(12) OPIS PATENTOWY (19) PL (11)
|
|
|
- Kacper Bielecki
- 8 lat temu
- Przeglądów:
Transkrypt
1 RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) (21) Numer zgłoszenia: (22) Data zgłoszenia (86) Data i numer zgłoszenia międzynarodowego , PCT/IB98/00821 (87) Data i numer publikacji zgłoszenia międzynarodowego: , W098/51800, PCT Gazette nr 46/98 (13) B1 (5 1) IntCl7 C12N 15/82 Opis patentow y p rzedrukow ano ze w zględu na zauw ażone błędy (54) Sposób modyfikowania kwitnienia w roślinach (30) Pierwszeństwo: ,GB, (73) Uprawniony z patentu: ADVANTA SEEDS B V, Kapelle, NL (43) Zgłoszenie ogłoszono: BUP 14/00 (72) Twórcy wynalazku: Sjef Smeekens, Dnebergen, NL Peter Weisbeek, Den Dolder, NL Marcel Proveniers, Utrecht, NL (45) O udzieleniu patentu ogłoszono: WUP 11/05 (74) Pełnomocnik: Ja nina Kossowska, PATPOL Sp. z o.o. (57) 1. Sposób modyfikowania kwitnienia w roślinach, znamienny tym, że obejmuje transformowanie roślin konstruktem zawierającym pełną albo częściową sekwencję DNA kodującą produkt ATH1 pod kontrolą promotora działającego w roślinach. PL B1
 Uprawniony z patentu: ADVANTA SEEDS B V, Kapelle, NL (43) Zgłoszenie ogłoszono: 17.07.")
2 Sposób m odyfikowania kwitnienia w roślinach Zastrzeżenia patentowe 1. Sposób modyfikowania kwitnienia w roślinach, znamienny tym, ze obejmuje transformowanie roślin konstruktem zawierającym pełną albo częściową sekwencję DNA kodującą produkt ATH1 pod kontrolą promotora działającego w roślinach. 2. Sposób według zastrz. 1, znamienny tym, ze proces kwitnienia w roślinach jest promowany przez transformowanie roślin konstniktem, który hamuje wytwarzanie białka ATH1. 3. Sposób według zastrz. 2, znamienny tym, że konstrukt jest przystosowany do ekspresji antysensownego RNA do RNA wytwarzanego przez gen ATH1. 4. Sposób według zastrz. 1, znamienny tym, że proces kwitnienia w roślinach jest opóźniany przez transformowanie roślin konstruktem, który promuje wytwarzanie zrekombinowanego białka ATH1. * * * Przedmiotem wynalazku jest sposób modyfikowania kwitnienia w roślinach obejmujący transformowanie roślin konstruktem zawierającym pełną albo częściową sekwencję DNA kodującą produkt ATH1 pod kontrolą promotora działającego w roślinach. Rośliny różnią się od zwierząt. Ciało dojrzałej rośliny tworzy się postembrionalnie przez ciągłą aktywność merystemów wierzchołkowych pędu i korzenia. Merystem wierzchołkowy pędu tworzy się w czasie embriogenezy rośliny i razem z liścieniami, hipokotylem, korzeniem zarodkowym i merystemem korzeniowym wytycza podstawowy plan budowy rośliny. Merystem wierzchołkowy korzenia powstaje jako zgrupowanie około stu komórek i jest źródłem całej nadziemnej części rośliny. W czasie wegetatywnej fazy rozwoju rośliny merystem ten daje początek liściom (rozeta), łodydze i spoczynkowym merystemom bocznym. Po tym następuje tworzenie się kwiatostanów drugorzędowych, liści łodygowych i ostatecznych merystemów kwiatowych po indukcji kwitnienia. Kwitnienie obejmuje kompleks oddziaływań produktów genów, które regulują zmianę tożsamości merystemu pędu. Czynnikami określającymi poziomy ekspresji tych genów są genotyp i bodźce środowiskowe, takie jak fotoperiod, temperatura i jakość światła. W dużej mierze jest nadal nieznane, w jaki sposób jest uzyskiwana ta zmiana pod wpływem bodźców. Jednym z najważniejszych zdarzeń w cyklu życiowym rośliny jest decyzja wejścia w fazę rozrodczą. Szeroki zakres sygnałów środowiskowych i endogennych kontroluje przejście z fazy wegetatywnej do fazy rozrodczej. Ważnymi sygnałami są długość dnia, temperatura (wernalizacja), dostępność wody i czynników odżywczych oraz szereg fitohormonów, a w szczególności giberelin (GA). Sygnały te indukują zmianę tożsamości wegetatywnego merystemu wierzchołkowego, zwaną przejściem do kwitnienia i to przejście stanowi o powstaniu merystemu kwiatowego. Podczas gdy produktem wegetatywnego merystemu wierzchołkowego są zawiązki liści, merystem kwiatowy wytwarza zawiązki, które różnicują się w kwiatostany drugorzędowe w czasie wczesnego wzrostu generatywnego i w kwiaty później w tym stadium. W badaniach dotyczących hodowli roślin, kontrola tych procesów stanowi najważniejszy problemem dla wielu roślin uprawnych. Ma to szczególne znaczenie dla roślin rozetowych, takich jak sałata, szpinak i burak cukrowy, u których następuje szybkie wydłużanie łodygi (wystrzeliwanie) po przejściu do kwitnienia, co czyni uprawę bezużyteczną. Przejście od wzrostu wegetatywnego do reproduktywnego jest krytycznym etapem rozwojowym, a ponieważ jest to pierwszy etap rozmnażania płciowego, ma ogromne znaczenie w rolnictwie, ogrodnictwie i hodowli roślin. Rolnicy mogą chcieć przyspieszyć lub opóźnić czas kwitnienia lub całkowicie mu zapobiec: przykładowo w celu zapobiegania wystrzeliwaniu np. u sałaty lub buraka cukrowego. Lepsze zrozumienie biologii molekularnej kwitnienia

3 roślin umożliwi jego kontrolowanie lub wpływanie nań w różny sposób, dostarczając rolnictwu ważnych praktycznych korzyści. W publikacji PCT W096/14414, ujawniono zastosowanie genu Constans (CO) do modyfikowania mechanizmów kwitnienia. Niniejszy wynalazek proponuje sposób wpływania na przejście roślin od wzrostu wegetatywnego do reproduktywnego przez dostarczenie transformowanych roślin, w których przejście jest opóźnione lub przyspieszone przez ekspresję specyficznych transgenów wpływających na ten proces. Takie geny mogą być wyrażane konstytutywnie lub wyrażane jedynie w odpowiedzi na bodźce zewnętrzne, na przykład środowiskowe lub chemiczne. ATH1 jest genem Arabidopsis thaliana zawierającym homeobox. Gen ten jest opisany przez Quaedvlieg i wsp., w Plant Celi 7, , 1995 (włączone tu jako odniesienie): jego sekwencja jest podana w fig. 1 w tej publikacji. Został on wyizolowany z kolekcji czynników transkrypcyjnych indukowanych światłem. Podlega ekspresji w młodych siewkach i kwiatach. Poziomy mrna ATH1 w etiolowanych siewkach są bardzo zależne od światła (fitochrom) i wykazują również adaptację świetlną. Obecnie ustaliliśmy, że produkt białkowy ATH1 jest zaangażowany w zmianę rozwojową od wzrostu wegetatywnego do generatywnego. W rezultacie badań nad ATH1::GUS i wstępnych badań nad 35S::ATH1 wywnioskowaliśmy, że ATH1 pełni funkcję w przemianie wegetatywnego merystemu wierzchołkowego w merystem kwiatowy. Konkretnie, ATH1 działa jako antygiberelina, przez represję syntezy GA lub możliwe szlaku odpowiedzi na GA: ilustruje to Przykład 6. Nasze badania nad konstruktami ATH1::GUS wykazały, że w młodych, rosnących na świetle siewkach ATH1 jest wyrażany we wszystkich trzech warstwach merystemu wierzchołkowego pędu i zawiązkach liści. W młodych, ciągle rozwijających się liściach, ATH1 podlega ekspresji w tkance naczyniowej. Ekspresja ta zanika w liściach rozwiniętych. Ekspresja ATH1 w merystemie jest w znacznej mierze ograniczona do wegetatywnej fazy rozwoju. Jak tylko Arabidopsis zaczyna kwitnąć (zmiana rozwoju wegetatywnego na generatywny) i merystem wierzchołkowy pędu staje się merystemem kwiatowym, ekspresja ATH1 w merystemie obniża się. W czasie fazy kwitnienia ATH1 jest wytwarzany na niskim poziomie w rozwijającej się tkance naczyniowej łodygi. Później w rozwoju rośliny, kiedy pojawiają się kwiaty, ATH1 ulega ekspresji w różnych częściach młodej rośliny (dno kwiatowe, działka kielicha lub tkanka naczyniowa pręcika). Nasza hipoteza, że ATH1 jest zaangażowany w kontrolowanie przemiany faz od wzrostu wegetatywnego do generatywnego, jest dalej potwierdzona przez fenotypy czasu kwitnienia przy nadekspresji ATH1 w formie sensownej i antysensownej. Rośliny ektopikalnie wyrażające antysensowny ATH1 wykazują fenotyp wczesnego kwitnienia; przeciwnie, większość roślin przenoszących konstrukty do nadekspresji sensownego ATH1 są późno kwitnące. Mała część roślin przenoszących konstrukty do nadekspresji jest, na skutek obniżenia ATH1 przez kosupresję, wczesno kwitnąca, tak jak w przypadku nadekspresji antysensownego ATH1, a fenotyp tych roślin przypomina mutanta terminal flower (Shannon i Meeks- Wagner, 1991) i fenotypy przy nadekspresji LEAFY (Weigel i Nilsson, 1995), APETALA 1 (Mandel i Yanowski, 1995) i CONSTANS (Putteril i wsp., 1995). W oparciu o te wyniki, w połączeniu z danymi dla ATH1::GUS, wywnioskowaliśmy, że ATH1 jest zaangażowany w kontrolowanie fazy przejścia od wzrostu wegetatywnego do generatywnego u Arabidopsis thaliana i prawdopodobnie jest genem regulującym czas kwitnienia. W konsekwencji, przejście można promować przez hamowanie ekspresji genu ATH1, bądź tez opóźniać lub zapobiegać mu przez promowanie takiej ekspresji. Przedmiotem wynalazku jest sposób modyfikowania kwitnienia roślin, który obejmuje transformowanie roślin konstruktem zawierającym pełną albo częściową sekwencję DNA kodującą produkt ATH1 pod kontrolą promotora działającego w roślinach. Korzystnie proces kwitnienia w roślinach jest promowany przez transformowanie roślin konstruktem, który hamuje wytwarzanie białka ATH1, korzystniej konstrukt jest przystosowany do ekspresji antysensownego RNA do RNA wytwarzanego przez gen ATH1. W kolejnym korzystnym wykonaniu wynalazku proces kwitnienia w roślinach jest opóźniany przez transformowanie roślin konstruktem, który promuje wytwarzanie zrekombinowanego białka ATH1.

4 Stosowanie sekwencji do hamowania albo promowania ekspresji genu jest całkiem dobrze poznane. Pełna sekwencja genu, pod kontrolą promotora, który działa efektywnie w roślinie, będzie generalnie dawać nadekspresję produktu genu, prowadząc do wzmocnienia efektu działania wytwarzanego w ten sposób białka. Czasami poziom produktu genu jest obniżany: zjawisko to jest nazywane kosupresją. Obniżenie poziomu produktu genu uzyskuje się również generalnie przez zastosowanie mutacji dominujących negatywnych lub przez odwrócenie orientacji sekwencji genu w stosunku do promotora tak, że wytwarzany jest antysensowny informacyjny RNA. Konstrukt DNA stosowany w sposobie według wynalazku może być konstruktem antysensownym wytwarzającym RNA antysensowny lub konstruktem sensownym (kodującym, co najmniej część funkcjonalnego białka) wytwarzającym RNA sensowny. RNA antysensowny jest sekwencją RNA, która jest komplementarna do sekwencji zasad w odpowiadającym mu mrna: komplementarna w tym znaczeniu, że każda zasada (lub większość zasad) w sekwencji antysensownej (czytanej w kierunku 3' do 5) ma zdolność do tworzenia par z odpowiadającymi zasadami (G z C, A z U) w sekwencji mrna czytanej w kierunku od 3' do 5'. Taki antysensowny RNA może być wytwarzany w komórce przez transformowanie odpowiednim konstruktem genowym zorganizowanym tak, aby wytwarzać transkrypt, którego co najmniej część sekwencji jest komplementarna do co najmniej części nici kodującej odpowiedniego genu (lub sekwencji DNA wykazującej do niej zasadniczą homologię). RNA sensowny jest sekwencją RNA, która jest zasadniczo homologiczna do co najmniej części odpowiadającego jej konstruktu DNA w prawidłowej orientacji, tak aby wytwarzać transkrypt o sekwencji identycznej z co najmniej częścią nici kodującej odpowiedniego genu (lub sekwencji DNA wykazującej do niego zasadniczą homologię). Odpowiednie konstrukty sensowne można zastosować do hamowania ekspresji genu (jak opisano w Międzynarodowym zgłoszeniu patentowym W001/08299) lub konstrukt sensowny kodujący i wyrażający funkcjonalne białko można transformować do rośliny w celu nadprodukcji białka. Konstrukty DNA stosowane w sposobie według wynalazku mogą zawierać sekwencję zasad o długości co najmniej 10 zasad (korzystniej co najmniej 35 zasad) do transkrypcji do RNA. Teoretycznie nie istnieje żadna górna granica dla sekwencji zasad - może być ona tak długą, jak odpowiednie mrna wytwarzane przez komórkę, ale dla wygody na ogół będzie wystarczało zastosowanie sekwencji o długości pomiędzy 100 i 1000 zasad. Wytwarzanie takich konstruktów jest opisane bardziej szczegółowo poniżej. Jako źródło sekwencji zasad DNA do transkrypcji, można zastosować odpowiedni cdna lub DNA genomowy albo syntetyczny polinukleotyd. Izolowanie odpowiednich sekwencji ATH1 z Arabidopsis jest opisane w Quaedvlieg i wsp., powyżej; podobne metody można zastosować do izolowania sekwencji ATH1 z innych roślin. Mogą mieć one większy lub mniejszy stopień homologii z sekwencjami ATH1 z Arabidopsis. Można w ten sposób otrzymać sekwencje kodujące kompletne lub zasadniczo kompletne białko. Sekwencje DNA odpowiedniej długości można wyciąć w celu ich użycia za pośrednictwem enzymów restrykcyjnych. Kiedy stosuje się genomowy DNA jako źródło częściowej sekwencji zasad do transkrypcji, możliwe jest zastosowanie zarówno regionów intronowych, jak i eksonowych lub kombinację obydwu. W celu otrzymania konstruktu odpowiedniego do modyfikowania ekspresji ATH1 w komórkach roślinnych, można zastosować sekwencję cdna taką, jaką znajduje się w cdna dla białka lub sekwencję genu, jaka znajduje się w chromosomie rośliny. Zrekombinowane konstrukty DNA można wytworzyć przy zastosowaniu standardowych technik. Przykładowo, sekwencję DNA do transkrypcji można wytworzyć przez traktowanie wektora zawierającego taką sekwencję enzymem restrykcyjnym w celu wycięcia odpowiedniego fragmentu. Sekwencję DNA do transkrypcji można również wytworzyć przez przyłączenie i ligację syntetycznych oligonukleotydów lub przez reakcję łańcuchową polimerazy (PCR) w celu wytworzenia odpowiednich miejsc restrykcyjnych na każdym końcu. Sekwencję DNA klonuje się następnie do wektora za promotorem i przed terminatorem. Jeżeli wymagany jest DNA antysensowny, klonowanie przeprowadza się tak, aby wycięta sekwencja była odwrócona w stosunku do swojej orientacji w nici, z której została wycięta. W konstrukcie wyrażającym RNA antysensowny nić, która poprzednio była nicią matrycową staje się nicią kodującą i odwrotnie. Konstrukt będzie zatem kodował RNA o sekwencji

5 zasad, która jest komplementarna do części albo całej sekwencji mrna dla białka. A zatem dwie nici RNA są komplementarne nie tylko w swojej sekwencji zasad, ale również orientacji (5' do 3'). W konstrukcie wyrażającym RNA sensowny, nici, matrycowa i kodująca, zachowują przeznaczenie i orientację wyjściowego genu roślinnego. Konstrukty wyrażające RNA sensowny kodują RNA z sekwencją zasad, która jest homologiczna do części albo całej sekwencji mrna. W konstruktach, które wyrażają funkcjonalne białko, cały region kodujący genu jest połączony z sekwencjami kontrolującymi transkrypcję zdolnymi do ekspresji w roślinach. Przykładowo, konstrukty stosowane w sposobie według wynalazku mogą być wytworzone jak następuje. Odpowiedni wektor zawierający pożądaną sekwencję zasad do transkrypcji (taki jak klon cdna pathl) traktowany jest enzymem restrykcyjnym w celu wycięcia tej sekwencji. Otrzymaną w ten sposób nić DNA klonuje się (jeśli trzeba, w odwrotnej orientacji) w drugim wektorze zawierającym pożądaną sekwencję promotorową i pożądaną sekwencję terminatorową. Odpowiednie promotory obejmują promotor 35S wirusa mozaiki kalafiora i promotor genu poligalakturonazy pomidora (Bird i wsp., 1988, Plant Molecular Biology. 11: ) oraz inne promotory roślinne regulowane w rozwoju. Odpowiednie sekwencje terminatorowe obejmują taką sekwencję z genu syntazy nopalinowej Agrobacterium tumefaciens (koniec 3' nos). W konstrukcie DNA stosowanym w sposobie według wynalazku region inicjacji transkrypcji może pochodzić z dowolnego promotora działającego w roślinach. Region inicjacji transkrypcji może być usytuowany tak, aby uzyskać transkrypcję sekwencji DNA kodującej mrna, które jest komplementarny do odpowiednio dużego ciągu zasad w mrna kodującym białko ATH1 (czyniąc konstrukt DNA pełnym lub częściowym konstruktem antysensownym). Regionem inicjacji transkrypcji (lub promotorem) działającym w roślinach może być promotor konstytutywny (taki jak promotor 35S wirusa mozaiki kalafiora) lub promotor indukowany lub regulowany w rozwoju, zależnie od wymagań. Przykładowo, może być pożądane modyfikowanie aktywności białka na pewnych etapach rozwoju rośliny. Zastosowanie konstytutywnego promotora będzie wpływać na poziomy i funkcje białka we wszystkich częściach rośliny, podczas gdy zastosowanie promotora specyficznego dla tkanki umożliwi bardziej selektywną kontrolę ekspresji genu i związanych z tym funkcji. A zatem RNA antysensowny lub sensowny jest wytwarzany jedynie w narządzie, w którym jego działanie jest wymagane. Konstrukty DNA stosowane w sposobie według wynalazku można wstawiać do roślin w celu regulowania ekspresji genu ATH1 prowadząc do modyfikacji cech roślin (w szczególności kwitnienia). W zależności od natury konstruktu, wytwarzanie produktu genu ATH1 może być zwiększane lub zmniejszane, bądź przez cały czas, bądź w określonym stadium życia rośliny. Generalnie, jak można się spodziewać, wytwarzanie białka jest zwiększane jedynie przez konstrukty, które wyrażają RNA homologiczny do zasadniczo kompletnych endogennych mrna dla białka. Sensowne konstrukty pełnej długości mogą również hamować ekspresję białka. Konstrukty zawierające niekompletną sekwencję DNA, krótszą niż sekwencja odpowiadająca kompletnemu genowi, generalnie hamują ekspresję genu i wytwarzanie białek niezależnie od tego, czy są zorganizowane do ekspresji RNA sensownego czy antysensownego. Konstrukt DNA stosowany w sposobie według wynalazku transformuje się do docelowej komórki roślinnej. Docelowa komórka roślinna może być częścią całej rośliny lub może być izolowaną komórką lub częścią tkanki, którą można regenerować do całej rośliny. Docelową komórkę roślinną można wybrać z dowolnego gatunku roślin jednoliściennych lub dwuliściennych. Rośliny mogą pochodzić z transformowanej komórki roślinnej przez regenerację transformantów i przez wytwarzanie kolejnych pokoleń potomstwa transformantów. Konstrukty stosowane w sposobie według wynalazku można stosować do transformowania dowolnej rośliny przy zastosowaniu odpowiedniej techniki transformacji w celu wytworzenia roślin według wynalazku. Zarówno komórki roślin jednoliściennych, jak i dwuliściennych można transformować wieloma sposobami znanymi w tej dziedzinie. W wielu przypadkach takie komórki roślinne (szczególnie wtedy, kiedy są to komórki roślin dwuliściennych), można hodować w celu zregenerowania całej rośliny, która następnie rozmnaza się dając kolejne pokolenia zmodyfikowanych genetycznie roślin. Można zastosować dowolną

6 odpowiednią metodę transformacji roślin. Przykładowo, rośliny dwuliścienne, takie jak pomidor i melon można transformować przy zastosowaniu technologii plazmidu Ti Agrobacterium, takiej jak opisana przez Bevan (1984, Nucleic Acid Research, 12: ) lub Fillatti i wsp. (Biotechnology, Czerwiec 1987, 5: ). Tak stransformowane rośliny można rozmnażać płciowo albo przez hodowle komórkowe lub tkankowe. Rośliny jednoliścienne można transformować przy zastosowaniu działa genowego. Inne sposoby transformacji roślin obejmują mikro wstrzyknięcie i elektroporację. Przykłady genetycznie zmodyfikowanych roślin według wynalazku obejmują zboza, przykładowo ryz i kukurydzę, pszenicę, jęczmień, owies i zyto. Innymi ważnymi produktami nasiennymi są rzepak oleisty (kanola), burak cukrowy, słonecznik, soja i sorgo. Większość roślin uprawnych hoduje się corocznie z nasion i wytwarzanie nasion dowolnego rodzaju zalezy od zdolności rośliny do kwitnienia, do jej zapylenia i wytworzenia nasion. Kontrola czasu kwitnienia jest istotna w ogrodnictwie. Rośliny ogrodnicze, których kwitnienie może podlegać kontroli, obejmują sałatę, cykorię, warzywa z rodzaju Brassica, włączając w to kapustę, brokuły, kalafior; oraz goździki i pelargonie. Główną właściwością zmodyfikowanych roślin uzyskanych sposobem według wynalazku jest wczesne lub opóźnione kwitnienie. Genotypy, w których wytwarzanie białka ATH1 jest hamowane generalnie kwitną wcześnie; genotypy, w których jest on stymulowany kwitną późno. Można obserwować również inny wpływ na fenotyp, np. karłowacenie np. u tytoniu. Kontrolowanie czasu kwitnienia może być użyteczne z kilku powodów. Przykładowo, kwitnienie można kontrolować w celu dostarczenia kwiatów lub owoców w czasie najbardziej odpowiednim do sprzedaży. Przy wytwarzaniu hybryd, można koordynować kwitnienie męskich i żeńskich rodziców. Najdogodniej jest dokonać tego przez zastosowanie indukowalnych promotorów genów, odpowiadających na bodźce zewnętrzne, na przykład zastosowanie związków chemicznych. Przykładem takiego promotora jest promotor genu izoformy II S-transferazy glutationowej, aktywowany przez zastosowanie znanego czynnika zabezpieczającego przed herbicydem (WO 93/01294 dla ICI). Kontrola wystrzeliwania może być ekonomicznie ważna dla wielu gatunków roślin uprawnych. Przykładowo, w przypadku buraka cukrowego ogromne zastosowanie będzie miało wytwarzanie odmian, które mają zmniejszoną tendencję do wystrzeliwania po zadziałaniu zimnem. Przetwórnie będą mogły wydłużyć w czasie okres aktywności, ze znaczącymi oszczędnościami w ogólnych kosztach. Odmiany oporne na wystrzeliwanie mogłyby być siane bardzo wcześnie w sezonie (luty), a nawet poprzedniego roku jesienią (zakładając, ze zimowe przymrozki nie stanowią problemu). Ponadto, odmiany, w których wystrzeliwanie jest zwiększone mogą być szybciej hodowane: krzyżowanie można przeprowadzać corocznie zamiast raz na dwa lata jak dotychczas. Wczesnokwitnący słonecznik mógłby zwiększyć zasięg geograficzny. Można by go hodować bardziej na północ (na północ od Paryża) i możliwe w bardziej suchych regionach, np częściach Hiszpanii, unikając okresów suszy późnym latem. W przypadku warzyw, wystrzeliwanie można kontrolować na przykład dla sałaty i cykorii. Umożliwiłoby to łatwiejsze hodowanie rośliny uprawnej w czasie lata. Istniejące odmiany mają tendencję do gwałtownego wystrzeliwania w warunkach letnich. W przypadku traw zredukowanie (lub brak) wystrzeliwania jest korzystne dla typów paszowych»oprawia jakość karmy) i typów rekreacyjnych (lepsza jakość trawników). Czasami korzystne jest kontrolowanie ekspresji transgenu w czasie tak aby zatrzymać ją po rozpoczęciu się kwitnienia albo hamować naturalnie występujące geny aż do czasu rozpoczęcia kwitnienia. Można tego dokonać przez zastosowanie promotora ATH1 do kontrolowania ekspresji transgenu lub transkrypcji DNA homologicznego do naturalnego genu. A zatem dalszą odrębną częścią wynalazku jest dostarczenie konstruktu DNA zawierającego promotor ATH1 połączony z heterologicznym DNA w taki sposób, że powoduje jego transkrypcję w komórkach roślinnych; oraz komórki roślinne transformowane takimi konstruktami DNA. ATH1 podlega ekspresji w wegetatywnym merystemie wierzchołkowym i obniżenie tej ekspresji koreluje z przejściem do kwitnienia. Wymuszona konstytutywna ekspresja ATH1 prowadzi w rezultacie do dramatycznej represji przejścia do kwitnienia zarówno u Arabidopsis, jak i u tytoniu: a zatem w przypadku Arabidopsis wystrzeliwanie jest opóźnione.

7 Odwrotnie, represja ATH1 prowadzi w rezultacie do fenotypu wczesnego kwitnienia. Nasze wyniki sugerują, ze ATH1 rozciąga swoją funkcję na modulowanie biosyntezy lub odpowiedzi na GA. Spodziewamy się, ze gen ATH1 będzie stanowił podstawę szczególnie użytecznego systemu kontrolowania wystrzeliwania. Długość dnia i przejście do kwitnienia Przejście do kwitnienia zostało szczególnie dobrze zbadane u Arabidopsis thaliana. Ten gatunek stał się modelowym systemem do badania przejścia do kwitnienia: na poziomie genetycznym przez izolację mutantów czasu kwitnienia, na poziomie molekularnym przez klonowanie genów, których produkty biorą udział w kontrolowaniu przejścia do kwitnienia. Arabidopsis jest typową rośliną rozetową, w której liście wegetatywne są ściśle upakowane na skutek zredukowanego wydłużania się międzywęźli. Po przejściu do kwitnienia nowo utworzone międzywęźla gwałtownie się wydłużają ( wystrzeliwanie ). W większości ekotypów Arabidopsis długość dnia ma największe znaczenie przy wyznaczaniu przejścia do kwitnienia. Arabidopsis jest fakultatywną rośliną długiego dnia (LD, ang. Long Day), co oznacza, że przejście do kwitnienia jest przyspieszane przez długie dni (cykl 16 godzin światła/8 godzin ciemności), ale nie stanowi to bezwzględnego wymagania. W warunkach długiego dnia (LD) przejście do kwitnienia jest zapoczątkowywane szybko i tworzy się jedynie kilka rozetowych liści (~7 liści dnia dla ekotypu Col-0). W czasie hodowli w warunkach krótkiego dnia (SD, ang. Short Day), np. 8 godz. światła/l 6 godz. ciemności, przejście do kwitnienia trwa duzo dłużej (~60 dni) i tworzą się pełne rozetki liściowe, które mają ponad ~30 liści w rozetce (ekotyp Col-0). Kwas giberelinowy (GA) i przejście do kwitnienia Wiadomo od dawna, że działanie giberelinami promuje przejście do kwitnienia u wielu gatunków roślin. Większość gatunków, u których zastosowanie GA może indukować kwitnienie, to rośliny długiego dnia lub wymagające zimna i wiele z nich normalnie rośnie w formie rozet w warunkach braku indukcji. Ponadto, kilka doświadczeń sugeruje, że poziom endogennych GA ma udział w kontrolowaniu przejścia do kwitnienia: warunki, które indukują przejście do kwitnienia mogą wywierać wpływ przez podniesienie poziomu endogennych GA prawdopodobnie w obrębie albo w pobliżu merystemu wierzchołkowego. Mutanty Arabidopsis defektywne w biosyntezie GA (seria GA) lub niewrażliwe na ten hormon (seria GAI) wykazują fenotyp późnego kwitnienia w warunkach braku indukcji, a ponadto poważnie zaburzony mutant GAI-3 jest również późno kwitnący w warunkach indukcji (Wilson i wsp., 1992). Udział GA w kontrolowaniu przejścia do kwitnienia przez ATH1 Testowaliśmy, czy egzogenny GA może przezwyciężyć hamujący efekt konstytutywnej ekspresji ATH1 u tytoniu. Najwyraźniej, opryskiwanie GA miało zdolność do ratowania fenotypu wczesnego kwitnienia u roślin tytoniu konstytutywnie wyrażających ATH1. Udział GA wykazano również przez fenotyp obniżonego wydłużania międzywęźli roślin tytoniu konstytutywnie wyrażających ATH1. Odkrycia te sugerują, że ATH1 działa jako represor biosyntezy GA, lub, alternatywnie, odpowiedzi na GA. Dominujący wpływ nadekspresji ATH1 na przejście do kwitnienia w kombinacji z odwracaniem tego efektu przez endogennie dodawane GA sugerują wiele zastosowań u różnych roślin uprawnych. Jest to szczególnie interesujące, ponieważ deregulacja ekspresji ATH1 nie prowadzi do plejotropowych fenotypów, a odwrócenie fenotypu jest całkowite. Całkowite odratowanie oznacza, że nie będzie problemu dotyczącego rozmnażania albo powielania transformantów ATH1, a zatem utrzymywanie linii transgenicznych, co może być poważnym problemem dla mutantów kwitnienia, jest proste. Stosując przemianę GA, rośliny można przywracać do rozwoju typu dzikiego w dowolnym momencie, rośliny kwitną normalnie i normalnie powstają nasiona. Możliwe jest również hamowanie nadekspresji ATH1 w roślinach zmodyfikowanych genetycznie uzyskanych sposobem według wynalazku przez traktowanie roślin gibereliną, na przykład gibereliną A3 lub A4/A7. Wzrost indeksu zbiorów Kiedy rośliny rosną blisko siebie, występuje syndrom unikania cienia, w którym rośliny reagują na promieniowanie dalekiej czerwieni odbijane od sąsiadów. Najbardziej widocznym

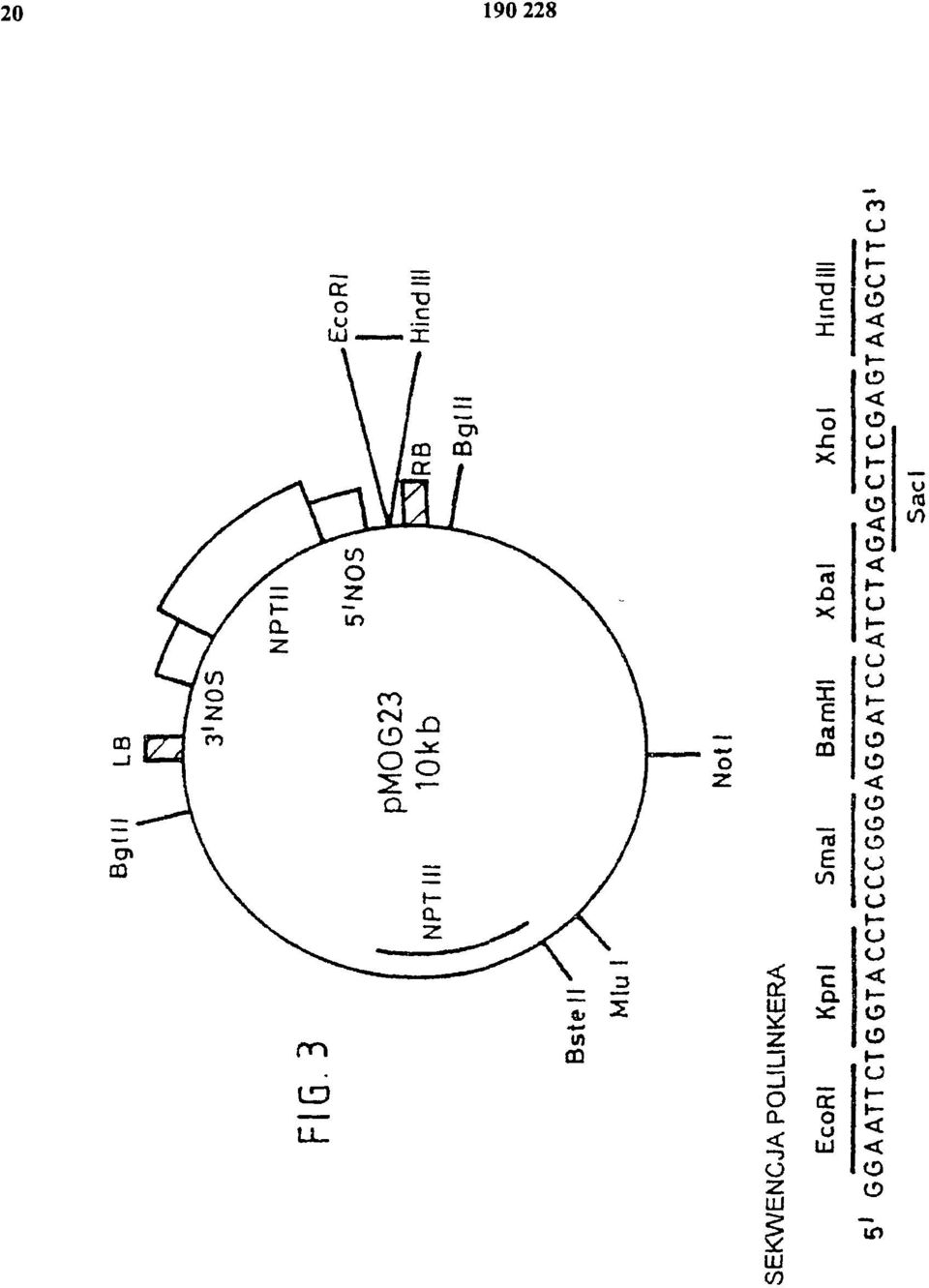
8 tego wynikiem jest szybki i dramatyczny wzrost tempa wzrostu łodyg i ogonków liściowych przy wzroście liści, wytwarzanie narządów spichrzowych i rozwój reproduktywny. Wiadomo, ze przez nadekspresję genów phya w tytoniu można przezwyciężyć odpowiedź unikania cienia, prowadząc do zwiększonego indeksu zbiorów (Robson i wsp., 1996). Indeks zbiorów wyraża się jako biomasa liści w proporcji do całej biomasy. Nadekspresja ATH1 w tytoniu powoduje obniżenie wzrostu łodyg, natomiast wzrost i liczba liści pozostają niezmienione, a nawet zmniejszone w porównaniu do typu dzikiego. Fenotypy nadekspresji ATH1 i nadekspresji phya są podobne, co sugeruje możliwość zastosowania nadekspresji ATH1 do zwiększenia indeksu zbiorów dla roślin uprawnych. Wynalazek będzie dalej opisany w odniesieniu do następujących przykładów i doświadczeń, które ilustrują niektóre aspekty niniejszego wynalazku; oraz w odniesieniu do rysunków, gdzie: Figura 1 przedstawia sekwencję DNA dla cdna ATH1; Figura 2 jest diagramem plazmidu pwp90; Figura 3 jest diagramem plazmidu pmog23; Figura 4 jest diagramem plazmidu pvdh275; Figura 5 jest wykresem słupkowym przedstawiającym karłowacenie powodowane przez konstytutywną ekspresję ATH1 w roślinach tytoniu w wieku 90 dni; Figura 6 jest wykresem pokazującym wpływ działania giberelin na wysokość roślin tytoniu z nadekspresją ATH1; Figura 7 jest wykresem słupkowym przedstawiającym czas kwitnienia (jako liczbę tworzonych liści rozetowych) dla roślin ze zmniejszoną i zwiększoną ekspresją ATH1 w porównaniu z Arabidopsis typu dzikiego (ekotyp C24). Metody ogólne Materiał roślinny i warunki wzrostu Używanymi genotypami Arabidopsis thaliana typu dzikiego były Columbia i C24. Gen ATH1 jest położony na chromosomie 4 pomiędzy markerami mi431 (96,9 cm) i (97,9 cm). Arabidopsis thaliana Columbia stosowano w doświadczeniach z transformacją roślin przy zastosowaniu protokołu infiltracji pod próżnią, natomiast Arabidopsis thaliana C24 stosowano w doświadczeniach z transformacją roślin przy zastosowaniu protokołu dla transformacji korzeni. Rośliny hodowano w pomieszczeniu hodowlanym w świetle fluorescencyjnym z 16 godzinnym okresem świetlnym, po którym następował 8 godzinny okres ciemności w stałej temperaturze 22 C. W celu pomiaru czasu kwitnienia, nasiona namaczano i umieszczano w 4 C na 4 dni w celu przełamania okresu spoczynkowego i siano do gleby. Kiełkujące siewki zwykle przykrywano pokrywą przez pierwsze 1-2 tygodnie w celu zapobiegania wysychaniu. Transformacja roślin Arabidopsis Binarne konstrukty zawierające chimerowe geny ATH1-GUS i geny 35S-antysensowny ATH1 transformowano do Arabidopsis thaliana ekotyp C24 przy zastosowaniu metody transformacji korzeni za pośrednictwem Agrobacterium tumefaciens według Valvekens i wsp. (1988). Transformanty selekcjonowano na pożywce zawierającej 50 mg/l kanamycyny. Binarne konstrukty zawierające chimerowe geny 35S-ATH1 transformowano do Arabidopsis thaliana ekotyp Columbia przy zastosowaniu protokołu infiltracji pod próżnią (Bent i wsp. (1994); Betchold i wsp. (1993)) z pewnymi modyfikacjami. Rośliny hodowano oddzielnie w 5,5 cm naczyniach. Rośliny transformowano po pojawieniu się pierwszych strąków na drugorzędowych pędach kwiatowych. 900 ml hodowli Agrobacterium tumefaciens zawierających odpowiedni konstrukt hodowano przez noc przed dniem infiltracji, komórki zbierano przez wirowanie i zawieszano w równiej objętości pożywki do infiltracji, zawierającej 2% zamiast 5% sacharozy. Rośliny infiltrowano przez zanurzenie całych rozetek i pędów kwiatowych na 10 minut pod ciśnieniem 100 mm Hg. Nasiona transformantów selekcjonowano na pożywce zawierającej 50 mg/l kanamycyny.

9 Przykład 1 Analiza ekspresji ATH1 Izolacja całkowitego RNA Całkowity RNA z roślin izolowano według De Vries i wsp. (1988) z pewnymi niewielkimi modyfikacjami: (1) tkankę roślinną mielono w ciekłym azocie w obecności połowy objętości mieszaniny fenol/bufor do ekstrakcji i ogrzewano w 65 C w łaźni wodnej i (2) RNA wytrącano etanolem/octanem sodu przed i po wytrąceniu LiCl. Analiza ochrony przed RNAazą Fragment HindIII-XhoI z fagmidu ATH1 sklonowano w pblu-scriptsk(-) (Stratagene) i strawiono HinI II w ceiu wytworzenia matrycy dla polimerazy RNA T7. Sonda RNA ATH1 chroniła fragment 140 nukleotydów. Sondę RNA zsyntetyzowano przy zastosowaniu polimerazy RNAT7 (Pharmacia) i buforu opisanego przez producenta, z tą różnicą, że użyto 160 μci [32P] UTP (800 Ci/mmmol). Ochronę przed RNAzą wykonano przy zastosowaniu 10 μg całkowitego RNA i 10 μg trna według protokołu opisanego przez Sambrook i wsp. (1989). Strawiona mieszanina zawierała 600 jedn./ml RNazy Tl (Gibco BRL) i 20 μg/ml RNazy A (Boehringer). Hybrydy RNA:RNA analizowano przez elektroforezę w żelach do sekwencjonowania (6% poliakryloamid/7 M mocznik) i uwidaczniano przez autoradiografię. Wytwarzanie chimerowych konstruktów ATH1-GUS Wyizolowano fragment Spel-Ncol zawierający około 1300 nukleotydów z sekwencji promotora ATH1. Po wypełnieniu końców w miejscu Ncol przez polimerazę Klenowa fragment ten wstawiono do pojedynczych miejsc Smal/Xbal binarnego wektora pbi101.1, który zawierał gen GUS (Jefferson i wsp., 1987), tworząc fuzję translacyjną pomiędzy promotorem ATH1 i genem GUS. Białko kodowane przez ten chimerowy gen składa się z 42 aminokwasów ATH1 połączonych z białkiem GUS. Binarny konstrukt został nazwany th1.4. th1.4 transformowano do kompetentnych komórek Agrobacterium tumefaciens LBA4404 (Gelvin i Schilperoort, 1988). Linie Arabidopsis (ekotyp C24) transformowano jak opisano poniżej. Lokalizacja in situ aktywności GUS w liniach Arabidopsis thaliana transgenicznych pod względem ATH1 -GUS Siewki i tkanki roślinne zbierano i barwiono przez 1do 16 godzin w 37 C w roztworze zawierającym 0,5 mg/ml X-Gluc (Biosynth AG) rozpuszczonego w N-dimetyloformamidzie, 0,1% Triton X-100, 0,5 mm K4Fe(CN)6 H2O, 0,5 mm K.3Fe(CN)6 i 50 mm bufor fosforanowy, ph 7,2. Po barwieniu X-Gluc, tkanki roślinne utrwalano przez noc w roztworze zawierającym 1% glutaraldehyd i 4% formaldehyd w 50 mm fosforanie sodu ph 7,2. Następnie siewki odwadniano w kolejnych etapach: w 10%, 30%, 50%, 70%, 90% i 2 x 100% etanolu. Tkanki dużych roślin zatapiano wstępnie w 1% agarozie (Sigma). Infiltrację i zatapianie w Techno vite 7100 (Kulzer, Hereaus) wykonywano zgodnie z instrukcją producenta. Skrawki 4 pm sporządzano przy zastosowaniu urządzenia obrotowego Reichert-Jung 1140 z jednorazowym stalowym nożem Adams. Skrawki barwiono 0,1% czerwienią rutenową (Sigma) w wodzie destylowanej przez 2 minuty w temperaturze pokojowej i fotografowano w Zeiss Axioskop przy zastosowaniu błony Kodak Professional Ektar 25. Siewki utrwalano i odwadniano jak wyżej. Technovit 7100 infiltrowano przez 1 dzień. Siewki przenoszono następnie do konstrukcji z folii celuloidowej (Amvois), taśmy dwustronnie klejącej, folii, dwustronnej taśmy. W ostatnich trzech warstwach wycinano centralny region w ceiu umieszczenia w nim siewek. Następnie nanoszono siewki w roztworze Technovitu 7100 i centralny region nakrywano inną folią. Po spolimeryzowaniu przez noc w temperaturze pokojowej otrzymano plastykową płytkę zawierającą siewki. W celu uzyskania skrawków zatopionych w płytce siewek, usuwano materiał folii celuloidowych i płytkę cięto wzdłuż przekroju podłużnego odpowiednich regionów siewek. Sporządzanie skrawków, barwienie i fotografowanie wykonywano jak opisano powyżej. Lokalizacja ekspresji ATH1 Ekspresję genu ATH1 analizowano przy zastosowaniu analizy ochrony przed RNAzą (Quaevlieg i wsp., 1995). Wysokie poziomy mrna ATH1 wykrywano w czasie wczesnego rozwoju siewek (dni 2-6) oraz w kwiatach dojrzałych roślin Arabidopsis. Lokalizację komór-
 (Stratagene) i strawiono HinI II w ceiu wytworzenia matrycy dla polimerazy RNA T7.")
10 kową ekspresji genu ATH1 określano przez wprowadzenie chimerowego konstruktu ATH1-GUS th1.4 do Arabidopsis thaliana. Różne tkanki barwiono X-gluc oraz sporządzono preparaty całej rośliny i skrawki tkanek dla uwidocznienia aktywności GUS (patrz poniżej). Ekspresja ATH1 w czasie rozwoju wegetatywnego Wierzchołek wzrostu 5-dniowej siewki hodowanej na świetle jest płaski i składa się z dwuwarstwowej tuniki otaczającej leżący pod nią korpus. W tym stadium, merystem ma zapoczątkowane zawiązki pierwszej pary liściowej (Mischke i Brown, 1965). W roślinach transformowanych th1.4, wysokie poziomy aktywności GUS były obecne w wierzchołkach pędów. Przekroje wierzchołka pędu pokazały, że wysoka aktywność GUS jest widoczna we wszystkich trzech warstwach merystemu wierzchołkowego pędu i rozciąga się przez region podwierzchołkowy, sięgając dalej, tam gdzie wiązka naczyniowa hipokotyla rozgałęzia się do liścieni. Wysokie aktywności GUS były również obecne w zawiązkach pierwszej pary liści. Ekspresja ATH1 w czasie przejścia do kwitnienia i rozwoju kwiatostanu. Wyjściowo, w czasie fazy kwitnienia, merystem wierzchołkowy pędu rozwija się w łodygę, liście łodygowe i wtórne kwiatostany. W miarę rozwoju kwiatostanu, merystem kwiatowy wytwarza zawiązki kwiatów. W roślinach transformowanych th1.4, aktywność GUS obniżała się w merystemie kwiatowym w czasie fazy przejścia. W merystemie nie było wykrywalnej aktywności GUS. Niskie poziomy aktywności GUS były obecne w strefie żeberka. Później, kiedy pojawiały się kwiaty, aktywność GUS była obecna w różnych częściach młodych kwiatów (dno kwiatowe, działka kielicha lub tkanka naczyniowa pręcika). Przykład 2 Konstrukcja fuzji promotorów z otwartą ramką odczytu ATH1 cdna ATH1 sklonowano do unikalnych miejsc restrykcyjnych EcoRI/XhoI do dobrze znanego i komercyjnie dostępnego wektora pbluescript SK(-) (Stratagene) Fuzja promotora 35S z CaMV z otwartą ramką odczytu ATH1 Fragment BamHI/SnaBI zawierający 1573 nukleotydy sekwencji cdna ACTH1 (miejsce BamHI zostało utworzone przez mutagenezę PCR, 35 nukleotydów za miejscem startu translacji) wyizolowano i wstawiono do pojedynczych miejsc do klonowania BamHI/SmaI w wektorze pwp90, który zawiera podwójny promotor 35S CaMV i terminator NOS (patrz fig. 2), uzyskując w rezultacie fuzję transkrypcyjną pomiędzy podwójnym promotorem 35S CaMV i cdna ATH1. Konstrukt ten, nazwany ch1.24, przecięto następnie enzymami restrykcyjnymi SstI/EcoRV, po czym uzyskaną w rezultacie wstawkę SstI/EcoRV wstawiono do pojedynczych miejsc restrykcyjnych BamHI/Smal w wektorze binarnym pmog23 (patrz fig. 3). Binarny konstrukt nazwano th1.2. th1.2 transformowano do kompetentnych komórek Agrobacterium tumefaciens pgv2260 (Caplan i wsp., 1985). Linie Arabidopsis (ekotyp Col-0) transformowano przy zastosowaniu infiltracji pod próżnią tak, jak opisano poniżej. 2.2 Konstuowanie fuzji promotora 35S z CaMV z antysensowną ramką ATH1 Fragment EcoRI/SnaBI zawierający około 1830 nukleotydów sekwencji cdna ACTH1 wyizolowano i wstawiono do pojedynczych miejsc do klonowania Smal/EcoRI w wektorze pwp90 (patrz fig. 2), uzyskując w rezultacie fuzję transkrypcyjną pomiędzy podwójnym promotorem 35S CaMY i antysensowną ramką ATH1. Uzyskany konstrukt nazwano ch1.22. Wstawkę EcoRV/SstI z ch1.22 sklonowano następnie w pojedynczych miejscach restrykcyjnych Smal/SacI binarnego wektora pmog23 (MOGEN)(patrz Fig. 3). Ten binarny konstrukt, nazwany th 1.1, transformowano do kompetentnych komórek Agrobacterium tumefaciens LBA4404. Linie Arabidopsis (ekotyp C24) transformowano tak, jak opisano poniżej Fuzja promotora szoku cieplnego z otwartą ramką odczytu ATH1 Przez mutagenezę PCR, utworzono dodatkowe miejsce BamHI w ptt19, wektorze zawierającym promotor, lider i 77 nukleotydów sekwencji kodującej genu szoku cieplnego Hsp 18.2 z Arabidopsis thaliana (Takahashi i Komeda, 1989). Dodatkowe miejsce BamHI jest zlokalizowane w niepodlegającym translacji liderze w pozycji nukleotydowej -710 od startu translacji Hsp18.2. Przez trawienie restrykcyjne BamHI usunięto niepodlegający translacji lider 5' i 77 nukleotydów sekwencji kodującej Hsp18.2. Pozostały konstrukt nazwano bezliderowym ptt19.
. W roślinach transformowanych th1.")
11 Fragment HindIII/BamHI tego bezliderowego ptt19, zawierający jedynie sekwencję promotorowąhsp18.2, połączono z fragmentem BamHI/EcoRI zawierającym całą sekwencję cdna ATH1, co dało w rezultacie fuzję transkrypcyjną promotora Hsp.18.2 z niepodlegającym translacji liderem i sekwencją kodującą. Miejsca BamHI i EcoRI utworzono przez mutagenezę PCR, w wyniku czego powstało miejsce restrykcyjne BamHI na początku sekwencji cd- NA ATH1 i miejsce restrykcyjne EcoRI zaraz za kodonem stop TAA. Otrzymany w rezultacie fragment HindIII/EcoRI wstawiono do pojedynczych miejscach restrykcyjnych HindIII/EcoRI wektora pwp90 (patrz fig. 2) i ten nowy konstrukt strawiono następnie częściowo enzymami restrykcyjnymi HindIII i EcoRV. Największy fragment HindIII/EcoRV wstawiono następnie do binarnego wektora pbin 19 przeciętego HindIII/SmaI (Frisch i wsp., 1995). Konstrukt ten nazwano HspH1. Wykonano również fuzję transkrypcyjną pomiędzy promotorem Hsp18.2 i sekwencją kodującą ATH1 bez sekwencji liderowej. Dodatkowe miejsce BamHI w cdna ATH1 utworzono przez mutagenezę PCR bezpośrednio przed startem translacji. Trawienie tego miejsca BamHI w połączeniu z trawieniem pojedynczego miejsca Xhol w cdnaathl daje w rezultacie fragment o długości około 680 nukleotydów, zawierający sekwencję kodującą ATHI. Ten fragment złożony z 680 nukleotydów został zamieniony na fragment o długości około 980 nukleotydów, który tworzy się po trawieniu HspHl enzymami restrykcyjnymi BamHI/XhoI. Rezultatem tego jest HspHIB, fuzja transkrypcyjna pomiędzy bezliderową sekwencją kodującą ATH1 i promotorem Hspl8.2. Zarówno HspH1, jak i HspH1B transformowano do kompetentnych komórek Agrobacterium tumefaciens LBA4404. Linie Arabidopsis (ekotyp C24) transformowano tak, jak opisano poniżej. 2.4 Fuzja promotora plastocyjaniny grochu z otwartą ramką odczytu ATH1 Fuzję transkrypcyjną pomiędzy promotorem plastocyjaniny grochu i sekwencją kodującą ATH1 można utworzyć przez wstawienie sekwencji kodującej ATHI do pojedynczych miejsc restrykcyjnych BamHI i SaII pvdh275 (Pwee i Gray, 1993; Last i Gray, 1989) (patrz również fig. 4). W sekwencji kodującej ATH1 dodatkowe miejsce Sali (bezpośrednio przed startem ATG w ATH1) i BamHI (bezpośrednio po sygnale stop TAA w ATH1) można utworzyć przez metagenezę PCR. Otrzymany w rezultacie konstrukt, w którym sekwencja kodująca ATH1 jest wstawiona pomiędzy promotor plastocyjaniny grochu i terminator nos można transformować do komórek Agrobacterium tumefaciens, a następnie dokonać transformacji roślin. Wprowadzenie dodatkowych kopii ATH1 do Arabidopsis Dodatkowe kopie ATH1 można wprowadzić do roślin Arabidopsis przez transformowanie ich dodatkowymi loci ATH1 zawierającymi promotor ATH1 i sekwencję kodującą ATH1. Można tego dokonać przez fuzję fragmentu SnaBI/Ncol cdna ATH1 o wielkości około 1000 nukleotydów z fragmentem restrykcyjnym Sstl/EcoRI cdna ATHl o wielkości około 250 nukleotydów z pbi101.1, zawierającym terminator nos Agrobacterium (Jefferson i wsp., 1987). Otrzymany w rezultacie fragment można połączyć z fragmentem restrykcyjnym Ncol o wielkości około 3,5 kb z genomowego klonu ATH1 (Quaedvlieg i wsp., 1995). Utworzony w ten sposób fragment NcoI/EcoRI o wielkości około 4750 nukleotydów zawierający promotor ATHI, sekwencję kodującą ATH1 i terminator nos, można wstawić do wektora do klonowania pmtl23 przeciętego Ncol/EcoRI (Chambers i wsp., 1988). Fragment restrykcyjny StuI/EcoRI otrzymanego w rezultacie konstruktu można następnie wstawić do binarnego wektora pmog23, transformować do komórek Agrobacterium, a następnie dokonać transformacji roślin. Przykład 3 Wpływ na cechy kwitnienia przez zastosowanie fuzji genowej promotor 35S CaMV/ATH1. Pomiar czasu kwitnienia Czas kwitnienia mierzono przez liczenie liczby liści, włączając w to liścienie, w rozecie w czasie pojawiania się pączka kwiatowego. Uprzednio wykazano ścisłą korelację pomiędzy liczbą liści i czasem kwitnienia (Koorneef i wsp., 1991; Bagnall (1993)). Nadekspresja ATH1 prowadzi do opóźnienia kwitnienia

12 W celu uzyskania dalszej wiedzy o roli ATH1 w rozwoju rośliny, sekwencję cdna ATH1 pełnej długości przyłączono do konstytutywnego promotora 35S wirusa mozaiki tytoniu i utworzony w ten sposób chimerowy gen 35S:ATH1 transformowano do Arabidopsis ekotyp Col-0 stosując metodę infiltracji pod próżnią. Otrzymano w ten sposób sześć niezależnych pierwotnych transformantów. Wszystkie te transgeniczne linie krzyżowano wsobnie. Z każdej niezależnej linii transgenicznej 40 odrębnych nasion poddano kiełkowaniu w glebie i sprawdzano pod kątem zmienionych fenotypów w porównaniu z roślinami typu dzikiego. Cztery spośród sześciu linii wykazywało fenotyp zmieniony pod względem czasu kwitnienia. W trzech z tych linii wszystkie rośliny miały opóźnione kwitnienie (około 14 liści rozetowych do czasu kwitnienia w porównaniu do 10 liści rozetowych u roślin typu dzikiego Col-0). W pozostałych liniach około 85% roślin wykazywało ten sam fenotyp opóźnionego kwitnienia, podczas gdy 15% roślin wykazywało fenotyp wczesnego kwitnienia (po około 7 liściach rozetowych), jak wykazano, na skutek braku RNA ATH1. Te wcześnie kwitnące rośliny wykazywały również fenotyp terminal flower, często z niekompletnymi kwiatami i zmutowanymi narządami kwiatowymi. Przykład 4 Wczesne kwitnienie przez ekspresją antysensownego ATH1. Podobnie jak ektopowa nadekspresja ATH1, hamowanie funkcji genu ATH1 może być również zastosowane do wpływania na czas kwitnienia. Hamowanie funkcji genu odbywa się przez konstytutywną nadekspresję antysensownego ATH1. Antysensowny pełnej długości cdna ATH1 połączono z konstytutywnym promotorem 35S wirusa mozaiki kalafiora i utworzony w ten sposób chimerowy gen 35S:antysensowny ATH1 transformowano do Arabidopsis ekotyp C24 stosując protokół transformacji korzeniowej według Valvekens. Otrzymano dwadzieścia dwa niezależne transformaty i wszystkie krzyżowano wsobnie. Z każdej linii 10 odrębnych nasion poddano kiełkowaniu w glebie i sprawdzano pod kątem zmienionych fenotypów w porównaniu z roślinami typu dzikiego C24. W pięciu z tych linii rośliny wykazywały fenotyp wczesnego kwitnienia: kwitnienie rozpoczynało się po utworzeniu od sześciu do dziesięciu liści rozetowych w porównaniu do około dwudziestu liści u roślin typu dzikiego. Przykład 5 Zmiana czasu kwitnienia u Nicotiana tabacum przez nadekspresję ATH1 Tak jak u Arabidopsis, ektopowa nadekspresja cdna ATH1 (kierowana przez promotor 35S wirusa mozaiki kalafiora) w tytoniu (Nicotiana tabacum cv. Samsun) prowadzi również do opóźnienia w czasie kwitnienia w porównaniu z roślinami tytoniu typy dzikiego. W roślinach tytoniu 35S::ATH1 czas kwitnienia był opóźniony o tygodnie lub miesiące. Rośliny te były również karłowate. Ta cecha karłowacenia, podobnie jak fenotyp kwitnienia, jest wyraźnie skorelowany z poziomem ekspresji transgenu. W najcięższych przypadkach, rośliny nie kwitły wcale i osiągały jedynie jedną piątą normalnej wysokości, podczas gdy w mniej poważnych przypadkach rośliny miały opóźnione kwitnienie o jedynie tydzień lub dwa i osiągały około czterech piątych normalnej wysokości. Liczba i kształt liści były normalne we wszystkich transformowanych roślinach. Przykłady 6-8 Następujące przykłady ilustrują wpływ GA na transgeniczne rośliny według wynalazku. Jak zaznaczono powyżej, nadekspresja ATH1 efektywnie hamuje wystrzeliwanie (indukcję kwiatów). Postawiliśmy hipotezę, że ATH1 może być represorem syntezy GA lub szlaku odpowiedzi na GA (sądzimy, ze pierwszej). Następujące przykłady przedstawiają i przemawiają za tą hipotezą. Metody ogólne Rośliny tytoniu (Nicotiana tabacum L. cv. Samsun NN) transformowano przy zastosowaniu procedury dysków liściowych (Horsch i wsp., 1985). Transgeniczne rośliny selekcjonowano na pożywce MS (Murashige i Skoog, 1962) zawierającej 300 mg/ml kanamycyny i 2% sacharozę. Po przeniesieniu do gleby, rośliny hodowano w szklarni w 22 C w warunkach świetlnych 16 godzin światła dziennego, kiedy trzeba uzupełnianego światłem sztucznym. Wpływ giberelin (GA) testowano przez stosowanie na liście (opryskiwanie) 100 mm GA3

13 w roztworze zawierającym 100 ml/l Tritonu X-100. Rośliny kontrolne opryskiwano roztworem zawierającym jedynie Triton X-100. Opryskiwanie rozpoczynano 60 dni po zasiewie, kiedy rośliny typu dzikiego miały wysokość około 2,5 cm i kontynuowano w odstępach 3 do 4 dni. Wysokość roślin mierzono, co 3 do 4 dni i kontynuowano do czasu wystąpienia kwitnienia, co określano na podstawie pojawienia się zawiązków kwiatowych. Przykład 6 Konstytutywna nadekspresja sensownego ATH1 prowadzi do opóźnionego kwitnienia 6.1 Nadekspresja ATH1 w tytoniu W celu uzyskania konstytutywnej nadekspresji genu ATH1 w transgenicznych roślinach, jego region kodujący umieszczono pod kontrolą promotora 35S CaMV i otrzymany w rezultacie konstrukt transformowano do tytoniu (Nicotiana tabacum cv. Samsun NN). Otrzymano czterdzieści niezależnych roślin opornych na kanamycynę, z których jedynie pięć wykazywało wykrywalną ekspresję transgenu. Poziomy mrna ATH1 były zróżnicowane od wysokiego u roślin H10E#4, #10, #30 do średniego/niskiego poziomu u roślin H10E#35 i #37. W zależności od poziomu ekspresji ATH1, kwitnienie u tych roślin było opóźnione od tygodni do miesięcy w porównaniu z roślinami typu dzikiego, które kwitną po 3-5 miesiącach w zależności od sezonu. W najpoważniejszym przypadku (H10E#4) rośliny nigdy nie kwitły do czasu zestarzenia się (>15 miesięcy po wysianiu). Rośliny H10E#10 i #35, które wykazywały wysoką ekspresję ATH1, kwitły po 15 miesiącach, podczas gdy rośliny wykazujące średnią/niską ekspresję, H10E#35 i #37 nie kwitły do 6 miesięcy. Podobnie jak zmieniony fenotyp kwitnienia, rośliny z nadekspresją ATH1 wykazywały zmniejszony wzrost łodygi, prowadzący w rezultacie do karłowacenia roślin. Występuje tu wyraźna korelacja pomiędzy stopniem karłowatości wzrostu i poziomem ekspresji transgenu (patrz fig. 5). W najpoważniejszym przypadku rośliny osiągały jedynie jedną piątą swojej normalnej wysokości. Liczba liści wykazywała zróżnicowanie od dwukrotnie wyższej od typu dzikiego do normalnej we wszystkich transgenach. 6.2 Nadekspresję ATH1 można odwrócić przez GA3 Fenotyp nadekspresji ATH1 można odwrócić do fenotypu typu dzikiego przez zastosowanie GA3. Stosowanie GA3 na liście wobec roślin tytoniu z Przykładu 6.1 (opryskiwanie 100 mm GA3 z przerwami trzy-czterodniowymi) prowadzi w rezultacie do całkowitego przywrócenia długości łodyg typu dzikiego (fig. 6). Dotyczyło to również fenotypu późnego kwitnienia (dane nie pokazane). Przykład 7 Nadekspresja ATH1 u Arabidopsis W celu uzyskania dalszych danych odnośnie roli ATH1 w rozwoju rośliny, pełnej długości cdna ATH1 połączono z konstytutywnym promotorem 35S wirusa mozaiki kalafiora i chimerowy gen 35S:ATH1 transformowano do Arabidopsis ekotyp Col-0 stosując metodę infiltracji pod próżnią. Otrzymano w ten sposób sześć niezależnych pierwotnych transformantow i wszystkie transgeniczne linie krzyzowano wsobnie. Z każdej niezależnej linii transgenicznej 40 odrębnych nasion poddano kiełkowaniu w glebie i sprawdzano pod kątem zmienionych fenotypów w porównaniu z roślinami typu dzikiego. Cztery spośród sześciu linii wykazywało fenotyp zmieniony pod względem czasu kwitnienia. Nasiona z tych roślin nie kiełkowały dobrze, a jeżeli miało to miejsce, rośliny zatrzymywały się na etapie siewek. Obydwa efekty można było odwrócić przez przeniesienie roślin do pożywki hodowlanej zawierającej 10-5 M GA3 i hodowanie ich na tej pożywce przez trzy dni. Po odratowaniu i przeniesieniu do gleby rośliny rozwijały się normalnie, z wyjątkiem fenotypu późnego kwitnienia. W warunkach krótkiego dnia, rośliny transgeniczne tworzą znacznie więcej liści rozetowych (ilości wegetatywnych) niż rośliny typu dzikiego (około 40 liści rozetowych i 100 dni po wykiełkowamu, a rośliny nadal nie kwitną, w porównaniu do około 30 liści rozetowych przed kwitnieniem dla roślin typu dzikiego). W warunkach LD w większości tych roślin następuje częściowa rewersja wzrostu generatywnego do wegetatywnego, widoczna przez tworzenie się napowietrznych rozetek (liści wegetatywnych) na łodydze kwiatostanowej. Rośliny (ekotyp C24) zawierające dodatkową kopię cdna ATH1 pod kontrolą promotora genu szoku cieplnego Hsp 18.2 (rośliny HspH1B) również wykazywały fenotyp późnego kwitnienia. Nawet bez szoku cieplnego (wiadomo, ze ten promotor ma również podstawową aktywność bez indukcji) rośliny zawierające ten konstrukt kwitną znacznie później w warunkach LD niż rośliny typu

14 dzikiego (30,5 liści rozetowych wytworzonych w typie dzikim wobec 61 liści rozetowych wytworzonych w roślinach HspHlB-patrz fig. 7). Przykład 8 Wczesne kwitnienie przez ekspresją antysensownego ATH1 Tak jak ektopiczna nadekspresja ATH1, zaburzenie funkcji genu ATH1 może również rzucić światło na funkcję ATH1 w rozwoju rośliny. Zaburzenie funkcji genu uzyskano przez konstytutywną nadekspresję antysensownego ATH1. Antysensowny pełnej długości cdna ATH1 połączono z konstytutywnym promotorem 35S wirusa mozaiki kalafiora i chimero wy gen 35S:antysensowny ATH1 transformowano do Arabidopsis ekotyp C24 stosując transformację korzeniową według Valvekens. Otrzymano dwadzieścia dwa niezalezne transformaty i wszystkie krzyzowano wsobnie. Z każdej linii 10 odrębnych nasion poddano kiełkowaniu w glebie i sprawdzano pod kątem zmienionych fenotypów w porównaniu z roślinami typu dzikiego C24. W pięciu z tych linii rośliny wykazywały fenotyp wczesnego kwitnienia: kwitnienie rozpoczynało się po utworzeniu od sześciu do dziesięciu liści rozetowych w porównaniu do około trzydziestu liści w roślinach typu dzikiego (patrz fig. 7). Przykład 9 Odpowiedź unikania cienia Kiedy rośliny rosną blisko siebie, objawia się syndrom unikania cienia, w którym rośliny reagują na obniżone stosunki świetlne czerwień/daleka czerwień powodowane przez filtrowanie światła czerwonego przez baldachim liści. Prowadzi to w rezultacie do szybkiego i dramatycznego zwiększenia tempa wzrostu łodyg i. ogonków liściowych przy wzroście liści, wytwarzania narządów spichrzowych i rozwoju reproduktywnego, powodując w ten sposób zwiększenie indeksu zbiorów (gdzie indeks zbiorów jest wyrażany jako biomasa liści w proporcji do całej biomasy). Uważa się, że w odpowiedź unikania cienia odbywa się głównie za pośrednictwem fitochromu B i wykazano, ze nadprodukcja fitochromu eliminuje odpowiedź unikania cienia, prowadząc w rezultacie do wzrostu indeksu zbiorów tytoniu rosnącego na polach (Robson i wsp., 1996). Brak fitochromu B prowadzi również do utraty odpowiedzi unikania cienia i w warunkach indukcji prowadzi to do zmniejszenia wydłużania się łodygi w porównaniu z warunkami nieindukcyj nymi. W laboratorium, sytuacje powodujące odpowiedź unikania cienia można naśladować przez dodawanie różnych natężeń strumienia światła dalekiej czerwieni (Frc) do stałego światła białego (Wc). W tych warunkach rośliny Arabidopsis typu dzikiego (C24 wt) wykazują typową odpowiedź unikania cienia (wydłużone hipokotyle przy Wc + Frc w porównaniu z długością hipokotyli przy tylko Wc, podczas gdy mutanty receptora fitochromu B, u których nie ma odpowiedzi unikania cienia, wykazują odpowiedź przeciwną (zmniejszenie długości hipokotyli). Rośliny z antysensownym AtHl również wykazują zmniejszenie długości hipokotyli, a w przypadkach najpoważniejszych u roślin z antysensownym AtHl (takich, jak AtHl#3) to zmniejszenie jest podobne, jak w nasionach mutantów phyb. A zatem można wnioskować, że podobnie jak brak aktywnego fitochromu B, utrata Athl prowadzi w rezultacie do utraty odpowiedzi unikania cienia. Analiza unikania cienia u roślin z antysensownym AtHl (takich, jak AtHl), roślin C24 typu dzikiego (C24 wt) i mutantów fotoreceptora (phyb) jest przedstawiona poniżej. Siewki hodowano przez 2 dni przy ciągłym białym świetle, a następnie 4 dni w tych samych warunkach świetlnych lub 4 dni przy świetle białym uzupełnionym daleką czerwienią. Długość hipokotyli mierzono po 6 dniach wzrostu. Uzyskano następujące wyniki. C24wt Wc = długość hipokotyla 5,5 mm Wc+Frc = długość hipokotyla 7,5 mm PhyB Wc = długość hipokotyla 9,5 mm Wc+Frc = długość hipokotyla 7,3 mm Antysensowny Wc = długość hipokotyla 9,0 mm Athl #3 Wc+Frc = długość hipokotyla 5,0 mm Antysensowny Wc = długość hipokotyla 9,8 mm

15 Athl#7 Wc+Frc = długość hipokotyla 8,0 mm Antysensowny Wc = długość hipokotyla 7,5 mm Athl#23 Wc+Frc = długość hipokotyla 7,0 mm Cytowana literatura: Bagnall, 1993: Ann. Bot Bechtold i wsp., 1993: C. R. Acad. Sci. Paris, Life Sciences 316: Bent i wsp., 1994: Sciece 265: Caplan i wsp., 1985: J. Bacteriology 161: Chambers i wsp., 1988: Gene 68: Frisch i wsp., 1995: Plant Mol. Biol. 27: Gelvin i Schilperoot, 1988: Plant Molecular Biology Manual: Dordrecht: Kluwer AcademicPublisher. Jefferson i wsp., 1987: EMBO J. 6: Koomeef i wsp., 1991: Mol, Gen. Genet. 229:57-66 Last i Gray, 1989: Plant Mol. Biol. 12: Mandel i Yanowski, 1985: Nature 377: Putteril i wsp., 1995: Cell 80: Pwee i Gray, 1993: Plant J. 3: Quaedvlieg i wsp., 1995: The Plant Cell 7: Shannon i Meeks-Wagner, 1991: The Plant Cell: Takahashi i Komeda, 1989: Mol. Gen. Genet. 219:365 Takahashi i wsp., 1992: Plant J. 2: Weigel i Nilsson, 1995: Nature 377: Horsch, R., Fry, F., Hoffman, N., Eichholtz, D., Rogers, S. i Fraley, R. (1985). A simple and general method for transferring genes into plants. Science 227, Murshige, T. i Skoog, F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15, Robson, P.R.H., McCormac, A.C., Irvine, A.S., i Smith, H. (1996). Genetic engineering of harvest index in tobacco through overexpression of a phytochrome gene. Nature Biotechnology 14 (8), Wilson, R.N., Heckman, J.W., i Sommerville, C.R. (1992). Gibberellin is requred for flowering in Arabidopsis thaliana under short days. Plant. Phys. 100,

16 FIG.1
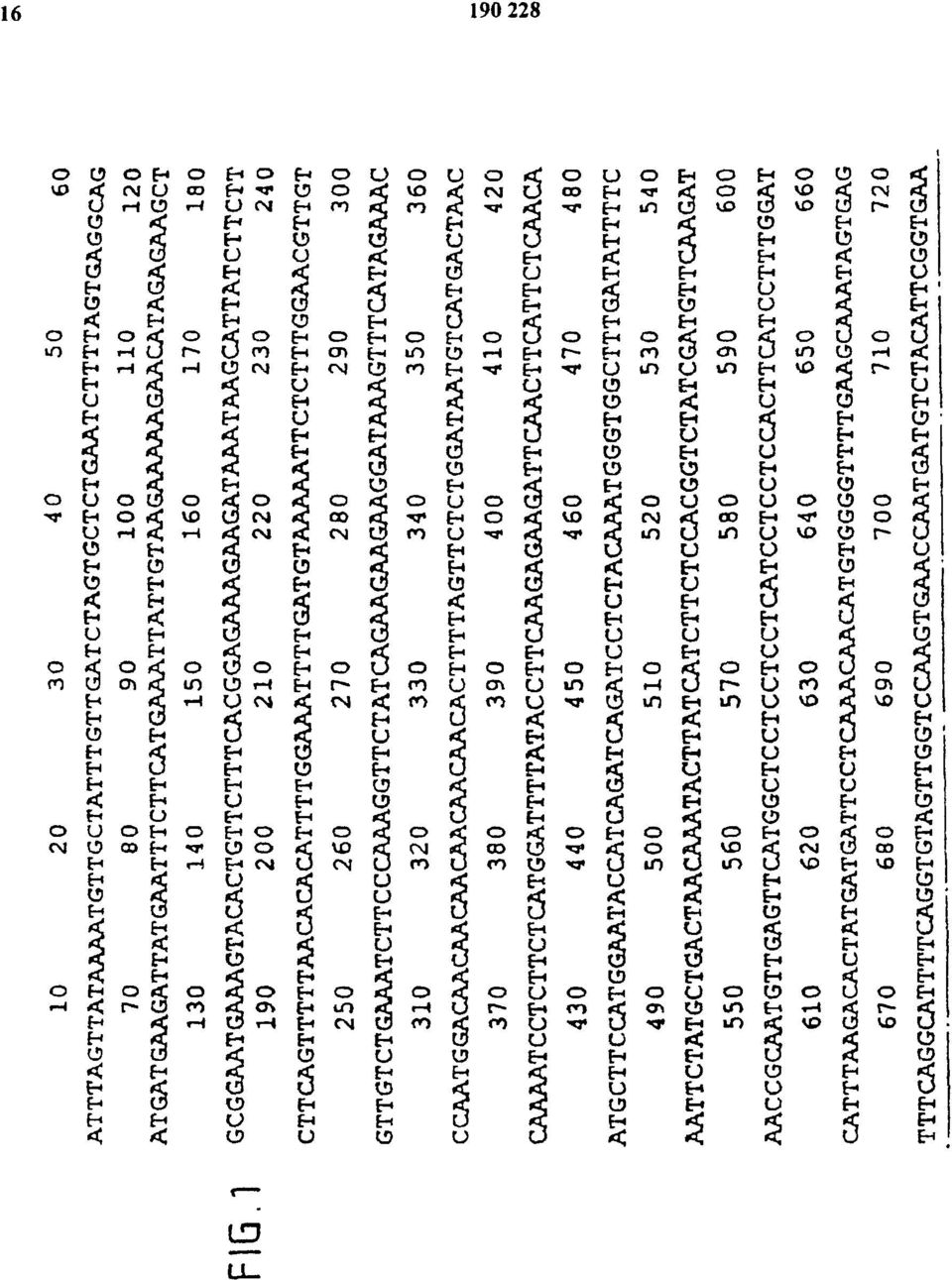
17 FIG.1 (C.D)
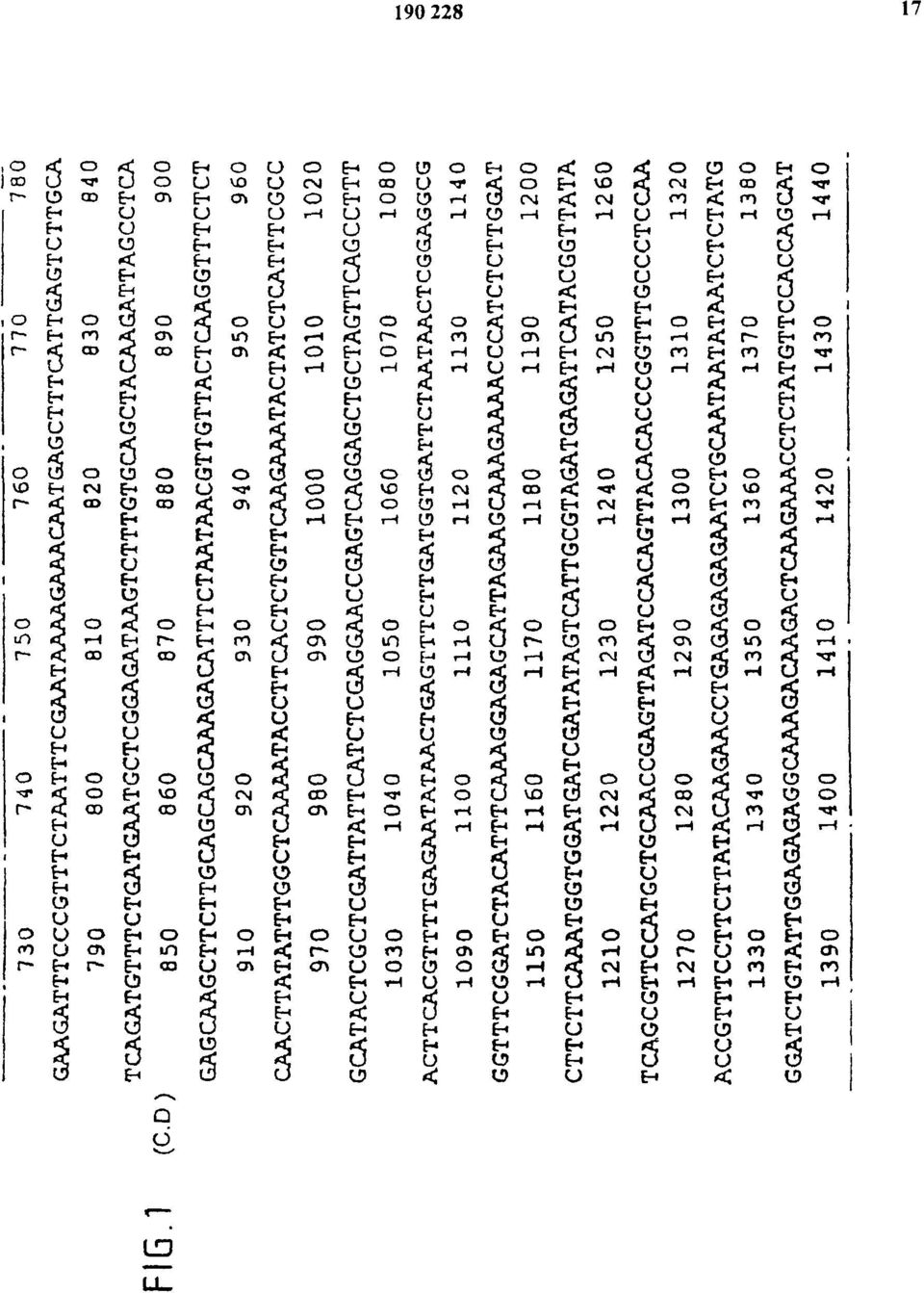
18 FIG. 1 (C-D.)
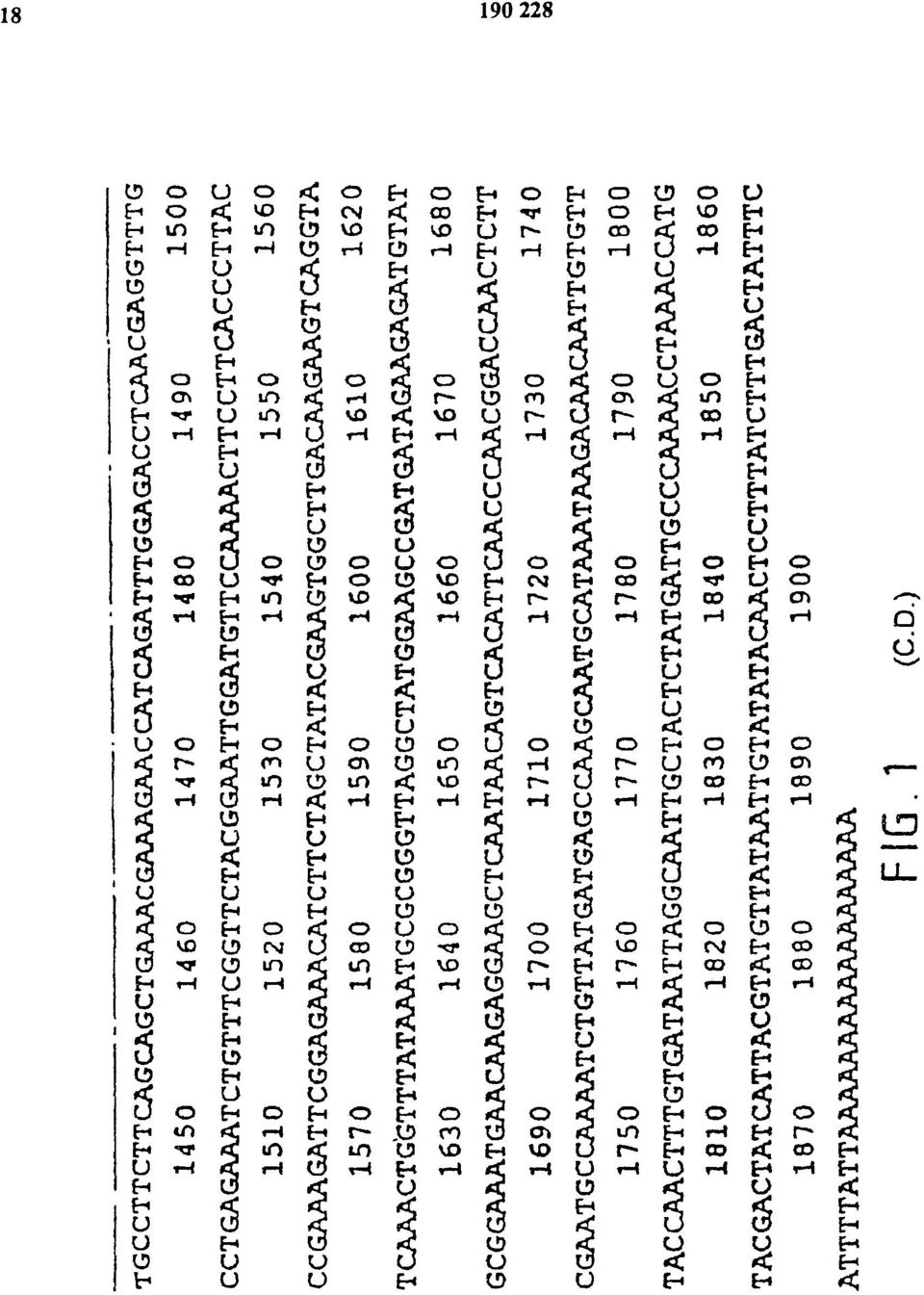
19 FIG
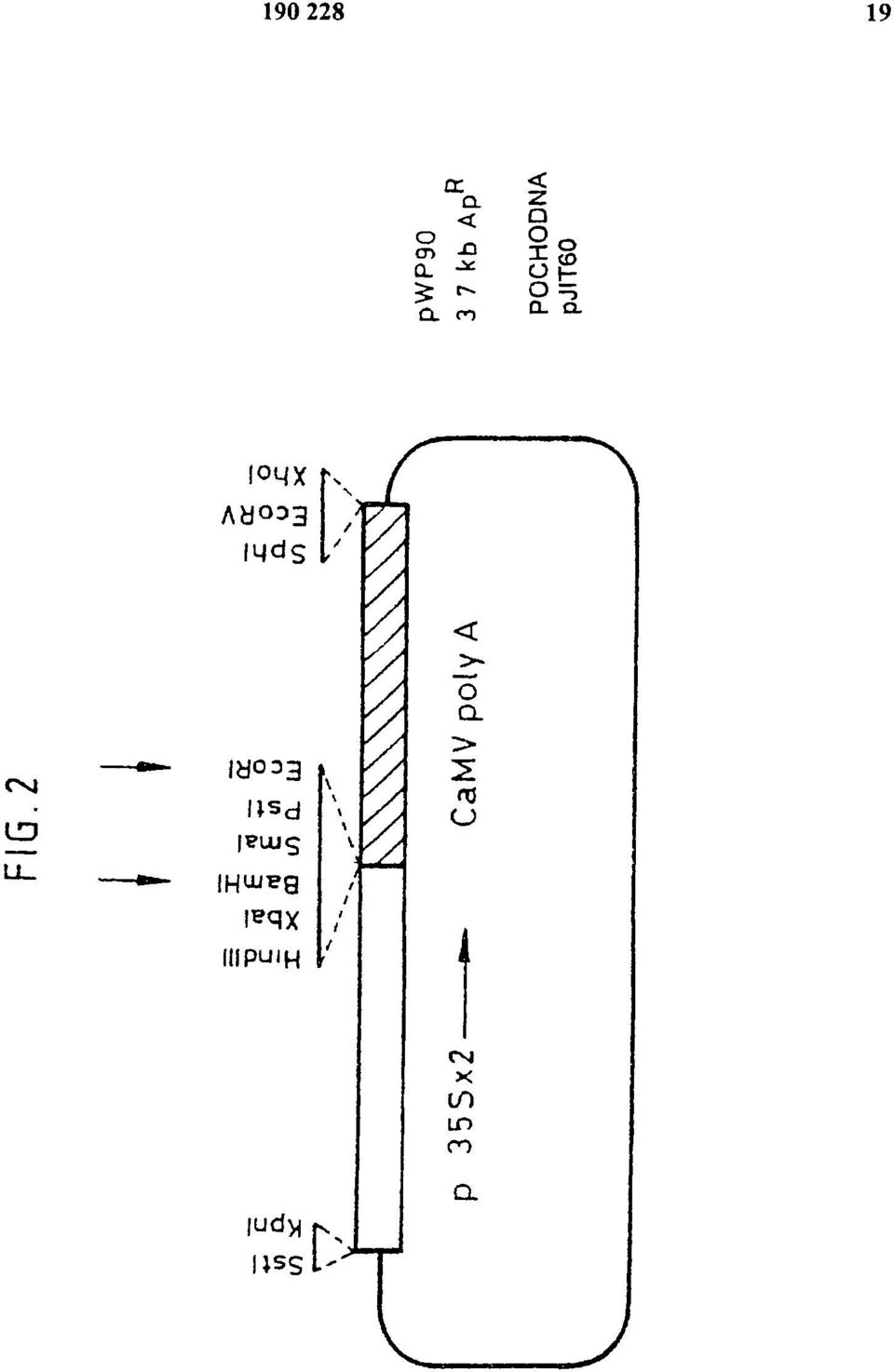
20 FIG. 3
Klonowanie molekularne Kurs doskonalący. Zakład Geriatrii i Gerontologii CMKP
 Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Klonowanie molekularne Kurs doskonalący Zakład Geriatrii i Gerontologii CMKP Etapy klonowania molekularnego 1. Wybór wektora i organizmu gospodarza Po co klonuję (do namnożenia DNA [czy ma być metylowane
Hormony roślinne ( i f t i o t h o or o m r on o y n )
") Hormony roślinne (fitohormony) Hormony roślinne: To związki chemiczne syntetyzowane w pewnych częściach rośliny służące do "komunikacji" pomiędzy poszczególnymi jej częściami. Działają w bardzo małych
Hormony roślinne (fitohormony) Hormony roślinne: To związki chemiczne syntetyzowane w pewnych częściach rośliny służące do "komunikacji" pomiędzy poszczególnymi jej częściami. Działają w bardzo małych
Zagadnienia: Wzrost i rozwój
 Zagadnienia: Wzrost i rozwój 1. Definicja wzrostu i rozwoju. 2. Fazy wzrostu i rozwoju (embrionalna, juwenilna, wegetatywna, generatywna). 3. Wpływ czynników środowiska na wzrost i rozwój roślin. 4. Kiełkowanie
Zagadnienia: Wzrost i rozwój 1. Definicja wzrostu i rozwoju. 2. Fazy wzrostu i rozwoju (embrionalna, juwenilna, wegetatywna, generatywna). 3. Wpływ czynników środowiska na wzrost i rozwój roślin. 4. Kiełkowanie
Inżynieria Genetyczna ćw. 3
 Materiały do ćwiczeń z przedmiotu Genetyka z inżynierią genetyczną D - blok Inżynieria Genetyczna ćw. 3 Instytut Genetyki i Biotechnologii, Wydział Biologii, Uniwersytet Warszawski, rok akad. 2018/2019
Materiały do ćwiczeń z przedmiotu Genetyka z inżynierią genetyczną D - blok Inżynieria Genetyczna ćw. 3 Instytut Genetyki i Biotechnologii, Wydział Biologii, Uniwersytet Warszawski, rok akad. 2018/2019
WYKŁAD XIII ROŚLINY WZROST I ROZWÓJ
 WYKŁAD XIII ROŚLINY WZROST I ROZWÓJ Podstawowe objawy życia: Przemiana materii (metabolizm) WZROST I ROZWÓJ Wzrost - nieodwracalny przyrost rozmiarów rośliny Rozwój - zmiany jakościowe zachodzące w ciągu
WYKŁAD XIII ROŚLINY WZROST I ROZWÓJ Podstawowe objawy życia: Przemiana materii (metabolizm) WZROST I ROZWÓJ Wzrost - nieodwracalny przyrost rozmiarów rośliny Rozwój - zmiany jakościowe zachodzące w ciągu
Inżynieria genetyczna- 6 ECTS. Inżynieria genetyczna. Podstawowe pojęcia Część II Klonowanie ekspresyjne Od genu do białka
 Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Inżynieria genetyczna- 6 ECTS Część I Badanie ekspresji genów Podstawy klonowania i różnicowania transformantów Kolokwium (14pkt) Część II Klonowanie ekspresyjne Od genu do białka Kolokwium (26pkt) EGZAMIN
Dr. habil. Anna Salek International Bio-Consulting 1 Germany
 1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
1 2 3 Drożdże są najprostszymi Eukariontami 4 Eucaryota Procaryota 5 6 Informacja genetyczna dla każdej komórki drożdży jest identyczna A zatem każda komórka koduje w DNA wszystkie swoje substancje 7 Przy
Nowoczesne systemy ekspresji genów
 Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
Nowoczesne systemy ekspresji genów Ekspresja genów w organizmach żywych GEN - pojęcia podstawowe promotor sekwencja kodująca RNA terminator gen Gen - odcinek DNA zawierający zakodowaną informację wystarczającą
TATA box. Enhancery. CGCG ekson intron ekson intron ekson CZĘŚĆ KODUJĄCA GENU TERMINATOR. Elementy regulatorowe
 Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
Promotory genu Promotor bliski leży w odległości do 40 pz od miejsca startu transkrypcji, zawiera kasetę TATA. Kaseta TATA to silnie konserwowana sekwencja TATAAAA, występująca w większości promotorów
TRANSKRYPCJA - I etap ekspresji genów
 Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
Eksparesja genów TRANSKRYPCJA - I etap ekspresji genów Przepisywanie informacji genetycznej z makrocząsteczki DNA na mniejsze i bardziej funkcjonalne cząsteczki pre-mrna Polimeraza RNA ETAP I Inicjacja
października 2013: Elementarz biologii molekularnej. Wykład nr 2 BIOINFORMATYKA rok II
 10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
10 października 2013: Elementarz biologii molekularnej www.bioalgorithms.info Wykład nr 2 BIOINFORMATYKA rok II Komórka: strukturalna i funkcjonalne jednostka organizmu żywego Jądro komórkowe: chroniona
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ 1. Gen to odcinek DNA odpowiedzialny
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ 1. Gen to odcinek DNA odpowiedzialny
Badanie funkcji genu
 Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Auksyna,,oczami roślin transgenicznych
 Auksyna,,oczami roślin transgenicznych dr Justyna Wiśniewska, UNIWERSYTET MIKOŁAJA KOPERNIKA w TORUNIU ZAKŁAD BIOTECHNOLOGII Auksyny naturalne i sztuczne Naturalne auksyny: IAA - kwas indolilo-3-octowy
Auksyna,,oczami roślin transgenicznych dr Justyna Wiśniewska, UNIWERSYTET MIKOŁAJA KOPERNIKA w TORUNIU ZAKŁAD BIOTECHNOLOGII Auksyny naturalne i sztuczne Naturalne auksyny: IAA - kwas indolilo-3-octowy
Imię i nazwisko...kl...
 Gimnazjum nr 4 im. Ojca Świętego Jana Pawła II we Wrocławiu SPRAWDZIAN GENETYKA GR. A Imię i nazwisko...kl.... 1. Nauka o regułach i mechanizmach dziedziczenia to: (0-1pkt) a) cytologia b) biochemia c)
Gimnazjum nr 4 im. Ojca Świętego Jana Pawła II we Wrocławiu SPRAWDZIAN GENETYKA GR. A Imię i nazwisko...kl.... 1. Nauka o regułach i mechanizmach dziedziczenia to: (0-1pkt) a) cytologia b) biochemia c)
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE. Ewa Waszkowska ekspert UPRP
 WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
WYNALAZKI BIOTECHNOLOGICZNE W POLSCE Ewa Waszkowska ekspert UPRP Źródła informacji w biotechnologii projekt SLING Warszawa, 9-10.12.2010 PLAN WYSTĄPIENIA Umocowania prawne Wynalazki biotechnologiczne Statystyka
Rośliny modyfikowane genetycznie (GMO)
") Rośliny modyfikowane genetycznie (GMO) Organizmy modyfikowane genetycznie Organizm zmodyfikowany genetycznie (międzynarodowy skrót: GMO Genetically Modified Organizm) to organizm o zmienionych cechach,
Rośliny modyfikowane genetycznie (GMO) Organizmy modyfikowane genetycznie Organizm zmodyfikowany genetycznie (międzynarodowy skrót: GMO Genetically Modified Organizm) to organizm o zmienionych cechach,
Bezpośrednia embriogeneza somatyczna
 Bezpośrednia embriogeneza somatyczna Zarodki somatyczne formują się bezpośrednio tylko z tych komórek roślinnych, które są kompetentne już w momencie izolowania z rośliny macierzystej, czyli z proembriogenicznie
Bezpośrednia embriogeneza somatyczna Zarodki somatyczne formują się bezpośrednio tylko z tych komórek roślinnych, które są kompetentne już w momencie izolowania z rośliny macierzystej, czyli z proembriogenicznie
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii
 Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Możliwości współczesnej inżynierii genetycznej w obszarze biotechnologii 1. Technologia rekombinowanego DNA jest podstawą uzyskiwania genetycznie zmodyfikowanych organizmów 2. Medycyna i ochrona zdrowia
Rośliny z probówki. Jak powstają? Alina Trejgell & Agata Stawicka, UMK
 Rośliny z probówki Jak powstają? I. Dojrzałe i niedojrzałe nasiona szarotka (Leontopodium alpinum) II. Inne organy roślin wyka (Vicia sepium) zarodki zygotyczne pąki kwiatowe wilca (Pharbitis nil) korzeń
Rośliny z probówki Jak powstają? I. Dojrzałe i niedojrzałe nasiona szarotka (Leontopodium alpinum) II. Inne organy roślin wyka (Vicia sepium) zarodki zygotyczne pąki kwiatowe wilca (Pharbitis nil) korzeń
(86) Data i numer zgłoszenia międzynarodowego: ,PCT/GB00/00248 (87) Data i numer publikacji zgłoszenia międzynarodowego:
 Data i numer zgłoszenia międzynarodowego: ,PCT/GB00/00248 (87) Data i numer publikacji zgłoszenia międzynarodowego:") RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 202462 (21) Numer zgłoszenia: 349486 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 28.01.2000 (86) Data i numer zgłoszenia
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 202462 (21) Numer zgłoszenia: 349486 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 28.01.2000 (86) Data i numer zgłoszenia
Badanie funkcji genu
 Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Badanie funkcji genu Funkcję genu można zbadać różnymi sposobami Przypadkowa analizy funkcji genu MUTACJA FENOTYP GEN Strategia ukierunkowanej analizy funkcji genu GEN 1. wprowadzenie mutacji w genie 2.
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu np. w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna
 Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Wybrane techniki badania białek -proteomika funkcjonalna Proteomika: umożliwia badanie zestawu wszystkich (lub prawie wszystkich) białek komórkowych Zalety analizy proteomu w porównaniu z analizą trankryptomu:
Powodzenie reakcji PCR wymaga właściwego doboru szeregu parametrów:
 Powodzenie reakcji PCR wymaga właściwego doboru szeregu parametrów: dobór warunków samej reakcji PCR (temperatury, czas trwania cykli, ilości cykli itp.) dobór odpowiednich starterów do reakcji amplifikacji
Powodzenie reakcji PCR wymaga właściwego doboru szeregu parametrów: dobór warunków samej reakcji PCR (temperatury, czas trwania cykli, ilości cykli itp.) dobór odpowiednich starterów do reakcji amplifikacji
Tematyka zajęć z biologii
 Tematyka zajęć z biologii klasy: I Lp. Temat zajęć Zakres treści 1 Zapoznanie z przedmiotowym systemem oceniania, wymaganiami edukacyjnymi i podstawą programową Podstawowe zagadnienia materiału nauczania
Tematyka zajęć z biologii klasy: I Lp. Temat zajęć Zakres treści 1 Zapoznanie z przedmiotowym systemem oceniania, wymaganiami edukacyjnymi i podstawą programową Podstawowe zagadnienia materiału nauczania
KLONOWANIE DNA REKOMBINACJA DNA WEKTORY
 KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
KLONOWANIE DNA Klonowanie DNA jest techniką powielania fragmentów DNA DNA można powielać w komórkach (replikacja in vivo) W probówce (PCR) Do przeniesienia fragmentu DNA do komórek gospodarza potrzebny
The Role of Maf1 Protein in trna Processing and Stabilization / Rola białka Maf1 w dojrzewaniu i kontroli stabilności trna
 Streszczenie rozprawy doktorskiej pt. The Role of Maf1 Protein in trna Processing and Stabilization / Rola białka Maf1 w dojrzewaniu i kontroli stabilności trna mgr Tomasz Turowski, promotor prof. dr hab.
Streszczenie rozprawy doktorskiej pt. The Role of Maf1 Protein in trna Processing and Stabilization / Rola białka Maf1 w dojrzewaniu i kontroli stabilności trna mgr Tomasz Turowski, promotor prof. dr hab.
Metody analizy genomu
 Metody analizy genomu 1. Mapowanie restrykcyjne. 2. Sondy do rozpoznawania DNA 3. FISH 4. Odczytanie sekwencji DNA 5. Interpretacja sekwencji DNA genomu 6. Transkryptom 7. Proteom 1. Mapy restrykcyjne
Metody analizy genomu 1. Mapowanie restrykcyjne. 2. Sondy do rozpoznawania DNA 3. FISH 4. Odczytanie sekwencji DNA 5. Interpretacja sekwencji DNA genomu 6. Transkryptom 7. Proteom 1. Mapy restrykcyjne
Przedmowa 9 Początki hodowli i oceny odmian roślin warzywnych w Polsce Hodowla roślin kapustnych Znaczenie gospodarcze Systematy
 Przedmowa Przekazywana czytelnikowi książka jest podręcznikiem szczegółowej hodowli wybranych, uprawianych w Polsce gatunków roślin warzywnych. Do tej pory wydano w Polsce w 1967 roku jeden podręcznik
Przedmowa Przekazywana czytelnikowi książka jest podręcznikiem szczegółowej hodowli wybranych, uprawianych w Polsce gatunków roślin warzywnych. Do tej pory wydano w Polsce w 1967 roku jeden podręcznik
Dane mikromacierzowe. Mateusz Markowicz Marta Stańska
 Dane mikromacierzowe Mateusz Markowicz Marta Stańska Mikromacierz Mikromacierz DNA (ang. DNA microarray) to szklana lub plastikowa płytka (o maksymalnych wymiarach 2,5 cm x 7,5 cm) z naniesionymi w regularnych
Dane mikromacierzowe Mateusz Markowicz Marta Stańska Mikromacierz Mikromacierz DNA (ang. DNA microarray) to szklana lub plastikowa płytka (o maksymalnych wymiarach 2,5 cm x 7,5 cm) z naniesionymi w regularnych
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Transkrypcja RNA
 SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Transkrypcja RNA SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy. 1. Karta
SCENARIUSZ LEKCJI BIOLOGII Z WYKORZYSTANIEM FILMU Transkrypcja RNA SPIS TREŚCI: I. Wprowadzenie. II. Części lekcji. 1. Część wstępna. 2. Część realizacji. 3. Część podsumowująca. III. Karty pracy. 1. Karta
Roślinne kultury tkankowe in vitro hodowla roślin, części roślin, tkanek lub pojedynczych komórek na sztucznych pożywkach w sterylnych warunkach.
 Roślinne kultury tkankowe in vitro hodowla roślin, części roślin, tkanek lub pojedynczych komórek na sztucznych pożywkach w sterylnych warunkach. TOTIPOTENCJA Zdolności do odtworzenia poszczególnych organów,
Roślinne kultury tkankowe in vitro hodowla roślin, części roślin, tkanek lub pojedynczych komórek na sztucznych pożywkach w sterylnych warunkach. TOTIPOTENCJA Zdolności do odtworzenia poszczególnych organów,
CROPVIT - ŹRÓDŁO NIEZBĘDNYCH MIKROSKŁADNIKÓW MIKROELEMENTY SCHELATOWANE ZAPEWNIAJĄ SZYBKIE WCHŁANIANIE PRZEZ ROŚLINĘ SZYBKO POPRAWIAJĄ KONDYCJĘ
 CROPVIT - ŹRÓDŁO NIEZBĘDNYCH MIKROSKŁADNIKÓW MIKROELEMENTY SCHELATOWANE ZAPEWNIAJĄ SZYBKIE WCHŁANIANIE PRZEZ ROŚLINĘ SZYBKO POPRAWIAJĄ KONDYCJĘ ROŚLIN UPRAWNYCH UCZESTNICZĄ W REAKCJACH SYNTEZY BIAŁEK,
CROPVIT - ŹRÓDŁO NIEZBĘDNYCH MIKROSKŁADNIKÓW MIKROELEMENTY SCHELATOWANE ZAPEWNIAJĄ SZYBKIE WCHŁANIANIE PRZEZ ROŚLINĘ SZYBKO POPRAWIAJĄ KONDYCJĘ ROŚLIN UPRAWNYCH UCZESTNICZĄ W REAKCJACH SYNTEZY BIAŁEK,
Praca kontrolna z biologii LO dla dorosłych semestr V
 Praca kontrolna z biologii LO dla dorosłych semestr V Poniższa praca składa się z 15 zadań. Przy każdym poleceniu podano liczbę punktów możliwą do uzyskania za prawidłową odpowiedź. Za rozwiązanie zadań
Praca kontrolna z biologii LO dla dorosłych semestr V Poniższa praca składa się z 15 zadań. Przy każdym poleceniu podano liczbę punktów możliwą do uzyskania za prawidłową odpowiedź. Za rozwiązanie zadań
Prezentuje: Magdalena Jasińska
 Prezentuje: Magdalena Jasińska W którym momencie w rozwoju embrionalnym myszy rozpoczyna się endogenna transkrypcja? Hipoteza I: Endogenna transkrypcja rozpoczyna się w embrionach będących w stadium 2-komórkowym
Prezentuje: Magdalena Jasińska W którym momencie w rozwoju embrionalnym myszy rozpoczyna się endogenna transkrypcja? Hipoteza I: Endogenna transkrypcja rozpoczyna się w embrionach będących w stadium 2-komórkowym
1. Biotechnologia i inżynieria genetyczna zagadnienia wstępne 13
 Spis treści Przedmowa 11 1. Biotechnologia i inżynieria genetyczna zagadnienia wstępne 13 1.1. Wprowadzenie 13 1.2. Biotechnologia żywności znaczenie gospodarcze i społeczne 13 1.3. Produkty modyfikowane
Spis treści Przedmowa 11 1. Biotechnologia i inżynieria genetyczna zagadnienia wstępne 13 1.1. Wprowadzenie 13 1.2. Biotechnologia żywności znaczenie gospodarcze i społeczne 13 1.3. Produkty modyfikowane
DR ŻANETA PACUD Zdolność patentowa roślin
 DR ŻANETA PACUD Zdolność patentowa roślin czyli rzecz o brokułach i pomidorach Sposoby ochrony prawnej roślin wprowadzenie Ochrona prawna odmian roślin - Międzynarodowa konwencja o ochronie nowych odmian
DR ŻANETA PACUD Zdolność patentowa roślin czyli rzecz o brokułach i pomidorach Sposoby ochrony prawnej roślin wprowadzenie Ochrona prawna odmian roślin - Międzynarodowa konwencja o ochronie nowych odmian
Regulacja wzrostu zbóż
 Regulacja wzrostu zbóż Kluczowe fazy rozwojowe Opracowanie dr hab. Kinga Matysiak, IOR-PIB, Poznań. Fot. Syngenta Ze środków ochrony roślin należy korzystać z zachowaniem bezpieczeństwa. Przed każdym użyciem
Regulacja wzrostu zbóż Kluczowe fazy rozwojowe Opracowanie dr hab. Kinga Matysiak, IOR-PIB, Poznań. Fot. Syngenta Ze środków ochrony roślin należy korzystać z zachowaniem bezpieczeństwa. Przed każdym użyciem
Ocena rozprawy doktorskiej mgr Justyny Kowalczyk
 Dr hab. Paweł Bednarek, prof. IChB PAN Instytut Chemii Bioorganicznej PAN ul. Noskowskiego 12/14 61-704 Poznań Ocena rozprawy doktorskiej mgr Justyny Kowalczyk Identyfikacja i charakterystyka nowego regulatora
Dr hab. Paweł Bednarek, prof. IChB PAN Instytut Chemii Bioorganicznej PAN ul. Noskowskiego 12/14 61-704 Poznań Ocena rozprawy doktorskiej mgr Justyny Kowalczyk Identyfikacja i charakterystyka nowego regulatora
gibereliny naturalna : GA 3 (kwas giberelowy)
") gibereliny naturalna : GA 3 (kwas giberelowy) Miejsce wytwarzania: w dojrzewających nasionach, owocach, zielonych częściach rośliny, w wierzchołkach wzrostu pędu, korzeniach i pręcikach. Biosynteza w plastydach
gibereliny naturalna : GA 3 (kwas giberelowy) Miejsce wytwarzania: w dojrzewających nasionach, owocach, zielonych częściach rośliny, w wierzchołkach wzrostu pędu, korzeniach i pręcikach. Biosynteza w plastydach
Celem projektu jest zbadanie mechanizmu aborcji kwiatów i zarodków gryki zwyczajnej pod wpływem stresu termicznego lub troficznego.
 Raport za rok 2018 z projektu NCN 2017/25/B/NZ9/00148 pt. Badanie mechanizmu degeneracji woreczkówzalążkowych i aborcji kwiatów jako przyczyny słabego zawiązywania nasion gryki zwyczajnej(fagopyrum esculentum
Raport za rok 2018 z projektu NCN 2017/25/B/NZ9/00148 pt. Badanie mechanizmu degeneracji woreczkówzalążkowych i aborcji kwiatów jako przyczyny słabego zawiązywania nasion gryki zwyczajnej(fagopyrum esculentum
METODYKA STOSOWANA W ZAKŁADZIE BIOLOGII ROZWOJU ROŚLIN
 METODYKA STOSOWANA W ZAKŁADZIE BIOLOGII ROZWOJU ROŚLIN Immunolokalizacja wybranych białek i polisacharydów Ksyloglukan u Arabidopsis Kaloza w gametofiach mszaków Immunocytochemia białek cytoszkieletu kortykalnego
METODYKA STOSOWANA W ZAKŁADZIE BIOLOGII ROZWOJU ROŚLIN Immunolokalizacja wybranych białek i polisacharydów Ksyloglukan u Arabidopsis Kaloza w gametofiach mszaków Immunocytochemia białek cytoszkieletu kortykalnego
Ćwiczenia 1 Wirtualne Klonowanie Prowadzący: mgr inż. Joanna Tymeck-Mulik i mgr Lidia Gaffke. Część teoretyczna:
 Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
Uniwersytet Gdański, Wydział Biologii Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią Biologia II rok ===============================================================================================
Fizjologiczne i molekularne markery tolerancji buraka cukrowego na suszę. Dr Danuta Chołuj
 Fizjologiczne i molekularne markery tolerancji buraka cukrowego na suszę Dr Danuta Chołuj Szacunkowe straty plonu buraków cukrowych w Europie na skutek suszy kształtują się pomiędzy 5 a 30 % W jakiej fazie
Fizjologiczne i molekularne markery tolerancji buraka cukrowego na suszę Dr Danuta Chołuj Szacunkowe straty plonu buraków cukrowych w Europie na skutek suszy kształtują się pomiędzy 5 a 30 % W jakiej fazie
Spis treści Część I. Genetyczne podstawy hodowli roślin 1. Molekularne podstawy dziedziczenia cech Dariusz Crzebelus, Adeta Adamus, Maria Klein
 Spis treści Część I. Genetyczne podstawy hodowli roślin 1. Molekularne podstawy dziedziczenia cech... 15 Dariusz Crzebelus, Adeta Adamus, Maria Klein 1.1. Budowa DNA i przepływ informacji genetycznej...
Spis treści Część I. Genetyczne podstawy hodowli roślin 1. Molekularne podstawy dziedziczenia cech... 15 Dariusz Crzebelus, Adeta Adamus, Maria Klein 1.1. Budowa DNA i przepływ informacji genetycznej...
TECHNIKI ANALIZY RNA TECHNIKI ANALIZY RNA TECHNIKI ANALIZY RNA
 DNA 28SRNA 18/16S RNA 5SRNA mrna Ilościowa analiza mrna aktywność genów w zależności od wybranych czynników: o rodzaju tkanki o rodzaju czynnika zewnętrznego o rodzaju upośledzenia szlaku metabolicznego
DNA 28SRNA 18/16S RNA 5SRNA mrna Ilościowa analiza mrna aktywność genów w zależności od wybranych czynników: o rodzaju tkanki o rodzaju czynnika zewnętrznego o rodzaju upośledzenia szlaku metabolicznego
Metody badania ekspresji genów
 Metody badania ekspresji genów dr Katarzyna Knapczyk-Stwora Warunki wstępne: Proszę zapoznać się z tematem Metody badania ekspresji genów zamieszczonym w skrypcie pod reakcją A. Lityńskiej i M. Lewandowskiego
Metody badania ekspresji genów dr Katarzyna Knapczyk-Stwora Warunki wstępne: Proszę zapoznać się z tematem Metody badania ekspresji genów zamieszczonym w skrypcie pod reakcją A. Lityńskiej i M. Lewandowskiego
Aneta Gerszberg i Andrzej K. Kononowicz. Zakład Cytogenetyki i Biologii Molekularnej Roślin Uniwersytet Łódzki
 Aneta Gerszberg i Andrzej K. Kononowicz Zakład Cytogenetyki i Biologii Molekularnej Roślin Uniwersytet Łódzki Molekularna uprawa Termin molekularna uprawa (ang. molecular farming), odnosi się do produkcji
Aneta Gerszberg i Andrzej K. Kononowicz Zakład Cytogenetyki i Biologii Molekularnej Roślin Uniwersytet Łódzki Molekularna uprawa Termin molekularna uprawa (ang. molecular farming), odnosi się do produkcji
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych
 Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
Proteomika: umożliwia badanie zestawu wszystkich lub prawie wszystkich białek komórkowych Zalety w porównaniu z analizą trankryptomu: analiza transkryptomu komórki identyfikacja mrna nie musi jeszcze oznaczać
PL B1 (12) OPIS PATENTOWY (19) PL (11)
 OPIS PATENTOWY (19) PL (11)") RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 188253 (21) Numer zgłoszenia: 335725 (13) B1 (22) Data zgłoszenia: 09.04.1998 (86) Data i numer zgłoszenia
RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (11) 188253 (21) Numer zgłoszenia: 335725 (13) B1 (22) Data zgłoszenia: 09.04.1998 (86) Data i numer zgłoszenia
Bloki licencjackie i studia magisterskie na Kierunkach: Biotechnologia, specjalność Biotechnologia roślinna oraz Genetyka
 Bloki licencjackie i studia magisterskie na Kierunkach: Biotechnologia, specjalność Biotechnologia roślinna oraz Genetyka INSTYTUT BIOLOGII EKSPERYMENTALNEJ W Katedrze Genetyki Ogólnej, Biologii Molekularnej
Bloki licencjackie i studia magisterskie na Kierunkach: Biotechnologia, specjalność Biotechnologia roślinna oraz Genetyka INSTYTUT BIOLOGII EKSPERYMENTALNEJ W Katedrze Genetyki Ogólnej, Biologii Molekularnej
SILVIT. Składniki pokarmowe [g/l lub g/kg] K2O SO3 B Zn SiO2 Aminokwasy 100 25 1,25 0,25 150 +
![SILVIT. Składniki pokarmowe [g/l lub g/kg] K2O SO3 B Zn SiO2 Aminokwasy 100 25 1,25 0,25 150 +](/thumbs/39/20204340.jpg "SILVIT. Składniki pokarmowe [g/l lub g/kg] K2O SO3 B Zn SiO2 Aminokwasy 100 25 1,25 0,25 150 +") SILVIT Biosytmulatory i antystresanty płynne. Preparat nawozowy o właściwościach stymulujących, zawiera aktywny krzem w pełni przyswajalny przez rośliny. Składniki pokarmowe [g/l lub g/kg] K2O SO3 B Zn
SILVIT Biosytmulatory i antystresanty płynne. Preparat nawozowy o właściwościach stymulujących, zawiera aktywny krzem w pełni przyswajalny przez rośliny. Składniki pokarmowe [g/l lub g/kg] K2O SO3 B Zn
Przeglądanie bibliotek
 Przeglądanie bibliotek Czyli jak złapać (i sklonować) ciekawy gen? Klonowanie genów w oparciu o identyczność lub podobieństwo ich sekwencji do znanego już DNA Sonda homologiczna (komplementarna w 100%)
Przeglądanie bibliotek Czyli jak złapać (i sklonować) ciekawy gen? Klonowanie genów w oparciu o identyczność lub podobieństwo ich sekwencji do znanego już DNA Sonda homologiczna (komplementarna w 100%)
Księgarnia PWN: Biotechnologia roślin, redakcja naukowa: Stefan Malepszy SPIS TREŚCI
 Księgarnia PWN: Biotechnologia roślin, redakcja naukowa: Stefan Malepszy SPIS TREŚCI 1 Wprowadzenie 15 2 Metoda kultury in vitro 19 2.1. Kultura komórek i tkanek............................... 19 2.1.1.
Księgarnia PWN: Biotechnologia roślin, redakcja naukowa: Stefan Malepszy SPIS TREŚCI 1 Wprowadzenie 15 2 Metoda kultury in vitro 19 2.1. Kultura komórek i tkanek............................... 19 2.1.1.
Basfoliar Kelp P-Max. Nawóz dolistny: Producent: COMPO Polska Sp. z o.o. Działanie:
 Nawóz dolistny: Basfoliar Kelp P-Max Producent: COMPO Polska Sp. z o.o. Grupa Rolnik jest wyłącznym dystrybutorem nawozu płynnego otrzymany z ekstraktu alg Ecklonia maxima z dodatkiem NPK (4++2). Basfoliar
Nawóz dolistny: Basfoliar Kelp P-Max Producent: COMPO Polska Sp. z o.o. Grupa Rolnik jest wyłącznym dystrybutorem nawozu płynnego otrzymany z ekstraktu alg Ecklonia maxima z dodatkiem NPK (4++2). Basfoliar
dr hab. Tomasz Pawłowski, prof. ID PAN Kórnik,
 dr hab. Tomasz Pawłowski, prof. ID PAN Kórnik, 14.03.2019 Ocena rozprawy doktorskiej mgr inż. Marleny Stawskiej Tytuł rozprawy: Rola wybranych elementów szlaku sygnałowego światła w regulacji kiełkowania
dr hab. Tomasz Pawłowski, prof. ID PAN Kórnik, 14.03.2019 Ocena rozprawy doktorskiej mgr inż. Marleny Stawskiej Tytuł rozprawy: Rola wybranych elementów szlaku sygnałowego światła w regulacji kiełkowania
Wykład 14 Biosynteza białek
 BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
BIOCHEMIA Kierunek: Technologia Żywności i Żywienie Człowieka semestr III Wykład 14 Biosynteza białek WYDZIAŁ NAUK O ŻYWNOŚCI I RYBACTWA CENTRUM BIOIMMOBILIZACJI I INNOWACYJNYCH MATERIAŁÓW OPAKOWANIOWYCH
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA
 Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa IZOLACJA DNA Z HODOWLI KOMÓRKOWEJ.
Zakład Biologii Molekularnej Materiały do ćwiczeń z przedmiotu: BIOLOGIA MOLEKULARNA Zakład Biologii Molekularnej Wydział Farmaceutyczny, WUM ul. Banacha 1, 02-097 Warszawa IZOLACJA DNA Z HODOWLI KOMÓRKOWEJ.
SCENARIUSZ LEKCJI. TEMAT LEKCJI: Podstawowe techniki inżynierii genetycznej. Streszczenie
 SCENARIUSZ LEKCJI OPRACOWANY W RAMACH PROJEKTU: INFORMATYKA MÓJ SPOSÓB NA POZNANIE I OPISANIE ŚWIATA. PROGRAM NAUCZANIA INFORMATYKI Z ELEMENTAMI PRZEDMIOTÓW MATEMATYCZNO-PRZYRODNICZYCH Autorzy scenariusza:
SCENARIUSZ LEKCJI OPRACOWANY W RAMACH PROJEKTU: INFORMATYKA MÓJ SPOSÓB NA POZNANIE I OPISANIE ŚWIATA. PROGRAM NAUCZANIA INFORMATYKI Z ELEMENTAMI PRZEDMIOTÓW MATEMATYCZNO-PRZYRODNICZYCH Autorzy scenariusza:
Wzrost i rozwój roślin
 Wykład 11 i 12 Wzrost i rozwój - definicje Wzrost roślin - powiększanie się rozmiarów rośliny, co można określić ilościowo (wzrost świeżej masy, wysokości, itp.). Zachodzi on poprzez podziały (wzrost embrionalny)
Wykład 11 i 12 Wzrost i rozwój - definicje Wzrost roślin - powiększanie się rozmiarów rośliny, co można określić ilościowo (wzrost świeżej masy, wysokości, itp.). Zachodzi on poprzez podziały (wzrost embrionalny)
Transformacja pośrednia składa się z trzech etapów:
 Transformacja pośrednia składa się z trzech etapów: 1. Otrzymanie pożądanego odcinka DNA z materiału genetycznego dawcy 2. Wprowadzenie obcego DNA do wektora 3. Wprowadzenie wektora, niosącego w sobie
Transformacja pośrednia składa się z trzech etapów: 1. Otrzymanie pożądanego odcinka DNA z materiału genetycznego dawcy 2. Wprowadzenie obcego DNA do wektora 3. Wprowadzenie wektora, niosącego w sobie
WYKŁAD: Klasyczny przepływ informacji ( Dogmat) Klasyczny przepływ informacji. Ekspresja genów realizacja informacji zawartej w genach
 Klasyczny przepływ informacji. Ekspresja genów realizacja informacji zawartej w genach") WYKŁAD: Ekspresja genów realizacja informacji zawartej w genach Prof. hab. n. med. Małgorzata Milkiewicz Zakład Biologii Medycznej Klasyczny przepływ informacji ( Dogmat) Białka Retrowirusy Białka Klasyczny
WYKŁAD: Ekspresja genów realizacja informacji zawartej w genach Prof. hab. n. med. Małgorzata Milkiewicz Zakład Biologii Medycznej Klasyczny przepływ informacji ( Dogmat) Białka Retrowirusy Białka Klasyczny
Temat: Budowa i funkcje korzenia.
 Temat: Budowa i funkcje korzenia. Korzeń to część podziemna organizmu roślinnego (organ wegetatywny) przystosowana do wypełniania określonych funkcji: Umocowania rośliny w podłożu. Pobierania z gleby wody
Temat: Budowa i funkcje korzenia. Korzeń to część podziemna organizmu roślinnego (organ wegetatywny) przystosowana do wypełniania określonych funkcji: Umocowania rośliny w podłożu. Pobierania z gleby wody
Biologia Molekularna z Biotechnologią ===============================================================================================
 Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Genetyki Molekularnej Bakterii Katedra Biologii i Genetyki Medycznej Przedmiot: Katedra Biologii Molekularnej Biologia
Biologia medyczna, materiały dla studentów
 Zasada reakcji PCR Reakcja PCR (replikacja in vitro) obejmuje denaturację DNA, przyłączanie starterów (annealing) i syntezę nowych nici DNA (elongacja). 1. Denaturacja: rozplecenie nici DNA, temp. 94 o
Zasada reakcji PCR Reakcja PCR (replikacja in vitro) obejmuje denaturację DNA, przyłączanie starterów (annealing) i syntezę nowych nici DNA (elongacja). 1. Denaturacja: rozplecenie nici DNA, temp. 94 o
Metody odczytu kolejności nukleotydów - sekwencjonowania DNA
 Metody odczytu kolejności nukleotydów - sekwencjonowania DNA 1. Metoda chemicznej degradacji DNA (metoda Maxama i Gilberta 1977) 2. Metoda terminacji syntezy łańcucha DNA - klasyczna metoda Sangera (Sanger
Metody odczytu kolejności nukleotydów - sekwencjonowania DNA 1. Metoda chemicznej degradacji DNA (metoda Maxama i Gilberta 1977) 2. Metoda terminacji syntezy łańcucha DNA - klasyczna metoda Sangera (Sanger
Klub Młodego Wynalazcy - Laboratoria i wyposażenie. Pracownia hodowli roślin i roślinnych kultur in vitro
 Klub Młodego Wynalazcy - Laboratoria i wyposażenie Zadbaliśmy o to, żeby wyposażenie w Klubie Młodego Wynalazcy było w pełni profesjonalne. Ważne jest, aby dzieci i młodzież, wykonując doświadczenia korzystały
Klub Młodego Wynalazcy - Laboratoria i wyposażenie Zadbaliśmy o to, żeby wyposażenie w Klubie Młodego Wynalazcy było w pełni profesjonalne. Ważne jest, aby dzieci i młodzież, wykonując doświadczenia korzystały
Zadania maturalne z biologii - 3
 Koło Biologiczne Liceum Ogólnokształcące nr II w Gliwicach 2015-2016 Zadania maturalne z biologii - 3 Zadania: Zad. 1(Wiktoria Wnuk, Weronika Żak, Tomasz Gojowy 2D) Na podstawie wykresu odpowiedz na pytania.
Koło Biologiczne Liceum Ogólnokształcące nr II w Gliwicach 2015-2016 Zadania maturalne z biologii - 3 Zadania: Zad. 1(Wiktoria Wnuk, Weronika Żak, Tomasz Gojowy 2D) Na podstawie wykresu odpowiedz na pytania.
Ćwiczenia 1 Wirtualne Klonowanie. Prowadzący: mgr Anna Pawlik i mgr Maciej Dylewski. Część teoretyczna:
 Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią ===============================================================================================
Uniwersytet Gdański, Wydział Biologii Biologia i Biologia Medyczna II rok Katedra Biologii Molekularnej Przedmiot: Biologia Molekularna z Biotechnologią ===============================================================================================
RT31-020, RT , MgCl 2. , random heksamerów X 6
 RT31-020, RT31-100 RT31-020, RT31-100 Zestaw TRANSCRIPTME RNA zawiera wszystkie niezbędne składniki do przeprowadzenia syntezy pierwszej nici cdna na matrycy mrna lub całkowitego RNA. Uzyskany jednoniciowy
RT31-020, RT31-100 RT31-020, RT31-100 Zestaw TRANSCRIPTME RNA zawiera wszystkie niezbędne składniki do przeprowadzenia syntezy pierwszej nici cdna na matrycy mrna lub całkowitego RNA. Uzyskany jednoniciowy
PL B1. UNIWERSYTET PRZYRODNICZY W LUBLINIE, Lublin, PL BUP 26/11
 PL 214501 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 214501 (13) B1 (21) Numer zgłoszenia: 391458 (51) Int.Cl. C12Q 1/68 (2006.01) C12N 15/29 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej
PL 214501 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 214501 (13) B1 (21) Numer zgłoszenia: 391458 (51) Int.Cl. C12Q 1/68 (2006.01) C12N 15/29 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej
Pobudliwość i koordynacja funkcji życiowych u roślin.
 Pobudliwość i koordynacja funkcji życiowych u roślin. Zadanie 1 A B C W doświadczeniu wykorzystano: syntetyczną auksynę i wodę. Jak zachowała się siewka A, B i C? Zadanie 2 I - Wyjaśnij jakiego czynnika
Pobudliwość i koordynacja funkcji życiowych u roślin. Zadanie 1 A B C W doświadczeniu wykorzystano: syntetyczną auksynę i wodę. Jak zachowała się siewka A, B i C? Zadanie 2 I - Wyjaśnij jakiego czynnika
Metody poprawy jakości nasion buraka cukrowego
 Metody poprawy jakości nasion buraka cukrowego Podlaski Sławomir Jubileusz 90-lecia urodzin Prof. dr hab. B. Geja i 90-lecia powstania Katedry Fizjologii Roślin Budowa handlowego nasienia buraka cukrowego
Metody poprawy jakości nasion buraka cukrowego Podlaski Sławomir Jubileusz 90-lecia urodzin Prof. dr hab. B. Geja i 90-lecia powstania Katedry Fizjologii Roślin Budowa handlowego nasienia buraka cukrowego
Organizmy Modyfikowane Genetycznie Rośliny transgeniczne
 Organizmy Modyfikowane Genetycznie Rośliny transgeniczne Co to GMO? GMO to organizmy, których genom został zmieniony metodami inżynierii genetycznej w celu uzyskania nowych cech fizjologicznych (lub zmiany
Organizmy Modyfikowane Genetycznie Rośliny transgeniczne Co to GMO? GMO to organizmy, których genom został zmieniony metodami inżynierii genetycznej w celu uzyskania nowych cech fizjologicznych (lub zmiany
KOD OPIS ROZWOJU Z BULWY OPIS ROZWOJU Z NASION
 Ziemniak Solanum tuberosum L. KOD OPIS ROZWOJU Z BULWY OPIS ROZWOJU Z NASION Główna faza rozwojowa 0: Kiełkowanie (formowanie pędów) 00 000 Bulwa w stanie spoczynku, nasiona suche kiełki niewidoczne 01
Ziemniak Solanum tuberosum L. KOD OPIS ROZWOJU Z BULWY OPIS ROZWOJU Z NASION Główna faza rozwojowa 0: Kiełkowanie (formowanie pędów) 00 000 Bulwa w stanie spoczynku, nasiona suche kiełki niewidoczne 01
Zawartość składników pokarmowych w roślinach
 Zawartość składników pokarmowych w roślinach Poszczególne rośliny różnią się zawartością składników pokarmowych zarówno w organach wegetatywnych, jak i generatywnych. Wynika to z różnych funkcji, jakie
Zawartość składników pokarmowych w roślinach Poszczególne rośliny różnią się zawartością składników pokarmowych zarówno w organach wegetatywnych, jak i generatywnych. Wynika to z różnych funkcji, jakie
"Dlaczego NIE dla GMO w środowisku rolniczym" Prof. zw. dr hab. inż. Magdalena Jaworska
 "Dlaczego NIE dla GMO w środowisku rolniczym" Prof. zw. dr hab. inż. Magdalena Jaworska Kierownik Katedry Ochrony Środowiska Rolniczego Uniwersytet Rolniczy w Krakowie Ekspert EU Biotechnology in Agriculture
"Dlaczego NIE dla GMO w środowisku rolniczym" Prof. zw. dr hab. inż. Magdalena Jaworska Kierownik Katedry Ochrony Środowiska Rolniczego Uniwersytet Rolniczy w Krakowie Ekspert EU Biotechnology in Agriculture
PL B1. UNIWERSYTET PRZYRODNICZY W LUBLINIE, Lublin, PL BUP 04/14. SYLWIA OKOŃ, Dąbrowica, PL KRZYSZTOF KOWALCZYK, Motycz, PL
 PL 220315 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 220315 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 400252 (22) Data zgłoszenia: 06.08.2012 (51) Int.Cl.
PL 220315 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 220315 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (21) Numer zgłoszenia: 400252 (22) Data zgłoszenia: 06.08.2012 (51) Int.Cl.
Pamiętając o komplementarności zasad azotowych, dopisz sekwencję nukleotydów brakującej nici DNA. A C C G T G C C A A T C G A...
 1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
1. Zadanie (0 2 p. ) Porównaj mitozę i mejozę, wpisując do tabeli podane określenia oraz cyfry. ta sama co w komórce macierzystej, o połowę mniejsza niż w komórce macierzystej, gamety, komórki budujące
Sukces kultur in vitro oparty jest na zjawisku totipotencji, czyli nieograniczonej zdolności komórek do dzielenia się i odtwarzania całego organizmu
 Rośliny z probówki Kultury in vitro to uprawa części roślin, tkanek, pojedynczych komórek, a nawet protoplastów poza organizmem macierzystym, na sztucznych pożywkach w warunkach sterylnych Sukces kultur
Rośliny z probówki Kultury in vitro to uprawa części roślin, tkanek, pojedynczych komórek, a nawet protoplastów poza organizmem macierzystym, na sztucznych pożywkach w warunkach sterylnych Sukces kultur
Wykład 1. Od atomów do komórek
 Wykład 1. Od atomów do komórek Skład chemiczny komórek roślinnych Składniki mineralne (nieorganiczne) - popiół Substancje organiczne (sucha masa) - węglowodany - lipidy - kwasy nukleinowe - białka Woda
Wykład 1. Od atomów do komórek Skład chemiczny komórek roślinnych Składniki mineralne (nieorganiczne) - popiół Substancje organiczne (sucha masa) - węglowodany - lipidy - kwasy nukleinowe - białka Woda
(86) Data i numer zgłoszenia międzynarodowego: , PCT/US99/21960 (87) Data i numer publikacji zgłoszenia międzynarodowego:
 Data i numer zgłoszenia międzynarodowego: , PCT/US99/21960 (87) Data i numer publikacji zgłoszenia międzynarodowego:") RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 197408 (21) Numer zgłoszenia: 346772 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 21.09.1999 (86) Data i numer zgłoszenia
RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 197408 (21) Numer zgłoszenia: 346772 (13) B1 Urząd Patentowy Rzeczypospolitej Polskiej (22) Data zgłoszenia: 21.09.1999 (86) Data i numer zgłoszenia
Sprawdź swoją wiedzę i umiejętności TKANKI ROŚLINNE. 1. Uzupełnij schemat ilustrujący hierarchiczną budowę organizmu roślin. komórka...
 Sprawdź swoją wiedzę i umiejętności TKANKI ROŚLINNE. 1. Uzupełnij schemat ilustrujący hierarchiczną budowę organizmu roślin. komórka...... organizm 2. Na rysunku komórki roślinnej wskaż i podpisz następujące
Sprawdź swoją wiedzę i umiejętności TKANKI ROŚLINNE. 1. Uzupełnij schemat ilustrujący hierarchiczną budowę organizmu roślin. komórka...... organizm 2. Na rysunku komórki roślinnej wskaż i podpisz następujące
Disruption of c-mos causes parthenogenetic develepment of unfertilized mouse eggs. W.H Colledge, M.B.L. Carlton, G.B. Udy & M.J.
 Disruption of c-mos causes parthenogenetic develepment of unfertilized mouse eggs. W.H Colledge, M.B.L. Carlton, G.B. Udy & M.J.Evans Partenogeneza-dzieworództwo, to sposób rozmnażania polegający na rozwoju
Disruption of c-mos causes parthenogenetic develepment of unfertilized mouse eggs. W.H Colledge, M.B.L. Carlton, G.B. Udy & M.J.Evans Partenogeneza-dzieworództwo, to sposób rozmnażania polegający na rozwoju
Hybrydyzacja kwasów nukleinowych
 Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
2014-03-26. Analiza sekwencji promotorów
 2014-03-26 Analiza sekwencji promotorów 1 2014-03-26 TFy tworzą zawiły układ regulacyjny, na który składają się różne oddziaływania białko białko poprzez wytworzenie PĘTLI Specyficzne TFy Ogólne TFy Benfey,
2014-03-26 Analiza sekwencji promotorów 1 2014-03-26 TFy tworzą zawiły układ regulacyjny, na który składają się różne oddziaływania białko białko poprzez wytworzenie PĘTLI Specyficzne TFy Ogólne TFy Benfey,
wykład dla studentów II roku biotechnologii Andrzej Wierzbicki
 Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Ekspresja genów jest regulowana
Genetyka ogólna wykład dla studentów II roku biotechnologii Andrzej Wierzbicki Uniwersytet Warszawski Wydział Biologii andw@ibb.waw.pl http://arete.ibb.waw.pl/private/genetyka/ Ekspresja genów jest regulowana
PL B1. Sposób i układ do modyfikacji widma sygnału ultraszerokopasmowego radia impulsowego. POLITECHNIKA GDAŃSKA, Gdańsk, PL
 PL 219313 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 219313 (13) B1 (21) Numer zgłoszenia: 391153 (51) Int.Cl. H04B 7/00 (2006.01) H04B 7/005 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej
PL 219313 B1 RZECZPOSPOLITA POLSKA (12) OPIS PATENTOWY (19) PL (11) 219313 (13) B1 (21) Numer zgłoszenia: 391153 (51) Int.Cl. H04B 7/00 (2006.01) H04B 7/005 (2006.01) Urząd Patentowy Rzeczypospolitej Polskiej
Wymagania edukacyjne biologia klasa 1
 Wymagania edukacyjne biologia klasa 1 Dział programu Numer i temat lekcji ocena dopuszczająca ocena dostateczna ocena dobra ocena bardzo dobra ocena celująca 1 2 3 4 5 6 7 I. Podstawy biologii 1. Biologia
Wymagania edukacyjne biologia klasa 1 Dział programu Numer i temat lekcji ocena dopuszczająca ocena dostateczna ocena dobra ocena bardzo dobra ocena celująca 1 2 3 4 5 6 7 I. Podstawy biologii 1. Biologia
Hybrydyzacja kwasów nukleinowych
 Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja tego genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
Hybrydyzacja kwasów nukleinowych Jaka jest lokalizacja tego genu na chromosomie? Jakie jest jego sąsiedztwo? Hybrydyzacja - powstawanie stabilnych struktur dwuniciowych z cząsteczek jednoniciowych o komplementarnych
Zainwestuj w rozwój systemu korzeniowego!
 .pl https://www..pl Zainwestuj w rozwój systemu korzeniowego! Autor: Karol Bogacz Data: 20 kwietnia 2017 Fundamentem każdej rośliny uprawnej jest jej system korzeniowy. To właśnie od niego zależy ilość
.pl https://www..pl Zainwestuj w rozwój systemu korzeniowego! Autor: Karol Bogacz Data: 20 kwietnia 2017 Fundamentem każdej rośliny uprawnej jest jej system korzeniowy. To właśnie od niego zależy ilość
(12) OPIS PATENTOWY. (86) Data i numer zgłoszenia międzynarodowego , PCT/KR96/00238
 OPIS PATENTOWY. (86) Data i numer zgłoszenia międzynarodowego , PCT/KR96/00238") RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (21) Numer zgłoszenia: 327971 (22) Data zgłoszenia: 11.12.1996 (86) Data i numer zgłoszenia międzynarodowego 11.12.1996,
RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (21) Numer zgłoszenia: 327971 (22) Data zgłoszenia: 11.12.1996 (86) Data i numer zgłoszenia międzynarodowego 11.12.1996,
BUDOWA I FUNKCJA GENOMU LUDZKIEGO
 BUDOWA I FUNKCJA GENOMU LUDZKIEGO Magdalena Mayer Katedra i Zakład Genetyki Medycznej UM w Poznaniu 1. Projekt poznania genomu człowieka: Cele programu: - skonstruowanie szczegółowych map fizycznych i
BUDOWA I FUNKCJA GENOMU LUDZKIEGO Magdalena Mayer Katedra i Zakład Genetyki Medycznej UM w Poznaniu 1. Projekt poznania genomu człowieka: Cele programu: - skonstruowanie szczegółowych map fizycznych i
skróci wzmocni pogrubi
 skróci wzmocni pogrubi zapobiega wyleganiu roślin zbóż rozwija pędy boczne i system korzeniowy pogrubia ścianki źdźbła hamuje wzrost elongacyjny działa także w późnych fazach rozwojowych zbóż Po co regulować
skróci wzmocni pogrubi zapobiega wyleganiu roślin zbóż rozwija pędy boczne i system korzeniowy pogrubia ścianki źdźbła hamuje wzrost elongacyjny działa także w późnych fazach rozwojowych zbóż Po co regulować
Masowe występowanie koguciego ogona na plantacji Objawy koguciego ogona niedługo po posadzeniu Objawy koguciego ogona niedługo po posadzeniu
 Koguci ogon Jedną z chorób tytoniu, z którą niekiedy spotykają się plantatorzy tytoniu, a często nie potrafią powiązać obserwowanych objawów z konkretną przyczyną jest koguci ogon. Koguci ogon można czasem
Koguci ogon Jedną z chorób tytoniu, z którą niekiedy spotykają się plantatorzy tytoniu, a często nie potrafią powiązać obserwowanych objawów z konkretną przyczyną jest koguci ogon. Koguci ogon można czasem
(12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:
 TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP (96) Data i numer zgłoszenia patentu europejskiego:") RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2329028 (96) Data i numer zgłoszenia patentu europejskiego: 01.09.2009 09778245.2 (13) (51) T3 Int.Cl. C12N 15/82 (2006.01)
RZECZPOSPOLITA POLSKA (12) TŁUMACZENIE PATENTU EUROPEJSKIEGO (19) PL (11) PL/EP 2329028 (96) Data i numer zgłoszenia patentu europejskiego: 01.09.2009 09778245.2 (13) (51) T3 Int.Cl. C12N 15/82 (2006.01)
Geny i działania na nich
 Metody bioinformatyki Geny i działania na nich prof. dr hab. Jan Mulawka Trzy królestwa w biologii Prokaryota organizmy, których komórki nie zawierają jądra, np. bakterie Eukaryota - organizmy, których
Metody bioinformatyki Geny i działania na nich prof. dr hab. Jan Mulawka Trzy królestwa w biologii Prokaryota organizmy, których komórki nie zawierają jądra, np. bakterie Eukaryota - organizmy, których
Regulacja Ekspresji Genów
 Regulacja Ekspresji Genów Wprowadzenie o Ekspresja genu jest to złożony proces jego transkrypcji do mrna, o Obróbki tego mrna, a następnie o Translacji do białka. 4/17/2019 2 4/17/2019 3 E 1 GEN 3 Promotor
Regulacja Ekspresji Genów Wprowadzenie o Ekspresja genu jest to złożony proces jego transkrypcji do mrna, o Obróbki tego mrna, a następnie o Translacji do białka. 4/17/2019 2 4/17/2019 3 E 1 GEN 3 Promotor
(12) OPIS PATENTOWY (19) PL
 OPIS PATENTOWY (19) PL") RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (21) Numer zgłoszenia: 323031 (22) Data zgłoszenia: 07.11.1997 (11) 185976 (13) B1 (51) IntCl7 F25B 39/00 F25D
RZECZPOSPOLITA POLSKA Urząd Patentowy Rzeczypospolitej Polskiej (12) OPIS PATENTOWY (19) PL (21) Numer zgłoszenia: 323031 (22) Data zgłoszenia: 07.11.1997 (11) 185976 (13) B1 (51) IntCl7 F25B 39/00 F25D