011 Acylaza penicylinowa Enzym hydrolizuje wiązanie amidowe w penicylinach Reakcja przebiega wg schematu: acylaza Reszta: fenyloacetylowa
BIOSYNTEZA ACYLAZY PENICYLINOWEJ Ćwiczenia z Mikrobiologii Przemysłowej 2011 Acylaza penicylinowa Enzym hydrolizuje wiązanie amidowe w penicylinach Reakcja przebiega wg schematu: acylaza Reszta: fenyloacetylowa
Nowoczesne systemy ekspresji genów
 Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny
 Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny Zadanie 1 1 pkt. za prawidłowe podanie typów dla obydwu zwierząt oznaczonych literami A oraz B. A. ramienionogi, B. mięczaki A.
Zasady oceniania rozwiązań zadań 48 Olimpiada Biologiczna Etap centralny Zadanie 1 1 pkt. za prawidłowe podanie typów dla obydwu zwierząt oznaczonych literami A oraz B. A. ramienionogi, B. mięczaki A.
IZOLACJA KWASÓW NUKLEINOWYCH WARUNKI ZALICZENIA PRZEDMIOTU- 7 ECTS PRZEDMIOT PROGOWY!!!
 WARUNKI ZALICZENIA PRZEDMIOTU- 7 ECTS PRZEDMIOT PROGOWY!!! W1-4p W2-4p W3-4p W4-4p W5-4p W6-4p W7-4p W8-4p W9-4p W10-4p min 21p wyjściówka I 40p wyjściówka II 40p egzamin I egzamin II min 21p 60p 60p min
WARUNKI ZALICZENIA PRZEDMIOTU- 7 ECTS PRZEDMIOT PROGOWY!!! W1-4p W2-4p W3-4p W4-4p W5-4p W6-4p W7-4p W8-4p W9-4p W10-4p min 21p wyjściówka I 40p wyjściówka II 40p egzamin I egzamin II min 21p 60p 60p min
CEL ĆWICZENIA: Zapoznanie się z przykładową procedurą odsalania oczyszczanych preparatów enzymatycznych w procesie klasycznej filtracji żelowej.
 LABORATORIUM 3 Filtracja żelowa preparatu oksydazy polifenolowej (PPO) oczyszczanego w procesie wysalania siarczanem amonu z wykorzystaniem złoża Sephadex G-50 CEL ĆWICZENIA: Zapoznanie się z przykładową
LABORATORIUM 3 Filtracja żelowa preparatu oksydazy polifenolowej (PPO) oczyszczanego w procesie wysalania siarczanem amonu z wykorzystaniem złoża Sephadex G-50 CEL ĆWICZENIA: Zapoznanie się z przykładową
Bioinformatyka Laboratorium, 30h. Michał Bereta
 Laboratorium, 30h Michał Bereta mbereta@pk.edu.pl www.michalbereta.pl Zasady zaliczenia przedmiotu Kolokwia (3 4 ) Ocena aktywności i przygotowania Obecnośd Literatura, materiały i ewolucja molekularna
Laboratorium, 30h Michał Bereta mbereta@pk.edu.pl www.michalbereta.pl Zasady zaliczenia przedmiotu Kolokwia (3 4 ) Ocena aktywności i przygotowania Obecnośd Literatura, materiały i ewolucja molekularna
ĆWICZENIA Z BIOCHEMII
 ĆWICZENIA Z BIOCHEMII D U STUDENTfiW WYDZIAŁU LEKARSKIEGO Pod redakcją Piotra Laidlera, Barbary Piekarskiej, Marii Wróbel WYDAWNICTWO UNIWERSYTETU JAGIELLOŃSKIEGO ĆWICZENIA Z BIOCHEMII DLA STUDENTÓW WYDZIAŁU
ĆWICZENIA Z BIOCHEMII D U STUDENTfiW WYDZIAŁU LEKARSKIEGO Pod redakcją Piotra Laidlera, Barbary Piekarskiej, Marii Wróbel WYDAWNICTWO UNIWERSYTETU JAGIELLOŃSKIEGO ĆWICZENIA Z BIOCHEMII DLA STUDENTÓW WYDZIAŁU
Przemiana materii i energii - Biologia.net.pl
 Ogół przemian biochemicznych, które zachodzą w komórce składają się na jej metabolizm. Wyróżnia się dwa antagonistyczne procesy metabolizmu: anabolizm i katabolizm. Szlak metaboliczny w komórce, to szereg
Ogół przemian biochemicznych, które zachodzą w komórce składają się na jej metabolizm. Wyróżnia się dwa antagonistyczne procesy metabolizmu: anabolizm i katabolizm. Szlak metaboliczny w komórce, to szereg
Dr hab. Elżbieta Jankowska
 Dr hab. Elżbieta Jankowska 80-308 Gdańsk, ul. Wita Stwosza 63 tel. (+48 58) 5235044, e-mail: elzbieta.jankowska@ug.edu.pl Gdańsk, 25.05.2016 Recenzja rozprawy doktorskiej mgr Jadwigi Popow-Stellmaszyk,
Dr hab. Elżbieta Jankowska 80-308 Gdańsk, ul. Wita Stwosza 63 tel. (+48 58) 5235044, e-mail: elzbieta.jankowska@ug.edu.pl Gdańsk, 25.05.2016 Recenzja rozprawy doktorskiej mgr Jadwigi Popow-Stellmaszyk,
AMINOKWASY. I. Wprowadzenie teoretyczne. Aminokwasy są to związki, które w łańcuchu węglowym zawierają zarówno grupę aminową jak i grupę karboksylową.
 AMIKWASY I. Wprowadzenie teoretyczne Aminokwasy są to związki, które w łańcuchu węglowym zawierają zarówno grupę aminową jak i grupę karboksylową. 2 3 WZY GÓLE ATUALY AMIKWASÓW WYSTĘPUJĄY W BIAŁKA Zalicza
AMIKWASY I. Wprowadzenie teoretyczne Aminokwasy są to związki, które w łańcuchu węglowym zawierają zarówno grupę aminową jak i grupę karboksylową. 2 3 WZY GÓLE ATUALY AMIKWASÓW WYSTĘPUJĄY W BIAŁKA Zalicza
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2135881 Urząd Patentowy Rzeczypospolitej Polskiej (96) Data i numer zgłoszenia patentu europejskiego: 14.06.2006 09010789.7
Inżynieria Środowiska
 ROZTWORY BUFOROWE Roztworami buforowymi nazywamy takie roztwory, w których stężenie jonów wodorowych nie ulega większym zmianom ani pod wpływem rozcieńczania wodą, ani pod wpływem dodatku nieznacznych
ROZTWORY BUFOROWE Roztworami buforowymi nazywamy takie roztwory, w których stężenie jonów wodorowych nie ulega większym zmianom ani pod wpływem rozcieńczania wodą, ani pod wpływem dodatku nieznacznych
IZOLACJA KWASÓW NUKLEINOWYCH WARUNKI ZALICZENIA PRZEDMIOTU- 7 ECTS PRZEDMIOT PROGOWY!!!
 WARUNKI ZALICZENIA PRZEDMIOTU- 7 ECTS PRZEDMIOT PROGOWY!!! W1-4p W2-4p W3-4p W4-4p W5-4p W6-4p W7-4p W8-4p W9-4p W10-4p min 21p wyjściówka I 40p wyjściówka II 40p egzamin I egzamin II min 21p 60p 60p min
WARUNKI ZALICZENIA PRZEDMIOTU- 7 ECTS PRZEDMIOT PROGOWY!!! W1-4p W2-4p W3-4p W4-4p W5-4p W6-4p W7-4p W8-4p W9-4p W10-4p min 21p wyjściówka I 40p wyjściówka II 40p egzamin I egzamin II min 21p 60p 60p min
Dopasowanie sekwencji (sequence alignment)
") Co to jest alignment? Dopasowanie sekwencji (sequence alignment) Alignment jest sposobem dopasowania struktur pierwszorzędowych DNA, RNA lub białek do zidentyfikowanych regionów w celu określenia podobieństwa;
Co to jest alignment? Dopasowanie sekwencji (sequence alignment) Alignment jest sposobem dopasowania struktur pierwszorzędowych DNA, RNA lub białek do zidentyfikowanych regionów w celu określenia podobieństwa;
PL B1. UNIWERSYTET IM. ADAMA MICKIEWICZA W POZNANIU, Poznań, PL BUP 24/17
 RZECZPOSPOLITA POLSKA (2) OPIS PATENTOWY (9) PL () 229709 (3) B (2) Numer zgłoszenia: 49663 (5) Int.Cl. C07F 7/30 (2006.0) Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 05.2.206 (54)
RZECZPOSPOLITA POLSKA (2) OPIS PATENTOWY (9) PL () 229709 (3) B (2) Numer zgłoszenia: 49663 (5) Int.Cl. C07F 7/30 (2006.0) Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 05.2.206 (54)
Bioinformatyka Laboratorium, 30h. Michał Bereta
 Laboratorium, 30h Michał Bereta mbereta@pk.edu.pl www.michalbereta.pl Zasady zaliczenia przedmiotu Kolokwia (3 4 ) Ocena aktywności i przygotowania Obecność Literatura, materiały Bioinformatyka i ewolucja
Laboratorium, 30h Michał Bereta mbereta@pk.edu.pl www.michalbereta.pl Zasady zaliczenia przedmiotu Kolokwia (3 4 ) Ocena aktywności i przygotowania Obecność Literatura, materiały Bioinformatyka i ewolucja
Właściwości aminokwasów i białek
 Właściwości aminokwasów i białek el ćwiczenia Ćwiczenie ma na celu poznanie niektórych typowych reakcji aminokwasów i białek. Reakcje te pozwalają odróżnić wolne aminokwasy od białek i innych związków
Właściwości aminokwasów i białek el ćwiczenia Ćwiczenie ma na celu poznanie niektórych typowych reakcji aminokwasów i białek. Reakcje te pozwalają odróżnić wolne aminokwasy od białek i innych związków
MACIERZE MUTACYJNE W ANALIZIE GENOMÓW czy możliwa jest rekonstrukcja filogenetyczna? Aleksandra Nowicka
 MAIERZE MUTAYJNE W ANALIZIE GENOMÓW czy możliwa jest rekonstrukcja filogenetyczna? Aleksandra Nowicka Zadaniem FILOGENETYKI jest : zrekonstruowanie ewolucyjnej historii wszystkich organizmów odkrycie przodka
MAIERZE MUTAYJNE W ANALIZIE GENOMÓW czy możliwa jest rekonstrukcja filogenetyczna? Aleksandra Nowicka Zadaniem FILOGENETYKI jest : zrekonstruowanie ewolucyjnej historii wszystkich organizmów odkrycie przodka
Translacja i proteom komórki
 Translacja i proteom komórki 1. Kod genetyczny 2. Budowa rybosomów 3. Inicjacja translacji 4. Elongacja translacji 5. Terminacja translacji 6. Potranslacyjne zmiany polipeptydów 7. Translacja a retikulum
Translacja i proteom komórki 1. Kod genetyczny 2. Budowa rybosomów 3. Inicjacja translacji 4. Elongacja translacji 5. Terminacja translacji 6. Potranslacyjne zmiany polipeptydów 7. Translacja a retikulum
Związki biologicznie aktywne
 Związki biologicznie aktywne Tematyka wykładów 1. Chemia związków biologicznie aktywnych pojęcia ogólne, klasyfikacja leków. 2. Związki biologicznie aktywne jako związki pochodzenia naturalnego, półsyntetycznego
Związki biologicznie aktywne Tematyka wykładów 1. Chemia związków biologicznie aktywnych pojęcia ogólne, klasyfikacja leków. 2. Związki biologicznie aktywne jako związki pochodzenia naturalnego, półsyntetycznego
Pytania Egzamin magisterski
 Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
Pytania Egzamin magisterski Międzyuczelniany Wydział Biotechnologii UG i GUMed 1. Krótko omów jakie informacje powinny być zawarte w typowych rozdziałach publikacji naukowej: Wstęp, Materiały i Metody,
ĆWICZENIE 1 i 2 Modyfikacja geu wołowej beta-laktoglobuliny przy użyciu metody Overlap Extension PCR (wydłużania nakładających się odcinków)
") ĆWICZENIE 1 i 2 Modyfikacja geu wołowej beta-laktoglobuliny przy użyciu metody Overlap Extension PCR (wydłużania nakładających się odcinków) Celem ćwiczenia jest wprowadzenie mutacji punktowej do genu
ĆWICZENIE 1 i 2 Modyfikacja geu wołowej beta-laktoglobuliny przy użyciu metody Overlap Extension PCR (wydłużania nakładających się odcinków) Celem ćwiczenia jest wprowadzenie mutacji punktowej do genu
RT31-020, RT , MgCl 2. , random heksamerów X 6
 RT31-020, RT31-100 RT31-020, RT31-100 Zestaw TRANSCRIPTME RNA zawiera wszystkie niezbędne składniki do przeprowadzenia syntezy pierwszej nici cdna na matrycy mrna lub całkowitego RNA. Uzyskany jednoniciowy
RT31-020, RT31-100 RT31-020, RT31-100 Zestaw TRANSCRIPTME RNA zawiera wszystkie niezbędne składniki do przeprowadzenia syntezy pierwszej nici cdna na matrycy mrna lub całkowitego RNA. Uzyskany jednoniciowy
Making the impossible possible: the metamorphosis of Polish Biology Olympiad
 Making the impossible possible: the metamorphosis of Polish Biology Olympiad Takao Ishikawa Faculty of Biology, University of Warsaw, Poland Performance of Polish students at IBO Gold Silver Bronze Merit
Making the impossible possible: the metamorphosis of Polish Biology Olympiad Takao Ishikawa Faculty of Biology, University of Warsaw, Poland Performance of Polish students at IBO Gold Silver Bronze Merit
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA
 Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa IZOLACJA DNA Z HODOWLI KOMÓRKOWEJ.
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa IZOLACJA DNA Z HODOWLI KOMÓRKOWEJ.
48 Olimpiada Biologiczna Pracownia biochemiczna zasady oceniania rozwiązań zadań
 48 Olimpiada Biologiczna Pracownia biochemiczna zasady oceniania rozwiązań zadań Część A. Rozdzielanie i identyfikacja aminokwasów (0 20 pkt) Zadanie A.1 (0 6 pkt) Wskazanie kolejności użycia roztworów
48 Olimpiada Biologiczna Pracownia biochemiczna zasady oceniania rozwiązań zadań Część A. Rozdzielanie i identyfikacja aminokwasów (0 20 pkt) Zadanie A.1 (0 6 pkt) Wskazanie kolejności użycia roztworów
Struktura i funkcja białek (I mgr)
") Struktura i funkcja białek (I mgr) Dr Filip Jeleń fj@protein.pl http://www.protein.pl/ Jeremy M. Berg, John L. Tymoczko, Lubert Stryer Biochemia Carl Branden, John Tooze Introduction to Protein Structure
Struktura i funkcja białek (I mgr) Dr Filip Jeleń fj@protein.pl http://www.protein.pl/ Jeremy M. Berg, John L. Tymoczko, Lubert Stryer Biochemia Carl Branden, John Tooze Introduction to Protein Structure
Laboratorium 5. Wpływ temperatury na aktywność enzymów. Inaktywacja termiczna
 Laboratorium 5 Wpływ temperatury na aktywność enzymów. Inaktywacja termiczna Prowadzący: dr inż. Karolina Labus 1. CZĘŚĆ TEORETYCZNA Szybkość reakcji enzymatycznej zależy przede wszystkim od stężenia substratu
Laboratorium 5 Wpływ temperatury na aktywność enzymów. Inaktywacja termiczna Prowadzący: dr inż. Karolina Labus 1. CZĘŚĆ TEORETYCZNA Szybkość reakcji enzymatycznej zależy przede wszystkim od stężenia substratu
-1- Preparat o właściwościach immunoregulatorowych
 -1- Preparat o właściwościach immunoregulatorowych Przedmiotem wynalazku jest preparat o właściwościach immunoregulatorowych. Wynalazek może znaleźć zastosowanie w przemyśle spożywczym i farmaceutycznym,
-1- Preparat o właściwościach immunoregulatorowych Przedmiotem wynalazku jest preparat o właściwościach immunoregulatorowych. Wynalazek może znaleźć zastosowanie w przemyśle spożywczym i farmaceutycznym,
PL B1. Sposób usuwania zanieczyszczeń z instalacji produkcyjnych zawierających membrany filtracyjne stosowane w przemyśle spożywczym
 PL 214736 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 214736 (13) B1 (21) Numer zgłoszenia: 388142 (51) Int.Cl. B01D 65/06 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia:
PL 214736 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 214736 (13) B1 (21) Numer zgłoszenia: 388142 (51) Int.Cl. B01D 65/06 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia:
Struktura biomakromolekuł chemia biologiczna III rok
 truktura biomakromolekuł chemia biologiczna III rok jak są zbudowane białka? dlaczego białka są tak zbudowane? co z tego wynika? 508 13 604 liczba struktur dostępnych w Protein Data Bank wynosi aktualnie
truktura biomakromolekuł chemia biologiczna III rok jak są zbudowane białka? dlaczego białka są tak zbudowane? co z tego wynika? 508 13 604 liczba struktur dostępnych w Protein Data Bank wynosi aktualnie
CHARAKTERYSTYKA PRODUKTU LECZNICZEGO
 CHARAKTERYSTYKA PRODUKTU LECZNICZEGO 1. NAZWA PRODUKTU LECZNICZEGO Vaminolact, roztwór do infuzji 2. SKŁAD JAKOŚCIOWY I ILOŚCIOWY 1000 ml roztworu zawiera: Substancje czynne: L-alanina L-arginina L-asparaginowy
CHARAKTERYSTYKA PRODUKTU LECZNICZEGO 1. NAZWA PRODUKTU LECZNICZEGO Vaminolact, roztwór do infuzji 2. SKŁAD JAKOŚCIOWY I ILOŚCIOWY 1000 ml roztworu zawiera: Substancje czynne: L-alanina L-arginina L-asparaginowy
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii
 Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
Inwestycja w przyszłość czyli znaczenie ochrony własności przemysłowej dla współczesnej biotechnologii Zdolność patentowa wynalazków biotechnologicznych dr Małgorzata Kozłowska Ekspert w UPRP dr Ewa Waszkowska
(Tekst mający znaczenie dla EOG)
") L 53/134 PL 23.2.2018 ROZPORZĄDZENIE WYKONAWCZE KOMISJI (UE) 2018/249 z dnia 15 lutego 2018 r. dotyczące na stosowanie tauryny, beta-alaniny, L-alaniny, L-argininy, kwasu L-asparaginowego, L-histydyny,
L 53/134 PL 23.2.2018 ROZPORZĄDZENIE WYKONAWCZE KOMISJI (UE) 2018/249 z dnia 15 lutego 2018 r. dotyczące na stosowanie tauryny, beta-alaniny, L-alaniny, L-argininy, kwasu L-asparaginowego, L-histydyny,
Novabeads Food DNA Kit
 Novabeads Food DNA Kit Novabeads Food DNA Kit jest nowej generacji narzędziem w technikach biologii molekularnej, umożliwiającym izolację DNA z produktów spożywczych wysoko przetworzonych. Metoda oparta
Novabeads Food DNA Kit Novabeads Food DNA Kit jest nowej generacji narzędziem w technikach biologii molekularnej, umożliwiającym izolację DNA z produktów spożywczych wysoko przetworzonych. Metoda oparta
WPROWADZENIE DO GENETYKI MOLEKULARNEJ
 WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
(13) B1 PL B1. Hoechst Aktiengesellschaft, Frankfurt nad Menem, DE. Gugała Barbara, PATPOL Spółka z o. o.
 B1 PL B1. Hoechst Aktiengesellschaft, Frankfurt nad Menem, DE. Gugała Barbara, PATPOL Spółka z o. o.") RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 169178 (13) B1 (2 1) Numer zgłoszenia 289953 Urząd Patentowy (22) Data zgłoszenia 19.04.1991 Rzeczypospolitej Polskiej (51) IntCl6 C12N 15/62 C12N
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 169178 (13) B1 (2 1) Numer zgłoszenia 289953 Urząd Patentowy (22) Data zgłoszenia 19.04.1991 Rzeczypospolitej Polskiej (51) IntCl6 C12N 15/62 C12N
Definicja immobilizacji
 Definicja immobilizacji Immobilizacja technika unieruchamiania biokatalizatorów / enzymów na nośnikach, stosowana powszechnie w badaniach naukowych i przemyśle (chemicznym, spożywczym) Problemy związane
Definicja immobilizacji Immobilizacja technika unieruchamiania biokatalizatorów / enzymów na nośnikach, stosowana powszechnie w badaniach naukowych i przemyśle (chemicznym, spożywczym) Problemy związane
WPROWADZENIE DO GENETYKI MOLEKULARNEJ
 WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
WPROWADZENIE DO GENETYKI MOLEKULARNEJ Replikacja organizacja widełek replikacyjnych Transkrypcja i biosynteza białek Operon regulacja ekspresji genów Prowadzący wykład: prof. dr hab. Jarosław Burczyk REPLIKACJA
Ćwiczenie nr 7. Aminokwasy i peptydy. Repetytorium. Repetytorium
 Repetytorium Ćwiczenie nr 7 dr Mariola Krawiecka Aminokwasy i peptydy 1. Podział aminokwasów. 2. Właściwości aminokwasów-aminokwasy jako jony obojnacze. 3. Reaktywność aminokwasów. 4. Biologicznie ważne
Repetytorium Ćwiczenie nr 7 dr Mariola Krawiecka Aminokwasy i peptydy 1. Podział aminokwasów. 2. Właściwości aminokwasów-aminokwasy jako jony obojnacze. 3. Reaktywność aminokwasów. 4. Biologicznie ważne
Drożdżowe systemy ekspresyjne
 Drożdże Drożdżowe systemy ekspresyjne Zalety: możliwość uzyskania dużej biomasy modyfikacje postranslacyjne eksprymowanych białek transport eksprymowanych białek do pożywki Duża biomasa W przypadku hodowli
Drożdże Drożdżowe systemy ekspresyjne Zalety: możliwość uzyskania dużej biomasy modyfikacje postranslacyjne eksprymowanych białek transport eksprymowanych białek do pożywki Duża biomasa W przypadku hodowli
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA
 Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa tel. 22 572 0735, 606448502
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa tel. 22 572 0735, 606448502
Łódź Łódź, ul. Żeromskiego 116 Prof. dr hab. inż. Aleksandra Olma
 90-924 Łódź Łódź, 10. 10. 2016 ul. Żeromskiego 116 Prof. dr hab. inż. Aleksandra Olma e-mail: aleksandra.olma@p.lodz.pl Recenzja rozprawy doktorskiej mgr Agaty Gitlin-Domagalskiej DESIGNING AND CHEMICAL
90-924 Łódź Łódź, 10. 10. 2016 ul. Żeromskiego 116 Prof. dr hab. inż. Aleksandra Olma e-mail: aleksandra.olma@p.lodz.pl Recenzja rozprawy doktorskiej mgr Agaty Gitlin-Domagalskiej DESIGNING AND CHEMICAL
PL B1. AKADEMIA GÓRNICZO-HUTNICZA IM. STANISŁAWA STASZICA W KRAKOWIE, Kraków, PL BUP 14/12
 PL 218561 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 218561 (13) B1 (21) Numer zgłoszenia: 393413 (51) Int.Cl. G01N 27/02 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia:
PL 218561 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 218561 (13) B1 (21) Numer zgłoszenia: 393413 (51) Int.Cl. G01N 27/02 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia:
Oznaczanie aktywności proteolitycznej trypsyny metodą Ansona
 Oznaczanie aktywności proteolitycznej trypsyny metodą Ansona Wymagane zagadnienia teoretyczne 1. Enzymy proteolityczne, klasyfikacja, rola biologiczna. 2. Enzymy proteolityczne krwi. 3. Wewnątrzkomórkowa
Oznaczanie aktywności proteolitycznej trypsyny metodą Ansona Wymagane zagadnienia teoretyczne 1. Enzymy proteolityczne, klasyfikacja, rola biologiczna. 2. Enzymy proteolityczne krwi. 3. Wewnątrzkomórkowa
Ćwiczenie laboratoryjne nr 4 dla e-rolnictwa (20 i 22 - skrypt).
.") Ćwiczenie laboratoryjne nr 4 dla e-rolnictwa (20 i 22 - skrypt). Oznaczanie aktywności proteolitycznej metodą Ansona na przykładzie trypsyny. Cel ćwiczenia Celem ćwiczenia jest poznanie metody oznaczania
Ćwiczenie laboratoryjne nr 4 dla e-rolnictwa (20 i 22 - skrypt). Oznaczanie aktywności proteolitycznej metodą Ansona na przykładzie trypsyny. Cel ćwiczenia Celem ćwiczenia jest poznanie metody oznaczania
Bioinformatyka. z sylabusu...
 Bioinformatyka Wykład 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas z sylabusu... Wykład 1, 2008 1 Co to jest Bioinformatyka? Zastosowanie technologii informacji do
Bioinformatyka Wykład 1. E. Banachowicz Zakład Biofizyki Molekularnej IF UAM http://www.amu.edu.pl/~ewas z sylabusu... Wykład 1, 2008 1 Co to jest Bioinformatyka? Zastosowanie technologii informacji do
KLONOWANIE DNA REKOMBINACJA DNA WEKTORY
 KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
Model : - SCITEC 100% Whey Protein Professional 920g
 Białka > Model : - Producent : Scitec 100% Whey Protein Professional - jest najwyższej jakości, wolnym od laktozy, czystym koncentratem i izolat białek serwatkowych (WPC + WPI) o bardzo dobrej rozpuszczalności
Białka > Model : - Producent : Scitec 100% Whey Protein Professional - jest najwyższej jakości, wolnym od laktozy, czystym koncentratem i izolat białek serwatkowych (WPC + WPI) o bardzo dobrej rozpuszczalności
Aminokwasy, peptydy, białka
 Aminokwasy, peptydy, białka Aminokwasy KWAS 1-AMINOCYKLOPROPANOKARBOKSYLOWY α AMINOKWAS KWAS 3-AMINOPROPANOWY β AMINOKWAS KWAS 4-AMINOPROPANOWY γ AMINOKWAS KWAS 2-AMINOETANOSULFONOWY β AMINOKWAS Aminokwasy
Aminokwasy, peptydy, białka Aminokwasy KWAS 1-AMINOCYKLOPROPANOKARBOKSYLOWY α AMINOKWAS KWAS 3-AMINOPROPANOWY β AMINOKWAS KWAS 4-AMINOPROPANOWY γ AMINOKWAS KWAS 2-AMINOETANOSULFONOWY β AMINOKWAS Aminokwasy
OFERTA TEMATÓW PROJEKTÓW DYPLOMOWYCH (MAGISTERSKICH) do zrealizowania w Katedrze INŻYNIERII CHEMICZNEJ I PROCESOWEJ
 do zrealizowania w Katedrze INŻYNIERII CHEMICZNEJ I PROCESOWEJ") OFERTA TEMATÓW PROJEKTÓW DYPLOMOWYCH (MAGISTERSKICH) do zrealizowania w Katedrze INŻYNIERII CHEMICZNEJ I PROCESOWEJ Badania kinetyki utleniania wybranych grup związków organicznych podczas procesów oczyszczania
OFERTA TEMATÓW PROJEKTÓW DYPLOMOWYCH (MAGISTERSKICH) do zrealizowania w Katedrze INŻYNIERII CHEMICZNEJ I PROCESOWEJ Badania kinetyki utleniania wybranych grup związków organicznych podczas procesów oczyszczania
TATA box. Enhancery. CGCG ekson intron ekson intron ekson CZĘŚĆ KODUJĄCA GENU TERMINATOR. Elementy regulatorowe
 Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
A. B. Co warto wiedzieć o aminokwasach?
 A. B. Co warto wiedzieć o aminokwasach? a) 1. Cele lekcji a. a) Wiadomości Uczeń zna pojęcia: asymetryczny atom węgla, L-aminokwas, peptyd, reakcja kondensacji, aminokwas białkowy, aminokwas egzogenny
A. B. Co warto wiedzieć o aminokwasach? a) 1. Cele lekcji a. a) Wiadomości Uczeń zna pojęcia: asymetryczny atom węgla, L-aminokwas, peptyd, reakcja kondensacji, aminokwas białkowy, aminokwas egzogenny
SPIS TREŚCI OD AUTORÓW... 5
 SPIS TREŚCI OD AUTORÓW... 5 BIAŁKA 1. Wprowadzenie... 7 2. Aminokwasy jednostki strukturalne białek... 7 2.1. Klasyfikacja aminokwasów... 9 2.1.1. Aminokwasy białkowe i niebiałkowe... 9 2.1.2. Zdolność
SPIS TREŚCI OD AUTORÓW... 5 BIAŁKA 1. Wprowadzenie... 7 2. Aminokwasy jednostki strukturalne białek... 7 2.1. Klasyfikacja aminokwasów... 9 2.1.1. Aminokwasy białkowe i niebiałkowe... 9 2.1.2. Zdolność
Metabolizm białek. Ogólny schemat metabolizmu bialek
 Metabolizm białek Ogólny schemat metabolizmu bialek Trawienie białek i absorpcja aminokwasów w przewodzie pokarmowym w żołądku (niskie ph ~2, rola HCl)- hydratacja, homogenizacja, denaturacja białek i
Metabolizm białek Ogólny schemat metabolizmu bialek Trawienie białek i absorpcja aminokwasów w przewodzie pokarmowym w żołądku (niskie ph ~2, rola HCl)- hydratacja, homogenizacja, denaturacja białek i
Wykład 14 Biosynteza białek
 BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
Bioinformatyka wykład 9
 Bioinformatyka wykład 9 14.XII.21 białkowa bioinformatyka strukturalna krzysztof_pawlowski@sggw.pl 211-1-17 1 Plan wykładu struktury białek dlaczego? struktury białek geometria i fizyka modyfikacje kowalencyjne
Bioinformatyka wykład 9 14.XII.21 białkowa bioinformatyka strukturalna krzysztof_pawlowski@sggw.pl 211-1-17 1 Plan wykładu struktury białek dlaczego? struktury białek geometria i fizyka modyfikacje kowalencyjne
Roztwory buforowe (bufory) (opracowanie: dr Katarzyna Makyła-Juzak)
 (opracowanie: dr Katarzyna Makyła-Juzak)") Roztwory buforowe (bufory) (opracowanie: dr Katarzyna Makyła-Juzak) 1. Właściwości roztworów buforowych Dodatek nieznacznej ilości mocnego kwasu lub mocnej zasady do czystej wody powoduje stosunkowo dużą
Roztwory buforowe (bufory) (opracowanie: dr Katarzyna Makyła-Juzak) 1. Właściwości roztworów buforowych Dodatek nieznacznej ilości mocnego kwasu lub mocnej zasady do czystej wody powoduje stosunkowo dużą
Technika fluorescencyjnego oznaczania aktywności enzymów. Wstęp:
 Technika fluorescencyjnego oznaczania aktywności enzymów Wstęp: Wśród technik biologii molekularnej jedną z najczęściej stosowanych jest technika fluorescencyjna. Stosując technikę fluorescencyjną można
Technika fluorescencyjnego oznaczania aktywności enzymów Wstęp: Wśród technik biologii molekularnej jedną z najczęściej stosowanych jest technika fluorescencyjna. Stosując technikę fluorescencyjną można
Ogólna budowa aminokwasów
 H w neutralnym ph H NH 3 + C COO - NH 2 C COOH R R grupa aminowa - NH 2 grupa karboksylowa - COOH Gly, G Ala, A Ogólna budowa aminokwasów Reguła CORN H R NH 3 + C L lewoskrętny (COO-R-N) COO - 1 20 aminokwasów
H w neutralnym ph H NH 3 + C COO - NH 2 C COOH R R grupa aminowa - NH 2 grupa karboksylowa - COOH Gly, G Ala, A Ogólna budowa aminokwasów Reguła CORN H R NH 3 + C L lewoskrętny (COO-R-N) COO - 1 20 aminokwasów
Mechanizm działania buforów *
 Mechanizm działania buforów * UNIWERSYTET PRZYRODNICZY Z doświadczenia nabytego w laboratorium wiemy, że dodanie kropli stężonego kwasu do 10 ml wody powoduje gwałtowny spadek ph o kilka jednostek. Tymczasem
Mechanizm działania buforów * UNIWERSYTET PRZYRODNICZY Z doświadczenia nabytego w laboratorium wiemy, że dodanie kropli stężonego kwasu do 10 ml wody powoduje gwałtowny spadek ph o kilka jednostek. Tymczasem
Wykład 2. Kinetyka reakcji enzymatycznych.
 Wykład 2 Kinetyka reakcji enzymatycznych. Kofaktory enzymów wd_2 2 Ryboflawina witamina B 2 Ryboflawina wit. B 2 FAD dinukleotyd flawinoadeninowy wd_2 3 Niacyna witamina PP (B 3 ) NAD + dinukleotyd nikotynamidoadeninowy
Wykład 2 Kinetyka reakcji enzymatycznych. Kofaktory enzymów wd_2 2 Ryboflawina witamina B 2 Ryboflawina wit. B 2 FAD dinukleotyd flawinoadeninowy wd_2 3 Niacyna witamina PP (B 3 ) NAD + dinukleotyd nikotynamidoadeninowy
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Ekspresja genów jest regulowana
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Ekspresja genów jest regulowana
Substancje o Znaczeniu Biologicznym
 Substancje o Znaczeniu Biologicznym Tłuszcze Jadalne są to tłuszcze, które może spożywać człowiek. Stanowią ważny, wysokoenergetyczny składnik diety. Z chemicznego punktu widzenia głównym składnikiem tłuszczów
Substancje o Znaczeniu Biologicznym Tłuszcze Jadalne są to tłuszcze, które może spożywać człowiek. Stanowią ważny, wysokoenergetyczny składnik diety. Z chemicznego punktu widzenia głównym składnikiem tłuszczów
PRZYKŁADOWE ZADANIA ORGANICZNE ZWIĄZKI ZAWIERAJĄCE AZOT
 PRZYKŁADOWE ZADANIA ORGANICZNE ZWIĄZKI ZAWIERAJĄCE AZOT Zadanie 1127 (1 pkt) Uszereguj podane związki według rosnącego ph w roztworze wodnym. Właściwy porządek podaj zapisując go wzorami półstrukturalnymi.
PRZYKŁADOWE ZADANIA ORGANICZNE ZWIĄZKI ZAWIERAJĄCE AZOT Zadanie 1127 (1 pkt) Uszereguj podane związki według rosnącego ph w roztworze wodnym. Właściwy porządek podaj zapisując go wzorami półstrukturalnymi.
AMINO MAX kaps - Trec Nutrition
 Dane aktualne na dzień: 27-01-2018 14:38 Link do produktu: https://sportowesuplementy.pl/amino-max-6800-320kaps-trec-nutrition-p-32.html AMINO MAX 6800 320kaps - Trec Nutrition Cena Dostępność Czas wysyłki
Dane aktualne na dzień: 27-01-2018 14:38 Link do produktu: https://sportowesuplementy.pl/amino-max-6800-320kaps-trec-nutrition-p-32.html AMINO MAX 6800 320kaps - Trec Nutrition Cena Dostępność Czas wysyłki
Lek od pomysłu do wdrożenia
 Lek od pomysłu do wdrożenia Lek od pomysłu do wdrożenia KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU
Lek od pomysłu do wdrożenia Lek od pomysłu do wdrożenia KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU KRÓTKA HISTORIA LEKU
Komputerowe wspomaganie projektowanie leków
 Komputerowe wspomaganie projektowanie leków wykład II Prof. dr hab. Sławomir Filipek Grupa BIOmodelowania Uniwersytet Warszawski, Wydział Chemii oraz Centrum Nauk Biologiczno-Chemicznych Cent-III www.biomodellab.eu
Komputerowe wspomaganie projektowanie leków wykład II Prof. dr hab. Sławomir Filipek Grupa BIOmodelowania Uniwersytet Warszawski, Wydział Chemii oraz Centrum Nauk Biologiczno-Chemicznych Cent-III www.biomodellab.eu